229 生物工学 第99巻 第5号(2021) 〔生物工学会誌 第99巻 第5号 229–232.2021〕 DOI: 10.34565/seibutsukogaku.99.5_229 はじめに 代謝物総体(メタボローム)を対象とするオーム科学 であるメタボロミクスは,代謝物の定量情報を網羅的に 取得することで生命システムの新たな代謝制御の発見の みならず,ゲノム情報に隣接した高解像度の表現型解析 手段としても活用できる.現在,メタボローム解析は, 生物工学をはじめとした,さまざまな研究分野で活用さ れており,さらに,トランスクリプトームやプロテオー ムなどの他の階層オミクスデータとメタボロームデータ を統合して解析することで,細胞機能や生命システム全 体の真の理解につながると期待されている.本稿では, 現在のメタボロミクスの技術的課題を整理するととも に,筆者らが開発を進めている次世代のメタボローム計 測技術について概説する. メタボローム計測技術の現状と課題 メタボロミクス(metabolomics)は,「代謝物」を表す metabolite,ギリシャ語で「総体」を意味するome,「学問」 を意味する接尾辞icsを組み合わせた造語であり,分子 量2000以下の低分子化合物を対象とした網羅的解析を 行うことを意味する1,2).代謝物はアミノ酸や有機酸な どの親水性化合物から脂質などの疎水性化合物までの極 性範囲が広い.また,陽イオン性,両イオン性,陰イオ ン性,非荷電化合物があり,物理化学的特性を大きく左 右する電荷も多様である(図1).さらに,代謝物には同 重体や構造・幾何異性体が数多く存在するため,代謝物 を包括的かつ正確に測定するためには各種クロマトグラ フィーと質量分析(mass spectrometry, MS)を組み合わ せた分離分析法が一般的に使用されている.しかし,1 回の分析ですべての代謝物を測定することは実質不可能 に近いため,代謝物のカバレッジを向上させるためには, 複数の分析システムを組み合わせることが常套手段と なっている3). メタボロミクスはMSの技術革新の恩恵を受け発展して きた学問領域であるともいえる.たとえば,液体クロマトグ ラフィー高分解能タンデム質量分析(liquid chromatography high-resolution tandem mass spectrometry, LC/HRMS/ MS)が発展したことで,観測される分子量関連イオン (m/z)の測定精度は飛躍的に向上した.一般的に,LC/ HRMS/MSを用いたメタボローム分析は「ノンターゲッ トメタボローム分析」と呼ばれており,本手法を用いる ことで,動物細胞抽出液から3万種以上の分子量関連イ オンピークが確認できる4).しかし,現行の最先端の MS測定技術を用いたとしても,検出された分子量関連 イオンピークに対して同定される代謝物の割合は∼10 % に過ぎない5).メタボロミクス研究の多くは同定された 代謝物の定量情報を代謝経路マップ上に投影することで 生命現象を解釈することから,大半のマススペクトル情 報は活用されていないのが現状である.すなわち,高解 像度表現型解析手段であるメタボロミクスの特性を最大 限に活かすには,「未知代謝物の同定法(構造推定法)の 開発」が重要研究課題の一つとして位置づけられる.ま た,未知代謝物が発見できれば,その反応を触媒する酵 素の存在が示唆されるため「機能ゲノミクス研究の進展」 にも貢献できる. また,従来の生体関連分子のオミクス計測は,技術的 限界から細胞集団(105–107 cells)の分析が主であり「時 間および空間で平均した細胞の応答」を観測してきた. しかし,近年の次世代シーケンサーの技術革新に伴い, 1細胞レベルでの核酸情報の取得が可能となり,生命シ ステムには多種多様な個性を持つ細胞が不均一に存在す ることが明らかになってきている6).一方,メタボロー ム分析は,PCRのような観測対象物の増幅操作ができ ないため,細胞内に存在する膨大な種類の分子種を識別 しながら,そのまま検出する必要がある.2016年の 著者紹介 九州大学生体防御医学研究所附属トランスオミクス医学研究センター(准教授) E-mail: [email protected]
次世代のメタボローム計測
和泉 自泰
図1.研究室で所有する600種の代謝物標準品の分子量とオク タノール/水分配係数(logPow)のプロット230 特 集 生物工学 第99巻 第5号(2021) Nature誌に掲載された記事では,「MSの高感度化と細 胞サンプリング技術の発展により1細胞での代謝解析が 可能となりつつあるがこの研究分野はまだ発達初期段階 である」と結論づけられた7).すなわち,1細胞メタボ ロミクスを実現可能にするためには,分析系の高感度化, 1細胞の単離技術および1細胞試料調製法の開発が重要 な技術課題としてあげられる.また,近年,マルチオミ クスデータと代謝モデルを活用した代謝ネットワーク研 究が盛んに行われるようになってきている.当該研究分 野でいつも議論となるのが代謝物の細胞内局在の問題で ある8).たとえば,真核細胞のクエン酸回路中間体のク エン酸やリンゴ酸などは,細胞質とミトコンドリアのど ちらにも存在していると想定されている.今後,各代謝 物の細胞小器官の分布を定量的に観測する技術が構築で きれば,代謝モデルの精度向上および代謝ネットワーク の制御のより深い理解につながることが予想される. 筆者らは,以上のメタボロミクスにおける課題を解決 するために,次世代のメタボローム計測技術の開発を進 めている(図2).本稿では,①ワイドターゲットメタボ ローム分析法の開発,②未知代謝物の構造推定法の開発, ③1細胞メタボローム分析法の開発,を中心にこれらの 開発状況について簡単に紹介する. ワイドターゲットメタボローム分析法の開発 主幹代謝(解糖系,ペントースリン酸経路,クエン酸回 路,核酸代謝,アミノ酸代謝など)は,生命のエネルギー の生成,細胞の維持ならびに修復プロセスに関わり,また, がんなどの代謝を理解するうえでも,もっとも重要な経路 である.また,これらの代謝中間体の多くはイオン性高極 性物質である.これまで親水性代謝物の測定は,主にキャ ピラリー電気泳動質量分析(capillary electrophoresis mass spectrometry, CE/MS)9)やイオンペア逆相液体クロマト グラフィー質量分析(ion-pairing reversed-phase liquid chromatography mass spectrometry, IP-RP-LC/MS)10)な どによって測定されてきた.しかし,親水性代謝物は極 性,電荷特性,分子量といった物性の範囲が幅広いため, 包括的かつ実用的な測定には至っておらず,第一選択と なる分析法の開発は未だ発展途上の段階である.筆者ら は,昭和電工マテリアルズ・テクノサービス株式会社と の共同研究により,1∼3級アミンおよび4級アンモニ ウム基を修飾したメタクリレートポリマー粒子を充填し た新規カラムを設計し,親水性相互作用と陰イオン交換 作用を連続的に組み合わせた親水性相互作用/陰イオン 交換クロマトグラフィー高分解能タンデム質量分析法 (unified hydrophilic interaction/anion exchange liquid
chromatography tandem mass spectrometry, unified HILIC/ AEX/MS/MS)を開発した.本手法は,陽イオン性,両イ オン性,陰イオン性の極性代謝物を一斉にクロマト分離, MS検出可能な唯一の手法であり,従来法よりも再現性, 網羅性の点で優れていることを実験的に示した(投稿中). 一方,脂質の包括的観測を主眼としたリピドーム分析 については,超臨界流体クロマトグラフィータンデム質 量分析(supercritical fluid chromatography tandem mass spectrometer, SFC/MS/MS)とin silico多重反応モニタリ ング(multiple reaction monitoring, MRM)ライブラリー による独自のワイドターゲット定量リピドーム分析法の 開発を行ってきた11,12).筆者らは,質量分析計の最大の 弱点である定量性の問題を克服するために脂質クラスご とに内部標準物質を設定し,内部標準物質と個々の脂質 分子を共溶出させるクロマト分離条件を用いることで, 脂質クラス内の個々の脂質分子のマトリクス効果を一斉 に標準化することを考えた.具体的には,極性ヘッドグ ループによる各脂質クラスのクロマト分離を達成できる 順相カラムを用いたSFC分離モードを選定し,SFCに 接続する質量分析には,脂質クラスごとに同時溶出する 個々の脂質分子を網羅的かつ高感度分析が可能な三連 四重極質量分析計(triple-quadrupole mass spectrometer, QqQMS)を採用した.各脂質クラス内の構造異性体(PC 16:0–20:4とPC 18:2–18:2など)の識別には,脂肪酸側鎖由 来のMRMトランジションを用いた.一方で,QqQMSに よる測定法はターゲット分析であるため,事前に測定試料 に含まれる脂質分子の情報が必要となる.生体内の脂質を 構成する脂肪酸の種類は限られており,MS/MSでのフラ グメンテーションは脂質クラスごとに規則性を有している ことから,仮想のMRMトランジションを作製した.そし て,各サンプルから等量ずつ混合したreference sampleの スクリーニング分析を実施し,その結果から測定試料中に 存在する脂質分子を選定することで,最終のSFC/MS/MS メソッドを構築した.以上の方法論に基づいてSFC/MS/ MS分析条件を検討したところ,エチレン架橋型ハイブ 図2.次世代メタボロミクスに向けた技術的課題
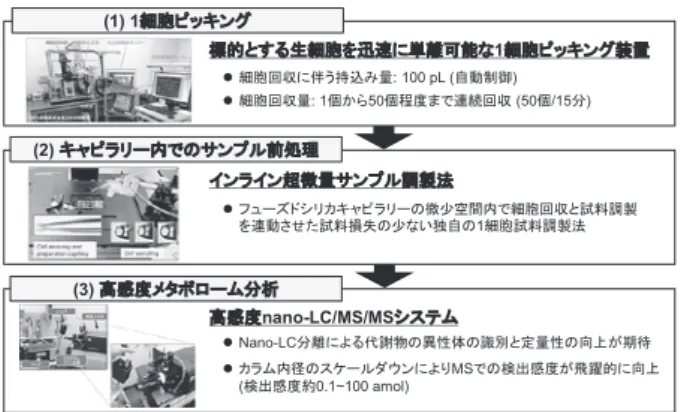
231 次世代のバイオ計測はこれだ!(前編)∼次世代,未来のバイオ計測技術∼ 生物工学 第99巻 第5号(2021) リッドシリカ粒子にジエチルアミン(diethylamine, DEA) を修飾したカラムを用いることで,中性脂質から極性脂 質までの幅広い脂質クラス22種をわずか20分で分離分 析することに成功した.また,各脂質クラスの代表的な 脂質分子を対象として添加回収試験を行ったところ,回 収率は64.9∼103.5 %であり,当該手法が定量性の高 い分析法であることが示された. 現在,当研究室での汎用メタボローム解析には,HILIC/ AEX/MS/MSおよびSFC/MS/MSを中心とした数種の 分析系を組み合わせたワイドターゲット分析法を実施し ており,微生物や動物細胞のバルク試料(>106 cells)か ら約500–800種の代謝物の定量情報の取得が可能となっ ている13). 未知代謝物の構造推定法の開発 LC/HRMS/MSによる生体内代謝物の同定は,標準品 を用いて,(1)プリカーサーイオンの一致度(<5 ppm), (2)プロダクトイオン(MS/MS)の類似性,(3)LCの 保持時間の一致度を基に決定する.未知の分子量関連イ オンピークに対する構造アノテーションとは,標準品が 入手できない化合物を対象としたものを指す.代謝物同 定については,メタボロミクス研究が開始された2000 年頃から大きな課題と認識されていた.この問題を解消 していくために,日本国内ではMassBank(http://www. massbank.jp/)14)と呼ばれる代謝物標準品のMSおよび MS/MSスペクトルデータベースが構築され,米国では ヒトに特化した代謝物データベースHuman Metabolome Database(HMDB, http://www.hmdb.ca/)15)の整備が行 われてきた.先行研究による未知の分子量関連イオンの 構造推定は,MassBankやHMDBなどの化合物データ ベースを活用し,上記の(1)および(2)によって実施 されてきた.しかし,これらの方法は,あくまで既知化 合物に対するアノテーション法であり,未知の低分子化 合物の構造に対してはMS/MSによる部分構造の類似性 の情報のみしか得ることができない.そのため,未知の 分子量関連イオンピークの網羅的な構造推定を実施する ためには,合理的な代謝物構造情報が格納されたライブ ラリーを開発する必要があった.そこで,筆者らは膨大 な数の内因性未知代謝物の構造推定を進める前に,外因 性化学物質をモデルケースとして,未知の分子量関連イ オンピークの構造推定法の開発に着手した. 薬物のような外因性化学物質は動植物の生体内に取り 込まれた後,各種酵素の作用によって代謝され,さまざ まな化学物質へと変換されることが知られている.筆者 ら は,13Cお よ び2H安 定 同 位 体 標 識 体 の 利 用,LC/ HRMS/MS,データマイニング技術,in silico代謝物予測, 代謝物プロファイルの経時的変化を解析することで,外 因性化学物質に由来する代謝物を高精度に同定するため の新しい方法論を提唱した.具体的には,古くから使用 されてきた除草剤,2,4-dichlorophenoxyacetic acid(2,4-D) をシロイヌナズナT87培養細胞に暴露した際に代謝され る化合物の網羅的探索を行った.その結果,in silico代謝 物予測およびHRMS/MSスペクトル情報をもとに,こ れまでに報告のある10種の代謝物に加えて16種の新規 2,4-D代謝物の同定に成功した16).同様の手法を用いて, コーヒー中のアクリルアミド(acrylamide, AA)代謝物 についても同定を試みた.コーヒー中に含まれる5000 種以上の分子量関連イオンピークの中から,AAと 13C 3-AAの精密質量差(Δm/z = 3.0102)および保持時間 の一致度をもとにAA付加体候補化合物の絞込みを行 い,最終的に2種の新規AA付加体を形成していること を明らかにした17). 以上の結果を踏まえ,本手法は一部の方法論を改変す ることで内因性の未知代謝物の同定にも適応できる可能 性が高いと判断した.未知代謝物の同定を行うための重 要な戦略の一つとして,未同定の代謝物の多くは,すで に生化学的手技によって解明された主幹となる代謝反応 の組合せの産物が含まれているという仮説を立てた.現 在,生物種を問わない既知の代謝物と酵素反応を多段階 に組み合わせた100万種以上のin silicoエピメタボライ トライブラリーを構築し,大腸菌や酵母,ヒト培養細胞 を対象とした未知の代謝物の包括的構造推定を実施して いる. 1細胞メタボローム分析法の開発 これまで筆者らは,一般的な動物1細胞(直径20 μm, 4 pL)からのメタボローム解析を達成するために,(1) 顕微鏡下で標的とする生細胞を迅速に単離・回収する技 術(ヨダカ技研株式会社の平藤衛氏,産業技術総合研究 所の山村昌平研究グループ長との共同開発),(2)サン プルロスを低減させた微小空間内での前処理および微量 試料導入技術(新潟大学大学院医歯学総合研究科の松本 雅記教授との共同開発),(3)ナノ液体クロマトグラ フィータンデム質量分析(nano-LC/MS/MS)による高 感度分析技術を開発してきた(図3).動物1細胞のメタ ボローム解析は,生体内に比較的豊富に存在するアミノ 酸,核酸塩基,ヌクレオシド,ヌクレオチドを測定対象 とした.開発した1細胞メタボローム分析法を用いて, ヒト頸がん由来細胞(HeLa細胞)の代謝レベルでの不 均一性を評価した.単一HeLa細胞のメタボローム分析
232 特 集 生物工学 第99巻 第5号(2021) を実施したところ,18種の親水性代謝物の検出に成功 した.さらに,培養したHeLa細胞集団から取り出した 22個の単一HeLa細胞を分析して得られた代謝物ピーク エリアを用いて階層的クラスター分析(hierarchical cluster analysis, HCA)を行った結果,細胞の間に,代 謝レベルで一定の類似度を示すサブクラスが存在するこ とを示した.今回の培養細胞を用いた1細胞メタボロー ム結果により観測されたサブクラスは,たとえば細胞周 期の違いあるいはその他の代謝機能の違いを捉えている 可能性が示唆された18). 本研究で使用した内径100 μmのnano-LCカラムによ る分析システムの感度では,典型的な動物1細胞から検 出可能な代謝物の数はまだまだ少ない.一方,筆者らは ハイブリッド型多孔質シリカ層を有する内径5 μmの オープンチューブラー型カラムの調製19)に成功してい る.今後,本カラムを用いた超高感度の1細胞メタボロー ム解析により,動物1細胞から200種以上の代謝物の検 出に挑戦する予定である. おわりに これまでのメタボロミクス研究は,研究者の興味の対 象となるサンプル群を用いて,既知の代謝経路上の代謝 物中間体の蓄積量,あるいは13Cグルコースなどを使用し た標識実験による代謝フラックスを実測し,表現型の違 い,バイオマーカーや代謝律速点の探索,あるいは代謝 制御機構を調べることが主な目的であった.一方,既知 代謝物の中のSAM(S-adenosylmethionine)やAcetyl-CoA は共有結合の転写調節,すなわちクロマチン修飾酵素の 補因子として働き,ヒストンのメチル化やアセチル化に よるエピジェネティックな制御にも関わっている20).ま た,近年,腫瘍が分泌する乳酸はM1マクロファージの ヒストン修飾(ラクチル化)を引き起こすことでM2マ クロファージの発現様式に誘導させ,腫瘍自身の増殖促 進に有利な環境を整えていることが示唆されている21). このように,いくつかの代謝物は,細胞のさまざまな生理 機能に大きな影響を与える制御因子としても働いている. 筆者らが開発を進めている次世代のメタボローム分析法 は,活性調節因子として働く新規代謝物の同定など,今 後のメタボロミクス研究の発展に大きく貢献できる実用 的な計測技術になりえると信じている. 謝 辞 本研究の一部は科学技術振興機構(JST)CREST「統合1細 胞解析のための革新的技術基盤」およびJST未来社会創造事業 「探索加速型(共通基盤領域)」の支援を受け実施した. 文 献
1) Fiehn, O.: Plant Mol. Biol., 48, 155 (2002).
2) Fukusaki, E. and Kobayashi, A.: J. Biosci. Bioeng., 100, 347 (2005).
3) Izumi, Y. et al.: Metabolites, 9, 257 (2019). 4) Ivanisevic, J. et al.: Anal. Chem., 85, 6876 (2013). 5) Creek, D. J. et al.: Anal. Chem., 84, 8442 (2012). 6) Saliba, A. E. et al.: Nucleic Acids Res., 42, 8845 (2014). 7) Fessenden, M.: Nature, 540, 153 (2016).
8) Volkova, S. et al.: Metabolites, 10, 303 (2020). 9) Harada, K. et al.: J. Biosci. Bioeng., 105, 249 (2008). 10) Kato, H. et al.: J. Biosci. Bioeng., 113, 665 (2012). 11) Takeda, H. et al.: J. Lipid Res., 59, 1283 (2018). 12) 和泉自泰,馬場健史:生物工学,98, 490 (2020). 13) Fushimi, T. et al.: J. Agric. Food Chem., 68, 11997 (2020). 14) Horai, H. et al.: J. Mass Spectrom., 45, 703 (2010). 15) Wishart, D. S. et al.: Nucleic Acids Res., 35, D521 (2007). 16) Takahashi, M. et al.: Anal. Chem., 90, 9068 (2018). 17) Yoshioka, T. et al.: J. Agric. Food Chem., 68, 3859 (2020). 18) Nakatani, K. et al.: Mass Spectrom., 9, A0080 (2020). 19) Hara, T. et al.: J. Chromatogr. A, 1580, 63 (2018). 20) Rinschen, M. M. et al.: Nat. Rev. Mol. Cell Biol., 20, 353
(2019).
21) Zhang, D. et al.: Nature, 574, 575 (2019). 図3.開発した1細胞メタボローム解析システム