「つばきとひきつり」から情報処理へ
―現代連合学習理論の50年―
中 島 定 彦
関西学院大学From “spit-and-twitch” to information processing:
Modern associative learning theories at fifty
Sadahiko Nakajima
Kwansei Gakuin University
This paper reviews the last 50 years of experimental and theoretical research on Pavlovian conditioning in ani-mals. It is the history of the movement from simple “spit-and-twitch” psychology to information processing views of associative learning. In 1962, Egger and Miller reported a pivotal study suggesting that information value is impor-tant in establishing an effective conditioned stimulus. In the late 1960s, Wagner, Rescorla, and Kamin published his-toric research papers demonstrating the importance of information value (i.e., predictability of the forthcoming sig-nificant event) by showing new phenomena in Pavlovian conditioning: relative cue validity, contingency effect, and associative blocking. The Rescorla–Wagner model came on stage in 1972 to explain these phenomena and success-fully predicted new phenomena, although this model had some shortcomings. Subsequent theories of Pavlovian conditioning have challenged to deal with these shortcomings, and the recent theoretical development is linked to computational modeling in a variety of ways.
Keywords: associative learning, Pavlovian conditioning, predictability, mathematical model
は じ め に 2013年はJ. B. Watsonによる「行動主義宣言」(Watson, 1913)からちょうど100年目であった。行動主義は,誕 生後すぐにPavlov(1927)の条件反射やThorndike(1911) の試行錯誤学習の研究を取り込み,動物の条件づけ研究 を主たる領域としつつ新行動主義者らによって他領域に も広がりを見せた。例えば,条件づけ研究の成果をもと に行動療法が編み出された(祐宗・春木・小林,1972)。 つまり,条件づけ研究は行動主義の本丸である。しか し,行動主義宣言以降今日まで一世紀の歴史の後半約 50年は,動物の条件づけ研究においても,刺激や反応 そのものの機械的連合学習ではなく,それらの「表象」 の間の連合学習が情報処理的な視点で論じられるように なった1。本稿では,動物の条件づけ研究におけるこう した学問的潮流について概観するが,紙幅の関係上,古 典的条件づけ(Pavlov型条件づけ,レスポンデント条件 づけ)に限定して紹介する。 情報概念の導入 強化力を持たない中性的な刺激(例えば音)は,生得 的に強化力を持つ餌のような無条件強化子(uncondi-tioned reinforcer)との接近対呈示により,反応を強化す
Copyright 2014. The Japanese Psychonomic Society. All rights reserved. Corresponding address: Department of Psychological Science,
Kwansei Gakuin University, 1–1–115 Uegahara, Nishinomiya, Hyogo 662–8501, Japan. E-mail: [email protected]
1 動物の古典的条件づけ研究において 1960年代に革 新的変化があったが,これを古典的条件づけ研究に おける「認知革命」と呼ぶべきかどうかは判断が難 しい。それ以前にも Tolman(1932)による条件づ けの認知的理論が存在していたし,1940∼1950年代 の心理学に大きな影響を及ぼしたHull(1943, 1952) の機械論的体系でさえ見方によっては認知的理論と いえる(Freeman, 1997)からである。したがって本 稿では 1960年代に古典的条件づけ研究において生 じた新たな動きを認知心理学への転換としてではな く情報処理心理学への変革として捉える。 DOI: http://dx.doi.org/10.14947/psychono.33.6
る力を持つようになる。これを条件強化子(conditioned reinforcer)と呼ぶ。Egger & Miller(1962)は,条件強化 子の成立に重要な要因は中性刺激と無条件強化子の接近 対呈示そのものではなく,中性刺激が無条件強化子の到 来を告げるという予報性(predictability)であることを 示した2。 彼らはラットのレバー押し行動を餌強化子で形成した 後,ラットを2群に分けてレバーのない装置内でFigure 1 左側のような手続きを実施した。第1群のラットには, S1の後にS2を呈示した後,餌を与えるという試行を繰 り返し行った。第2群のラットも同様の手続きを受けた が,S1の単独呈示試行が付加されていた。こうした処置 の後,レバーを取りつけた装置にラットを入れてしばら く餌強化子で反応を維持してから餌強化子を撤去した。 この状況で,餌強化子の代わりにS1またはS2をレバー 押し反応に随伴させて与えることで,S1とS2の反応強 化力をテストした。その結果,第1群ではS1の強化力が S2よりも高かったが,第2群ではほぼ同じ(数値的には S2の方がやや高い)であった(Figure 1右側)。また,S1 の強化力が第1群より第2群で低かった。これは,S1と 餌との関係が前者では100%対呈示,後者では部分的対 呈示となっている(S1が呈示されても餌が呈示される とは限らない)ことによるもので不思議ではない。しか し,S2の強化力が第1群より第2群で高いという事実は, 両群ともS2と餌の関係が同じ100%対呈示であることか ら,新たな説明を必要とする。それが「情報価」であっ た。つまり,第1群では餌の到来はS1によって完全に予 報されるためS2は冗長(情報価がない)である。一方, 第2群ではS1は時間的に先行しているとはいえ餌を伴わ ない試行もあって餌の予報性が低い。それに対して S2 は必ず餌を伴うので情報価を持つことになるから,反応 強化力を獲得することになる。 刺激の予報性
Egger & Miller(1962)の実験において重要な点は,S1 の予報性が低くなることによってS2の情報価が高まるこ とである。この問題は1960年代後半に発表された諸研 究によってさらに注目され,条件づけ研究の最もホット な話題となった。例えば,Wagner, Logan, Haberlandt, & Price(1968)はこれを手がかり妥当性(cue validity)と いう言葉で表現し,3つの実験を行った。Table 1にその うちの1つを示す。2種類の音刺激のうち1種類(Aまた はB)と光刺激(L)をウサギに複合同時呈示してから 眼周辺に微弱電流を流すという眼瞼条件づけ事態(古典 的条件づけの一種)である。無関連群ではAL試行とBL 試行の半数で電流が流され,関連群ではAL試行でのみ 電流が流された。訓練で用いた試行と各刺激要素につい てのテスト結果(眼瞼反射の生起率)をTable 1の右端 に示す。関連群では電流が流されなかったBL試行やそ の要素である刺激Bに対して低反応であり,2種類の音 刺激の弁別ができていることが見て取れる。なお,無関 連群では刺激 Aも刺激Bも比較的低反応であることか ら,音刺激 (A, B) よりも光刺激 (L) のほうが明瞭で あったことが窺える。興味深いのは刺激Lへの反応であ る。刺激Lの電流後続確率は両群ともに50%であるが, 無関連群では高反応であるのに対して関連群では比較的 低反応である。関連群では刺激Aが電流を100%予報す Figure 1. Procedure and results of Egger & Miller (1962, Experiment 1). D.C.=repeat from beginning.
2 彼らの実験はラットのレバー押しを餌強化子で訓練 する状況であるのでオペラント条件づけであるが, 反応の変容・維持過程を研究しようとしたものでは なく,2つの刺激(視聴覚刺激と餌刺激)の関係性 の研究であり,古典的条件づけをテーマとした実験 だといえる。
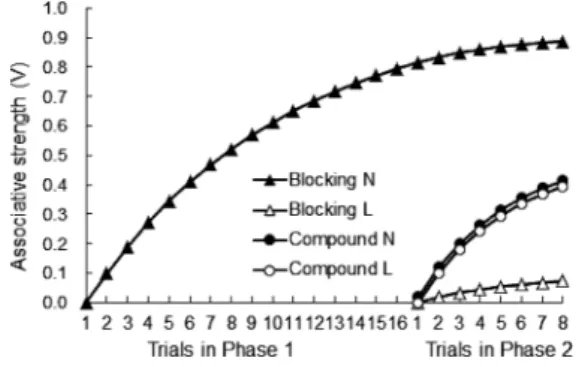
るため,刺激Lは冗長であり,刺激Lは明瞭度が高くて も反応誘発の手がかりとして相対的に妥当でないことが 原因であるとWagner et al.(1968)は論じた。 また,Rescorla(1968)は,ラットの恐怖条件づけ事 態で予報性を検討した実験を報告した3。恐怖条件づけ は古典的条件づけの一種で,視聴覚刺激を条件刺激 (conditioned stimulus, CS),嫌悪事象を無条件刺激(un-conditioned stimulus, US) とし,この2刺激の接近対呈示を 繰り返すことで,CSが恐怖や不安(凍結反応の生起や現在 行っている行動の抑制)といった条件反応(conditioned response, CR)を誘発するようになる。Rescorlaは,音刺 激を CS, 電撃をUSに用いて,CS呈示中のUS呈示確率 (P(US|CS))とCS非呈示中のUS呈示確率(P(US|noCS)) をさまざまに組み合わせた実験を行っている。Figure 2 に10群のラットの反応を示す。P(US|CS)が低くなれば CR が小さくなることは既知であったが,P(US|CS)が 高くてもP(US|noCS)が高ければCRが小さくなること, P(US|CS) とP(US|noCS)が等しければ(つまりCSがUS 到来に関する何らの情報を与えなければ)CRは見られな いことが新たに見出された。CRの大きさがP(US|CS)と P(US|noCS)の両者に依存するというRescorlaの発見は, 現在では随伴性効果(contingency effect)と呼ばれている。 刺激の予報性すなわち刺激が持つ情報価の重要性につ いては, Kamin(1968) による阻止 (ブロッキング, blocking)に関する一連の実験にもみることができる。 Table 2にラットの恐怖条件づけ事態での代表的な実験例 を示す。音(N)と光(L)の同時複合刺激を電撃(US) と対呈示した複合群はテストで単独呈示された刺激Lに 恐怖CRを示すが,複合条件づけ処置に先立って刺激N をUSと対呈示していた阻止群では刺激Lはテスト時に CRをほとんど引き起こさない(刺激Lを訓練時に用い なかった統制群とほぼ同じ値)。阻止群では第 1フェイ ズにおいて刺激NがUSの到来を十分に予報する信号と なるので,第2フェイズで刺激Nと複合呈示された刺激 LはUSの到来に関して新たな情報をもたらさない。つ まり,冗長な刺激Lは情報価を持つ刺激Nによって学習 を阻止される。なお,Table 2の最下段に示した逆行阻止 群の手続きや結果については後に述べる。 Table 1.
Design and results of Wagner et al. (1968, Experiment 3).
Group Training Test
Uncorrelated 304AL → US 304AL → noUS 304BL → US 304BL → noUS AL? (96.8%) BL? (93.8%) A? (21.9%) B? (28.1%) L? (81.2%) Correlated 608AL → US 608BL → noUS AL? (100%)BL? (12.5%) A? (78.1%) B? (9.4%) L? (21.9%) Note. A, B=tone and click, counterbalanced among the group,
L=flash light, US=electric shock. The number before the pair of alphabets indicates the number of compound training trials, and the number in parentheses shows the mean amount of eyelid responses in testing.
Figure 2. Mean amount of fear responses in testing (.50 minus suppression ratio) in 10 groups of rats having different combination of P(US|CS) and P(US|noCS). Created from the results of Rescorla (1968).
Table 2.
Design and results of Kamin (1968, first experiment). Group Phase 1 Phase 2 Test
Control 24N → US L?(.06)
Compound 8LN → US L?(.45) Blocking 16N → US 8LN → US L?(.05) Backward
blocking 8LN → US 16N → US L?(.25) Note. N, L=noise and light, respectively, US=electric shock.
The number before the pair of alphabets indicates the num-ber of training trials, and the numnum-ber in parentheses shows the mean amount of fear responses in testing (.50 minus sup-pression ratio). 3 筆者とRescorlaとの私的会話によれば,彼がスワス モア大学を卒業してペンシルベニア大学の大学院に 入学する際,口頭試問において「条件づけのように 下等な『つばきとひきつり(spit-and-twitch)』の心 理学を研究して,今更どんな意味があるのか」と問 われたという。これは1962年のことである。彼の大 学院での研究はそれに回答を与えるものであった。
Rescorla–Wagnerモデル 手がかり妥当性・随伴性効果・阻止効果といった現象 は古典的条件づけの研究者に,中性刺激がCRを引き起 こす有効なCSとなるには単にその刺激とUSの接近対呈 示だけでは不十分であり,USの到来に関する情報を持 つ必要があるという事実に気づかせた。Kamin(1968) はそれらを「注意」あるいは「注意様過程」という言葉 で表現し意外な結果によって生じる驚きが古典的条件づ けの形成には必要であると論じた。新しい情報をもたら さない冗長な刺激は動物の注意を喚起せず,その刺激は 有効な CSとなることかできない。こうした考え方を 「注意」という概念を直接用いず,USとの連合の強度は 複数のCS間で分配されるという形で数式表現したのが 次に示す Rescorla–Wagner モデル(以下,R–W モデル) である(Rescorla & Wagner, 1972; Wagner & Rescorla, 1972)。
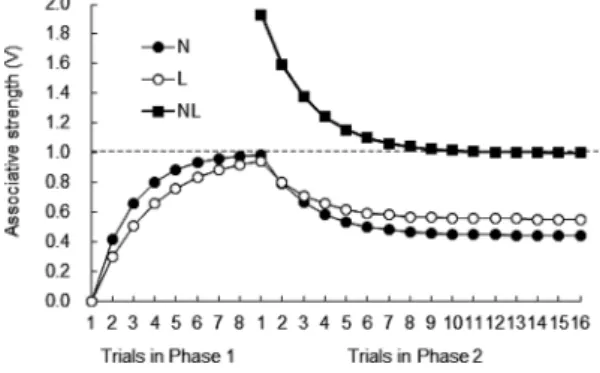
ΔVA=αA β(λ−VAX) ここで,ΔVAは刺激AのUSとの連合強度の1試行あた りの変化量(当該試行で新たに学習する量),αAは刺激A に依存する学習パラメータ (0≤αA≤1) で,一般に刺激の明 瞭度を表す。βはUSに依存する学習パラメータ(0≤β≤1) で一般にUSの強度である。なお,↑β(強化時のβ)≥↓β (消去時のβ) とすることが多い。λはUSが規定する限界 で,強化時はλ=1,非強化時はλ=0とすることが一般的 である。VAXはその試行で呈示される各刺激が持つ連合強 度の総和を示し,これを共有連合原理 (shared associative principle)と呼ぶ4。連合強度の CS間分配はこの原理に よってなされる。 言い換えれば,VAXはその試行で呈示される刺激全体 がUSを予報する程度であり,それとλとのずれ(λ−VAX) は動物が感じる「驚き」に相当する。R–Wモデルの最大 の特徴はこの点にあり,Hull(1943)など旧来の数理モ デルが刺激Aの新たな学習量はλとVA (刺激A自身のそれ までの学習量)との差分に基づいて行われるとしていた ものから,R–Wモデルではλ とVAX(その試行で呈示さ れる全刺激の学習量の総和)との差分に基づくとするこ とで,刺激の情報量を捉えることに成功したのである。 これによってR–Wモデルは前節で述べた3つの効果を 説明することが可能になり,その他さまざまな現象にも 適用された。紙幅の関係上,本稿では2点に絞って簡単 に紹介する。詳しい邦文解説は中島・獅々見(2003)を 参照されたい。 1)阻止効果への適用 Figure 3は先に紹介したKamin (1968)の実験で刺激N と刺激 Lが獲得する連合強度についてR–Wモデルでシ ミュレートしたものである。阻止群では第1フェイズに おいてUSが規定する学習限界近くまで刺激Nが達して いるため,第2フェイズで刺激Lが獲得可能な連合強度 はほとんど残されていない。一方,複合群は第2フェイ ズから始まるので刺激Nも刺激Lも同程度(最終的には USが規定する学習上限の半分ずつ)まで連合強度が獲 得される。したがって,第2フェイズ後のテストにおい て刺激Lが誘発するCRは阻止群よりも複合群において 大になる。 2)過剰予期効果の発見 科学的に優れた理論は既存の諸現象を説明できるだけ でなく,未知の現象の存在を正しく予測できねばならな い。R–Wモデルはそれまで知られていなかったいくつ かの現象の発見に貢献している。例えば,第1フェイズ でA→US試行とB→US試行をそれぞれ複数回実施して, 刺激 A と刺激 B がともに十分な CR を誘発するように なった後,第2フェイズで両刺激を用いて複合条件づけ (AB→US)を繰り返し行う。最後に,刺激Aと刺激Bを それぞれ呈示して CRを測定すると,第2フェイズを行 わなかった場合に比べて各刺激が誘発するCRが小さく なるという現象がある。この現象は,いったん形成され たCRが訓練を継続しているのに減弱してしまうという 極めて直観に反するものであるが,Rescorla(1970)が
Figure 3. Simulation of Kamin’s (1968) blocking ex-periment by the Rescorla–Wagner model with αN=αL=
0.2, β=0.5, λ=1. 4 ここでXは刺激Aと同時に呈示される以外のすべて の刺激を指す。刺激Aを含めて合計2つの刺激が用 いられる場合は刺激Xを意味するが,3つ以上の刺 激を用いた実験ではこの表記は紛らわしい。このた め,VAXではなくVΣと表記されることもある。なお, 刺激が単独で用いられる場合はVAで,共有連合原 理を持たないHull (1943)の習慣強度形成の式など 旧来の数理モデルと実質的に同じである。
ラットの恐怖条件づけ事態で報告5して以降,ラットの食 餌性条件づけ事態(Lattal & Nakajima, 1998),ハトのキー つつき条件づけ(サイントラッキング)事態(Khallad & Moore, 1996), ウ サ ギ の 瞬 膜 条 件 づ け 事 態(Kehoe & White, 2004)などさまざまな古典的条件づけの実験事態 で確認されており,以下に述べる理由により過剰予期効 果(overexpectation effect)と呼ばれている。 Figure 4は,Kremer (1978) がラットの恐怖条件づけ事 態で行った実験を参考例として筆者がR–Wモデルでシ ミュレーションしたものである。第1フェイズ終了時に は音刺激 (N)も光刺激 (L)もλ=1により規定される最 大値にほぼ達している。第2フェイズで刺激Nと刺激L を同時複合呈示するとその連合強度の総和VNLは約2と なるが,US強度には変更がなくλ=1であるから(λ− VAX)はマイナスの値となり,λ=VNLつまりVNL=1とな るまで刺激複合 NLの連合強度は減少する。したがっ て,その構成要素である刺激Nや刺激Lの連合強度も低 下する。ここで両刺激の明瞭度が等しいなら,第2フェ イズ終了時にはVN=VL=0.5となる。これを擬人的に表 現すれば,第1フェイズ終了時に各刺激について十分な US到来予期を形成していたラットは,第2フェイズ開 始時には刺激複合 NLに対して約2倍のUS予期を示す (刺激Nと刺激Lは単独でも怖いので刺激複合NLだと2 倍怖い)が,実際にはこれまでと同強度のUSであるた めこの予期は過剰となり,予期が1となるまで減少修正 されるということである。この考えに従えば,一見する と直観に反するような現象も納得されるであろう。 なお,Figure 4で興味深いのは,第2フェイズ終了時 にN<Lとなっていることである。第1フェイズにおけ る獲得をみるとN>Lであるので,刺激Nは刺激Lより も明瞭であることがわかる。R–Wモデルでは明瞭な刺 激は連合強度の1試行あたりの変化量が大きい。いった ん形成されたCRが訓練を継続しているのに減弱してし まうという過剰予期効果そのものだけでなく,その際に 強い刺激の減弱が大きい(弱い刺激の方が大きなCRを 誘発する)という直観的にはさらに理解しづらい特徴も R–Wモデルは正しく予測する。 その後の理論的発展 R–Wモデルは比較的単純な数理モデルであるにもかか わらず古典的条件づけにかかわる多くの現象を説明でき るため,米国の学習心理学の教科書では古典的条件づけ の章で最もページを割いて解説されている(今田・中島, 1995)。しかし,R–Wモデルは問題点も数多く抱えてい る (Miller, Barnet, & Grahame, 1995; Walkenbach & Haddard, 1980)。邦文解説として中島・獅々見 (2003) を参照され たい。ここでは,その後の理論的発展とのかかわりに着 目して4つに限って取り上げる。 1)制止学習をめぐる諸現象 R–Wモデルは消去をCS–US連合強度の喪失,つまり 学習解除であると捉える。したがって,消去後の時間経 過によって生じるCRの自然回復(自発的回復)を説明 できない。自然回復はPavlov(1927)の研究室で発見さ れて以降,多くの実験で確認されている極めて頑健な現 象であり (Rescorla, 2004),それを説明できないのは古典 的条件づけの理論として致命的ともいえる。 また,R–Wモデルでは,CSとUS間のプラスの連合強 度(興奮連合)だけでなくマイナスの連合強度(制止連 合) を想定するが,制止連合は興奮連合と同じ式に従うと されている (Wagner & Rescorla, 1972)。例えば,Figure 5の 第1フェイズは音刺激(N)にはUSが後続するが,音と 光の刺激複合 (NL) にはUSが後続しないというPavlovの 条件制止手続きによって,刺激Nがプラスの連合強度, 刺激 Lがマイナスの連合強度を獲得することをR–Wモ デルで示したものである。制止性の CS(この場合は刺 激L)は一緒に形成された興奮性のCS(この場合は刺激 N)のCR誘発を抑制するだけでなく,他の場面で形成 された興奮性CSによるCR誘発も抑制し,刺激L自身が 興奮性CSとなることも遅らせる。こうした制止性CSの 特徴をR–Wモデルは容易に説明できる。しかしながら, R–Wモデルは制止性CSを単独で呈示すればマイナスの Figure 4. Simulation of Kremer’s (1978)
overexpecta-tion experiment by the Rescorla–Wagner model with
αN=0.7, αL=0.5, ↑β=0.6, ↓β=0.3, λ=1. 5 最初の実験報告(Rescorla, 1970)やそれに続く報告 (Wagner, 1971) はR–Wモデルの出版年よりも早いが, 同モデルは出版の3年前にマクマスター大学で開催 された会議で発表されており,上記2報告の中でも 同モデルからの新たな予測として述べられている。
連合強度は消失すると予測するが(Figure 5の第2フェ イズ),実際にはマイナスの連合強度は維持されるか (例えば,Zimmer-Hart & Rescorla, 1974),むしろ強まる (例えば,DeVito & Fowler, 1987)。
このほか,R–Wモデルでは単一の刺激が興奮的特徴と 制止的特徴を同時に有することはできない(1つの刺激 は1つの連合強度しか取り得ないため)が,実際には両 特徴を持つ刺激を形成することは可能である(例えば, Tait & Saladin, 1986)。また,他の刺激の興奮連合が消失 すると過去にそれとともに訓練されていた刺激の制止連 合が自動的に弱まるという報告(例えば,Lysle & Fowler, 1985)もR–Wモデルでは説明できない。これらの発見 によって,R–Wモデル以降の古典的条件づけの理論の 多く(ただし,後述のコンパレータ仮説を除く)は,興 奮連合とは独立のメカニズムとして制止連合を想定する ことになった。 2)CS事前呈示効果 古典的条件づけにおいてCS→USの接近対呈示訓練に 先立って CSを事前に単独で経験させておくと訓練に よって生じる CR獲得が遅れる。このCS事前呈示効果 (CS-preexposure effect)は潜在制止(latent inhibition)と も呼ばれ,Lubow & Moore(1959)によってヤギやヒツ ジの屈曲反射条件づけ事態で報告されて以降,多くの動 物種においてさまざまな条件づけ事態で確認されている (Lubow, 1989)。何の情報ももたらさない冗長なCSには 注意が向けられなくなる(無視するようになる)ため, そのCSが情報価を持つ(USを予報する)ようになって も,ただちに連合学習が進まないと解釈されることが多 い。前述のように,R–Wモデルの特徴は複数のCS間の 相互作用を「注意」という概念を用いず,USとの連合 強度が複数のCS間で分配されるとした点にある。この ため,USを呈示せずCSだけ呈示してもCS–US連合強度 は変化しない(連合強度はゼロのままである)ので,R–W モデルではCS事前呈示効果が説明できない。R–Wモデ ルでCS事前呈示効果を説明するための最も簡便な方法 は,単独呈示を行うとCSの明瞭度α が低下するとみな すことである(Frey & Sears, 1978)。
しかし,CSの明瞭度が試行ごとに変化すると考えると, USとの連合強度が複数のCS間で分配されるという仮定 は必ずしも必要ではない。Mackintosh (1975) の注意モデ ルはこの立場に基づいている。ある試行でUSをよく予 報した(情報価の高い)CSは明瞭度が上昇し,次の試行 における連合強度の変化が大きくなる。逆に,冗長だっ たCSは明瞭度が低くなり,次試行での連合強度変化が 小さくなる。言い換えれば,情報価の高いCSは注意さ れ,低いCSは無視される。このモデルでは,例えば前 述の阻止効果は次のように説明される。まず,第1フェ イズで刺激NはUSに関する唯一の情報源であるため注 意が払われ,その連合強度は増加する。第2フェイズで 追加される刺激Lについては冗長であることが第1試行 終了時に判明するため,それ以降は注意が払われず条件 づけが妨げられる。 Mackintosh(1975)の注意モデルでは情報価の高いCS への注意は条件づけ手続きが変わらない限り維持される が,常に同じUSを予報し続けると知覚処理が自動化し, 積極的な注意が向けられなくなることに着目したのが Pearce & Hall (1980)のモデルであり,この2つのモデル を融合する試みも行われている (Esber & Haselgrove, 2011; Le Pelley, Esber, & Haselgrove, 2012; Pearce & Mackintosh 2010)。 一方,Wagner(1978)はプライミング理論を提唱して 事前呈示効果を説明した。実験装置に入れられた動物は そこで繰り返しCSの単独呈示を受ける。これによって 実験装置文脈とCSの連合が生じるので,その装置内で 呈示されたCSは驚きを喚起しなくなる(CSの馴化が生 じる)。このためCS→USの対呈示が開始されてもCSは 十分な処理がなされず,条件づけの進行が遅れるのであ る。プライミング理論は共有連合原理についてはR–Wモ デルを踏襲しつつ,条件づけ試行内の事象の情報処理過 程については記憶の二重貯蔵モデル (Atkinson & Shiffrin, 1968)を参考にして,リハーサルや検索といった概念を 取り込んだ理論であり,その後,情動の相反過程理論 Figure 5. Simulation of conditioned inhibition training
with N→US (reinforcement) trials and NL→noUS (nonreinforcement)trials by the Rescorla–Wagner
model with αN=αL=0.8, ↑β=0.5, ↓β=0.3, λ=1 for
reinforcement and λ=0 for nonreinforcement. The re-inforcement and nonreinfocement trials are alternately presented until Trial 30. The Rescorla–Wagner model successfully predicts acquisition of conditioned inhibi-tion for L, but it incorrectly predicts extincinhibi-tion of conditioned inhibition when L is presented alone with no US for the last 10 trials.
(opponent-process theory: Solomon & Corbit, 1974)を吸収 してSOP(standard operating procedure in memory)理論 (Wagner, 1981) となり,さらにAESOP (affective extension
of SOP)理論(Wagner & Brandon, 1989)へと発展した。
3)形態化学習
刺激Aと刺激BはそれぞれUSを伴うが,刺激複合AB はUSを伴わないという手続き (A→US, B→US, AB→noUS) をランダムな順序で繰り返すと,次第に動物はA試行と B試行で反応し,AB試行では反応しなくなる。しかし, R–Wモデルにしたがえば,この負パターニング(nega-tive patterning)課題を動物は解決できないはずである。 なぜなら,学習の最終段階でVA=1, VB=1, VAB=0となる はずであるが,共有連合原理によりVAB=VA+VBであるの で,0=1+1となってしまうからである。そこでRescorla (1972a, 1973) とWhitlow & Wagner (1972) は,Spence (1952)
に倣って刺激複合では刺激間相互作用によって特有の形 態刺激(unique configural cue)が生じると仮定した(た だし,形態刺激の明瞭度は低いため,負パターニング課 題などの特殊な弁別学習以外の複合条件づけ事態ではこ の要因は無視できるとした)。これによってR–Wモデル で負パターニング課題の学習を説明できることになっ た。つまり,刺激複合ABをABqとみなせば,学習の最 終段階でVA=1, VB=1, VABq=0となるので,形態刺激qの 連合強度をVq=−2とすればよいことになる。 負パターニング課題をより単純に説明しようとするな ら,刺激複合 ABは刺激Aと刺激Bと形態刺激qからな ると仮定するよりも,刺激複合AB全体を1つの形態刺 激と見なすほうが単純である(ただし,刺激Aや刺激B は形態刺激ABと類似しているので般化要因に配慮する 必要がある)。Pearce(1987, 1994)の提唱する刺激形態 化/般化理論(Pearceモデル)はそうした考えに基づい ており,共有連合原理を用いずに,古典的条件づけの諸 現象に対して R–Wモデルとほぼ同等の説明力を持つ。 一例としてTable 3にPearceモデルによる阻止効果の説明 を示す。 しかし,PearceモデルはしばしばR–Wモデルと異なっ た結果を予想する。例えば,刺激AはUSを伴うが,刺 激複合ABはUSを伴わず,刺激複合ABCはUSを伴うと いう弁別訓練(A→US, AB→noUS, ABC→US)を行うと, 動物はA試行とABC試行で反応し,AB試行では反応し なくなる。弁別が完成した後,C試行とBC試行でテス トする(この2つは動物が初めて経験する試行である)。 R–Wモデルでは弁別訓練後にVA=1, VAB=0, VABC=1となっ ているはずであるから,VB =−1, VC=1であり,ゆえに VBC=0となる。したがってC試行で大きなCRが生じ,BC 試行ではCRは生じないことが予想される。一方,Pearce モデルではFigure 6に示すようにVBC=0.73, VC=0.60で,BC 試行がC試行よりやや大きなCRを誘発することを予想 する 。実験の結果はPearceモデルに合致していた(Naka-jima, 1997)。 また,刺激 AはUSを伴うが,刺激複合ABはUSを伴 わず,刺激複合BCはUSを伴うという弁別課題(A→US, Table 3.
Prediction of blocking effect by the Pearce model.
Group Phase 1 Phase 2 Test
Compound NL → US L?(0.50)
Blocking N → US NL → US L?(0.25)
Pearce model calculation
[Group Blocking]
VN comes to 1.
EN also comes to 1, because there is no gener-alization from other stimuli (eN=0).
VNL comes to 1.
VNL=ENL+eNL
[Group Compound]
ENL comes to 1, because there is no generalization from other stimuli.
[Group Blocking]
ENL comes to 0.5, because generalization from E(eN NL)
is 0.5.
VL=EL+eL
Because EL=0,
generaliza-tion from a similar stimu-lus (ENL) directly
deter-mines the amount of CR. [Group Compound]
Generalization from ENL(eL)
is 0.5
[Group Blocking] Generalization from ENL(eL)
is 0.25
Note. VN, VL, and VNL=net associative strength of Stimuli N, L, and NL, respectively. EN, EL, and ENL=proper associative strength of
Stimuli N, L, and NL, respectively. eN, eL, and NL=generalized associative strength of Stimuli N, L, and NL, respectively. The
amount of generalization (e) is the product of the inter-stimulus similarity and the proper associative strength of the similar stim-ulus. The similarity between Stimuli N (or L) and NL is 0.5.
AB→noUS, BC→US)の場合,訓練後にはVA=1, VAB=0, VBC=1となるはずであるから,R–WモデルではVB=−1, VC=2となり,C試行ではA試行やBC試行よりも大きな CRを誘発するはずである。一方,Pearceモデルに従っ て計算するとVC=0.64となり中程度のCRとなる。実験
結果(Pearce & Wilson, 1990)はこれを支持した6。
4)回顧的再評価 Table 1の最終行に示した群のラットは,第1フェイズ で音刺激(N)と光刺激(L)の同時複合刺激を電撃(US) と対呈示する訓練(NL→US)をまず行ってから,第2 フェイズで刺激 NとUSを対呈示する訓練(N→US)を 受けた。その結果,テスト時に刺激Lが誘発するCRは 統制群と比べて小さかった(逆行阻止効果)。しかし, NL→US 訓練からテストまでの日数が統制群よりも長 かったことがこの群差の原因であるとKamin(1968)は 考察した。つまり,逆行阻止効果は真正のものではな く,アーティファクトに過ぎないと結論したことにな る。その後,多くの研究者により逆行阻止効果の実証が 行われたが,そのほとんどは失敗に終わった(1990年代 半ばまでの研究展望としてNakajima & Kawai(1997)を 参照されたい)。しかし,その後Miller & Matute (1996)
がUSの代わりに生物的重要性の低い刺激を用いること で逆行阻止効果を確実に示すことに成功し,Urushihara & Miller(2010)はテスト刺激のCR誘発力を第2フェイ ズ時に弱くしておくことで通常の恐怖条件づけ事態にお いても逆行阻止効果を示すことに成功した。 また,NL→US訓練後にN→US訓練を行うことで刺激 L の誘発する CR が弱まるという逆行阻止効果に比べ, NL→US訓練後にN→noUS訓練を行うことで刺激Lの誘 発する CRが強まるという隠蔽解除(unovershadowing) 効果は容易に観察される (例えば,Matzel, Schachtman, & Miller, 1985)。逆行阻止や隠蔽解除といった現象は,CSの 反応誘発力が他のCSの価値変化によって逆行性の干渉 を受けることを示しており,まとめて負の回顧的再評価 (negative retrospective revaluation)と呼ばれる7。R–Wモ
デルでは呈示されていないCSの連合強度は変化しない ので,こうした現象を説明することはできない。この問 題を克服するためにR–Wモデルの改訂(Van Hamme & Wasserman, 1994) や,SOP理論の改訂 (Dickinson & Burke, 1996)が行われているが,そうした試みとは異なる視点 で説明するのがコンパレータ仮説(Miller & Matzel, 1988) である。 R–Wモデルをはじめとして,これまで紹介してきた すべての理論モデルはCS間の競合現象を学習(連合獲 得)の問題(情報価の高いCSが低いCSの学習を妨げる) と考えるのに対し,コンパレータ仮説では接近対呈示だ けが学習の要件であるとする。阻止効果などのCS間の Figure 6. Graphical representations of the Pearce
mod-el’s predictions for Experiment 1 of Nakajima(1997). Adapted from Nakajima (1998) with some modifcatri-ons. The similarity of two unitary stimuli is equal to the proportion of shared elements between them. For example, the similarity between AB and BC is 1/4, be-cause each of them shares a half of all elements (i.e., 1/2 by 1/2 results in 1/4). The similarities of stimuli are in boxes. The net associative strength of each unitary stimulus is shown below the stimulus, and its own as-sociative strength is in brackets.
6 この種の弁別課題を拡張するとR–Wモデルはとんで もない予測をしてしまう。例えば,7つの刺激A∼G を用い,「A→US」「AB→noUS」「BC→US」「CD→noUS」 「DE→US」「EF→noUS」「FG→US」の各試行をラン ダム順序で繰り返すと,訓練後にはVA=1, VAB=0, VBC=1, VCD=0, VDE=1, VEF=0, VFG=1になるはずなの で,VB=−1, VC=2, VD=−2, VE=3, VF=−3, VG=4とな る。つまり,A試行でのCRに比べてC試行では2倍, E試行では3倍,G試行では4倍の正の連合強度にな り,制止についてもBよりDは2倍,Eは3倍の負の 連合強度になる。これは極めて非現実的である。 7 これに対し,CSの反応誘発力が他のCSの価値変化 によって促進する場合を正の回顧的再評価と呼ぶ。 回顧的再評価全般に関する解説は澤(2007)に詳し い。正の回顧的再評価は古くから2次条件づけなど の媒介過程により説明されてきたため,R–Wモデル にとって脅威ではなかった。これに対して負の回顧 的再評価はR–Wモデルの基盤を揺るがす重要な現象 であることから,単に「回顧的再評価」といえば負 の回顧的再評価のことを指すことが多い。なお,回 顧的再評価と推論の関係について川合・久保(川合) (2008)も参照されたい。
競合現象は,連合獲得時ではなくCR表出時の問題であ ると考えるのである。Figure 7にコンパレータ仮説の概 念図を示す。CSによるCR誘発量は,そのCS(ターゲッ トCS)と US との連合強度(ターゲットCS が直接喚起 するUS表象の強さ)だけでなく,ターゲットCSと共生 起したことのある他のCS(コンパレータCS)を介して 間接的に喚起されるUS表象の強さにも依存する。前者 が後者をはるかに凌駕していれば大きなCRが出力され るが,間接的に喚起されるUS表象が強ければCR出力は 小さくなる。 例えば,阻止効果ではL–USの連合学習とともにN–US の連合学習も生じる。また,LとNは同時呈示されてい るのでL–N間の連合学習も生じる。テストで刺激Lが呈 示されると,L–US連合によりUS表象が直接喚起される が,L–N連合も賦活し,さらにN–US連合が賦活して, US表象が間接的に喚起される。N→USの訓練が十分に なされていればN–US連合は強固であり,したがって間 接的に喚起されるUS表象も強い。したがって,L–US連 合が直接喚起するUS表象の影響が小さくなり,誘発さ れるCRが小さなものとなる。コンパレータ仮説はこう した説明を行うために,NL→US訓練後にN→US訓練を 行う場合(逆行阻止)も,N→US訓練後にNL→US訓練 を行う場合(順行阻止)と同様に,CR低下が生じると 予想する。 コンパレータ仮説は,その後Williams (1996) の批判や Miller自身の研究チームでの実験結果を経て,比較過程 を2重にした拡張版コンパレータ仮説 (Denniston, Savas-tano, & Miller, 2001)に発展し,さらに数式での表現を 実装して SOCR (sometimes-competing retrieval)モデル (Stout & Miller, 2007)に進化して,古典的条件づけにお けるさまざまな新現象の予測に成功している(Miller & Witnauer, 2012)。 お わ り に 本稿では行動主義の1世紀の歴史の後半に生じた,条 件づけにおける単純機械論から情報処理論への移行を概 観した。紙幅の関係で各理論については簡単な説明にと どまらざるを得なかった。また,読者の理解を容易にす るため,情報概念の導入から予報性に関する諸現象の発 見,それを説明するために生み出された R–Wモデル, それが抱える問題点をきっかけとした理論的発展,とい う流れで論を構成したため,R–Wモデル以外の各理論 間の関係について紹介できず,R–Wモデルと各理論との 接点もそれぞれ1つのテーマに絞らざるを得なかった。 実際のところ,各理論は古典的条件づけの諸現象をそれ ぞれユニークな形で説明し,R–Wモデルと多くの接点を 持つ。詳しくは今田・中島(2003)の解説書を参照され たい。また,中島(2013)は連合学習の流れについて, 本稿と一部重複しながらやや異なった,より広い視点で 論じているので併読されたい。 連合学習研究における情報処理的視点は半世紀の歴史 を持つ。その誕生から間もない頃,Rescorla(1972b)は 古典的条件づけにおける情報変数の重要性について当時 最新のデータに基づいて論じた。半世紀の歴史の半ば (つまり,今から四半世紀前)に,Rescorla(1988)は古 典的条件づけに関する心理学界の固陋な見識を嘆き,啓 蒙論文を著した。なお,その前年には,古典的条件づけの 連合構造を明らかにするために開発された研究法を道具 的条件づけ(オペラント条件づけ)研究に適用した諸実 験を平易に紹介した論文も発表している (Rescorla, 1987)。 連合学習研究のこの半世紀の歩みは心理学界において いまだ十分に認識されているとは言えないが,古典的条 件づけに関する連合理論は着実に発展している。理論は 精緻になり,融合し,計算モデル構築(computational modeling)への進化が始まっている。古典的条件づけを 研究する実験心理学者が計算モデルを本格的に用い始め たのは形態化学習をコネクショニストモデルで説明しよ うとしたKehoe (1988, 1989) がおそらく最初だと思われる が,その後,多彩なモデル化の試みがWagnerら (Wagner, 2008; Wagner & Brandon, 2001), McLaren & Mackintosh (2000, 2002), Schmajukら(Buhusi & Schmajuk, 1999; Grossberg & Figure 7. The comparator hypothesis. The amount of
conditioned responses (CR) elicited by the target con-ditioned stimulus (CS) is determined not only by the representation of the unconditioned stimulus (US) di-rectly evoked by the target CS (Link 1) but also by the indirect US representation evoked by the association between the target CS and its comparator CS (Link 2) and the association between the comparator CS and the US representation (Link 3). Adapted from Miller and Matzel (1988).
Schmajuk, 1989; Kutlu & Schmajuk, 2012; Schmajuk & Buhusi, 1997; Schmajuk & DiCarlo, 1991; Schmajuk, Lam, & Gray, 1996), Harris(2006)によってなされてきた。神経科学やコン ピュータサイエンスの研究者を巻き込みつつ進化する古 典的条件づけの計算モデルの現状については,近年出版さ れた専門書Computational models of conditioning (Schmajuk, 2010) やLearning & Behavior誌の特集号 (Alonso & Schmajuk, 2012)を参照されたい。
引用文献
Alonso, E., & Schmajuk, N. (2012). Computational models of classical conditioning: Guest editors’ introduction.
Learn-ing & Behavior, 40, 231–240.
Atkinson, R. C., & Shiffrin, R. M. (1968). Human memory: A proposed system and its control processes. In K. W. Spence & J. T. Spence (Eds.), The psychology of learning and motivation,
Vol. 2. New York: Academic Press, pp. 89–195.
Buhusi, C. V., & Schmajuk, N. A. (1999). Timing in simple conditioning and occasion setting: A neural network approach.
Behavioural Processes, 45, 33–57.
Denniston, J. C., Savastano, H. I., & Miller, R. R. (2001). The extended comparator hypothesis: Learning by contiguity, responding by relative strength. In R. R. Mowrer & S. B. Klein (Eds.), Handbook of contemporary learning theories. Mahwah, NJ: Erlbaum, pp. 65–117.
DeVito, P. L., & Fowler, H. (1987). Enhancement of conditioned inhibition through extinction training. Animal Learning &
Behavior, 15, 448–454.
Dickinson, A., & Burke, J. (1996). Within-compound associa-tions mediate retrospective revaluation of causality judge-ments. Quarterly Journal of Experimental Psychology, 49B, 60–80.
Egger, M. D., & Miller, N. E. (1962). Secondary reinforcement in rats as a function of information value and reliability of the stimulus. Journal of Experimental Psychology, 64, 97–104. Esber, G. R., & Haselgrove, M. (2011). Reconciling the influence
of predictiveness and uncertainty on stimulus salience: A model of attention in associative learning. Proceedings of the
Royal Society B, 278, 2553–2561.
Freeman, J. E. (1997). Pavlov in the classroom: An interview with Robert A. Rescorla. Teaching of Psychology, 24, 283–286. Frey, P. W., & Sears, R. J. (1978). Model of conditioning incor-porating the Rescorla–Wagner associative axiom, a dynamic attention process, and a catastrophe rule. Psychological
Review, 85, 321–340.
Grossberg, S., & Schmajuk, N. A. (1989). Neural dynamics of adaptive timing and temporal discrimination during asso-ciative learning. Neural Networks, 2, 79–102.
Harris, J. A. (2006). Elemental representations of stimuli in associative learning. Psychological Review, 113, 584–605. Hull, C. L. (1943). Principles of behavior. New York: Appleton. Hull, C. L. (1952). A behavior system. New Haven, CT: Yale
University Press.
今田 寛・中島定彦(1995).学習心理学テキスト(英 語)に見る近年の古典的条件づけ理論の傾向 基礎心 理学研究,13, 97–104.
(Imada, H., & Nakajima, S. (1995). Recent trends in theo-ries of classical conditioning: Analyses of textbooks on learning published during the last decade. Japanese Journal
of Psychonomic Science, 13, 97–104.)
今田 寛(監修)・中島定彦(編)(2003).学習心理学 における古典的条件づけの理論―パヴロフから連合学 習研究の最先端まで― 培風館
(Imada, H., & Nakajima, S.)
Kamin, L. J. (1968). Attention-like processes in classical condi-tioning. In. M. R. Jones (Ed.), Miami Symposium on the
pre-diction of behavior: Aversive stimulation. Coral Gables, FL:
University of Miami Press, pp. 9–31.
川合伸幸・久保(川合)南海子(2008).ヒトと動物の 回顧的推論について 認知科学,15, 378–391. (Kawai, N., & Kubo-Kawai, N. (2008). Retrospective
infer-ence in humans and nonhuman animals. Cognitive studies:
Bulletin of the Japanese Cognitive Science Society, 15, 378–
391.)
Kehoe, E. J. (1988). A layered network model of associative learning: Learning-to-learn and configuration. Psychological
Review, 95, 411–433.
Kehoe, E. J. (1989). Connectionist models of conditioning: A tutorial. Journal of the Experimental Analysis of Behavior,
52, 427–440.
Kehoe, E. J., & White, N. E. (2004). Overexpectation: Response loss during sustained stimulus compounding in the rabbit nictitating membrane preparation. Learning & Memory, 11, 476–483.
Khallad, Y., & Moore, J. (1996). Blocking, unblocking and overexpectation in autoshaping with pigeons. Journal of the
Experimental Analysis of Behavior, 65, 575–591.
Kremer, E. F. (1978). The Rescorla–Wagner model: Losses in associative strength in compound conditioned stimuli. Journal
of Experimental Psychology: Animal Behavior Processes, 4,
22–36.
Kutlu, M. G., & Schmajuk, N. A. (2012). Solving Pavlov’s puzzle: Attentional, associative, and flexible configural mechanisms in classical conditioning. Learning & Behavior, 40, 269–291. Lattal, K. M., & Nakajima, S. (1998). Overexpectation in appe-titive Pavlovian and instrumental conditioning. Animal
Learning & Behavior, 26, 351–360.
Le Pelley, M. E., Haselgrove, M., & Esber, G. R. (2012). Model-ing attention in associative learnModel-ing: Two processes or one? Learning & Behavior, 40, 292–304.
Lubow, R. E. (1989). Latent inhibition and conditioned attention
theory. Cambridge: Cambridge University Press.
Lubow, R. E., & Moore, A. U. (1959). Latent inhibition: The ef-fect of non-reinforced preexposure to the conditioned stim-ulus. Journal of Comparative and Physiological Psychology,
52, 415–419.
Lysle, D. T., & Fowler, H. (1985). Inhibition as a “slave pro-cess”: Deactivation of conditioned inhibition through ex-tinction of conditioned excitation. Journal of Experimental
Psychology: Animal Behavior Processes, 11, 71–94.
Mackintosh, N. J. (1975). A theory of attention: Variations in the associability of stimuli with reinforcement. Psychological
Review, 82, 276–298.
Matzel, L. D., Schachtman, T. R., & Miller, R. R. (1985). Recov-ery of an overshadowed association achieved by extinction of the overshadowing stimulus. Learning and Motivation,
16, 398–412.
McLaren, I. P. L., & Mackintosh, N. J. (2000). An elemental model of associative learning: I. Latent inhibition and per-ceptual learning. Animal Learning & Behavior, 28, 211–246. McLaren, I. P. L., & Mackintosh, N. J. (2002). Associative
learning and elemental representation: II. Generalization and discrimination. Animal Learning & Behavior, 30, 177–200. Miller, R. R., Barnet, R. C., & Grahame, N. J. (1995).
Assess-ment of the Rescorla–Wagner model. Psychological Bulletin,
118, 363–386.
Miller, R. R., & Matute, H. (1996). Biological significance in forward and backward blocking: Resolution of a discrepancy between animal conditioning and human causal judgment.
Journal of Experimental Psychology: General, 125, 370–386.
Miller, R. R., & Matzel, L. D. (1988). The comparator hypothe-sis: A response rule for the expression of associations. In G. H. Bower (Ed.), The psychology of learning and motivation,
Vol. 22. San Diego, CA: Academic Press, pp. 51–92.
Miller, R. R., & Witnauer, J. E. (2012). Comparator hypothesis of associative learning. In N. M. Seel (Ed.), Encyclopedia of
the sciences of learning Vol. 3. Heidelberg, Germany: Springer,
pp. 661–665.
Nakajima, S. (1997). Failure of inhibition by B over C after A+, AB−, ABC+training. Journal of Experimental Psychology:
Animal Behavior Processes, 23, 482–490.
Nakajima, S. (1998). Further investigation of responding elicit-ed by BC and C after A+, AB−, ABC+ training. Quarterly
Journal of Experimental Psychology, 51B, 289–300.
中島定彦(2013).連合学習理論 藤永 保(監修) 最 新心理学事典 平凡社 pp. 747–751.
(Nakajima, S.)
Nakajima, S., & Kawai, N. (1997). Failure of retrospective in-ference in the rats’ taste aversion. Japanese Psychological
Research, 39, 87–97.
中 島 定 彦・ 獅 々 見 照(2003).Rescorla–Wagner モ デ ル 今田 寛(監修)・中島定彦(編) 学習心理学におけ る古典的条件づけの理論―パヴロフから連合学習研究 の最先端まで― 培風館 pp. 31–58.
(Nakajima, S., & Shishimi, A.)
Pavlov, I. P. (1927). Conditioned reflexes (G. V. Anrep, Transl.). London: Oxford University Press.
Pearce, J. M. (1987). A model of stimulus generalization in Pavlovian conditioning. Psychological Review, 94, 61–73. Pearce, J. M. (1994). Similarity and discrimination: A selective
review and a connectionist model. Psychological Review,
101, 587–607.
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian condi-tioning: Variations in the effectiveness of conditioned but not of unconditioned stimuli. Psychological Review, 87, 532–552.
Pearce, J. M., & Mackintosh, N. J. (2010). Two theories of at-tention: A review and a possible integration. In C. J. Mitch-ell & M. E. Le PMitch-elley (Eds.), Attention and associative
learn-ing: From brain to behaviour. Oxford: Oxford University
Press, pp. 11–40.
Pearce, J. M., & Wilson, P. N. (1990). Configural associations in discrimination learning. Journal of Experimental Psychology:
Animal Behavior Processes,16, 250–261.
Rescorla, R. A. (1968). Probability of shock in the presence and absence of CS in fear conditioning. Journal of Comparative
and Physiological Psychology, 66, 1–5.
Rescorla, R. A. (1970). Reduction in the effectiveness of rein-forcement after prior excitatory conditioning. Learning and
Motivation, 1, 372–381.
Rescorla, R. A. (1972a). “Configural” conditioning in discrete-trial bar pressing. Journal of Comparative and Physiological
Psychology, 79, 307–317.
Rescorla, R. A. (1972b). Informational variables in Pavlovian conditioning. In G. H. Bower (Ed.), The psychology of learning
and motivation, Vol. 6. New York: Academic Press, pp. 1–46.
Rescorla, R. A. (1973). Evidence for unique stimulus account of configural conditioning. Journal of Comparative and
Physiological Psychology, 85, 331–338.
Rescorla, R. A. (1987). A Pavlovian analysis of goal-directed behavior. American Psychologist, 42, 119–129.
Rescorla, R. A. (1988). Pavlovian conditioning: It’s not what you think it is. American Psychologist, 43, 151–160. Rescorla, R. A. (2004). Spontaneous recovery. Learning &
Memory, 11, 501–509.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and non-reinforcement. In A. H. Black & W. F. Prokasy (Eds.), Classical conditioning II: Current research and theory. New York: Appleton, pp. 64–99.
澤 幸祐(2007).古典的条件づけ理論におけるabsent cueの処理 専修人文論集,80, 245–263.
(Sawa, K. (2007). Information processing of “absent cue” in theories of classical conditioning. Studies in the Humanities:
Journal of the Senshu University Research Society, 80, 245–
263)
Schmajuk, N. (Ed.) Computational models of conditioning. Cambridge: Cambridge University Press.
Schmajuk, N. A., & Buhusi, C. V. (1997). Occasion setting, stimulus configuration, and the hippocampus: A neural net-work approach. Behavioral Neuroscience,111, 235–258. Schmajuk, N. A., & DiCarlo, J. J. (1991). A neural network
ap-proach to hippocampal function in classical conditioning.
Behavioral Neuroscience, 105, 82–110.
Schmajuk, N. A., Lam, Y., & Gray, J. A. (1996). Latent inhibi-tion: A neural network approach. Journal of Experimental
Psychology: Animal Behavior Processes, 22, 321–349.
Solomon, R. L., & Corbit, J. D. (1974). An opponent-process theory of motivation: I. Temporal dynamics of affect.
Psy-chological Review, 81, 119–145.
Spence, K. W. (1952). The nature of the response in discrimi-nation learning. Psychological Review, 59, 89–93.
Stout, S. C., & Miller, R. R. (2007). Sometimes-competing re-trieval (SOCR): A formalization of the comparator hypoth-esis. Psychological Review, 114, 759–783.
祐宗省三・春木 豊・小林重雄(編著)(1972).行動療 法入門 川島書店
(Sukemune, S., Haruki, Y., & Kobayashi, S.)
Tait, R. W., & Saladin, M. E. (1986). Concurrent development of excitatory and inhibitory associations during backward conditioning. Animal Learning & Behavior, 14, 133–137. Thorndike, E. L. (1911). Animal intelligence. New York:
Mac-millan
Tolman, E. C. (1932). Purposive behavior in animals and men. Berkeley, CA: University of California Press.
Urushihara, K., & Miller, R. R. (2010). Backward blocking in first-order conditioning. Journal of Experimental Psychology:
Animal Behavior Processes, 36, 281–295.
Van Hamme, L. J., & Wasserman, E. A. (1994). Cue competition in causality judgments: The role of nonpresentation of com-pound stimulus elements. Learning and Motivation, 25, 127– 151.
Wagner, A. R. (1971). Elementary associations. In H. H. Kend-ler & J. T. Spence (Eds.), Essays in neobehaviorism: A
memo-rial volume to Kenneth W. Spence. New York: Appleton, pp.
187–213.
Wagner, A. R. (1978). Expectancies and the priming of STM. In S. H. Hulse, H. Fowler & W. K. Honig (Eds.), Cognitive
processes in animal behavior. Hillsdale, NJ: Erlbaum, pp.
177–209.
Wagner, A. R. (1981). SOP: A model of automatic memory processing in animal behavior. In N. E. Spear & R. R. Miller (Eds.), Information processing in animals: Memory
mecha-nisms. Hillsdale, NJ: Erlbaum, pp. 5–47.
Wagner, A. R. (2008). Evolution of an elemental theory of Pav-lovian conditioning. Learning & Behavior, 36, 253–265. Wagner, A. R., & Brandon, S. E. (1989). Evolution of a structured
connectionist model of Pavlovian conditioning (AESOP). In
S. B. Klein and R. R. Mowrer (Eds.), Contemporary learning
theories: Pavlovian conditioning and the status of traditional learning theory. Hillsdale, NJ: Erlbaum, pp. 149–189.
Wagner, A. R., & Brandon, S. E. (2001). A componential theory of Pavlovian conditioning. In. R. R. Mowrer & S. B. Klein (Eds.), Handbook of contemporary learning theories. Mahwah, NJ: Erlbaum, pp. 23–64.
Wagner, A. R., Logan, F. A., Haberlandt, K., & Price, T. (1968). Stimulus selection in animal discrimination learning. Journal
of Experimental Psychology, 76, 171–180.
Wagner, A. R., & Rescorla, R. A. (1972). Inhibition in Pavlovian conditioning: Application of a theory. In R. A. Boakes & M. S. Halliday (Eds.), Inhibition and learning. London: Academic Press, pp. 301–336.
Wagner, A. R., & Vogel, E. H. (2010). Associative modulation of US processing: Implications for understanding of habituation. In N. Schmajuk (Ed.), Computational models of associative
learning. Cambridge: Cambridge University Press, pp.150–
185.
Walkenbach, J., & Haddad, N. (1980). The Rescorla–Wagner theory of conditioning: A review of the literature. The
Psy-chological Record, 30, 497–509.
Watson, J. B. (1913). Psychology as the behaviorist views it.
Psychological Review, 20, 158–177.
Whitlow, J. W., Jr., & Wagner, A. R. (1972). Negative patterning in classical conditioning: Summation of response tendencies to isolable and configural components. Psychonomic Science,
27, 299–301.
Williams, B. A. (1996). Evidence that blocking is due to asso-ciative deficit: Blocking history affects the degree of subse-quent associative competition. Psychonomic Bulletin &
Re-view, 3, 71–74.
Zimmer-Hart, C. L., & Rescorla, R. A. (1974). Extinction of Pavlovian conditioned inhibition. Journal of Comparative