4-Phenylbutyrate suppresses the unfolded protein response without restoring protein folding in Saccharomyces cerevisiae
30
0
0
全文
(2) 22. ABSTRACT. 23. Accumulation of unfolded secretory proteins in the endoplasmic reticulum (ER),. 24. namely ER stress, is hazardous to eukaryotic cells and promotes the unfolded protein. 25. response (UPR). Ire1 is an ER-located transmembrane protein that senses ER stress and. 26. triggers the UPR. According to previous in vitro experiments, 4-phenylbutyrate (4-PBA). 27. works as a chemical molecular chaperone. Since 4-PBA attenuates the UPR in. 28. mammalian tissue cultures, this chemical may have clinical potential for restoring ER-. 29. stressing conditions. In the present study, we investigated 4-PBA’s mode of action using. 30. the yeast Saccharomyces cerevisiae as a model organism. Although 4-PBA blocked a. 31. dithiothreitol (DTT)-induced UPR, it did not appear to restore impairment of ER protein. 32. folding that was caused by DTT. Moreover, even under non-stress conditions, 4-PBA. 33. attenuated UPR that was induced by an Ire1 mutant that exhibits a substantial activity. 34. without sensing ER accumulation of unfolded proteins. We also found that 4-PBA. 35. drastically promotes the degradation of Ire1. These observations indicate that at least in. 36. the case of yeast cells, 4-PBA suppresses the UPR not through restoration of the ER. 37. function to correctly fold proteins. Instead, the accelerated degradation of Ire1 possibly. 38. explains the reason why the UPR is attenuated by 4-PBA.. 39 40. INTRODUCTION. 41. The endoplasmic reticulum (ER) is the cellular compartment where secretory and. 42. transmembrane proteins are folded. Misfolding of proteins can be hazardous to cells. 43. through various reasons. Aberrant ER client proteins, such as mutants of cystic fibrosis. 44. transmembrane conductance regulator (CFTR), are unable to reach their final. 45. destination and exert their proper function because they are accumulated in the ER and. 46. degraded through the ER-associated protein degradation (Pranke and Sermet-Gaudelus,. 47. 2014). Moreover, misfolded proteins and their aggregates per se are toxic to cells (Soto,. 48. 2003). The ER stress response or the unfolded protein response (UPR) is triggered by. 49. ER accumulation of misfolded proteins, so-called ER stress, commonly in eukaryotic 2.
(3) 50. organisms (Mori, 2009; Walter and Ron, 2011). Although the ER stress response. 51. generally protects cells against protein aggregation, in the case of metazoan cells, it can. 52. also lead to apoptosis (Shore et al., 2011). Because these issues are tightly related to. 53. various human diseases, which include diabetic mellitus and some neurodegenerative. 54. diseases, chemical compounds that suppress ER protein aggregation and the ER stress. 55. response may be valuable sources for drug development (Oakes and Papa, 2015).. 56. It is well known that 4-phenylbutyrate (4-PBA) is such a compound (Kolb et al.,. 57. 2015; Ranga Rao et al., 2018). While being soluble in aqueous environments, 4-PBA. 58. has a hydrophobic moiety, implying that it can inhibit aggregation of proteins as a. 59. chemical chaperone. Intriguingly, 4-PBA suppresses the UPR in both mammalian and. 60. yeast cells (Kubota et al., 2006; Ozcan et al., 2006; Pineau et al., 2009; Le et al., 2016).. 61. Ire1 is an ER-located type-I transmembrene protein commonly found in all. 62. eukaryotic species. In the budding yeast Saccharomyces cerevisiae, Ire1 is believed to. 63. serve as a sole ER-stress sensor that initiates the UPR, which is a dramatic. 64. transcriptome change that occurs in response to ER stress. Ire1 has an endoribonuclease. 65. domain in its cytosolic moiey (Sidrauski and Walter, 1997). When yeast cells are. 66. exposed to ER stress, Ire1 promotes the splicing of the HAC1 mRNA (Cox and Walter,. 67. 1996), which is then translated into a transcription factor protein that changes the yeast. 68. transcriptome to restore ER functions (Travers et al., 2000; Kimata et al., 2006).. 69. As reviewed in our previous publication (Kimata and Kohno, 2011), the luminal. 70. domain of Ire1 plays critical roles in regulating the activity of Ire1 in response to ER. 71. stress. The ER-located molecular chaperone BiP is associated with the luminal domain. 72. of Ire1 to inhibit self-association and activation of Ire1 (Bertolotti et al., 2000; Okamura. 73. et al., 2000; Kimata et al., 2003). ER-accumulated misfolded proteins cause dissociation. 74. of BiP from Ire1 and enable the self-association of Ire1, which alone is not sufficient for. 75. its activation (Kimata et al., 2004; Oikawa et al., 2007). We and others have also. 76. reported that unfolded proteins are directly captured by the luminal domain of Ire1 for. 77. complete induction of the UPR (Kimata et al., 2007; Promlek et al., 2011; Gardner and 3.
(4) 78 79. Walter, 2011). These insights explain the molecular mechanism by which ER-accumulated. 80. misfolded proteins activate Ire1 and trigger the UPR. It is thus widely believed that 4-. 81. PBA may attenuate the UPR by restoring the protein-folding status in the ER. Indeed, 4-. 82. PBA is likely to assist the proper folding of ER client proteins such as a CFTR mutant. 83. (Rubenstein et al., 1997; Lim et al., 2004; Kubota et al., 2006).. 84. In this study, using yeast as a model organism, we addressed the molecular. 85. process through which 4-PBA suppresses the UPR. Our findings indicate that contrary. 86. to the theory described above, 4-PBA does not seem to assist protein folding in the ER.. 87. Here we propose that 4-PBA induces the degradation of Ire1, which is likely to result in. 88. the attenuation of the UPR.. 89 90. MATERIAL AND METHODS. 91. Yeast strains. 92. S. cerevisiae ire1 (ire1::TRP1) strain KMY1015 was derived from a standard. 93. laboratory strain KMY1005 (MAT leu2-3,112 ura3-52 his3200 trp1901 lys2-801). 94. and transformed with the UPR lacZ-reporter plasmid pCZY1 (Mori et al., 1992). The. 95. resulting strain was further transformed with a single copy plasmid pRS313-IRE1. 96. (Kimata et al., 2004), which has the endogenous IRE1 gene (the coding sequence plus. 97. the 5’- and 3’-untranslated regions), and was used as a wild-type IRE1 strain TKY001.. 98. Strain TKY002 is a derivative of TKY001 that has the bZIP-mutant version of the IRE1. 99. gene (Promlek et al., 2011) instead of the wild-type IRE1 gene. Strain TKY003 was. 100. obtained by transforming KMY1005 with the eroGFP expression plasmid pPM28. 101. (Merksamer et al., 2008).. 102. Insertion of an HA epitope sequence into the IRE1 gene was previously described. 103. (Kimata et al., 2003). The resulting gene was inserted into the single copy plasmid. 104. vector (to generate pRS315-IRE1-HA; Kimata et al., 2004) or into the multi-copy. 105. plasmid vector (to generate pRS423-IRE1-HA; Kimata et al., 2003) and was used for 4.
(5) 106. expression of the C-terminally HA-tagged version of Ire1 (Ire1-HA). We transformed. 107. KMY1015 with pCZY1 and pRS315-IRE1-HA to obtain strain TKY004 and with. 108. pCZY1 and pRS423-IRE1-HA to obtain strain TKY005. TKY006 is a derivative of. 109. TKY004 that has the bZIP mutant version of the IRE1-HA gene (Promlek et al., 2011).. 110. Strain BY4742hrd1 (MAT his31 leu20 lys20 ura30 hrd1::kanMX4) was. 111. purchased from EUROSCARF (http://www.euroscarf.de). The DOA10 gene on. 112. BY4742hrd1 was disrupted by the HIS5MX marker, and the resulting hrd1 doa10. 113. strain was further transformed with pRS315-IRE1-HA and named TKY007.. 114. For all experiments shown in this study, yeast cells were cultured at 30°C in. 115. standard synthetic dextrose medium containing 2% glucose, 0.66% yeast nitrogen base. 116. without amino acids (MP Biomedicals) and appropriate auxotrophic requirments.. 117 118. Chemicals. 119. Dithiothreitol (DTT; catalog code: D0632), tunicamycin (catalog code: T7765), and 4-. 120. PBA (catalog code: SML0309) were purchased from Sigma-Aldrich.. 121 122. Protein analyses. 123. After being harvested by centrifugation at 1,600 ×g for 1 min, 5.0 OD600 cells were. 124. disrupted by agitation with glass beads (425-600 µm) in 100 µl of the lysis buffer. 125. containing 50 mM Tris-Cl (pH 7.9), 5 mM ethylenediaminetetraacetic acid, 1% Triton. 126. X-100 and protease inhibitors (2 mM phenylmethylsulfonyl fluoride, 100 µg/ml. 127. leupeptin, 100 µg/ml aprotinin, 20 µg/ml pepstatin A and Calbiochem Protease Inhibitor. 128. cocktail Set III (100x dilution)). The cell lysates were then clarified by centrifugation at. 129. 8,000 ×g for 10 min, fractionated using standard SDS-DTT polyacrylamide gel. 130. electrophoresis (PAGE), and transferred onto polyvinylidene difluoride-based Western. 131. blotting membrane (Hybond-P; GE Healthcare). The semi-dry electrophoretic transfer. 132. and subsequent treatment of the blot membranes were performed as indicated by the. 133. manufacturer’s instruction. As primary antibodies, we used 12CA5 mouse monoclonal 5.
(6) 134. anti-HA antibody (Roche), 22C5D8 mouse monoclonal anti-Pgk1 antibody (Abcam). 135. and rabbit polyclonal anti-yeast BiP antibody (Kimata et al., 2003). After treatment of. 136. the blot membranes with the horseradish peroxidase-conjugated secondary antibodies. 137. and the enhanced chemiluminescence (ECL) reagent (ECL prime, GE healthcare), the. 138. ECL signal was quantitatively detected using the luminescence imager LAS4000 (Fuji. 139. Film).. 140. For the BiP aggregation assay, cells were disrupted as described in the previous. 141. paragraph and subjected to sequential centrifugations at 720 ×g for 30 sec and 8,000 ×g. 142. for 15 min. The pellet fractions obtained by the second centrifugation were then washed. 143. twice using the lysis buffer and analyzed as the “pellet” samples with the anti-BiP. 144. Western blotting.. 145. For the pulse-chase experiment, cells were labeled by culturing with 35S-labeled. 146. methionine and cysteine (400 kBq/ml; EXPRE35S35S Protein Labeling Mix,. 147. PerkinElmer) for 2 hr. Chase incubation was initiated by adding non-labeled methionine. 148. at a final concentration of 0.004% and cysteine at a final concentration of 0.003%. Cell. 149. lysates were then obtained as mentioned above and subjected to anti-HA. 150. immunoprecipitation, as previously described (Kimata et al., 2003). Standard SDS-DTT. 151. PAGE was performed to fractionate the immunoprecipitation products, and the. 152. radioactive signal was detected on an autoradiograph. For autoradiography, a storage. 153. phosphor screen BAS-MS2040 (Fuji Film) and its reader BAS2500 (Fuji Film) were. 154. employed.. 155 156. RNA analyses. 157. The hot phenol method was employed for extracting total RNA from yeast cells (Kimata. 158. et al., 2003). To monitor the HAC1 mRNA splicing, we subjected the total RNA sample. 159. to reverse transcriptase (RT)-PCR analysis using a 17-mer polyA primer for the RT. 160. reaction and the HAC1-specific primer sets for PCR (Promlek et al., 2011). The. 161. resulting RT-PCR products were then run on a 2% agarose gel, stained with ethidium 6.
(7) 162 163. bromide, and observed with the fluorescence imager GelDoc XR+ system (BioRad). For quantification of the IRE1-gene transcript, total RNA samples were subjected. 164. to RT reaction with the 17-mer polyA primer. We then analyzed the RT reaction. 165. products by qPCR using SYBR Premix Ex Taq™ II (Tli RNaseH Plus; Takara) in the. 166. real-time PCR machine LightCycler 96 (Roche). The primer sets used are as. 167. follows: 5’-AGT CAG AAT TTT CCA TCT TTA GTC G-3’/5’-CGG TCA CTG GAA. 168. GCG TAT CT-3’ for the amplification of the IRE1-gene product and 5’-ATA TTC CAG. 169. GAT CAG GTC TTC CGT AGC-3’/5’-GTA GTC TTC TCA TTC TGT TGA TGT TGT. 170. TGT TG-3’ for the amplification of the TAF10-gene product, which served as an. 171. internal control.. 172 173. Microscopic observation. 174. We observed yeast cells carrying the eroGFP expression plasmid using the Keyence BZ-. 175. 9000E microscope and the objective lens CFI Plan Apo 100xH (Nikon). Florescence. 176. images were observed using the filter set OP-79301 (excitation wavelength, 472.5/30;. 177. dichroic mirror wavelength, 495; emission, 520/35) for excitation with blue light (1.0-. 178. sec exposure) and a custom-made filter set (excitation wavelength, 395/25; dichroic. 179. mirror wavelength, 495; emission, 510/20) for excitation with UV/violet light (1.3-sec. 180. exposure).. 181 182. Data quantification. 183. In this study, we analyzed gel images using the image processing software Image J. 184. (https://imagej.nih.gov/ij/) to obtain quantitative data, which are presented here as. 185. averages and standard deviations of the values from multiple cultures. The following. 186. formula was used to obtain the “HAC1 mRNA-splicing efficiency” from fluorescent. 187. images of the HAC1 RT-PCR products: [(band intensity of the spliced form)/{(band. 188. intensity of the spliced form) + (band intensity of the unspliced form)}].. 189 7.
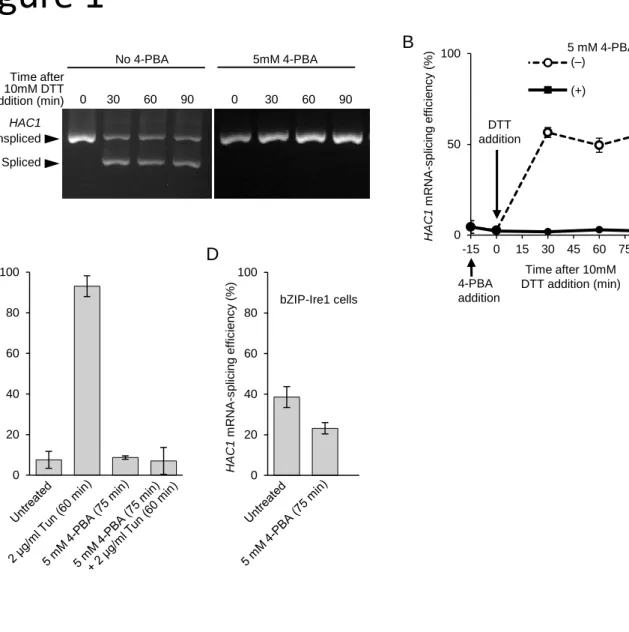
(8) 190. RESULTS. 191. 4-PBA suppresses yeast UPR.. 192. In this study, we monitored the UPR in yeast cells through checking the HAC1-mRNA. 193. splicing, which was measured using the RT-PCR technique that amplify the HAC1. 194. mRNA species. In the experiment shown in Figs 1A and B, yeast cells were cultured in. 195. the presence of 10 mM DTT, a disulfide-reducing reagent that damages ER protein. 196. folding and causes a potent UPR. As we reported previously (Le et al., 2016), 5 mM 4-. 197. PBA almost completely abolished the UPR when added into the culture 15 min before. 198. DTT imposition. 4-PBA also attenuates the UPR induced by an N-glycosylation. 199. inhibitor tumicamycin (Fig. 1C), as initially indicated by Pineau et al. (2009).. 200. In a mutant form of yeast Ire1, which is named as bZIP-Ire1, the full-length. 201. luminal domain was replaced with a basic leucine zipper (bZIP) dimer-forming peptide. 202. that was derived from the yeast bZIP-family transcription factor protein Gcn4 (Liu et. 203. al., 2000; Promlek et al., 2011). This mutant is located on the ER membrane and. 204. exhibits a modest UPR-inducing activity even in the absence of external ER-stressing. 205. stimuli. As shown in Fig. 1D, 4-PBA attenuated, although not drastically, the HAC1-. 206. mRNA splicing induced by bZIP-Ire1 in non-stressed yeast cells.. 207 208. 4-PBA cannot restore ER damage induced by DTT.. 209. Because 4-PBA attenuated activity of bZIP-Ire1, which lacks the misfolded protein-. 210. sensing luminal domain, we questioned the idea that 4-PBA suppresses the UPR by. 211. restoring protein-folding capacity of the ER. We thus investigated protein-folding status. 212. in the ER of yeast cells in the presence and absence of 4-PBA.. 213. In the experiment shown in Fig. 2, we cultured cells with or without DTT and/or. 214. 4-PBA, lysed them in the presence of the mild detergent Triton X-100. The total lysate. 215. samples were then fractionated by centrifugation. Fig. 2A shows that in the absence of. 216. 4-PBA, DTT treatment modestly increased the amount of BiP in the total lysate. 217. samples, indicating an induction of the UPR (compare lane 2 with 1). Importantly, 8.
(9) 218. similarly to our previous observation (Promlek et al., 2011), the cellular DTT treatment. 219. caused a more drastic increment of the BiP level in the pellet samples (Fig. 2B, compare. 220. lane 2 with 1). This is presumably because in yeast ER, DTT leads to the formation of. 221. protein aggregates that incorporate BiP. Since 4-PBA attenuates DTT-induced UPR. 222. (Figs 1A and B), it suppressed the increase in BiP levels by cellular DTT treatment in. 223. the total lysate samples (Fig. 2A, compare lane 4 with 2). However, 4-PBA did not. 224. appear to affect the BiP level in the pellet samples (Fig. 2B, compare lane 3 with 1 and. 225. lane 4 with 2). This observation indicates that 4-PBA does not diminish protein. 226. aggregation in the ER of non-stressed cells and of DTT-treated cells. We then performed. 227. a similar experiment using an ire1 strain, and found that not only DTT but also 4-PBA. 228. enhances BiP sedimentation in this strain (Fig. S1).. 229. Because a mutant form of GFP, which is named as eroGFP, has an N-terminal. 230. ER-translocation signal and a C-terminal ER-retention signal (His-Asp-Glu-Leu), it is. 231. located in the ER (Merksamer et al., 2008). Moreover, eroGFP changes its excitation. 232. spectrum upon formation of a single intramolecular disulfide bond (Merksamer et al.,. 233. 2008). Using a fluorescent microscope having two different excitation-light filters, we. 234. observed yeast cells that produce eroGFP (Fig. 3). As expected, treatment of cells with. 235. DTT changed the ratio of fluorescent intensity of eroGFP subjected to two different. 236. excitation wavelengths (compare column 2 with 1), indicating an impaired disulfide-. 237. bond formation of eroGFP. Here we showed that this ratio was not affected by 4-PBA. 238. (compare column 3 with 1 and column 4 with 2). This observation indicates that 4-PBA. 239. does not restore the ability of the ER to form protein disulfide bonds, which is the so-. 240. called oxidative protein folding and is an important part in the maturation of ER client. 241. proteins.. 242 243. 4-PBA promotes degradation of Ire1 in yeast cells.. 244. To explore the mechanism by which 4-PBA attenuates the UPR, we next measured the. 245. cellular amount of Ire1, which is a key factor for the HAC1-mRNA splicing and the 9.
(10) 246. UPR induction. In the experiment shown in Fig. 4A, yeast cells that produce C-. 247. terminally HA-tagged Ire1 (Ire1-HA; Kimata et al., 2003) under the control of the IRE1. 248. authentic promoter were assessed for their cellular Ire1-HA level through anti-HA. 249. Western blot analysis of their cell lysates. We then found that cellular treatment with. 250. DTT slightly induced Ire1-HA (Fig. 4A; compare lane 2 with 1). Moreover, and. 251. importantly, 4-PBA treatment severely decreased the cellular Ire1-HA level both in the. 252. presence and absence of DTT (Fig. 4A; compare lane 3 with 1 and lane 4 with 2). Fig.. 253. 4A also shows that the cellular amount of a cytosolic Pgk1 was almost unaffected by. 254. DTT or 4-PBA. Using the qRT-PCR technique (Fig. 4B), we observed that 4-PBA did. 255. not appear to considerably decrease the mRNA expression level of the IRE1 gene. As. 256. shown in Fig. 4C, 4-PBA also decreased the cellular amount of the HA-tagged version. 257. of bZIP-Ire1, albeit modestly.. 258. We next monitored the stability of Ire1-HA in yeast cells through the 35S. 259. methionine/cysteine pulse labeling and chase experiment. In the experiments shown in. 260. Fig.5, yeast cells carrying a multi-copy (2µ) plasmid vector fused with the HA-tagged. 261. version of the IRE1 gene (IRE1-HA) were labeled with 35S methionine/cysteine for 2hr.. 262. We then added or did not added 4-PBA into the labeled cultures simultaneously with the. 263. addition of non-labeled methionine and cysteine, which started the chase period (Fig.. 264. 5A). According to the data shown in Fig. 5B, Ire1-HA signal was almost undetectable. 265. after a 75-min chase in the presence of 4-PBA (lane 3), whereas Ire1-HA appeared to be. 266. quite stable in untreated cells (compare lane 2 with 1). This finding indicates that 4-PBA. 267. dramatically accelerates the degradation of Ire1-HA.. 268. ER-resident transmembrane proteins are often digested through their. 269. ubiquitination and proteasomal degradation. This process is named the ER-associated. 270. degradation (ERAD), for which Hrd1 and Doa10 serve as the ubiquitin ligases. As. 271. shown in Fig. S2, 4-PBA drastically reduced the amount of Ire1-HA even in. 272. hrd1doa10 cells. This observation indicates that the ERAD is not involved in the. 273. degradation of Ire1 promoted by 4-PBA. 10.
(11) 274. If 4-PBA attenuates the UPR via reduction of the cellular level of Ire1,. 275. overproduction of Ire1 may, at least partly, suppresses the effect of 4-PBA to repress the. 276. UPR. In order to validate this idea, we again employed yeast cells carrying the 2µ-. 277. plasmid-borne IRE1-HA gene. The cellular level of overproduced Ire1-HA was. 278. decreased by cellular treatment with 4-PBA (Fig. 6A). As shown in Fig. 6B,. 279. overproduction of Ire1-HA per se slightly caused the HAC1 mRNA splicing (column 1),. 280. which was enhanced by cellular treatment with DTT (column 2), and 4-PBA lowered,. 281. but did not completely abolished, the level of HAC1 mRNA splicing both in the. 282. presence or absence of DTT (compare column 3 to 1 and column 4 to 2). Importantly. 283. and as expected, the UPR was induced even in the presence of 4-PBA, although weakly. 284. (Fig. 6B, compare column 4 to 3).. 285 286. DISCUSSION. 287. According to in vitro experiments presented by Kubota et al. (2006), 4-PBA has an. 288. ability to function as a chemical chaperone to prevent the aggregation of denatured. 289. proteins. As mentioned in the Introduction section, this is presumably because of the. 290. chemical property of 4-PBA, a water-soluble molecule albeit with a hydrophobic. 291. moiety. Moreover, 4-PBA suppresses cellular responses against the accumulation of. 292. misfolded proteins in the ER, such as the UPR, in both yeast and mammalian cells. 293. (Kubota et al., 2006; Ozcan et al., 2006; Pineau et al., 2009; Le et al., 2016). These. 294. observations led to an idea that under some stressing conditions, 4-PBA may work as a. 295. chemical chaperone in vivo with an ability to totally restore the protein-folding status in. 296. the ER. 4-PBA is thus expected to have a therapeutic potential (Kolb et al., 2015; Ranga. 297. Rao et al., 2018) because misfolding and aggregation of ER client proteins are linked to. 298. numerous human diseases (Wang and Kaufman, 2016). This theory may lead to an. 299. expansion of the clinical usage of 4-PBA, which is already used to alleviate the urea. 300. cycle disorder.. 11.
(12) 301. However, here we show that the suppression of the UPR by 4-PBA is not due to. 302. restoration of the ER protein-folding status at least in the case of yeast cells. While cells. 303. carrying bZIP-Ire1 evokes the UPR independently of the accumulation of misfolded. 304. proteins in the ER (Promlek et al., 2011), 4-PBA suppresses the UPR also in bZIP-Ire1. 305. cells (Fig. 1D). Moreover, while 4-PBA almost completely abolishes the HAC1-mRNA. 306. splicing induced by DTT in IRE1 cells (Figs 1A and B), it does not restore the DTT. 307. damage of ER protein folding (Figs 2 and 3). Conversely, 4-PBA seemed to worsen. 308. protein folding status in the ER of ire1 cells (Fig. S1). According to Bicknell et al.. 309. (2007), Ire1 is not absolutely required but assists maintenance of integrity of ER. 310. functions even in non-stressed cells. It is thus likely that, even under normal culturing. 311. conditions, ER conditions in IRE1 cells and ire1 cells are somehow different, which. 312. explains the reason why 4-PBA facilitates BiP sedimentation not in IRE1 cells but in. 313. ire1 cells.. 314. According to our observations shown in Figs 4 and 5, 4-PBA induces a rapid. 315. degradation of Ire1, which possibly leads to the attenuation of the UPR shown in this. 316. study. Consistently to this idea, 4-PBA did not completely abolish the UPR in cells. 317. overproducing Ire1-HA (Fig. 6). By considering its fast degradation shown in Fig. 5, we. 318. deduce that the Ire1-HA molecules are mostly new-borne in these cells treated with 4-. 319. PBA. It should be noted that, in yeast cells treated with DTT, the Ire1 molecules are. 320. carried along actin cables to form the Ire1 clusters, which shows a potent HAC1-mRNA. 321. splicing activity (Ishiwata-Kimata et al., 2013). This insight may explain the reasons. 322. why the new-borne Ire1-HA poorly triggers the UPR strongly and why, in the. 323. experiment shown in Fig. 6, the UPR is only modest in the presence of both DTT and 4-. 324. PBA. The molecular mechanism by which 4-PBA accelerates the degradation of Ire1. 325. remains unknown, since the cellular amount of Ire1-HA was considerably reduced by 4-. 326. PBA even in cells carrying the hrd1doa10 mutation (Fig. S2), which are beleaved to. 327. completely abolish the ERAD (Carvalho et al., 2006).. 12.
(13) 328. Nevertheless, it should also be noted that, in certain cases, 4-PBA does exhibit an. 329. ability to restore the protein-folding condition in the ER. In mammalian cells, 4-PBA. 330. supports the folding and trafficking of ER client proteins, including the F508 CFTR. 331. mutant (Rubenstein et al., 1997; Lim et al., 2004). Moreover, 4-PBA is reported to. 332. improve the viability of IRE1-gene knockout yeast cells under the presence of. 333. tunicamycin (Kubota et al., 2006). Our findings presented here indicate that 4-PBA not. 334. only has these positive effects, but also attenuates the UPR in a different manner. We. 335. speculate that 4-PBA can assist folding of particular proteins, though not able to restore. 336. global protein folding status in cells, possibly because of substrate specificity of 4-PBA.. 337. As aforementioned, since misfolding and aggregation of ER client proteins link to. 338. a wide variety of diseases, chemicals that assist protein folding and maturation in the. 339. ER are intriguing targets for drug discovery. The screening and evaluation of such drugs. 340. can be readily performed through monitoring the ER protein-folding status using. 341. reporter assays to assess the cellular level of the UPR (Iwawaki et al. 2004). However,. 342. this convenient methodology requires cautious interpretation because, as indicated here,. 343. the suppression of the UPR is not always due to an improvement of ER conditions.. 344 345. Acknowledgements. 346. We thank Dr. Kazutoshi Mori (Kyoto University) for providing materials. This work is. 347. supported by MEXT/JSPS KAKENHI grants 16K14726 to Y.K. and 24228002,. 348. 26116006 and 17H01468 to K.K.. 349 350. Conflict of interest. None declared.. 351 352. REFERENCES. 353. Bertolotti A, Zhang Y, Hendershot LM, Harding HP, Ron D. Dynamic interaction of BiP. 354. and ER stress transducers in the unfolded-protein response. Nat Cell Biol 2000; 2: 326-. 355. 32. 13.
(14) 356 357. Bicknell AA, Babour A, Federovitch CM, Niwa M. A novel role in cytokinesis reveals a. 358. housekeeping function for the unfolded protein response. J Cell Biol 2007; 177: 1017-. 359. 27.. 360 361. Carvalho P, Goder V, Rapoport TA. Distinct ubiquitin-ligase complexes define. 362. convergent pathways for the degradation of ER proteins. Cell 2006; 126: 361-73.. 363 364. Cox JS, Walter P. A novel mechanism for regulating activity of a transcription factor. 365. that controls the unfolded protein response. Cell 1996; 87: 391-404.. 366 367. Gardner BM, Walter P. Unfolded proteins are Ire1-activating ligands that directly induce. 368. the unfolded protein response. Science 2011; 333: 1891-4.. 369 370. Ishiwata-Kimata Y, Yamamoto YH, Takizawa K, Kohno K, Kimata Y. F-actin and a. 371. type-II myosin are required for efficient clustering of the ER stress sensor Ire1. Cell. 372. Struct Funct 2013; 38: 135-43.. 373 374. Iwawaki T, Akai R, Kohno K, Miura M. A transgenic mouse model for monitoring. 375. endoplasmic reticulum stress. Nat Med 2004; 10: 98-102.. 376 377. Kimata Y, Ishiwata-Kimata Y, Ito T, Hirata A, Suzuki T, Oikawa D, Takeuchi M, Kohno. 378. K. Two regulatory steps of ER-stress sensor Ire1 involving its cluster formation and. 379. interaction with unfolded proteins. J Cell Biol 2007; 179: 75-86.. 380 381. Kimata Y, Ishiwata-Kimata Y, Yamada S, Kohno K. Yeast unfolded protein response. 382. pathway regulates expression of genes for anti-oxidative stress and for cell surface. 383. proteins. Genes Cells 2006; 11: 59-69. 14.
(15) 384 385. Kimata Y, Kimata YI, Shimizu Y, Abe H, Farcasanu IC, Takeuchi M, Rose MD, Kohno. 386. K. Genetic evidence for a role of BiP/Kar2 that regulates Ire1 in response to. 387. accumulation of unfolded proteins. Mol Biol Cell 2003; 14: 2559-69.. 388 389. Kimata Y, Kohno K. Endoplasmic reticulum stress-sensing mechanisms in yeast and. 390. mammalian cells. Curr Opin Cell Biol 2011; 23: 135-42.. 391 392. Kimata Y, Oikawa D, Shimizu Y, Ishiwata-Kimata Y, Kohno K. A role for BiP as an. 393. adjustor for the endoplasmic reticulum stress-sensing protein Ire1. J Cell Biol 2004;. 394. 167: 445-56.. 395 396. Kolb PS, Ayaub EA, Zhou W, Yum V, Dickhout JG, Ask K. The therapeutic effects of 4-. 397. phenylbutyric acid in maintaining proteostasis. Int J Biochem Cell Biol 2015; 61: 45-52.. 398 399. Kubota K, Niinuma Y, Kaneko M, Okuma Y, Sugai M, Omura T, Uesugi M, Uehara T,. 400. Hosoi T, Nomura Y. Suppressive effects of 4-phenylbutyrate on the aggregation of Pael. 401. receptors and endoplasmic reticulum stress. J. Neurochem 2006; 97: 1259-68.. 402 403. Le QG, Ishiwata-Kimata Y, Kohno K, Kimata Y. Cadmium impairs protein folding in. 404. the endoplasmic reticulum and induces the unfolded protein response. FEMS yeast res. 405. 2016; 16: fow049.. 406 407. Lim M, McKenzie K, Floyd AD, Kwon E, Zeitlin PL. Modulation of deltaF508 cystic. 408. fibrosis transmembrane regulator trafficking and function with 4-phenylbutyrate and. 409. flavonoids. Am J Respir Cell Mol Biol 2004; 31: 351-7.. 410. 15.
(16) 411. Liu CY, Schroder M, Kaufman RJ. Ligand-independent dimerization activates the stress. 412. response kinases IRE1 and PERK in the lumen of the endoplasmic reticulum. J Biol. 413. Chem 2000; 275: 24881-5.. 414 415. Merksamer PI, Trusina A, Papa FR. Real-time redox measurements during endoplasmic. 416. reticulum stress reveal interlinked protein folding functions. Cell 2008; 135: 933-47.. 417 418. Mori K. Signalling pathways in the unfolded protein response: development from yeast. 419. to mammals. J Biochem 2009; 146: 743-50.. 420 421. Mori K, Sant A, Kohno K, Normington K, Gething MJ, Sambrook JF. A 22 bp cis-. 422. acting element is necessary and sufficient for the induction of the yeast KAR2 (BiP). 423. gene by unfolded proteins. EMBO J 1992; 11: 2583-93.. 424 425. Oakes SA, Papa FR. The role of endoplasmic reticulum stress in human pathology.. 426. Annu Rev Pathol 2015; 10: 173-94.. 427 428. Oikawa D, Kimata Y, Kohno K. Self-association and BiP dissociation are not sufficient. 429. for activation of the ER stress sensor Ire1. J Cell Sci 2007; 120: 1681-8.. 430 431. Okamura K, Kimata Y, Higashio H, Tsuru A, Kohno K. Dissociation of Kar2p/BiP from. 432. an ER sensory molecule, Ire1p, triggers the unfolded protein response in yeast. Biochem. 433. Biophys Res Commun 2000; 279: 445-50.. 434 435. Ozcan U, Yilmaz E, Ozcan L, Furuhashi M, Vaillancourt E, Smith RO, Gorgun CZ,. 436. Hotamisligil GS. Chemical chaperones reduce ER stress and restore glucose. 437. homeostasis in a mouse model of type 2 diabetes. Science 2006; 313: 1137-40.. 438 16.
(17) 439. Pineau L, Colas J, Dupont S, Beney L, Fleurat-Lessard P, Berjeaud JM, Berges T,. 440. Ferreira T. Lipid-induced ER stress: synergistic effects of sterols and saturated fatty. 441. acids. Traffic 2009; 10: 673-90.. 442 443. Pranke IM, Sermet-Gaudelus I. Biosynthesis of cystic fibrosis transmembrane. 444. conductance regulator. Int J Biochem Cell Biol 2014; 52: 26-38.. 445 446. Promlek T, Ishiwata-Kimata Y, Shido M, Sakuramoto M, Kohno K, Kimata Y.. 447. Membrane aberrancy and unfolded proteins activate the endoplasmic reticulum stress. 448. sensor Ire1 in different ways. Mol Biol Cell 2011; 22: 3520-32.. 449 450. Ranga Rao S, Subbarayan R, Ajitkumar S, Murugan Girija D. 4PBA strongly attenuates. 451. endoplasmic reticulum stress, fibrosis, and mitochondrial apoptosis markers in. 452. cyclosporine treated human gingival fibroblasts. J Cell Physiol 2018; 233: 60-6.. 453 454. Rubenstein RC, Egan ME, Zeitlin PL. In vitro pharmacologic restoration of CFTR-. 455. mediated chloride transport with sodium 4-phenylbutyrate in cystic fibrosis epithelial. 456. cells containing delta F508-CFTR. J Clin Invest 1997; 100: 2457-65.. 457 458. Shore GC, Papa FR, Oakes SA. Signaling cell death from the endoplasmic reticulum. 459. stress response. Curr Opin Cell Biol 2011; 23: 143-9.. 460 461. Sidrauski C, Walter P. The transmembrane kinase Ire1p is a site-specific endonuclease. 462. that initiates mRNA splicing in the unfolded protein response. Cell 1997; 90: 1031-9.. 463 464. Soto C. Unfolding the role of protein misfolding in neurodegenerative diseases. Nat Rev. 465. Neurosci 2003; 4: 49-60.. 466 17.
(18) 467. Travers KJ, Patil CK, Wodicka L, Lockhart DJ, Weissman JS, Walter P. Functional and. 468. genomic analyses reveal an essential coordination between the unfolded protein. 469. response and ER-associated degradation. Cell 2000; 101: 249-58.. 470 471. Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic. 472. regulation. Science 2011; 334: 1081-6.. 473 474. Wang M, Kaufman RJ. Protein misfolding in the endoplasmic reticulum as a conduit to. 475. human disease. Nature 2016; 529: 326-35.. 476. 18.
(19) 477. FIGURE LEGENDS. 478 479. Fig. 1. Attenuation of the HAC1 mRNA splicing by 4-PBA.. 480. (A) We added (or did not add) 4-PBA in the cultures of the wild-type IRE1 strain. 481. TKY001, and after 15 min, DTT was added. The cells were then harvested at the. 482. indicated time points, and their total RNA samples were analyzed through RT-PCR. 483. amplification of the HAC1 transcripts, which was followed by agarose gel. 484. electrophoresis and ethidium bromide staining. (B) Data from the experiment shown in. 485. panel A are numerically expressed as the HAC1 mRNA-splicing efficiency. (C) A. 486. similar experiment as in panels A and B was performed using tunicamycin instead of. 487. DTT. For dual treatment with 4-PBA and tunicamycin, tunicamycin was added into the. 488. cultures 15 min after adding 4-PBA. (D) After culturing with 4-PBA or under untreated. 489. conditions, TKY002 cells that had bZIP-Ire1 were assessed for HAC1 mRNA splicing.. 490 491. Fig. 2. 4-PBA does not diminish BiP sedimentation induced by cellular treatment. 492. with DTT.. 493. The IRE1 cells TKY001 were cultured with 4-PBA and/or DTT (or none of both. 494. (untreated)), as in Figs 1A and B. For dual treatment with 4-PBA and DTT, DTT was. 495. added into the cultures 15 min after adding 4-PBA. Total cell lysates were then obtained. 496. and fractionated by centrifugation for the BiP sedimentation assay. Panels A and B. 497. represent anti-BiP Western blot images of the total cell lysates (equivalent to 0.05 OD600. 498. cells) and the pellet samples (equivalent to 1.1 OD600 cells), respectively. Band densities. 499. were then quantified and normalized against that of the untreated samples, which are set. 500. at 1.00, before calculation of the means and standard deviations of the values from. 501. multiple cultures.. 502 503. Fig. 3. DTT impairs disulfide-bond formation in the ER even in the presence of 4-. 504. PBA. 19.
(20) 505. The IRE1 cells TKY003 producing eroGFP were cultured with 4-PBA and/or DTT (or. 506. none of both (untreated)), as in Figs 1A and B, and then observed under a florescent. 507. microscope as described in the Materials and Method section. For dual treatment with. 508. 4-PBA and DTT, DTT was added into the cultures 15 min after adding 4-PBA.. 509. Fluorescent intensities of a cell illuminated by UV/violet light (395 nm) and blue light. 510. (472.5 nm) were quantified (100 cells were observed in each sample) and used for the. 511. calculation of the indicated values.. 512 513. Fig. 4. A decrease of Ire1 in 4-PBA-treated cells at the protein level but not at the. 514. mRNA level.. 515. (A) Yeast cells producing Ire1-HA (TKY004) were cultured with 4-PBA and/or DTT (or. 516. none of both (untreated)), as performed in Figs 1A and B. For dual treatment with 4-. 517. PBA and DTT, DTT was added into the cultures 15 min after adding 4-PBA. Total cell. 518. lysates (equivalent to 0.13 OD600 cells) were then analyzed through anti-HA and anti-. 519. Pgk1 Western blotting. Band densities were quantified and are presented as in Fig. 2.. 520. (B) Wild-type IRE1 cells (TKY001) were cultured with 4-PBA or remained untreated. 521. for measuring the cellular amount of the IRE1-gene transcript. The data were obtained. 522. as relative values against the cellular TAF10-mRNA levels, which served as inner. 523. controls, and normalized against the value of the untreated sample, which is set at 1.00.. 524. (C) A similar assay as performed in panel A was performed using yeast cells producing. 525. the bZIP mutant version of Ire1-HA (YKY006).. 526 527. Fig. 5. 4-PBA promotes the degradation of Ire1.. 528. (A) For the 35S methionine/cysteine (Met/Cys) pulse label-chase experiments, cells. 529. highly producing Ire1-HA (TKY005) were cultured in SD medium as illustrated here.. 530. (B) Harvested cells from 5-ml cultures were then lysed and subjected to anti-HA. 531. immunoprecipitiation, the products of which were analyzed using SDS-DTT PAGE and. 532. autoradiography. 20.
(21) 533 534. Fig. 6. Ire1 overproduction mitigates the effect of 4-PBA to suppress the UPR.. 535. (A) Cells highly producing Ire1-HA (TKY005) were checked for the cellular Ire1-HA. 536. level as done in Fig. 4A. (B) The TKY005 cells were cultured as indicated and checked. 537. for the HAC1-mRNA splicing as done in Figs 1A and B.. 21.
(22) Graphical abstract 4-phenylbutyrate (chemical chaperone) _. Assistance of protein folding in the endoplasmic reticulum Attenuation of the unfolded protein response Previous theory. Degradation of Ire1 Attenuation of the unfolded protein response Insight from this study.
(23) A. B No 4-PBA Time after 10mM DTT addition (min). 0. 30. 60. 5mM 4-PBA 90. 0. 30. 60. 90. HAC1 Unspliced Spliced. C. 40 20 0. (+). 50. DTT addition. 0. 0. 100. HAC1 mRNA-splicing efficiency (%). HAC1 mRNA-splicing efficiency (%). 100. 60. 5 mM 4-PBA (–). 100. -15. D 80. HAC1 mRNA-splicing efficiency (%). Figure 1. bZIP-Ire1 cells 80 60 40. 20 0. 4-PBA addition. 15. 30. 45. 60. 75. Time after 10mM DTT addition (min). 90.
(24) Figure 2 A Total cell lysates. BiP. Lane number. 1. BiP-band intensity. 1.00. 2. 3. 4. 1.68 1.06 0.85 ±0.32 ±0.29 ±0.27. B Pellets. Lane number BiP-band intensity. BiP. 1. 2. 1.00. 3.31 ±1.02. 3 1.21 ±0.33. 4 3.37 ±0.38.
(25) 0 Column number. eroGFP fluorescent intensity ratio (472.5 nm light excitation / 395 nm light excitation). Figure 3 3. 2. 1. 1 2 3 4.
(26) Figure 4 A. B. C. IRE1 mRNA level. 1. Ire1-HA. bZIPIre1-HA Lane number Ire1-HA band intensity. 1. 2. 3. 4. 1.00. 1.52 ±0.32. 0.26 0.20 ±0.08 ±0.08. 0.95 ±0.37. 0.98 0.92 ±0.36 ±0.31. Pgk1. Pgk1 band intensity. 1.00. 0 bZIP-Ire1-HA band intensity. 1.00. 0.54 ±0.12.
(27) Figure 5 A 35. 4-PBA – or + Cold Met/Cys. S Met/Cys Labeling (120 min). Chase (75 min) Harvest Harvest “Before chase” “Chase 75 min”. Before chase. B. Chase 75 min. –. +. 2. 3. Ire1-HA. Lane number. 1. 5mM 4-PBA.
(28) Figure 6 A Ire1-HA. Ire1-HA 1.00 band intensity. 0.44 ±0.07. B HAC1 mRNA-splicing efficiency (%). 100 80 60. 40 20 0. Ire1-HA O.E..
(29) Figure S1 ire1D cells. Total cell lysates BiP-band intensity. BiP. 1.00. 1.11 0.86 0.76 ±0.15 ±0.19 ±0.19. BiP. Pellets. BiP-band intensity. 1.00. 1.50 1.66 ±0.28 ±0.33. 2.32 ±0.27. Fig. S1. The BiP sedimentation analysis using ire1D cells. The KMY1015 (ire1D) cells were cultured as indicated and subjected to the BiP sedimentation analysis as done in Fig. 2..
(30) Figure S2 hrd1D doa10D cells. Ire1-HA. Ire1-HA 1.00 band intensity. 0.35 ±0.07. Fig. S2. Cellular Ire1-HA level in hrd1Ddoa10D cells. TKY007 (hrd1D doa10D IRE1-HA) were checked for the cellular Ire1-HA level as done in Fig. 4A..
(31)
図



+3


関連したドキュメント
PD-L1 expression was upregulated in macrophages and dendritic cells (DCs) in high-grade invasive human OSCC tissues or co-cultured with mesenchymal-phenotype OSCC cells in
Methods: Angiopoietin-like protein-3 (ANGPTL3), LPL activity, HTGL activity, remnant lipoproteins (RLP-C & RLP-TG), small dense LDL-Cholesterol (sd LDL-C) were measured in
Liver microsomes of mouse, rat, dog and monkey (1 mg protein/mL) were incubated with 1 µM sumatriptan (A) and rizatriptan (B) with or without NADPH in the presence or absence of
In addition, more than 50% of fluorescence positive cells exhibited shrinkage and rounding even in the absence of anti-Fas antibodies (about 56, 65, and 56% of PKR-, dN-,
performed 4 h and 8 h euglycemic (5.5 mmol/l) clamps with 3 different insulin concentrations (basal, medium postprandial and high postprandial, ranging from ~ 35 to ~ 1450 pmol/l)
In immunostaining of cytokeratin using monoclonal antibodies, the gold particles were scattered in the cytoplasm of the hepatocytes and biliary epithelial cells
Consistent with the results of echocardiographic and histo- logical measurement, the mRNA expression levels of these cardiac remodeling markers were significantly decreased
It follows from [4] that a dual ovoidal subspace of H(K) is either the set of lines at distance at most 3 from a given point (type P), or the set of lines of an ideal
