リン ゴ果 実 の エ チ レ ン生 合 成 制 御 に
関す る遺伝子工学
(研究課題番号 :
09
660001 )
平成
9
年度〜平成1 1
年度科学研究費補助金 基盤研究(C) (2)
研究成果報告書平成
12 年 3 月
研究代表者:原(弘前大 田 竹 雄
リン ゴ果実 のエ チ レン生 合 成制御 に関す る遺伝 子工 学
研 究組 織
研 究 代 表 者 :原 田竹 雄 (弘 前 大 学 農 学 生 命 科 学 部 助 教 授 ) 研 究 分 担 者 :な し
研 究 経 費 :
平 成 9年 度 平 成
1 0
年 度 平 成11
年 度計
1,700
千 円600
千 円800
千 円3,1 00
千 円研 究 発 表 等
(1) 学 会 誌 等
Suna koT
,Sa kur a baW, Se ndaM
,AkadaS,I s hi ka waR
,Ni i ze kiM a ndHa r adaT:
Ana l l el eoft her l pe nl ng‑ S pe ci f i c1‑ a mi noc yc l opr opa ne1‑ c a r boxyl i ca ci ds ynt ha s e gene( ACSl )i na ppl ef r ui twi t hal ongs t or a gel i f e. Pl antPh ys i o
l.1 19:1 2971 1303 1999.
Ha r adaT
,Suna koT
,Wa ka s a
Y,Soe J l ma
∫,Sa t ohT
,Ni i ze ki
M:Ana l l el eof t he1‑ a mi noc yc l opr opa ne ‑ l l Ca r boxyl a t es ynt ha s ege ne
(Md‑ACSl)a cc ount sf or t hel ow l e velofe t hyl e ne pr oduc t i oni nc l i ma c t e r i cf r ui t sofs omea ppl ec ul t i va r s . The o r . Appl .Ge ne t . i npr e s s2000・
Suna koT
,I s hi ka waR
,Se nda
M,Aka daS
,Ni i ze ki
Ma ndHa r a daT ( 2000) MdACS‑ 5A ( Ac c e s s i onNot ABO34992)a nd5B ( Ac c e s s i onNo. ABO34993)
,Two Wound‑ Re s pons i veGe ne sEnc odi ng1 ‑ Ami noc yc l opr opa ne ‑
1‑ Ca r boxyl a t e Synt ha s ei nAppl e . ( PGR00‑ 030) Pl antPh ys i o
l・1 22:620.2000.
( 2)
口頭 発 表桜 庭 和 歌 子 ・砂 子 智 美 ・原 田竹 雄 ・千 田 峰生 ・赤 田辰 治 ・石 川 隆 二 ・新 関 稔 : リ ン ゴ
ACC
合 成 酵 素 遺 伝 子 の5'
隣接 領 域 に見 出 され たsI NE
につ い て . 冒 本 育種 学 会 第92
回講 演 会 (鳥 取 大 学 )1 997
年9
月.砂子智美 ・原田竹雄 ・千 田峰生 ・赤 田辰治 ・石川隆二 ・新関 稔 :リンゴ果実内の
A C C
合成酵素遺伝子 (A CS )
の発現解析. 日本育種学会第93
回講演会 (横 浜市立大学)1 9 97
年4 月.
∴' ; : : . , ; ・7
/′‑r† /*′‑一一一一̲I
∫)㌔
原田竹雄 ・砂子智美 ・佐久間智宏 ・千田峰生 ・赤田辰治 ・石川隆二 ・新関 稔 :リ ンゴ栽培品種の
A C S l
遺伝子型 と果実内エチ レン濃度. 日本育種学会第93回講演会(横浜市立大学)1997年
4
月.砂子智美 ・石川隆二 ・千田峰生 ・赤田辰治 ・新関 稔 ・原田竹雄 :リンゴ各組織 に お ける
A C C
合成酵素遺伝子 (ACS)の発現解析 日本育種学会第94回講演会(岩手大学) 1998年9月.
桜 庭 和 歌子 ・神 修 平 ・石川 隆二 ・千 田峰生 ・赤 田辰 治 ・新 関 稔 ・原 田竹 雄 :リ ン ゴに見 出 され た レ トロポ ゾ ン Md‑SINElにつ い て. 日本 育種 学 会 第 94回講 演会 (岩手 大 学 ) 1998年 9月.
原 田竹 雄 ・砂 子 智美 ・佐 藤 耕 ・副 島淳一 ・新 関 稔 :リ ン ゴにお け る
A C C
合 成 酵 素 遺 伝 子 (Md‑
A CS l )
の対 立 遺 伝 子 型 とラ イ ブニ ン グ果 実 内 エ チ レ ン 量 .日本 育種 学会 第 95回講演会 (宇都 宮 大 学 )1999年 4月.砂 子 智 美 ・石 川 隆二 ・千 田峰生 ・赤 田辰 治 ・新 関 稔 ・原 田竹 雄 :リン ゴ
A C C
合 成酵 素 遺伝 子
( A C S )
フ ァ ミリーの発 現解析 .日本 育種 学会 第 95回講 演 会 (宇 都 宮 大 学 )1999年4月.Harada,T,SunakoT,SoeJlma∫,SatoT,NiizekiM∴Low ethyleneproductionin thefruitofsomeapplecultivarsiscausedbyamutationintherlpenlngSpecific ACCsynthasegene.XVIInternationalBotanicalCongressAbstracts,604,
(St.Rouis),1999・
若 佐 雄 也 ・神 修平 ・石川 隆 二 ・赤 田辰 治 ・新 関 稔 ・原 田竹 雄 :リン ゴ果 皮 赤色 化 遺伝 子 Rfに連鎖 す る DNAマ ー カー に兄 い だ された トラ ンス ポ ゾ ン Md‑Tnlにつ い て. 日本 育種 学会 第 96回講演会 (岡山大 学 )1999年 9 月 .
(3) 出版 物
原 田竹 雄 :リ ン ゴの貯 蔵 性 を高 め る
A C C
合 成 酵 素 の変 異 遺伝 子 .化 学 と生 物 .37: 429‑431.1999年 7月 .HaradaT,SunakoT,SakurabaW,SendaM,AkadaS,IshikawaRandNiizekiM:
Md‑ACSl,afruitripenlng1‑aminocylopropane‑1‑carboxylatesynthasegeneof apple・BreedingandBiotechnologyforFruitTrees・NIFTS/CISROp42‑45・1998
は じめ に
リンゴは世界 にお ける生産高が約
4 0 0 0
万 トンに達 し,ブ ドウ,バ ナナ, カ ンキツに次 ぐ果実 である.わが国 においては約1 0 0
万 トンが生産 されてお り, その5
割 は青森県 に よる ものである (農林水 産省統計情報部1 9 9 8 ).
リンゴは 他の果実 に比べ収穫 後の 日持 ち性 が高 く,長期の貯蔵が可能であ る とい う特徴 があ る. しか し,品種 に よって収穫後の商品価値 を保持で きる期 間が異 な り, 短い ものは長期貯蔵 に不適当であ って,収穫後す ぐに出荷 しなければな らない.一方,長 い もの は常 温貯蔵 ,冷蔵 ,
c A
貯蔵 な どを行 うこ とで,出荷 時期 を延 長す ることがで きる. この ような品種 の多様性 と産地 に よる熟期 のず れな どから, リンゴで は周年 に近い出荷体制がで きている.
ライブニ ングは果実の肥大生長が止 まってか らの有機酸の消失 ,細胞 の軟弱 化 な どの生理現象 をい う. リンゴの成熟 はライブニ ングに伴 って呼吸量が急激 に上昇す るク リマ クテ リック型であ り,エチ レンが ライブニ ング過程へ の プロ グラムス イ ッチであることが判 ってい ることか ら, リンゴの成熟や貯蔵性 はエ チ レンの生合成 と密接 な関係 にある と思われる」実際に,品種 に よ りライブニ ング過程 でのエチ レン生成量 はか な り異 な り, ライブニ ングが ゆっ くりと進行 す る品種 で はエチ レン生成量 が比較 的少量 である とい う報告 が あ る こ とか ら
( c h u1 9 8 8
,G es s ma ne tal .1 9 9 3
,毒松木 ら1 9 9 7 )
, この品種 の もつ遺伝 的素 質が注 目されて きた. しか し, この様 なエチ レン生成量 の品種 間差 の分子機構については不 明であ った.
エ チ レンの生合成 は,メチオニ ン
‑S‑
アデ ノシルメチオニ ン( S A M )‑1‑
ア ミノシクロプロパ ン‑1
‑カルボン酸( A C C )
‑エチ レンの経路で行 われ( Y a n g a n dH of f ma n1 9 8 4 )
,S A M
‑A C C
を触 媒 す るA C C
合 成 酵 素( S‑ a d e n o s yl ‑ L一 m et h yl t hi o a de n osi nel y a s e
,E C 4. 4. 1 . 1 4 )
が この経路の律速酵素であるこ とが明 らか にされている.エチ レンは果実の ライブニ ングのみな らず,植物生 長の 多 くの過程 を調節 しているホルモ ンである.その生成 は,時期 お よび組織特異 的に行 われ, また,傷害,病原菌の感染,高温 などのス トレス に よって も一過 的に活性化す る
( Ya n ga n dH of f ma n1 9 8 4 ).
この様 な多 くの生理現象 に対応す る形で,A C C
合成酵素( A C S )
遺伝子 は多重遺伝子族 を構成 してい る. トマ ト では8
つ( S hi ue ta l .1 9 9 8 )
,アラビ ドプシスでは5
つ( Li a n geta l .1 9 9 6 )
の
A C S
が報告 されてお り,それぞれの遺伝子 は異なる発現調節機構 で制御 され てい るこ とが判 って きた. リンゴにおいては,M d‑ A C S l ( D o n ge ta l . 1 9 91
,L a y‑ Y e ea n dK ni g ht o n1 9 9 5 )
,M d‑ A C S 2
,M d‑ A C S 3 ( R os e nfi el de tal .1 9 9 6 )
,M d‑ A C S 4 ( Ki neta l . 1 9 9 2 )
の4
つのA C S
遺伝子 のc D N A
塩基配列が既 に報 告 されてお り,この うち, M d‑ A C S l
についてはゲノ ミックD N A
の配列( H a r a dae t a l . 1 9 9 7 )
が明 らか にされている. しか し, これ らM d‑ A C S
の発現制御 につ い てはほ とん ど報告が ない.本研 究では, リンゴの ライブニ ング果実での これ ら
M d‑ A C S
の発現 について‑ 1‑
解析 を進 めた. また, これ らの解析 結 果 とライブニ ング時のエ チ レン生成量 の 関係 につ いて も詳細 に検討 した. さ らに,本研 究 を通 して新 た に単 離 された Md‑ACSに関す る特徴 づ け を進 めた.
Ⅰ リ ン ゴ果 実 内 の
ACC
合 成 酵 素 遺伝 子( ACS)
の発 現 解 析リンゴ果実の ク リマ クテ リック型 ライブニ ングにおけるエチ レンの 関与 は古 くか ら知 られてお り,果実の貯 蔵性 とエチ レンに関す る様 々な研 究が精力 的 に 進 め られて きた
( c h u1 9 8 8
,G e s s ma netal .1 9 9 3
,蓑松木 ら1 9 9 7 ).
しか し, エ チ レン生合 成 の制御 機構 につ いて は末 だ不 明の 点が多 い( S a y m o u reta l . 1 9 9 3 ).
ライブニ ング時の リン ゴ果実 か らは, これ までエチ レン生合成系 の律 速酵素であるA C S
遺伝子 のc D N A
が3
クロー ン (Md‑ACSl,Md‑ACS2,Md‑ACS3)報
告 されてい る( D o n getal .1 9 9 1
,L a y‑ Y e ea n dK ni g ht o n1 9 9 5
,R os e nfi el de t a l . 1 9 9 6 ).
しか し, これ らの発現 につ いてはほ とん ど明 らか にされてい ない.我 々は Md‑ACSlの ゲ ノ ミック構 造解析 を通 して,Md‑ACSlには基本 型 であ る
1‑ 1
型 と,その プロモー ター領域 に1 6 2 b p
のM d‑ SI N El
が挿入 した1‑ 2
型 の2
つ の対 立 遺伝 子 が存在 す る こ とを明 らか に した
( H a r a d ae ta l . 1 9 9 8 ).
既 報 の Md‑ACSlのc D N A
の塩基配列( D o n geta l .1 9 9 1
,L a y‑ Y e ea n dX ni g ht o n1 9 9 5 )
は遺伝子型が
1‑ 1 / 1‑ 2
へ テロ型 であ る リンゴ品種 ̀ゴールデ ンデ リシ ャス ' よ り単離 されてい るが,いず れのc D N A も 1‑ 1
型 である. この ことか ら, この プ ロモー ター領域 の差異 に よ り1‑ 1
型 と1‑ 2
型 では発現 に違 いが見 られるのでは ないか と考 え られた.そ こで本 章で は これ らの遺伝 子 の発現様 式 につ いて詳 しく検 討 した.す なわ
ち
,ACSlの対立遺伝子の発現 を調べ るために,リンゴ栽培品種 の中か ら,1‑ 2/ 1‑ 2
のホモ型 である ̀ふ じ' と
1‑ 1 / 1‑ 2
のヘ テ ロ型 である ̀ゴールデ ンデ リシ ャス' を用いて,Md‑ACSlと Md‑AC53をプローブ として ノーザ ン解析 を行 った.また,1‑ 2
型遺伝 子の発現 をR T‑ P C R
解析 に よ り検討 した.材料お よび方法
1 .
実験材料弘前大 学農学部付属藤崎農場 よ りサ ンプ リング された, リンゴ栽培 品種 ̀ゴ ールデ ンデ リシ ャス'お よび ̀ふ じ'の プ レク リマ クテ リック とク リマ クテ リ
ックの果実 を用 いた.
2 .
内部エチ レン濃度 (I E C )
の測定果実内エチ レン濃度 は,果実 の陽光面 とその反対側か ら果実 を切 り取 り,水 の入 ったデシケー ター内で果実内の気体 を回収 し,注射器で採取 した 1
ml
を 用 いて , ガス ク ロマ トグ ラフ ィー( S HI M A D Z UG C‑ 8 AG A SC H R O M A T O G R A P H )
に より測定 した.測定値 は
5
個体 の果実の平均 に よ り算 出 した.3.
サザ ンハ イブ リダイゼ ーシ ョンゲノ ミック
D N A
は成業か らv a r a d a r aj a ne ta l . ( 1 9 91 )
の方法 に よって抽 出 し,C s Cl 2 ‑ Et Br
法で分画 した.各D N A5F Lg
に制 限酵素 を加 え,全量1 0 0F Ll
の 反応液で消化 した.電気泳動 は,0. 8%
水平平板 アガロースゲルを用 いて2 3 V 1 5
時 間行 った.泳動終 了後 ,
Al k al it r a ns f e rb uf f e r ( 0 . ̀ 4NN a O H )
で ナイ ロ ン‑ 3I
メ ンブ レンフィル ター
( H y b o n d
T"N + ;A me r s h a m )
に約2 4
時間ブロッテ ィングを 行 い,その後 ,5×S S C
溶液で5
分 間,2×S S C
溶液で5
分 間メ ンブ レンを洗浄した.ハ イブ リダイゼーシ ョンは,
H y b ri di z ati o nb uf f e r ( 6×S S C
,2m ME D T A p H8. 0
,1 0m MTri s‑ H Clp H7 . 5
,5×D e n h a r d s
,0 . 2m gS al m o nS p e r m a r yD N A
,1 0m MN a 3 P
O。 , 1
%N aN‑ L a u r o ylS al c o si n a t e )
へP ri me‑ I tⅡR a n d o mP ri me r La b el i n gKi t( S T R A T A G E N E )
を用 いて3 2 p
で標 識 したD N A
プローブ( 2 0 0n g )
を 加 え,6 5℃1 2
時間以上行 った.洗浄 は,w a s hi n gb uf f e r( 2×S S C
,0 . 1 %S D S )
で 室温2 0
分 間の条件 で2
回行 った. さ らに,( 0. 2×S S C
,0 . 1 0 / O S I ) S )
で6 5o C
,2 0
分 間 2 回洗浄 した.その後,オー トラジオグラフィーに よるシグナル検 出 を行 った.
4.R N A
抽 出D o n geta l . ( 1 9 9 1 )
の方法 を改良 して, リンゴ果実 よ り全R N A
を調製 した.5g
の 試 料 を 液 体 窒 素 中 で 磨 砕 後 ,4 0ml
の 緩 衝 液 (4. 0 Mg u a ni di n e i s ot hi o d y a n a t e
,1 0m ME D T A
,3 0 0m MTri sp H 7 . 5
,1% 2‑ me r c a pt o et h a n ol
,0. 5% s o di u ml a t l r O yls a r c o si n e )
を加 えた.室温で1 0
分 間放置後,p ol y vi n yl p yr r ol i d o n e( 1 / 1 0F W )
を加 え,氷上 で2
時間放置 した. これ を4℃
,8 , 0 0 0r p m
で
1 5
分 間遠心 し,上澄 み を濠過 した. これにC s C1 2
を加 え,2 0℃
,2 5 , 0 0 0r p m
で
2 4
時 間平衡 密 度勾 配遠 心 した.沈殿 物 を滅菌水 に とか し, フェノール/ク ロロフ ォルム処理 した後 ,エ タノール沈殿 させ たR N A
分 画 を滅菌水 に とか した.5.
ノーザ ン分析全
R N A2 0〃g
をR N A
試 料 媛 衝 液( 1×M O P S
,5 0% f or m a mi d e
,2 . 2M f or mal d e h y d e
,1 0m ME D T A )
に とか し,6 5℃ 1 5
分 間変性 させ た後 ,氷上 で急 冷 させ た.その 中に,染色 液( 0 . 01 % B P B
,0. 0 2 5% X C
,0 . 0 5m g / mlEt B r )
を 加 え,1 . 2%
ゲル( 1
×M O P S
,0 . 6 6Mf or mal d e h y d e
,1 . 2%
アガロース)で4 0 V
,2. 5
時 間電気 泳動 を行 った.泳動終 了後 , ゲルを1 0×S S C
で2 0
分 間,2
回 洗 浄 した後 ,2 0×S S C
に よ り一晩ブロ ッテ ィングを行 い,変性R N A
をナイロン メ ンブ ランフィル ター( H y b o n d‑ N )
上 にブロ ッテ ィング した.U v
で固定 した後 ,5 0% f or ma mi d e
,5×D e n h a r dt ' ss ol uti o n
,0. 5%S D S
,5×S S P E
,2 0〃g/ mls al m o n s p e r mD N A
を含 む溶液 中でプ レハ イブ リダイゼーシ ョンを行 い,3 2 p
で ラベ ル し た プローブD N A
を加 え,4 2℃
で一晩ハ イブ リダイゼーシ ョンを行 った.その後 , メ ンブ レンは2×S S C / 0 . 1 % S D S
で室温 ,5
分 を2
回,0. 2×S S C / 0 . 1 % S D S
で 室温1 0
分 ,0. 2×S S C / 0 . 1 % S D S
で6 5℃
,1
時間洗浄 した.6.R T‑ P C R
反応T ot alR N AI O〃g
を2 0〃1
の逆転写反応液( 5 0m MK C
l,1 0 m MTri s‑ H Clp H8 ・ 3
,5m M M g Cl
Z,im Md N T P s
,lF LM Ol i g o( d T ) p ri m e r
,5 0 u ni t sM u L V r e v e r s e
t r a ns 。 ri pt a s e;P E R KI NE L M E R , 2 0 u ni t sR N a s el n hi bi t or;P E R KI NE L … E R )
中 で ,4 2℃
,1
時 間‑9 4℃
,5
分 間反応 させ た後 ,氷上 で急冷 させ てc D N A
を合 成 した. この合 成 されたc D N A
をさらに1 0 0〃1
のP C R
反応液( 5 0m MK C
l,1 0m M
Tri s‑ H Clp H8. 3
,2 . 5
mMM g C1 2
,2 0 0〃Md N T P s
,0 . 4〃Mp ri me r s( A C S 1‑ 3 ' A )
,5unitsTaq polymerase)中で,94℃,2分‑45℃,1分‑72℃,3分 の反応 を 30サ イクル継続す るこ とに よ り,cDNA の増 幅 を行 った.増幅 した DNA は さ ら
に内側 の プ ライマ ー (ACS1‑3'B)を用 いて PCR 反応 を行 った.
7.PCR反応
ACSトA,ACS1‑Bの両 プローブはプラス ミ ド pAP1‑6を,ACS3プローブは リン ゴ品種 ̀旭 'の葉か ら抽 出 した DNAをテ ンプ レー トとした pcRの増 幅産物 を利 用 した. これ らの DNAは pcR反応 液 (10mMTris‑HCl pH8.3,50mMKCl,12.5 mMMgC12,0.2mMdNTPs,0.5FLMprimers,2unitsAmpliTaqGoldT";PERXINELMER)
中で 94℃,1分 反応 させた後 ,94℃ 1分 ,55℃ 2分 ,72℃ 3分 のサ イクル を 40回行 った.pcRと RT‑PCRに用 いた プ ライマ ーを下記 に示 す.尚,数値 は ACSl
に関す る もの は AccessionNo.U89156,ACS3に関す る もの は U73816に対応 す る番号 であ
ACS1‑AF ACS1‑AR ACSl‑BF ACSトBR ACS3‑F ACS3‑R ACSL3'AF ACS1‑3'AR ACSll3'BF ACS1‑3'BR
結果
る.
5'TACCATGAGGTCCACAACAC3' 5'GGTGAGCACTAAGTGGTTGGG3' 5'GATGAAAGGTAGCCTGGTCTGA3' 5'TACACTAATCACATTGTATAGAATC3I
5'GACAAATAGAAAGAGACCTGAGGACG3' 5'ccATCGATTATACAAACTGATTGTG3' 5'TCCACAACACAAACGGGATTATTCA3' 5'AGGCTACCTTTCATCTACCGGGA3' 5'ATCTTCTCGAGTCATGGCTGGC3'
5'TTTCATCTACCGGGAATAGGACCGCGG3'
2302‑2321 2813‑2793 4056‑4077 4513‑4489 1086‑1110
1573‑1549 3312‑3336 4070‑4048 2489‑2510 4062‑4036
1.Md‑ACSlお よび Md‑ACS3に対す る特 異的なプローブの作 製
pcRに よ り ACS1‑A,ACS1‑B,ACS3の DNA プローブの調製 を試 み た.ACSlプ ローブは Md‑ACSlが挿入 されているプ ラス ミ ド (pAP116)を, また,ACS3プロ ーブは品種 ̀旭 'の葉 か ら抽 出 した全 DNAをテ ンプ レー トとした pcRに よる増 幅産物 を用 いた.ACS3の PCR産物 は プラス ミ ドに挿入 し, シークエ ンスす るこ とで Md‑ACS3の一部 で あることを確 か めた.また,Md‑ACS2 (Rosanfieldeta
l .
1996)につ いて もプローブを作製す るため,複数の プライマ ーを作 製 L pCR を 行 ったが ,いず れ も 目的の長 さの産物 が得 られず , プローブ を得 るこ とがで きなか った.
作 製 された プ ローブはその特異性 を調べ るため に, リンゴ品種 ̀ふ じ', ̀ゴ ールデ ンデ リシ ャス', ̀印度'か ら抽 出 した全 DNAを
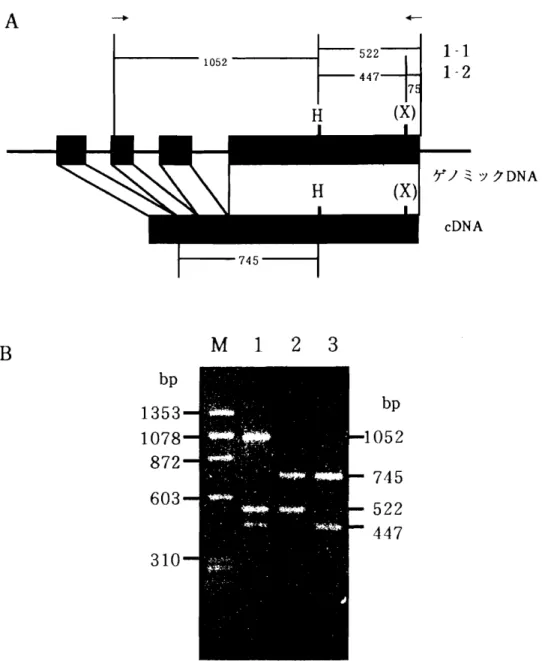
Ec o R I
で切 断 し,上記 の プ ローブに よるサザ ン解析 を行 った (図 211).Md‑ACSl5'末端側 の他 の Md‑ACS と比較 的相 同性 の高い ACS1‑Aをプローブ と した場合 ,Md‑ACSlで あ る 1.5kbの‑ 5‑
バ ン ド以外 に もシグナルが見 られた. また,
3 '
末端側 のA C S 1‑ B
をプローブ と した時 は ̀ふ じ'お よび ̀ゴールデ ンデ リシ ャス'で Md‑ACSlの2 . 1 k b
のバ ン ドのみが み られた.なお, ̀印度'では Md‑ACSl以外 のバ ン ドがみ られたので, 今 回の発現解析実験 では使用 しなか った.Md‑ACS3では Md‑ACSlお よび Md‑ACS3に対 し,比較的相 同性 の低 い
3'
末端 側 をプローブ としてサザ ン解析 を行 った (図21 2 ) .
その結果,Md‑ACS3と考 え られ るバ ン ドに加 え,弱 いシグナルのバ ン ドが検 出 された. しか し, ここで検 出 され るバ ン ドが Md‑ACSl由来の もの を含 まないことか ら, この プローブが完 全 な もので はないが,Md‑ACSl以外 の プローブ として使用で きる と判 断 され,A C S 3
プローブ として使用 した.2.
Md‑ACSlお よび Md‑ACS3の発現解析先 ず ,Md‑ACSlの対 立遺伝子 間の発現の違 い を検討 した (図 213).Md‑ACSl には基本 型の
1‑ 1
型 と5 '
隣接領域 にSI N E
を有す る1‑ 2
型 に分類 される.そ こ で,ト2/ 1‑ 2
のホモ型 である ̀ふ じ' と1‑ 1 / 1‑ 2
のヘテロ型 である ̀ゴールデ ンデ リシ ャス' を用 いて ノーザ ン解析 を行 った. ̀ゴールデ ンデ リシ ャス ' と̀ふ じ'の内部エチ レン濃度
( I E C )
は,それぞれ収穫 直後では1 6/ ∠1/1
,0 . 7
〃1/1
であ り,2 0℃
,1 2
日間静置後では2 9 8/ ∠1/1
,5 7〃1/1
とい う値 を 示 した.そ こで,収穫直後 をプ レク リマクテ リック,1 2
日間静置後 をク リマク テ リックの果実 とし,それぞれか らR N A
を抽 出 した.A C S l
をプローブ とした 場合 ,プ レクリマクテ リック果実では両品種 ともシグナルが認め られなか った.一方, ク リマ クテ リツク果実で は ̀ゴールデ ンデ リシ ャス'では強い発現がみ られ たが , ̀ふ じ'で は極微弱の シグナルが認 め られた. この メ ンブ レンを用 い て
A C S 3
をプローブ とした場合, プ レク リマ クテ リック果実 において も両品 種 で シグナルがみ られ, さ らに,クリマクテ リックでその量が明瞭 に上昇す る こ とは なか った. この こ とか ら, ライブニ ング時に発現す るA C S
遺伝 子族 の 中で,A C S 3
が常時発 現 を示す の に対 し,ACSl‑1はク リマ クテ リック特異 的 に 発現す ることが判 った.次 に, ̀ふ じ'で得 られたシグナルが実際に ACSl‑2型由来の ものであるか を 確 か め るた め に
R T‑ P C R
を行 った (図4 )
. 図4 A
の矢 印 で示 す プ ライ マ ー( A C S 1‑ 3' B F
お よびA C S 1‑ 3 ' B R )
を使用 した増幅産物 を H)'ndⅢと xhoI
で切 断 し た場合 ,1‑ 1
型 は5 2 2 b p
となるが,1‑ 2
型 は3 '
末端側 に XhoI
認識部位 が存 在 す るた め4 4 7 b p
と7 5 b p
に分 断 される. また,セ ンス プライマ ーか ら H)'ndⅢ
までの長 さはイ ン トロンを含 むゲノ ミックD N A
では1 0 5 2 b p
であるのに対 し,c D N A
で は7 4 5 b p
とな るので,得 られたR T‑ P C R
の産物が混入す るゲ ノ ミックD N A
由来の ものではない ことが判定で きる.図4B
で示 した通 り, ̀ゴールデ ン デ リシ ャス'で は両遺伝子型 を保有 しているに もかかわ らず ,1‑ 1
型のみが, 一方 の ̀ふ じ'では1‑ 2
型のみが確認 された.これ らの結果か ら,
1‑ 2
型 は1‑ 1
型 に比べ発現量が極端 に少 な く, ̀ゴールデ ンデ リシ ャス'の ように発現が認 め られない場合 もあるこ とが判 明 した.
考 察
リンゴ栽培 品種 ̀ゴールデ ンデ リシャス'には ライブニ ング時 に発現す る
A C S l
が
1
対 の対 立遺伝 子 と して存在す る.そ こで, これ ら両遺伝 子 (Md‑ACSl‑1 と Md‑ACSl‑2)の転写 について検討 した.R T‑ P C R
の結果 , ̀ゴールデ ンデ リシ ャス'で は1‑ 1
型の転写物が確 認 された が ,1‑ 2
型 は検 出 されなか った.同様 の結果 は増幅 プライマーの位置 を変 えたR T‑ P C R
で も得 られた( H a r a d aeta l .1 9 9 8 ) .D o n ge ta l . ( 1 9 9 1 )
お よびL a y‑ Y e e a n dK ni g ht o n ( 1 9 9 7 )
は ̀ゴールデ ンデ リシ ャス'由来の Md‑ACSIc D N A
の塩 基 配列 を報告 しているが,両者 とも1‑ 1
型であ り,本章の結果 と符合す る もの で ある.一方,Md‑ACS上2をホモ型 で有す る栽培 品種 ̀ふ じ' につ いて も同様 の良 T‑ P C R
解析 を行 った ところ,1‑ 2
型の転写物が確 認 された. しか し, ノーザ ン解析 に よるこれ らの転写量 を調べ てみ る と, ̀ふ じ'の1‑ 2
型 は ̀ゴールデ ンデ リシ ャス' に比べ ,圧倒 的 に少量 であ った.以上 の こ とか ら,ト2
型遺伝 子 はト1
型 に比べ発現が極端 に低 い こ とが明 らか となった.Md‑ACSl12にはプロモーター領域 に
M d‑ SI N El
が挿入 されているが,その部位 が転写 開始点か ら‑ 7 81 b p
であること,挿入配列 は1 6 2 b p
と短い こ とな どか ら, この挿入 の遺伝 子発現 レベ ル‑ の影響 が どの程度であ るか注 目され る.5'
上 流 領域 に トランスポゾンが挿入 されたA C O ( Bl u mee ta l . 1 9 9 7 )
やA C S ( S hi net a l . 1 9 9 8 )
が報告 されているが , これ らは トランスポ ゾ ン非挿入型 との比較 がない こ とか ら挿入‑ の影響 は不 明であ る.両対立遺伝 子 間には 5'上 流域 に塩 基 置換が複数あ ることか ら, シス因子の 1塩基変異 に よって転写活性が異 なる こ とも予想 され る.
1‑ 2
型の転写能低下の原 因を確 か めるにはト2
型 か らsI N E
を除去 した際の プロモー ター活性 を測定す る解析 が必要 となるだろ う.
トマ トでは8つある ACSファミリーの 中で 2つが ライブニ ング時 に発現 して い る
( Li nc ol ne tal .1 9 9 3
,N a k at s u k ae tal .1 9 9 8 ).
リンゴでは3
つの ACS が ライブニ ング果実か ら単離 されてい る( R o s e nfi el deta l . 1 9 9 6 )
が , これらの 中で Md‑ACSlのみ ノーザ ンハ イブ リダイゼ ーシ ョンに よる転写解析 が行 わ れている
( D o n ge ta l .1 9 9 1 ).
我 々の ノーザ ン解析 の結果,Md‑ACSlはD o n get a l . ( 1 9 91 )
の報告 同様 , ライブニ ング時特異的 に発現 し,Md‑ACS3は構成的 に 発 現す るこ とが 明 らか となった. しか し,Md‑ACS2の プローブ を得 る こ とが できなか ったため, この遺伝子 の発現 については検討 しなか った.
‑ 7‑
Ⅲ
Md‑ ACSl
の対 立 遺伝 子 型 と リ ンゴ果 実 内エ チ レ ン濃 度 の 関係リンゴ栽培 品種 ̀ゴールデ ンデ リシ ャス'の ライブニ ング時 に発現 す る
A C C
合成酵素遺伝子
( A C S l )
は基本 遺伝子型 であるト1
型 と, この5'
上流域 に レ トロポゾ ンMd‑ SI NEl
が挿入 した対立遺伝子ト2
型の両者がヘ テ ロ型 の状 態 で 存在 す る. この品種 の ク リマ クテ リック果実 にはト1
型の強 い発現 は認 め られ るが,ト2
型 か らの転 写物 は検 出 され ない. また,1‑ 2
型 ホモであ る ̀ふ じ' で は1‑ 2
型 の転 写物 が確 認 され るが ,その転 写量 は低 く, 同時 にク リマ クテ リック時果実 の 内部 エ チ レン量 も少 ない.以上 の こ とか ら ト2
型遺伝 子 は プ ロモー ター領域 の変異 に よってA C Sl
の転写活性 が低下 してい る もの と考 え ら れた.リンゴ果実のエ チ レン生成量 は品種 に よってか な り異 なるこ とが知 られてい る
( C h u1 9 8 8
,G e s s ma netal .1 9 9 3
,毒松 木 ら1 9 9 7 ).
一 方 , リン ゴの栽培 品種 はpcR
法 に よ りMd‑ SI NEl
挿入 の有無 を診 断す ることで,M d‑ A C S l
の1‑
1
型 と1‑ 2
型 の各 ホモ型 お よび両者か らなるヘ テ ロ型 の3
群 に分類 す るこ と が で きる.そ こで ,M d‑ A C S l
の遺伝子型 とライブニ ング時の果実 内エ チ レン量との関係 につ いて検 討 す るため,熟期 が異 なる
3 0
種 以上 の栽培 品種 を対 照 に 果実内エチ レン量 を計測 した. また,A C S l
のヘ テ ロ型 同士 , ̀王林 '×
̀グ ラ ニ ース ミス'の交雑実生群か ら得 られた果実 について も同様 の検 討 を進 めた.材料 お よび方法
1.
実験材料弘前大学 農学部 附属藤 崎農場 お よび青森県 りん ご試験場 で栽植 されてい る リ ンゴ栽培種 を材料 と して使用 した.また, ̀王林 '
×
̀グラニ ース ミス'のF
l個 体 は外観 ,食 味, 日持 ち性等 の優 れた晩生優 良品種 の育種 事業 の一環 と して, 農林水 産省 果樹 試験場 りん ご支場 (盛 岡) にて1 9 8 9
年 に交配 された実生個 体 群 を使用 した. これ らの個体か ら適期 収穫 された果実 は2 0℃
暗所 に1 2
日間 静置す る こ とで, ク リマ クテ リック段 階へ と誘導 し,エチ レン測定の材料 と し た.以上 の調査 は1 9 9 8
年 に行 った.一万,1 9 9 7
年 の栽培種 でのエ チ レン量 測定 には適期収穫 果実 を直 ちに4℃
の大型貯蔵庫 に保管 した もの を材料 と した.1 9 9 7
年1 2 月 8
日に全 品種 を一斉 に出庫 し2 0℃
,1 2
日間静置後 ,エチ レン量 を測定 した.2 .
果実内エ チ レン量 の測定ク リマ クテ リック果実 の内部エチ レン濃度の測定 は果実 の陽光面 とその反対 側 よ り果 肉の一部 を包丁 で切 り取 り (約
3 0 g)
, この ブロ ック をデシケ ー ター内の充分 に脱気 された蒸 留水 中 に沈 め,逆 さに したガ ラス製の ロー トを用 いて 減圧下 で回収 された果 肉内の ガス を注 射器 で
1ml
採 取 し, これ をサ ンプル と した.測定 は FII)‑ガス クロマ トグラフィー (島津 GC‑8A),直径 3mm,長 さ 2 mの活性 アル ミナカラムにて行 った.各 品種 ご と5
個 の果実 を使用 し,その平 均 を測定値 とした.3.Md‑ ACSl
の対立遺伝子型の判定各個 体 の成業 か ら全 DNA を抽 出 し (Varadarajam eta
l .
1992), これ をテ ンプ レー トとして PCR を行 った.用 いた プライマーは下記 に示 した.ACS1‑F 5'AGAGAGATGCCAm TTGTTCGTAC3' 861‑887 ACSLR 5'CcTACAAACTTGCGTGGGGATTATAAGTGT3' 1379‑1350
数値 は AccessionNo.U89156の塩基配列番号 に対応 す る.反応 液 の電気 泳動 像 よ り,増 幅産物 のサ イズの違 いか ら 1‑1型 ,1‑2型 を判定 し,各個体の
ACS l
の遺伝子型 を決定 した.結果
1.ACSl
遺伝子型 の分類Md‑ ACSl‑ 1
の転 写 開始 点 よ り上 流 781bp (この部位 にMd‑ ACSl1 2
で は Md‑sINElが挿入 されてい る) の前後 に設定 した プ ライマ ー (ACS1‑F,ACS1‑氏)を 用 いた pcR 実験 に よ り, リン ゴ栽培 品種 お よび野生種 の
ACSl
の遺伝 子 型 を 判定 した.検定 した材料 の総 てにおいて,基本型 (1‑1型)と sINE挿入型 (1‑2 型 ) に対応 す る 489bp と 655bp の PCR 産物 の どち らか一方 , も し くは両者 が得 られた (図 3‑2).これ らの結果か ら,供試材料 は 1‑1ホモ型 ,1‑1/1‑2ヘ テ ロ型 ,1‑2ホモ型の 3 つ に分類 された.調査 した 11種 の野生種 では 8種 が 1‑1ホモ,3種 が 1‑1/1‑2ヘ テ ロ型 であった.2.
リンゴ栽培 品種 の果実 内エチ レン量ACSl
の遺伝 子 型 と果実 内エ チ レン量 の 関係 を明 らか にす る 目的で ライ ブニ ング果実 内のエチ レン量 を測定 した.熟期 が異 なる( 7
月中旬か ら1 1
月上旬 ま で)栽培 品種 をそれぞれの適期 に収穫 し, さ らにク リマ クテ リック段 階‑ と到 達 させ るために 20℃,12日間静置 した後 に,1品種 につ き 5個 の果実 につ いて エ チ レン量 を測定 した. その平均 値 を示 したのが 図 3‑3であ る. また, ここ で得 られたデー タの一部 を 1997年 の測定値 と対比 させ ,その相 関 を表 した も のが図 3‑4であ る.両年の測定値 には高 い正 の相 関 (r=0.76)が得 られた こ とか ら,各 品種 間の内部エチ レン量 の高低 は遺伝 的要 因に よる もの と判 断 され た. さ らに,両 図か ら判 断 される ように,ACS l
の1‑ 2
型 をホモ型 で有す る品種 は1‑ 1
型 を有す る遺伝子型のそれ らに比べ,明 らか にエチ レン量が低 い こ とが 判 明 した.‑ 9‑
3.F
l個体 による解析̀王林'× ̀グラニース ミス'の
F
l個体群 を用いてエチ レン量の測定 を行 っ た.p c R
で遺伝子型が判定 された1 6
個体 中,1 3
個体 より果実が収穫 された.図
3‑ 5
は Fl個体か ら得 られた果実 ごとの測定値 をプロッ トした ものである.1‑ 2
型 と判定 された個体の果実 は明 らかにエチ レン量が少 なか った. また,ヘテロ 型 の個体 か らの果実 はト1
ホモ型 に比べ てエチ レン量が少 ない傾 向が認 め ら れた.以上の ことか ら,F
l個体群 において も1‑ 2
型 の リンゴは1‑ 1
型 を有す る個体 に比べエチ レン量が少 ないことが明 らかになった.考察
エチ レ㌢生合成系の
A C C
合成酵素遺伝子( M d‑ A C S l )
は対立遺伝子が存在 し, そのA C S l‑ 2
はプロモー ター領域 にMd‑ SI N El
が挿入 した こ とに よって,転写 量 が低下 した変異遺伝子 である もの と考 え られた. さらに,A C S l1 2
の変異遺 伝 子 は転写量 だけではな く,エチ レン量 に も影響す ることが明 らか となった.しか し, ̀ゴールデ ンデ リシャス'や ̀ふ じ' による
1‑ 2
型の転写量 の低下 はsI N E
の挿入が原 因であるとは断定 されない.1‑ 2
型の5'
隣接領域 にはsI N E
挿 入 以外 に も基本 の1‑ 1
型 と異 なる箇所が ある.仮 にそれが発現転写調節 の シ ス 因子 に関与す るボ ックス内の塩基置換 であった とす れば,その ことが転写能 に大 きな影響 を与 えることになる.A C S l‑ 2
の転写量低下 の原 因がSI N E
挿入 の結果で あるこ とを証明す るためには,詳細 なプロモーター解析 が必要 となろう.
果実 内エチ レン量の測定はリンゴの栽培品種 とFl個体群 を対照 とした. リン ゴは他殖 性 であ るため様 々な遺伝子の変異が蓄積 してい る.そ こで, ̀王林 '
× ̀グラニース ミス'の
F
l個体群 を用い,他の遺伝 的背景 を少 な くした実験材 料 を用 いた. この結果 において も,1‑ 2
ホモ型では明 らか にエチ レン量 が低 い とい う結論が得 られた (図3‑ 6).
さらに,1‑ 1 /1‑ 2
へ テロが上1
ホモに比べ エ チ レン量 が低 い傾 向 を示 した ことか ら,A C S l
の発現 は半優 性 であ る もの と 推測 された.トマ トでは
8
つのACS
遺伝子の内,2
つ( L EI A C S 2
,L E‑ A C S 4 )
が ライブニ ングに関与 している( O eti ke reta1 .1 9 97 ).
リンゴの場合,A C S
l,A C S 2
,A C S 3
の
c D NA
が果実か ら単離 されている( La y‑ Ye ea n dX ni gt h o n1 9 9 6
,Ros e nfi el d
etal .1 9 97 ). RN A
ゲ)I,プロッ ト解析 か ら,A C S l
はライブニ ング果実 に特異 的 に発現 してい るの に対 し,A C S 3
は構成 的であることが示 された.A C S 2
を含 め末単離 のACS
遺伝子 の存在 も予想 されることか ら,各A C S
の ライブニ ング 時のエチ レン生成‑ の寄与率 を明 らか にす るには発現解析 を慎 重 に行 う必要が あ ろ う. しか し,AC S l
の遺伝子型 の違 いのみでエチ レン量 に大 きな差が認め られた こ とは,A C S l
の産物が クリマ クテ l)ック果実のエチ レン生成 にお ける主要 な酵素 はであることを示 している.
トマ トでは
A C S
やA C O( A C C
酸化酵素)のア ンチセ ンスR N A
を発現 させ る こ とで ,果実 の 日持 ち性 を大 幅 に高めた( ° el l e reta l . 1 9 91 ).
リンゴにお い て もエ チ レン生成量が抑 え らると, 日持 ち性 (貯蔵性)が高 くなる と予想 さ れ る.実 際,1‑2型 と判定 された ものは ̀ふ じ' をは じめ として,一般 に貯 蔵 性 が良い とされている品種 である (佐藤1 9 9 9 ).
さらに,図3‑ 3
ではA C S l‑ 2
型 がト1
に比べ,晩生の品種 に多い傾向がみ られる.エチ レンの生成量 を抑制す ることで ライブニ ングが遅れ,収穫適期が後退す ることも考 え られることか ら,A C S
遺伝 子 は貯蔵性 だけではな く,収穫 時期 に も影響 している可能性がある.遺伝子型 を分類 した とき (表
1 )
,野生種 では1‑ 1
ホモ型が最 も多 く,ト1 / 1‑ 2
へ テ ロ型 は
3
種 でのみ認め られ,1‑ 2
ホモ型が存在 しなか った. この ことは, リンゴ野生種 では栽培種 に比べ全体的に112ホモ型が少 ない といえる.A C S l
の 変異体 は貯蔵性 には有効 であるが,ライブニ ングが ゆっ くり進行す るために, 香 りが少 な く, また成熟が遅れることか ら,野生種 その ものに とっては,なん ら有益 な形質ではなか った可能性 もある.一方,近年の育成品種 の中には, ̀さ ん さ'や ̀青 り9号'など 1‑2ホモ型が多 く,ト2ホモ型が育種 で優先的に選 抜 されて きた こ とを意味す る. この こ とは逆 に,A C S l
遺伝子 をT ) N A
マーカー とす る こ とで,貯蔵性の ̀良 し悪 し' を選抜 で きることも意味す る. リンゴは 交 配 してか ら結実す るまで数年 を要す ることか ら,果実形質 (果皮色 ,食味, 貯 蔵性 な ど)の調査 には膨大 な時間 と圃場 スペ ース を必要 と した.D N A
マー カ ー に よる貯蔵性診 断は実生 の段 階で可能であることか ら,今後 の リンゴ育種 に お ける有効 な選抜手段 となろう.‑ ll‑
Ⅲ リ ン ゴ各 組 織 にお け る
ACS
の発 現 解 析 お よび新 た に単 離 され たMd‑ ACS5 A , Md‑ ACS5 B
の解 析エチ レンは植 物 の さまざまな成長 を調節す るホルモ ンで あ り, また,傷害 や オーキシ ンな どの処理 に よ り,その生成量が一過的 に増加 す る. さ らに果実 の 追熟 (ライブニ ング) を促進す る作用 は古 くよ り知 られてい る現象 で ある.エ チ レン生 成系 の律 速酵 素 は 1‑ア ミノシ ク ロ プ ロパ ン‑1‑カル ボ ン酸 合 成酵 素
( A C S )
で あ るこ と, また,その遺伝 子 は ファ ミリー を構 成 し各遺伝子 が機能 を 分担 して い るこ とが判 って きた. リンゴで は, これまで3
種類 のA C S
遺伝 子( A C S
l,AC S 2
,AC S 3 )
が いずれ もライブニ ング時の果実か らc D N A
と して単離 さ れ,他 の1
種類 (A C S 4 )
は培養 シュー トでI A A
に よ り誘導 されるこ とがわか っ ているが ,その他 につ いての発現 はほ とん ど報告が ない. リンゴ果実 ではA C S 3
が クライマ クテ リックの前後で比較 的構成的 に発現す るの に対 し
,A C S l
はク ラ イマ クテ リック時 に強 く発現す るこ とを報告 した.本 章ではA C S l
お よびAC S 3
の両
A C S
遺伝子が果実以外の組織 で どの ように発現す るのか を調べ た. また, 傷害 に よるエチ レン生 成 には これ らの遺伝 子が 関与 しなか ったため,A C S
遺伝 子 のd e g e n e r a t ep ri m e r
を用いて新 たなA C S
遺伝子( M d‑ A C S S A
,M d‑ A C S 5 B )
を 単離 し,その発現パ ター ンの解析 も行 った.材料及 び方法
1 .
実験材料及 びエチ レン測定リンゴ栽培 品種 ̀王林 'の培 養 シュー トを用 いた. シュー トは継代培 地
( M S
培 地,
3 %s u c r o s e
,0 . 7m g /16‑ b e nz yl a mi n o p u ri n e
,1 0 0m g/1i n o si t ol
,0. 8
%a g a rp H5 . 8 )
で培養 した.約3
週 間後 の シュー トを試験管 に入 れ,2ml
のb u f f e r ( 5m MM ES
,0. 5 5M
マ ンニ トールp H5 . 8 )
を加 え, シ リコン栓 で密封 し2 5℃
にて静置 させ た.
2
時間 ごとに注射器 で1 ml
を採種 し, ガス クロマ トグラフ ィ ーでエチ レン生成量( n l / g / h r )
を測定 した.2 .5 ' R A C E
前章 と同様 の方法で,エ リシター処理 した成業か ら
R N A
を抽 出 し,1〃g
相 当 のT ot alR N A1 5〃1
を7 0℃
,1 0
分 間熟変性 し氷上 で急冷 した後 ,全量2 0〃1
の逆転 写反応液
( 5 0m MT ri s‑ H Clp H8. 3
,6m MM g C
12,4 0m MK C
l,1m MD T T
,1/ JMr e v e r s ep ri m e r , 1m Md N T P , 5 0u ni t sM u L Vr e v e r s et r a ns c ri pt a s e;P E R KI N E L … E R )
中で,4 2℃
,1
時間反応 させ てc D N A
を合成 した後 ,7 0℃
,1 5
分 で反応を停止 した. この合成 された
c D N A
に6 0u ni t s /〃1
のR N a s e H ( T a K a R a )
を1〃
1
加 え てR N A‑ D N A
ハ イ ブ リ ッ トのR N A
鎖 を分 解 した 後 ,QI A G E N
のP C R
p u ri fi c ati o nKi t
でD N A
を精製 した. これ を2 5/ ∠1
のT d T
反応 液( 0. 1M
カコジル酸 ナ トリウム,
0 . 1m MD T T
,2m MM n C1
2,0 . 4m Md C T P
,1 3u ni t sTe r mi n al D e o x y nu cl e oti d ylTr a ns f e r a s e;Ta K a R a )
中で3 7℃
,1 5
分‑6 5℃
,1 0
分 反応させ
c D N A
の5 '
末端 にC
を付加 させ た. この反応 液5〃1
を全量5 0〃1
のP C R
反応 液
( 1 0m MTri s‑ H Clp H8 . 3
,5 0m MX C
l,1 . 5m MM g C1
2,0 . 2m Md N T P
,0 . 4 F LMOl i g o‑ d Gpri me r , 0 ・ 4
mMr e v e r s epri m e r , 2 ・ 5u ni t sA m pl iT a qG ol d T " ; P E R KI N E L M E R )
中でP C R
反応 を行 った.増幅 したD N A
はさ らに内側 の プライマ ー を用いて
P C R
反応 を行 った.3. T A
クローニ ング増 幅 した
p c R
産物 はp C RP u ri f i c ati o nKi t( QI A G E N )
を用 いて精製 した後 ,T A
ク ローニ ングを行 った.T A
ベ クターはp Bl u e s c ri ptⅡK S 十
をEc o RV
で切 断 した後 , フェノール/クロロフォルム処理 し,その上層 をエ タノール沈殿 した.これ を
4 9〃1
の反応液( 1 0m MTri s‑ H Clp H8 ・ 3
,5 0m MX C
l,1 ・ 5m MM g C1
2,1m M d T TP
,2 . 5u ni t sA m pl iT a qG ol d
T";P E R XI NE L M E R )
中で7 5℃
,2
時間反応 さ せ て ベ ク ター にT
を付 けた .p c R
産物 と, ベ ク ター はD N ALi g a ti o nXi tⅡ
( Ta Xa R a )
を用 いて1 6o C
,一晩 ライゲ‑シ ョン反応 を行 った後 , コロニ ー トラ ンス フ ォーメー シ ョン法 に よって形 質転換 した.形質転換 させ た大腸 菌 はs o c
液体培 地
( 2%
パ ク トトリプ トン,0 . 5%
パ ク トイース トエ キス トラク ト,1 0m M Na C
l,2 . 5m MX C
l,1 0m MM g C1
2,1 0m MM g S
O。,2 0m M
グル コース) で3 7
℃,1
時 間振 盗培 養 を行 った後 ,ア ンピシ リン (終濃 度5 0/ ∠g/ ml )
,X‑ G al
(終濃 度2 0 0F Lg/ ml )
,I P T G
(終濃度2 0 0F Lg/ m
l) をL B
上層寒天培 地( 1 . 00 / .
パ ク トトリプ トン,
0 . 5%
パ ク トイース トエ キス トラク ト,1 . 0% N a C
l,0. 8%
寒天) 上 で3 7℃
一晩培養 し, コロニ ー を形 成 させ , この 内, 白色 コロニ ー を組 換 え 体 と して選抜 した.4.
プ ラス ミ ドD N A
調製目的の プラス ミ ドの入 った大腸菌 を
L B
液体培 地 (終濃度5 0〃g/ ml
ア ンピ シ リン) 中で3 7℃
一晩振盗培養 した.8 , 0 0 0r p m
で5
分 間遠心 し,集菌 した 級 ,上 清 を捨 て沈殿 をT E G ( 2 5m MTri s‑ H Clp H8. 0
,1 0m ME D T A
,5 0m M
グ ル コース) で懸 濁 し, リゾチ ームI T E G ( l on g / ml
リゾチ ー ム) を加 えて撹 拝した. アルカ リ
S D S
溶液( 0 . 2NN a O H
,1 0 / O S D S )
を加 え3M
酢酸 ナ トリウムbH 4. 5 )
を加 え1 0 , 0 0 0r p m1 0
分 間遠 心 した.上清 を回収 し等量 の イ ソプロパ ノール を加 えて,
‑ 2 0℃
に2 0
分 間放置後 ,1 5
,0 0 0r p m
で1 0
分 間遠心 し, プラ ス ミ ドD N A
を沈殿 させ た.沈殿物 をT Eb uf f e r( 1 0m MTri s‑ H Clp H8 . 0
,1m M ED T A)
に溶解 し,R N a s e ( 0 . 1m g / ml )
で3 7℃ 3 0
分 間R N A
を分解 した後 , フェ ノー ル/ク ロロフォルム処理 して,上層 を回収 し,エ タノール沈殿 させ ,
D N A
を
T Eb uf f e r
に洛解 した.5.
シークエ ンス反応‑1 31
プ ラス ミ ド
D N A
は,Se qui Ther m
TMc ycl eSe que nci ng Ki t‑ LC ( f orLトCO R s e que nci ng) ( Epi c e nt erTec h nol ogi es )
を用 いて シー クエ ンス 反応 させ た.シークエ ンス反応物 を
D NA s e que nc erM odel40 00 L ( LI‑ CO R)
に よって読 み と った後 ,Base I ma ge I RS oft war eVer si on 2. 1 0 ( LトCO R)
を用 い て解析 を行 った.6.
サザ ン解析 , ノーザ ン解析 及 びRT‑ P CR
サザ ン解析 , ノーザ ン解析 及 び
RTI P CR
は第2
章 と同様 に行 った.A C S 5
の プ ローブはエ リシター処理 した成葉か ら抽 出 したR N A
をR TI P CR
L,
それ をクロ ーニ ング したA C S 5 A
の プラス ミ ドをp cR
した もの を用いた. このPC R
で用 いた プライマー とR T‑ P CR
に用 いたde ge ner at e
図
4‑ 3
のA C S 5 ノ
=こ対応す る番号である.de ge ner at eF 5' TTYC ARG AYTA Y CAYGG3' de ge ner at eR 5' ACN ARNCC RAARC TNG AC AT3' ACS5 F 5' AC A TCG A A TG C AAC CG CA ACC TTA3' A CS5 R 5' TC TA AG TG G C TCG A AC AAG AG G 3'
pri mer
を下記 に示す.尚,数値 は7 9‑ 95
7 37‑ 71 8 5 92‑ 61 6 1 27 6‑ 1 255
結果
1.ACS
遺伝子の各組織 での発現A C S l
お よびA C S 3
の両ACS
遺伝子が果実以外の組織 で どの ように発現す るの か を調べ た.まず,リンゴの各組織 よ りR NA
を抽 出 し,ノーザ ン解析 を行 った (図4‑ 1 )
.す なわち, クライマ クテ リック時の果実 ,開花後約7 0
日 目の健 全葉 , 種子発芽時の幼根 ,継代培養 の シュー トの茎 , この シュー トの葉 ,結実後約 1ケ月後 の離層形成が認め られた果柄 をそれぞれ,果実,成葉,梶 ,茎 ,幼葉 , 離層 として解析 を行 った.その結果,
A C S l
は果実のみで発現が認 め られ,その 他 の組織 ではシグナルが得 られなか った.一方,A C S 3
をプローブ とした とき, 成葉,幼葉で最 も高 く,茎 での発現 も高か ったが,果実,梶 ,離層では低 い発 現 しかみ られなか った.次 に,成業 をみ じん切 りに した もの を傷害処理 ,細胞壁分解酵 素溶液 (セル ラーゼオノズ カ