2018 年 11 月 5 日発行
記録 ・ 調査報告 Note
1〒 041–8611 北海道函館市港町 3–1–1 北海道大学水産学部育種生物学講座 2〒 684–0034 鳥取県境港市昭和町 9–20 鳥取県境港水産事務所 (2017 年 12 月 13 日受付;2018 年 3 月 26 日改訂;2018 年 3 月 27 日受理;2018 年 6 月 1 日 J–STAGE 早期公開) キーワード:アメマス , 脊椎骨数 , 地域個体群 , 北海道 Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2018Masanori Oda*. 2018. Geographical variation in vertebral numbers of white-sppotted charr on Hokkaido Island, Japan. Japan. J. Ichthyol., 65(2): 187–190. DOI: 10.11369/jji.17-059. Abstract Vertebral numbers in white-spotted charr (Salvelinus leucomaenis), collected from four streams and two lakes on Hokkaido Island, Japan, were significantly higher in fish from streams flowing into the Sea of Japan compared with those flowing into the Pacific Ocean, a result compatible with previous allozyme studies. White-spotted charr in Lake Shikotsu had the lowest number of vertebrae, possibly resulting from long-term isolation. The maximum difference in mean vertebral numbers among charr populations was 1.6, twice that observed in chum salmon Onchorhynchus keta and a possible reflection of the absence of hatchery programs that can lead to population mixing. It is likely, therefore, that regional genetic structures have been maintained in Hokkaido Island white-spotted charr.
*Tottori Prefectural Fishery Office, 9–20 Showamachi, Sakaiminato, Tottori 684–0034, Japan (e-mail: [email protected] net.ne.jp)
脊
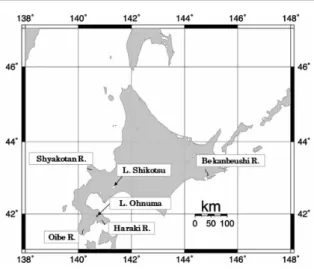
椎骨数は,古くから魚類の系群識別の指標 として用いられてきた(松宮,2000;和田, 2004).例えば,北海道におけるサケ Oncorhynchus keta の脊椎骨数は,河川(地域)間で異なってお り遺 伝的 要因の存 在が 示唆されている(久 保, 1956;Ando et al, 2011;安藤ほか,2015;安藤ほか, 2017).久保(1956)は,北海道南部の知内川のサ ケ稚魚の脊椎骨数が日本海に注ぐ石狩川水系千歳 川よりも多いことを報告している.安藤ほか(2015) は,胚発生期の水温が個体の脊椎骨数に及ぼす影 響を排除するため同じ水温条件で孵化飼育したサ ケ稚魚の脊椎骨数を複数の河川間で比較し,太平 洋に由来する河川のサケ稚魚の脊椎骨数は,日本 海に由来するサケ稚魚のものより多くなる傾向に あることを示し,脊椎骨数は遺伝的要因により決 定されることを示している.このように,脊椎骨 数は地域個体群の集団構造を推定する簡便な手法 として現在でも有効である. アメマス Salvelinus leucomaenis は,イワナ属魚 類の一種であり,分布域は朝鮮半島以北の沿海地 方,サハリン,千島列島,カムチャツカ半島など で,国内では最上川および利根川以北,北海道で の分布域は知床半島を除くほぼ全域である(鷹見, 2003).北海道では本種を専門とした漁業は行わ れておらず(鷹見,2003),種苗放流事業も行わ れていないため在来の集団構造が残っていると考 えられる.本研究は,北海道内の複数の河川およ び湖沼から採集したアメマスの脊椎骨数を比較し, 脊椎骨数が集団構造の推定に有用であるかどうか を検討した. 材 料 と 方 法 河川産アメマスの採集は 1999 年の 7 月から同 年の 11 月にかけて行った.採集河川は,北海道 渡島半島の原木川,及部川,積丹半島の積丹川,北海道のアメマスにみられた脊椎骨数の地理的変異
尾田昌紀
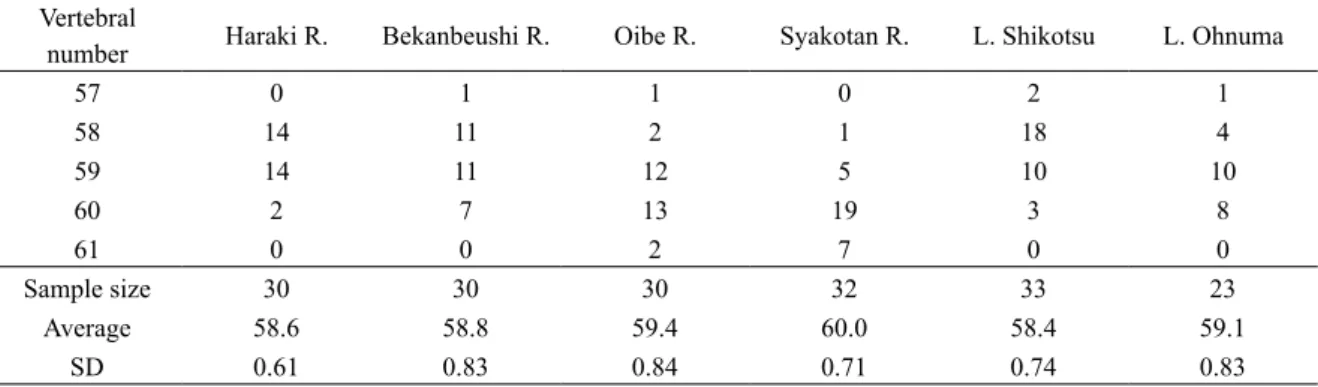
1, 2尾田昌紀 188 北海道東部の別寒辺牛川の 4 河川である(Fig. 1). このうち,原木川,及部川および積丹川は北海道 内水面漁業調整規則によって保護水面に指定され ており,堰堤には魚道が設置されているため魚類 の移動は妨げられていない.別寒辺牛川には魚類 の移動を妨げるような堰堤は設置されていない. これらの河川での標本の採集はエレクトリック ショッカー(スミス・ルート社製)を用いた. 湖沼産のアメマスの採集は,支笏湖および大沼 で行った(Fig. 1).支笏湖では 1999 年の 5 月か ら 9 月までの間に,刺網を用いて毎月 1 回ずつ採 集を行った.大沼では 1999 年の 5 月と 11 月に刺 網により大沼漁業協同組合が漁獲した個体の提供 を受けた.支笏湖は面積 76.2 km2,水深 363 m の カルデラ湖であり,約 42,000 年前に成立したと 考 え ら れ て い る( 元 田,1950; 小 野・ 五 十 嵐, 1991).支笏湖は唯一の流出河川である千歳川の 上流部には落差 5 m の滝が存在しており,また近 年には 5 つの水力発電用ダムが建設されたことか ら下流部からの魚類の遡上はない.千歳川は現在, 石狩川に合流して日本海に流出している.一方, 大沼は 2,000 年以上の駒ヶ岳の噴火により成立し たせき止め湖であり,その面積と水深はそれぞれ 5.1 km2,12 m である(元田,1950;瀬川,1974). これらの標本の採集は北海道の特別採捕許可を 得て行った.採集したアメマスは採集後,直ちに 約 10%ホルムアルデヒド水溶液で固定し,研究 室で標準体長を測定した.脊椎骨数は,軟X線写 真撮影機(ソフテックス)により写真撮影した後 に計数した.なお,2 個の尾鰭椎と尾鰭椎前第 1 椎 体 に つ い て は ま と め て 1 個 と 数 え た(藤 田, 1990).脊椎骨数について,Scheffe の方法により 各個体群間で対比較を行った. 結 果 計数した各個体群の脊椎骨数と平均値を Table 1 に示す.太平洋側河川の原木川の脊椎骨数の平 均値は 58.6 個,最頻値は 58,59 個,別寒辺牛川 の平均値は 58.8 個,最頻値は 58,59 個であった. 日本海側河川の及部川の脊椎骨数の平均値は 59.4 個,最頻値は 60 個,積丹川の平均値は 60.0 個, 最頻値は 60 個であった.支笏湖の脊椎骨数の平 均値は 58.4 個で最頻値は 58 個,大沼の平均値は 59.1 個で最頻値は 59 個であった.脊椎骨数の平 均値は最も多かった積丹川の 60 個と,最も少な かった支笏湖の 58.4 個の間で 1.6 個の差があった. 湖沼産を含む 6 個体群のアメマスの脊椎骨数に ついて個体群間で Scheffe の対比較を行った(Table 2).その結果,原木川と及部川,積丹川の間で有 意差が検出された.また,積丹川と別寒辺牛川, 支笏湖,大沼の間でも有意差が検出された.さら に,及部川と支笏湖の間にも有意差が検出された. 考 察 今回の調査結果から,北海道におけるアメマス の脊椎骨数には個体群間で変異があることが確認 された.4 河川で比較すると日本海側河川(積丹 川,及部川)のほうが太平洋側河川(別寒辺牛川, 原木川)より脊椎骨数の平均値および最頻値が高 く,原木川と及部川は地理的に近接しているにも 関わらず有意差が検出された.鷹見・木下(1990) は,北海道の函館市西方を流れる茂辺地川のアメ マスの脊椎骨数について平均値 59.3 個,最頻値 59 個と報告しており,及部川と近い値を示した. アイソザイムを用いた北海道南部(渡島半島) の河川におけるアメマスの集団遺伝学的研究によ ると,渡島半島のアメマスは太平洋側の河川集団 (桂川,川汲川)と日本海側の河川集団(知内川, 天の川,赤神川,厚沢部川)に大別される[小山 達也,未発表データ(昭和 60 年北海道大学水産 学部卒業論文)].このことから,本研究で得られ た太平洋側と日本海側の河川個体群の脊椎骨数の 差異は,遺伝的変異を反映している可能性が示唆 される.渡島半島の太平洋側と日本海側の河川集 団で遺伝的変異が検出された要因として,両河川 集団における降海型の出現頻度の違いや,津軽海 峡における日本海から太平洋に流れる強い潮流の
存在が降海型アメマスの移動にとって障壁となっ ている可能性が指摘されている[小山達也,未発 表データ(昭和 60 年北海道大学水産学部卒業論 文)].アメマスの母川回帰性について詳しいこと は分かっていないが,標識放流の結果から春に降 海するもので海洋における移動範囲は河口から 10 km 程度,初冬に降海するもので 70 km ほどと 報告されている(鷹見,2003). 本研究で確認されたアメマスの平均脊椎骨数の 差は 1.6 個で,北海道のサケの 0.7 から 0.8 個(安 藤ほか,2017)と比べても変異幅が大きい.これ は,本種を対象にした種苗放流事業が行われてい ないことに加えて,降海後も母川近くに留まるた め,サケよりも地域ごとに遺伝的分化を遂げた個 体群の存在を示唆している.そして,脊椎骨数は それらを識別する指標として有効であると考えら れる.特に支笏湖の個体群の平均脊椎骨数は 58.4 個と最も少なかった.支笏湖は約 42,000 年前に 成立し,現在は流出河川である千歳川との間に滝 やダムによって魚類の移動が妨げられているため, 支笏湖の個体群は他の個体群と比べて比較的高い 遺伝的分化を遂げている可能性がある.一方,鰓 耙数では支笏湖のアメマスは河川産の個体群より 有意に多いことが報告されており(鷹見・木下, 1990;尾田・後藤,2004),このことは長い湖沼 生活を経たことによるプランクトン食性への適応 的変異の可能性が示唆されている(尾田・後藤, 2004). 本研究から,6 個体群のみの比較ではあるが, アメマスの脊椎骨数に地理的変異が存在し,それ が遺伝的差異を反映している可能性が示された. Yamamoto et al. (2004)は,日本列島のイワナ 4 亜 種(アメマス,ニッコウイワナ,ヤマトイワナ, ゴギ)の分布域を網羅するようにサンプリングを 行い,ミトコンドリア DNA シトクロム b 領域の 部分塩基配列を決定し,それにもとづく系統地理 学的解析を行っている.その結果よると北海道の アメマスは大きく二つのクレードに分けられ,一 方のクレードは更に 3 つに細分化される.このよ うに北海道のアメマスは複雑な個体群構造を示す が,渡島半島の個体群は日本海側も太平洋側も同 一のグループにまとまり(Yamamoto et al., 2004), 脊椎骨数およびアイソザイムの解析結果と異なっ ていた.これは,解析に用いているマーカーの感 度の違い(小松・松井,2003;和田,2004)によ るものと考えられる.また,魚類の脊椎骨数は, 発生初期の水温によって変異する場合(Kobayashi, 1981;Ando et al., 2011)もあり,今回,検出され た脊椎骨数の変異は各河川の水温の影響を受けて いる可能性も排除できない.このことは,安藤ほ か(2015)が行った同一水温条件での孵化飼育実 験などを行い,更なる検証を行うことが今後の課 題である. 謝 辞 本研究を進めるうえで,元北海道大学水産学部 教授の後藤 晃博士には終始,懇切なるご指導と ご鞭撻を賜った.同大水産学部の矢部 衛特任教 Table 1. Vertebral numbers in Hokkaido Island white-spotted charr
Table 2. Multiple comparisons among white-spotted charr populations (Scheffeʼs test)
Vertebral
number Haraki R. Bekanbeushi R. Oibe R. Syakotan R. L. Shikotsu L. Ohnuma
57 0 1 1 0 2 1 58 14 11 2 1 18 4 59 14 11 12 5 10 10 60 2 7 13 19 3 8 61 0 0 2 7 0 0 Sample size 30 30 30 32 33 23 Average 58.6 58.8 59.4 60.0 58.4 59.1 SD 0.61 0.83 0.84 0.71 0.74 0.83 SD: standard deviation Population 1. 2. 3. 4. 5. 6. 1. Haraki R. – NS * ** NS NS 2. Bekanbeushi R. – – NS ** NS NS 3. Oibe R. – – – NS ** NS 4. Syakotan R. – – – – ** * 5. L. Shikotsu – – – – – NS 6. L. Ohnuma – – – – – – *P < 0.05, **P < 0.01, NS: not significant
尾田昌紀 190 授,星野浩一博士(現西海区水産研究所)にはソ フテックスの使用方法および脊椎骨の計数につい てご教示いただいた.また,地方独立行政法人北 海道立総合研究機構水産研究本部さけます・内水 面水産試験場道東センターの小山達也氏には未発 表データの引用について快諾いただいた.以上の 方々に厚く御礼申し上げる. 引 用 文 献
Ando, D., Y. Shinriki, Y. Miyakoshi, H. Urabe, T Aoyama and M. Nakajima. 2011. Seasonal variations in and effect of incubation water temperature on vertebral number in naturally spawning chum salmon Oncorhynchus keta. Fish. Sci., 77: 799–807. 安藤大成・神力義仁・下田和孝・安富亮平・佐々 木義隆・宮腰靖之・中島正道.2015.サケ稚魚の 脊椎骨数における地域差.日本水産学会誌,81; 843–845. 安藤大成・渡辺智治・下田和孝.2017.北海道の サケ稚魚にみられた脊椎骨数の地理的変異.水 産増殖,65: 89–91. 藤 田 清.1990. 魚 類 尾 部 骨 格 の 比 較 形 態 図 説. 東海大学出版会,東京.897 pp.
Kobayashi, T. 1981. Variation in number of vertebrae of sandfish, Arctoscopus japonicas (STEINDACHNER), correlated with water temperature in period of early development. Bull. Hokkaido. Reg. Fish. Res. Lab., 46: 57–67. 小池裕子・松井正文.2003.生物進化と保全生態 学.小池裕子・松井正文(編著),pp. 19–39.保 全遺伝学.東京大学出版会,東京. 久保達郎.1956.脊椎骨数に見られる知内川のサ ケ魚群の差異性.北海道大学水産学部研究彙報, 6: 266–270. 松宮義晴.2000.魚をとりながら増やす.成文堂 書店,東京.177 pp. 元田 茂.1950.北海道湖沼誌.北海道立水産孵 化場試験報告,5: 1–96. 尾田昌紀・後藤 晃.2004.湖沼産アメマス(Salvelinus leucomaenis)にみられる鰓耙数の変異.陸水生物 学報,19: 19–24. 小野有五・五十嵐八枝子.1991.北海道の自然誌. 北海道大学出版会,札幌.219 pp. 瀬川秀良.1974.日本地形誌-北海道地方.朝倉 出版,東京.303 pp. 鷹見達也.2003.アメマス.水島敏博・鳥澤 雅 (編著),pp. 118–121.新 北のさかなたち.北海 道新聞社,札幌. 鷹見達也・木下哲一郎.1990.北海道支笏湖およ び茂辺地川産のアメマスの形態比較.北海道大 学水産学部彙報,41: 121–130. 和田時夫.2004.系群.竹内俊郎・中田英昭・和 田時夫・上田 宏・有元貴文・渡部終五・中前 明(編著),pp. 141–142.水産海洋ハンドブック. 株式会社生物研究所,東京.
Yamamoto, S., K. Morita, S. Kitano, K. Watanabe, I. Koizumi, K. Maekawa and K. Takamura. 2004. Phylogeography of white-spotted charr (Salvelinus leucomaenis) infered from mitochondrial DNA sequences. Zool. Sci., 21: 229–240.
2018 年 11 月 5 日発行
記録 ・ 調査報告 Note
〒 811–3304 福岡県福津市津屋崎 4–46–24 九州大学大学院生物資源環境科学府附属水産実験所 (2018 年 3 月 23 日受付;2018 年 4 月 16 日改訂;2018 年 4 月 17 日受理;2018 年 6 月 19 日 J–STAGE 早期公開) キーワード:塩分 , 河口堰 , 河川汽水域 , 自然再生 , 生物多様性 Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2018Akihiko Koyama, Seiya Matsunaga, Tomonori Kawamoto, Kazuki Kanno, Kaito Sawa and Norio Onikura*. 2018. Spawning nests of six goby species in a newly-established fish-way on the Onga River estuary barrage. Japan. J. Ichthyol., 65(2): 191–197. DOI: 10.11369/ jji.18-014.
Abstract A new fish-way, comprising variously-sized bed materials on a gentle slope, established on the Onga River estuary barrage was found to have 69 cobbles and boulders used as spawning nests of six goby species, including a threatened species, during the period from June 2013 to May 2014. Male body sizes were positively correlated with spawning substrate. In addition, wide salinity variations recorded over the fish-way during high tides on either side of spring tide were also implicated in goby spawning site selection. *Corresponding author: Fishery Research Laboratory, Kyushu University, 4–46–24 Tsuyazaki, Fukutsu, Fukuoka 811–3304, Japan (e-mail: [email protected])
河
口域は陸水と海水の移行帯で,汽水環境を 形成するため,様々な生活様式の魚類がそ こを利用する(Potter et al., 2015).また,塩分は 汽水域の魚類群集構造や魚類相に影響を与える重 要 な 環 境 要 因 で あ る こ と か ら(Selleslagh et al., 2009;Contente et al., 2011),汽水に生息する魚類 の多様性を維持する上で塩分を含めた物理環境の 多様性を維持することが重要となる.つまり,河 口域において多種多様な魚類が生息するためには, 汽水から淡水という塩分の河川縦断的な変化が必 要不可欠である. 河口域には塩水の遡上を防ぐとともに湛水部を 作り出して水利用を安定させることを目的として, 河口堰がしばしば設置される(白川,2009).しか し,このような人工横断構造物は,通し回遊魚の 移動阻害を引き起こし,塩水遡上の遮断に伴い汽 水生物の生息に悪影響を及ぼす恐れもある(Sinha et al., 1996;玉井,2002;Yoon et al., 2016).例えば, 絶 滅 危 惧 II 類 に 選 定 さ れ て い る シ ロ ウ オ Leucopsarion petersii は(環境省,2015),大潮満潮 時のみ汽水となる場所を産卵場とする可能性が高 いことから(長谷川・小路,2017),河口堰による 塩水遡上の遮断はその産卵に負の影響を与える恐 れがある.河口堰では,魚道設置による回遊生物 の移動阻害を解消するだけでなく,汽水に生息す る生物への影響を緩和する策を講じるべきである. 福岡県の一級河川である遠賀川では,1980 年 代より河口堰が運用されている(古賀,2011). この堰は,河口から 2 km に位置しており,その 湛水区間は 9 km を超えている.つまり,その運 用によって 10 km 弱に及ぶ汽水域の環境を失った. そのため,堰の上下流で塩分が淡水から高塩分へ と劇的に変化すること,河口堰周辺は遡上魚の産 卵場所や待機場所が確保されていないことなどが, 長 年 に わ た り 問 題 視 さ れ て き た( 吉 永 ほ か, 2014).また,本堰に設置された階段式魚道は, 急勾配で流速が速く,遊泳力の小さい小型魚や底 生魚の遡上が困難であることも課題となっていた遠賀川河口部の多自然魚道で確認されたハゼ類と産卵巣
小山彰彦・松永誠弥・川本朋慶・菅野一輝・澤 海人・鬼倉徳雄
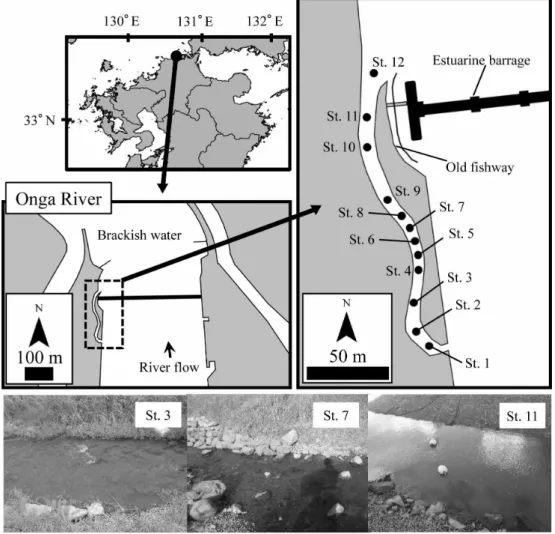
小山彰彦ほか 192 (吉永ほか,2014). これらの遠賀川河口堰を取り巻く課題を解決す るために,2008 年より遠賀川河口堰改良事業が 開始し,2012 年度に縦断勾配 1/200 程度,延長約 300 m の 多 自 然 魚 道 が 新 設 さ れ た( 吉 永 ほ か, 2014).この多自然魚道は,緩やかな蛇行を設け て延長を長くし,緩勾配とすることで,遊泳力の 乏しい小型魚の遡上を可能とするだけでなく,汽 水から淡水への塩分の連続性の確保や,魚道内に 転石を配置し,小さな瀬淵構造を形成することで, 多様な生物の生息を可能とする微環境の創出が試 みられた(吉永ほか,2014). 汽水域を利用する魚類の中で,特に定住性が高 い分類群としてハゼ亜目魚類(以下,ハゼ類とする) が挙げられる.河川感潮域,汽水域に生息する種 が多く(瀬能ほか,2004),その多様性の高さから 河口域一帯の環境指標生物として注目されている (鬼倉ほか,2008).ハゼ類のいくつかの種は,転 石やカキ殻などの硬い基質に産卵するため(石川・ 中村,1941;道津,1959,1961;松井,1986;Onikura et al., 2008,2009;鬼倉ほか,2009;Inui et al., 2010), ハゼ類が多自然魚道内に設置された多数の転石を 産卵場として利用することが期待された.また, 八代海に注ぐ球磨川河口域では,希少なハゼ類 8 種のうち,2 種がその生息を塩分に左右されること が示されており(Koyama et al., 2016),淡水から汽 水への移行帯が魚道内に形成されることで,絶滅 危惧種の定着や先のシロウオのような感潮域上端 部で産卵する種による利用も期待された.本稿では, 多自然魚道内で実施したハゼ類の産卵巣の調査結 果を報告するとともに,本河川の河口堰に新設さ れた多自然魚道の効果を考察する. 材 料 と 方 法 2013 年 6 月 か ら 2014 年 5 月 ま で, 毎 月 1 回, 調査を実施した.多自然魚道は大きく 3 蛇行して おり(Fig. 1),その蛇行部前後に自然石を設置す
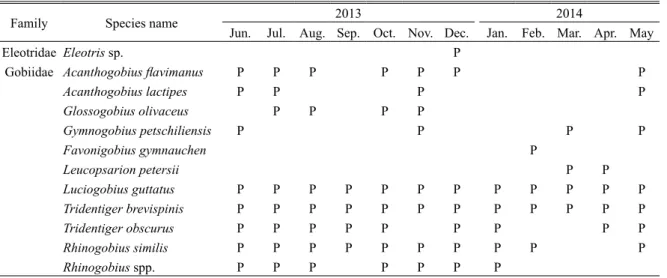
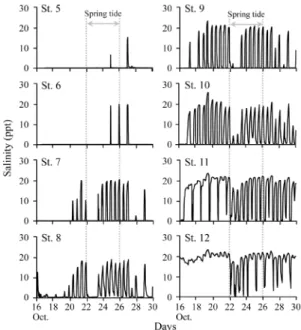
ることで,小さな瀬淵が複数形成されている.水 路幅は 2–3 m ほどの細流であり,水深は 4–30 cm と淵部でも浅いものの,流速は 5–70 cm/s と幅広 いレンジを示し,複雑な流れを有していた.満潮 時の海水遡上は 2 つ目の蛇行部までであり,それ より上流側(St. 1 から St. 4)は淡水域であった. 水温は,6 月から 10 月までは 20˚C を超え,11 月 に 12.8˚C を示し,その後 12 月から翌 2 月までは 10˚C を下回り,3 月から 4 月は再び 10˚C 以上と なり,5 月に 20˚C を超えた. まず,ハゼ類の産卵状況を把握するために,多 自然魚道内に 12ヶ所の調査地点を設置した(Fig. 1).既存の研究によって産卵基質の密度が特に高 い場所でハゼ類は産卵を行うことが報告されてい る(Inui et al., 2010).そのため,転石が特に多く 存在した St. 4 から St. 9 までの区間に調査地点を 密に設けた.各地点の上下流 2 m を調査範囲とし て,大潮の最干潮の前後 3 時間に,範囲内の転石 を調べた.ハゼ類の卵が付着した基質(産卵基質) は,定規と一緒に写真を撮影し,画像から卵塊の 面積と卵が付着している基質の底面積を計測した. また,卵塊を保護する親個体が採集された場合は, その標準体長を現地で記録した.これらの情報を 記録した後,基質と親個体を採集地に戻した.そ の後,多自然魚道内のハゼ類相を把握するために タモ網を用いた採集を実施した.採集したハゼ類 の種名を記録した後,すべて採集地に放流した. なお,カワアナゴ属の一種 Eleotris sp. は採集され た個体が小型であり,現場での同定が困難であっ たため,種を断定しなかった.また,明仁ほか (2000) に 掲 載 さ れ て い る ト ウ ヨ シ ノ ボ リ Rhinogobius sp. OR は,分類学的な精査が必要な 状況であるため(明仁ほか,2013),本研究では トウヨシノボリやシマヒレヨシノボリ Rhinogobius sp. BF(明仁ほか,2013)と思われる個体をヨシ ノボリ属の複数種 Rhinogobius spp. として扱った. 産卵場調査は干潮前後で行ったため,最下流部 を除けば,調査時には環境水から塩分を検出でき なかった.したがって,多自然魚道内の塩分の動 態を把握するため,2014 年 10 月 15 日に, 塩水 が遡上しない St. 1 から 4 を除く,8 地点の流心部 に塩分ロガー(Hobo U24 Conductivity logger, Onset Computer Corporation)を設置し,15 分間隔で塩分 を記録した.観測期間は 2 週間とし,同年 10 月 30 日に塩分ロガーを回収した. 魚道内の転石の大きさのばらつきが,様々な種 および体サイズによる産卵巣利用に寄与するかを 確かめるため,基質の底面積と卵保護個体の標準 体長について,Kendall の順位相関解析を行った. 結 果 多自然魚道内で採集されたハゼ類は,未同定種 を除くと 10 種であった(Table 1).このうち,ミ ミ ズ ハ ゼ Luciogobius guttatus と ヌ マ チ チ ブ Tridentiger brevispinis の 2 種は年間を通して採集さ れ,ゴクラクハゼ Rhinogobius similis が 10ヶ月連 続で採集された(Table 1). ハゼ類の卵の産着が確認された転石は,調査期 間中で 69 個(長径 17–57 cm)となり,このうち, Table 1. Gobioidei species observed in the new fish-way from June 2013 to May 2014
Family Species name 2013 2014
Jun. Jul. Aug. Sep. Oct. Nov. Dec. Jan. Feb. Mar. Apr. May
Eleotridae Eleotris sp. P
Gobiidae Acanthogobius flavimanus P P P P P P P
Acanthogobius lactipes P P P P Glossogobius olivaceus P P P P Gymnogobius petschiliensis P P P P Favonigobius gymnauchen P Leucopsarion petersii P P Luciogobius guttatus P P P P P P P P P P P P Tridentiger brevispinis P P P P P P P P P P P P Tridentiger obscurus P P P P P P P P P Rhinogobius similis P P P P P P P P P P Rhinogobius spp. P P P P P P P
小山彰彦ほか 194 32 個で卵保護中の雄個体も採集された(アシシ ロハゼ Acanthogobius lactipes;産卵巣 2ヶ所,ウロ ハゼ Glossogobius olivaceus,1ヶ所;シロウオ,3ヶ 所;ゴクラクハゼ,6ヶ所;ヌマチチブ,17ヶ所; チチブ Tridentiger obscurus,3ヶ所)(Fig. 2).また, 親個体が採集できなかった産卵巣 37ヶ所のうち の 2ヶ所については,卵の形状が著しく細長い棍 棒 状 で, ウ ロ ハ ゼ の 卵 の 形 状( 長 径 2.14–3.13 mm,短径 0.44–0.50 mm;鈴木ほか,1988)とよ く一致したが,同時期に産卵する他種の卵の形状 [アシシロハゼ(長径 2.4–2.6 mm,短径 0.48–0.56 mm;道津,1959),ゴクラクハゼ(長径 2.4 mm, 短径 0.45 mm;道津,1961),ヌマチチブ(長径 0.88 ± 0.07 mm, 短 径 0.58 ± 0.01 mm;Kishi, 1978), チチブ(長径 1.39 ± 0.05 mm,短径 0.72 ± 0.04 mm;Kishi, 1978)]とは大きく異なっていたため, ウロハゼと判断した. 各種の卵塊の面積を Fig. 3 に示した.種ごとに 卵塊面積は様々であったが,アシシロハゼ,シロ ウオおよびゴクラクハゼの 3 種の卵塊面積は 22 cm2以下であり,他の 3 種と比べて小さかった. 基質の底面積については,親個体が確認された 32 産卵巣のうち,基質全体が撮影されていた 31 について,親の体長との関係性を解析した結果, 有意な正の相関(Kendallʼs τ = 0.276, P < 0.05)が 認められた(Fig. 4).
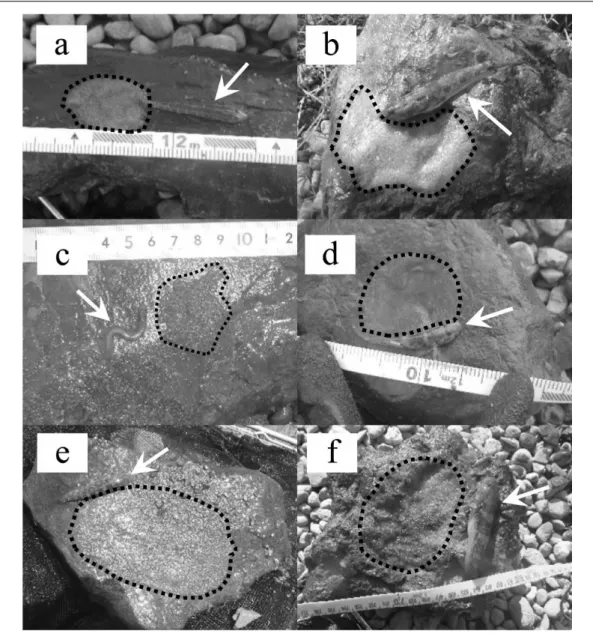
Fig. 2. Spawning nests with a male guarding the egg clutch, (a) Acanthogobius lactipes, (b)
Glossogobius olivaceus, (c) Leucopsarion petersii, (d) Rhinogobius similis, (e) Tridentiger brevispinis, (f) Tridentiger obscurus. Arrows and broken lines indicate the male goby and egg clutch of each species, respectively.
各月,各地点で確認された産卵巣の情報を Fig. 5 に整理した. 産卵巣は 2013 年 6 月から 10 月, お よ び 2014 年 4 月 か ら 5 月 に 確 認 さ れ, 特 に 2014 年 5 月に多かった.各種の産卵が確認でき た期間は異なり,複数回の調査で産卵巣を確認で きたハゼ類はヌマチチブ(5ヶ月),ゴクラクハゼ (3ヶ月),アシシロハゼ(2ヶ月)の 3 種だった. 一方,確認できた産卵巣の種類が最も多かったの は,2013 年 6 月から 8 月の 3ヶ月間であり,それ ぞれ 3 種の産卵巣が確認された.また,St. 1 から St. 9 の範囲で産卵巣が確認され,特に St. 5 で産 卵巣の確認数および産卵した種数が最も多かった. 塩分ロガーによる塩分の連続観測の結果,上流 部の地点ほど,塩水の遡上頻度が低いことが明ら かとなった(Fig. 6).特に,St. 5 と St. 6 は,大潮 から中潮にあたる 10 月 25 日から 27 日のわずか 3 日間のみ塩分の上昇が検出された.一方で,St. 9 より下流の地点では,潮汐の強弱に関係なく, 満潮時に海水の遡上が確認された. 考 察 本調査によって,10 種を超えるハゼ類が確認 され,そのうち,絶滅危惧種を含む 6 種の産卵を Fig. 3. Area of egg clutches (EA, cm2) for each species.
Al, Acanthogobius lactipes; Go, Glossogobius olivaceus; Lp, Leucopsarion petersii; Rs, Rhinogobius similis; Tb, Tridentiger brevispinis; To, Tridentiger obscurus; Un, unknown.
Fig. 5. Frequency of egg clutches (a) during the survey period, and (b) at the survey sites. Al, Acanthogobius lactipes; Go, Glossogobius olivaceus; Lp, Leucopsarion petersii; Rs, Rhinogobius similis; Tb, Tridentiger brevispinis; To, Tridentiger obscurus.
Fig. 6. Salinity at eight sites, monitored at 15 min intervals from October 16 through 30. Broken lines indicate the spring tide period.
Fig. 4. Relationship between standard length (SL; mm) and area of substrate (SA; cm2). Al, Acanthogobius lactipes; Go, Glossogobius olivaceus; Lp, Leucopsarion petersii; Rs, Rhinogobius similis; Tb, Tridentiger brevispinis; To, Tridentiger obscurus.
小山彰彦ほか 196 確認できた.ヌマチチブとゴクラクハゼの産卵期 間が他の種と比べて長く(Fig. 5a),産卵巣も多 く確認された.この 2 種は,いずれも感潮域上端 の低塩分域から淡水域を主な生息場としている(道 津,1961;瀬能ほか,2004).また,2014 年 4 月 には,シロウオの産卵巣が 3 例確認された.本種 は常時低塩分である感潮域上部,あるいは大潮満 潮時のみ汽水となる場所を産卵場として利用する 可能性が示されている(松井,1986;長谷川・小 路,2017).本調査でシロウオが産卵した地点は, St. 5 と St. 6 であり,いずれの地点も潮汐の大き い大潮から中潮の時期にのみ塩分が遡上する場所 であった(Fig. 6).2014 年 10 月に塩分の連続観 測を実施したため,シロウオの産卵期である春季 の塩分動態ではないものの,過去の知見(長谷川・ 小路,2017)と類似した結果が得られたと考えら れる. Inui et al.(2010)は,多様な産卵基質の密度を 維持することが,ハゼ類の多様性を保全する上で 重要であるとしている.今回の調査では,基質が 多く存在した St. 4 から St. 9 の範囲で産卵巣 53ヶ 所( 全 産 卵 床 の 77%) が 発 見 さ れ た(Fig. 5b). つまり,本魚道内でも,ハゼ類は基質の密度が高 い場所を産卵場として利用していると推察される. また,6 種の親個体の体長と産卵巣として利用し た基質の面積の間には,有意な正の相関が認めら れ(Fig. 4),また,卵塊面積の小さいアシシロハゼ, シロウオ,ゴクラクハゼと比べて,卵塊面積の大 きいウロハゼ,ヌマチチブ,およびチチブは,利 用する基質が大きかったことから(Fig. 3),様々 なサイズの転石の存在が複数種の産卵に寄与した と判断される. 今回の研究で魚道内に出現したにもかかわらず, 産卵巣が見つからなかった種のうち,ミミズハゼ についてはその出現時期と既知の産卵期が重なっ た.本種の産卵期は,福岡県内では 1–5 月である (道津,1957).今回の調査で,親を特定できなかっ た産卵巣は 35ヶ所あり,そのうち 13ヶ所は本種 の産卵期内である 4–5 月に見つかったことから (Fig. 5),ミミズハゼが魚道内で産卵していた可 能性があることを付記しておく. 魚道の効果を適切に評価するためには,その魚 道の設置に関わる条件,目的に応じた検討が必要 である(高橋,2000).遠賀川の多自然魚道の設 置の背景として,河口堰の上下流で塩分が劇的に 変化すること,河口堰周辺は遡上魚の産卵場所や 待機場所が確保されていないこと,また,既設の 魚道は急勾配で流速が速く,遊泳力の弱い小型魚 や底生魚の遡上が困難であることが挙げられてい る(吉永ほか,2014).そして,遠賀川河川事務 所の調査によって,多自然魚道は既設魚道よりも 出現する魚類の種数が多いこと,加えて,既設魚 道で確認されていない純淡水魚や通し回遊魚が多 く採集されたことが報告されている(吉永ほか, 2014).この報告に加えて,本調査の結果を踏ま えると,多自然魚道は,魚道の主目的である純淡 水魚や通し回遊魚の待機,移動通路としての機能 に加えて,数種のハゼ類の生息場および産卵場と して機能していることが示された.特に,魚道内 の勾配を緩やかにしたことで高塩分域から低塩分 域への移行域が創出されたこと,多様な大きさの 基質が多自然魚道内に存在したことが,基質のサ イズや塩分の選好性が異なる 6 種のハゼ類の産卵 を可能にしたと考えられる. 以上を総括すると,多自然魚道は,生物の移動 に着目した一般的な魚道の機能に加えて,河口堰 による塩水遡上遮断に伴う生態系への負の影響の 緩和に少なからず貢献していると評価できる.た だし,河口堰運用によって失われた感潮域は,10 km 弱に及ぶこと,失われた遠賀川の河川規模と 多自然魚道の規模を勘案したとき,本事業はこの 川での汽水域生態系の再生のための第一歩と位置 付けたい.そして,遠賀川と同様に,河口堰で感 潮域が減少している河川では,類似した試みを実 施することで,汽水域の生態系への負の影響を緩 和できるかもしれない. 謝 辞 本研究を進めるにあたり,当時九州大学大学院 生物資源環境科学府附属水産実験所の学生であっ た甲斐桑梓氏,大川卓也氏にご協力いただいた. また,本調査は,国土交通省遠賀川河川事務所の 許可のもと行われた.本研究の一部は,日本学術 振興会科研費萌芽研究(JP25660148)において実 施された. 引 用 文 献 明仁・坂本勝一・池田祐二・藍澤正宏.2013.ハ ゼ 亜 目. 中 坊 徹 次( 編 ),pp. 1347–1608,2109– 2170.日本産魚類検索 全種の同定 第三版.東海 大学出版会,秦野. 明仁・坂本勝一・池田祐二・岩田明久.2000.ハ ゼ亜目.中坊徹次(編),pp. 1139–1310.日本産
魚類検索 全種の同定 第二版.東海大学出版会, 東京.
Contente, R. F., M. F. Stefanoni and H. L. Spach. 2011. Fish assemblage structure in an estuary of the Atlantic Forest biodiversity hotspot (southern Brazil). Ichthyol. Res., 58: 38–50. 道津喜衛.1957.ミミズハゼの生活史.九州大学 農学部学芸雑誌,16: 93–100. 道 津 喜 衛.1959. ア シ シ ロ ハ ゼ の 生 態・ 生 活 史. 長崎大学水産学部研究報告, 8: 196–201. 道 津 喜 衛.1961. ゴ ク ラ ク ハ ゼ の 生 態・ 生 活 史. 長崎大学水産学部研究報告,10: 120–125. 長谷川拓也・小路 淳.2017.広島県三津大川に おけるシロウオの遡上および産卵場の環境特性. 日本水産学会誌,83: 574–579.
Inui, R., N. Onikura, M. Kawagishi, M. Nakatani, Y. Tomiyama and S. Oikawa. 2010. Selection of spawning habitat by several gobiid fishes in the subtidal zone of a small temperate estuary. Fish. Sci., 76: 83–91.
石川 昌・中村中六.1941.ウロハゼ Glossogobius brunneus (TEMMICK et SCHLEGEL) の生活史.水 産学会報,8: 1–16,pls. 1–2.
環境省.2015.レッドデータブック 2014―日本の 絶滅のおそれのある野生生物―4 汽水・淡水魚類. ぎょうせい,中央区.414 pp.
Kishi, Y. 1978. Egg size difference among three populations of the goby, Tridentiger obscurus. Jpn. J. Ichthyol., 24: 278–280.
古賀邦雄.2011.ダムの書誌あれこれ (91) 遠賀川 福智山ダム・遠賀川河口堰.月間ダム日本,801: 73–84.
Koyama, A., R. Inui, H. Iyooka, Y. Akamatsu and N. Onikura. 2016. Habitat suitability of eight threatened gobies inhabiting tidal flats in temperate estuaries: model developments in the estuary of the Kuma River in Kyushu Island, Japan. Ichthyol. Res., 63: 307–314. 松井誠一.1986.シロウオの生態と増殖に関する
研究.九州大学農学部学芸雑誌,40: 135–174. Onikura, N., R. Inui, K. Eguchi, T. Nishida and J.
Nakajima. 2008. Spawning site of " Kuroko-haze" (Drombus sp.) (Pisces, Gobiidae) in eastern Kyushu Island, Japan: New record from the field. Biogeography, 10: 13–16.
Onikura, N., J. Nakajima, T. Nishida, R. Inui, K. Eguchi, M. Nakatani and S. Oikawa. 2009. Spawning sites of Eleotris acanthopoma (Perciformes: Eleotridae) in the Kitagawa and Sumiegawa Rivers, Kyushu Island, Japan. Ichthyol. Res., 56: 105–107.
鬼倉徳雄,西田高志,乾 隆帝,中島 淳,江口 勝久,及川 信.2009.宮崎県北川の汽水域ワン ドに見られるハゼ亜目魚類の産卵場と横断分布. 日本生物地理学会会報,64: 29–39.
Potter, I. C., J. R. Tweedley, M. Elliott and A. K. Whitfield. 2015. The ways in which fish use estuaries: a refinement and expansion of the guild approach. Fish Fish., 16: 230–239.
Selleslagh, J., R. Amara, P. Laffargue, S. Lesourd, M. Lepage and M. Girardin. 2009. Fish composition and assemblage structure in three Eastern English Channel macrotidal estuaries: a comparison with other French estuaries. Estuar. Coast. Shelf Sci., 81: 149–159. 白 川 直 樹.2009. 河口堰. 高橋 裕・岩屋隆夫・ 沖 大幹・島谷幸宏・寶 馨・玉井信行・野々 村邦夫・藤芳素生(編),pp. 206–207.川の百科 事典.丸善,東京. 瀬能 宏・鈴木寿之・渋川浩一・矢野維畿.2004. 決定版 日本のハゼ.平凡社,東京.536 pp. Sinha, M., M. K. Mukhopadhyay, P. M. Mitra, M. M.
Bagchi and H. C. Karamkar. 1996. Impact of Farakka barrage on the hydrology and fishery of Hoogly estuary. Estuaries, 19: 710–722. 鈴木伸洋・柵瀬信夫・杉原拓郎.1988.ウロハゼ の 卵 発 生 と 仔 稚 魚 の 形 態 変 化. 水 産 増 殖,35: 203–212. 高橋剛一郎.2000.魚道の評価をめぐって.応用 生態工学,3: 199–208. 玉井信行.2002.長良川河口堰モニタリング調査 を評価する.応用生態工学,5: 1–3.
Yoon, J. D., M. H. Jang, H. B. Jo, K. S. Jeong, G. Y. Kim and G. J. Joo. 2016. Changes of fish assemblages after construction of an estuary barrage in the lower Nakdong River, South Korea. Limnology, 17: 183–197.
吉永勝彦・石坪昭二・深浦貴之・田中 聡.2014. 遠賀川河口堰多自然魚道における施工上の配慮 と工夫について.平成 26 年度 国土交通省国土 技術研究会論文集,291–295.
2018 年 11 月 5 日発行
記録 ・ 調査報告 Note
1〒 780–8520 高知県高知市曙町 2–5–1 高知大学大学院黒潮圏総合科学専攻 2〒 755–8611 山口県宇部市常磐大 2–16–1 山口大学理工学部 3〒 649–2211 和歌山県西牟婁郡白浜町 459 京都大学フィールド科学教育研究センター瀬戸臨海実験所 4現住所:〒 625–0052 京都府舞鶴市長浜番外地 京都大学フィールド科学教育研究センター舞鶴水産実験所 (2018 年 4 月 30 日受付;2018 年 6 月 7 日改訂;2018 年 6 月 9 日受理;2018 年 9 月 10 日 J–STAGE 早期公開) キーワード:ウキゴリ属 , 北限記録 , COI, 記載 Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2018Yumi Henmi*, Ryutei Inui, Ryutaro Goto and Gyo Itani. 2018. First record of Gymnogobius macrognathos on Akkeshi mud flat, Hokkaido, Japan and utilization of Upogebia major burrows. Japan. J. Ichthyol., 65(2): 199–203. DOI: 10.11369/jji.18-021.
Abstract Eight specimens of a threatened goby, Gymnogobius macrognathos (30.04– 38.87 mm in standard length), were collected on a mudflat characterized by abundant burrows of Upogebia major, at Akkeshi, Hokkaido, northern Japan in August 2017. Because G. macrognathos specimens were collected directly from U. major burrows by suction pump, and were also collected with U. major by digging sediment exposed at low tide, it was concluded that G. macrognathos utilized U. major burrows on the tidal flat. Gymnogobius mororanus and an unidentified species of Gymnogobius also utilized U. major burrows. The presence of G. macrognathos at Akkeshi represents the northernmost record of the species in Japanese waters, the isolated population being ca. 500 km north of the main distributional range of the species along the Pacific coast of Japan.
*Corresponding author: Maizuru Fisheries Research Station, Kyoto University, Nagahama, Maizuru, Kyoto, 625–0086 Japan (e-mail:[email protected])
エ
ドハゼ Gymnogobius macrognathos Bleeker, 1860 は日本およびロシアのオルガ湾,ピョートル 大帝湾,朝鮮半島西岸,中国山東省に分布する (Stevenson, 2002;明仁ほか,2013).本種は,国内 では宮城県から宮崎県の太平洋沿岸,瀬戸内海, 京都府および兵庫県,福岡県から熊本県の日本海・ 東シナ海沿岸に分布し,汽水域に生息する(乾, 2015).その生息場は,主に砂泥底で,アナジャコ 類やスナモグリ類の巣穴を隠れ家や産卵場として 利 用 す ること が 知 ら れ て い る(Kinoshita, 2002; Kinoshita et al., 2010;Koyama et al., 2017;Henmi et al., 2018).上記の分布域以外に,北海道やサハリンにおい て記録があるものの(Tanaka, 1908;Jordan et al., 1913),標本に基づいたものではないため,再検
討 す る こ と が 不 可 能 で あ る(Dyldin and Orlov, 2017).筆者らは,2017 年 8 月の干潮時に北海道 厚 岸 郡 の 干 潟 に お い て, ア ナ ジ ャ コ Upogebia major の巣穴からエドハゼを採集した.この採集 地は,エドハゼの北海道における新産地であり, 日本における分布の北限となった.よって,ここ に本標本の新産地記録および北海道における初め ての標本に基づく記録として記述する.本種は環 境省レッドリスト 2014 で絶滅危惧 II 類に指定さ れているが(乾,2015),干潟に生息するハゼ類は, 採集圧による個体数の減少よりも生息場所が知ら れないまま公共工事などで生息地の消失や改変の 可 能 性 が 高 い と 考 え ら れ て い る( 乾・ 小 山, 2014).そのため,本論文では採集地の風景を示 した(Fig. 1).
北海道厚岸郡におけるエドハゼ Gymnogobius macrognathos
の記録およびアナジャコの巣穴利用
邉見由美
1, 4・乾 隆帝
2・後藤龍太郎
3・伊谷 行
1邉見由美ほか 200 2017 年 8 月 9 日,10 日に北海道厚岸郡厚岸町 のアナジャコ Upogebia major が高密度に生息する 泥干潟で調査を行った.9 日は,干潮時に干潟海 側の潮下帯水深 10 cm 程度の箇所と干潟内のタイ ドプールの冠水した箇所で,ヤビーポンプを用い てアナジャコの巣穴入り口 1 つから各 3 回吸引し た.この作業を海側とタイドプール内で各 45 回 行った.10 日は,干潮時に干出した砂泥底にて おおよそ 1 m × 1 m × 50 cm の掘り返し採集を,シャ ベルを用いて 5 回行った. 標本の計数および計測方法は Stevenson(2002) に従った.計測値はデジタルノギスを用いて 0.01 mm 単位で測定した.標準体長は SL で,全長は TL でそれぞれ略記した . 本研究で観察した標本 は高知大学魚類標本コレクション(BSKU)に登 録されている . 採集時の水温は 24.6–25.9˚C,塩分 は 29–33 で あ っ た. 厚 岸 の 最 干 潮 は,2017 年 8 月 9 日 の 10 時 18 分 平 均 水 面 -69 cm,10 日 の 10 時 48 分平均水面 -66 cm であった(第一管区海上 保安本部海洋情報部,2018). 9 日の吸引採集では,海側からは,ヘビハゼ Gymnogobius mororanus Jordan and Snyder, 1901 が 1 個体採集された.タイドプール内では,エドハゼ が 1 個体,ヘビハゼが 10 個体,ウキゴリ属未同 定種が 1 個体採集された.10 日の掘り起こし採 集では,エドハゼが 7 個体,ヘビハゼが 12 個体, ウキゴリ属未同定種が 3 個体採集された. 得られたエドハゼ 8 個体を記載に用い,そのう ち 2 個体(BSKU 123559, 123560)を DNA 解析に 用いた.比較のため,愛知県名古屋市藤前干潟産 エドハゼ1個体(BSKU 123592,2017 年 9 月 6 日 . 邉見由美・伊谷 行採集)を含め,組織を一部切 り出し,DNeasy Blood & Tissue Kit(QIAGEN)を 用 い て DNA 抽 出 を 行 な っ た. ミ ト コ ン ド リ ア DNA の COI(cytochrome c oxidase subunit I) 遺 伝 子 の 部 分 領 域 を, プ ラ イ マ ー LCO1490 と HCO2198(Folmer et al., 1994)を用いて PCR 法に より増幅した.PCR 反応液の組成は 1 サンプル あたり,蒸留水 17.5 µL,10 × ExTaq Buffer 2.5 µL, dNTP Mixture 2.0 µL,プライマー(10 µM)各 0.3 µL,TaKaRa ExTaq 0.13 µL,鋳型 DNA 2.5 µL であ る.温度条件は,94 ˚C 変性 3 分を 1 回,94 ˚C 変 性 45 秒,45 ˚C アニーリング 1 分 30 秒,72 ˚C 伸 長 1 分を 35 回,72 ˚C 伸長 4 分を 1 回とした.得 られた PCR 産物はアガロースゲル電気泳動で増 幅 を 確 認 後,ExoSap-IT(Thermo Fisher Scientific) を 使 っ て 精 製 し た. シ ー ケ ン シ ン グ は Eurofins Genomics 社の DNA シーケンスサービスを利用し た.得られたシーケンスデータは Seaview(Galtier et al., 1996;Gouy et al., 2010)で確認し,659 bp に ついて決定し比較を行った[日本 DNA データバ ンク(DDBJ)のアクセッション番号:LC381670– LC381672].
Gymnogobius macrognathos Bleeker, 1860 エドハゼ(Fig. 2)
記載標本 8 個体(30.04–38.87 mm SL):BSKU Fig. 1. Collection site of Gymnogobius macrognathos at Akkeshi, Hokkaido, Japan, showing a tidal pool to the left and open water to the right.
123559,36.73 mm SL,45.92 mm TL,北海道厚岸 郡 厚 岸 湖(43˚03'N, 144˚51'E),2017 年 8 月 9 日, ヤビーポンプ,邉見由美・伊谷 行採集.BSKU 123560–123566,30.04–38.87 mm SL,36.60–47.32 mm TL,北海道厚岸郡厚岸湖(43˚03'N, 144˚51'E), 2017 年 8 月 10 日,シャベルと手網,邉見由美・ 伊谷 行採集. 記 載 計 数 値: 背 鰭 条 数 VI-I, 11–13( 最 頻 値 VI-I, 11);臀鰭条数 I, 9–10(I, 9 と I, 10);胸鰭軟 条数 19–20(20);腹鰭条数 I, 5;縦列鱗数 42-49 (44);横列鱗数 5-9(8). 体 各 部 の SL に 対 す る 割 合(%): 頭 長 27.5– 30.2;眼径 4.5–5.0;両眼間隔 2.2–3.7;吻長 7.3–8.1; 胸 鰭 基 部 で の 体 高 13.0–15.5; 尾 柄 長 16.3–20.4; 尾柄高 7.5–9.6. 体に鱗が不規則に並ぶ.上顎先端は下顎先端と ほぼ同じ位置にある.下顎峡部に肉質皮弁がない. 両眼間隔にある感覚孔の開口数は 2 個ある. 生鮮時の色彩はやや黄色みを帯びた褐色で腹部 は白い.頭部と体背側に雲状斑がある.体側に数 本の不明瞭な暗色横線があり腹部に斑紋がない. 尾鰭はやや黄色みを帯び,下部 1/3 を除き横縞模 様を持つ.第 1 背鰭,第 2 背鰭に縞模様の黒色斑 を有する.固定後の色彩は生鮮時よりも退色する が,体側にみられる数本の不明瞭な暗色横線は残 る. 分布(Fig. 3) 本種は,日本,ロシア(オルガ湾, ピョートル大帝湾),朝鮮半島(東津江,満頃江, 錦江),中国山東省(煙台市,青島市)から記録 がある(Stevenson, 2002;明仁ほか,2013).日本 国内では,北海道太平洋沿岸(厚岸),宮城県か ら宮崎県の太平洋沿岸,瀬戸内海,京都府および 兵庫県,福岡県から熊本県の日本海・東シナ海沿 岸 か ら 報 告 さ れ て い る( 明 仁 ほ か,2013; 乾, 2015;本研究). 備考 調査標本の形態は,G. macrognathus のホ ロタイプに基づいた Stevenson (2002)の記載によ く一致した.胸鰭条数は BSKU 123559,123561, 123562 が 19 であり,Stevenson(2002)の示した エドハゼの特徴である胸鰭条数 20 とは異なって いた.また,背鰭条数は BSKU 123564 のみが VI-I, 13 であり,Stevenson(2002)の示した背鰭条数 VI-I, 10-12 と は 異 な っ て い た. さ ら に,BSKU 123561 は縦列鱗数が 55 であり,Stevenson(2002) の示した縦列鱗数 45–50 とは異なっていた.しか し,これらの個体は,その他の特徴が Stevenson (2002) と 明 仁 ほ か(2013) が 示 す エ ド ハ ゼ G. Fig. 2. Gymnogobius macrognathos (BSKU 123565, 32.15 mm SL) from a mudflat at Akkeshi, Hokkaido, Japan (preserved specimen).
Fig. 3. Distribution of Gymnogobius macrognathos. Broken lines indicate distribution ranges of G. macrognathos. Black star indicates collection site of the present study.
邉見由美ほか 202 macrognathos と一致したこと,同属他種では胸鰭 軟 条 数 や 背 鰭 軟 条 数 に 変 異 が 見 ら れ る こ と (Stevenson, 2002;明仁ほか,2013),これらは個 体変異であると考えられる.同所的に採集された ヘビハゼは,上顎先端が下顎先端より後方に位置 すること,上顎後端が眼の後縁をはるかに超える ことなどによりエドハゼから識別できる(明仁ほ か,2013). DNA 解析の結果,厚岸産 2 個体の間には変異 はなく,厚岸産 2 個体と藤前干潟産個体の間では, 2 塩基の変異が見られた(約 0.3% の集団間変異). この値は,一般にハゼ類で確認されている種間変 異 の 値 に 比 べ て は る か に 小 さ く(Matsui et al., 2012;Knebelsbergera and Thiel, 2014),種内変異だ と考えられる.このことから,今回厚岸から採集 さ れ た 個 体 は, 遺 伝 的 に も エ ド ハ ゼ(G. macrognathos)であることが支持された.
エドハゼは,アナジャコ,ヨコヤアナジャコ Upogebia yokoyai の 巣 穴 を 隠 れ 家 と し て 利 用 し (Kinoshita, 2002;Kinoshita et al., 2010),産卵場と し て は 主 に ニ ホ ン ス ナ モ グ リ Nihonotrypaea japonica の巣穴を利用することが知られている (Koyama et al., 2017;Henmi et al., 2018).また,ヘ ビハゼもヨコヤアナジャコの巣穴を隠れ家として 利 用 す る こ と が 知 ら れ て い る(Kinoshita et al., 2010).本調査地には,砂泥表面の形状からアナ ジャコが構築したと考えらえる巣穴が多数確認さ れ(Kinoshita, 2002;鈴木ほか,2013),実際にア ナジャコの標本も採集されたことから,アナジャ コが高密度に生息していると考えられた.本調査 地において,エドハゼとヘビハゼが,ヤビーポン プによりアナジャコの巣穴から直接採集され,干 出時の掘り返しによりアナジャコの巣穴のみが確 認される場所から採集された.これらのことから, 既存の知見と同様に,本調査地においても,エド ハゼとヘビハゼは,干潮時にアナジャコの巣穴を 隠れ家として用いていたことが推察される. ヤビーポンプによる採集により,北海道厚岸湖 におけるエドハゼのアナジャコの巣穴への共生率 は 1%(計 1 個体 / 90 巣穴開口部),ヘビハゼの共 生率は 11%(計 10 個体 / 90 巣穴開口部)である ことが明らかになった.ただし,採集結果は 2, 3 回目の吸引個体も含まれる.2,3 回目の吸引に より,地下で巣穴が破壊されて付近の他の巣穴に いたハゼが採集された可能性もあるため,この値 は過大評価になるだろう.一方,共生性ハゼ類が 巣穴の下部にいた場合にはヤビーポンプで採集さ れないため,この手法で求めた共生率は過小評価 ともなりうる.これらの共生率の精度については, 今後,行動観察などにより詳細な研究を経た考察 が望まれる. これまでエドハゼの国内における分布北限とし て宮城県河北町長面浦が知られていた(鈴木・増 田,1993).本研究により,宮城県河北町よりお よそ 500 km 北方に更新され,本調査地が国内分 布の北限となった.今後の詳細な調査により,ア ナジャコ類の巣穴からの採集を行えば,京都以北 の日本海や,青森県および岩手県でエドハゼの分 布が確認されるかもしれない. 一方,近年,干潟の埋め立てや護岸整備など, 生息環境の悪化などによって本種の生息地は減少 しており,環境省レッドリスト 2014 では本種は 絶滅危惧 II 類に指定されている(乾,2015).逸 見ほか(2014)で指摘されているように,共生性 の種の保全には,宿主の保全が求められている. エドハゼの生息にはアナジャコ類やスナモグリ類 の巣穴が必要であるとすると,これら宿主が生息 する干潟自体の保全が必要である.より効果的な 保全のためには,本種による宿主の巣穴利用に加 えて,底質や塩分などどのような環境条件が本種 の生息に必要であるかを明らかにする必要がある (例えば,Koyama et al., 2016;Inui et al., 2018).
謝 辞 本研究を行うにあたり調査地の情報をくださっ た産業技術総合研究所の清家弘治博士,同定にご 協力いただいた京都大学の岩田明久博士,標本登 録に際して便宜を図っていただいた高知大学の遠 藤広光博士,施設を使用させていただいた北海道 大学北方生物圏フィールド科学センター水圏ス テーション厚岸臨海実験所の皆様に謹んで感謝の 意を表する.担当編集委員の甲斐嘉晃博士,なら びに,匿名査読者 2 名には,本稿の改訂に際し, 有益なコメントをいただいた.ここに記して,感 謝の意を表する . また,本研究の一部は JSPS 科 研費 15J12531,16K07233 の助成を受けて行われ た. 引 用 文 献 明仁・坂本勝一・池田祐二・藍澤正宏.2013.ハ ゼ亜目.中坊徹次(編著),日本産魚類検索全種 の同定 第三版.p. 1482.東海大学出版会,泰
野.
Bleeker, P. 1860. Zesde bijdrage tot de kennis der vischfauna van Japan, Acta. Soc. Reg. Sci. Indo-Neêrland., 8: 1–104.
第一管区海上保安本部海洋情報部.2018.潮汐カ レンダー2017 年 8 月厚岸:http://www1.kaiho.mlit. go.jp/KAN1/kaisyou/tide/suisan.cgi?mode=search&year =2017&month=08&ichi=52(参照 2018-05-31). Dyldin, Y. V. and A. M. Orlov. 2017. Ichthyofauna of fresh
and brackish waters of Sakhalin Island: An annotated list with taxonomic comments: 4. Pholidae–Tetraodontidae families. J. Ichthyol., 57: 183–218.
Folmer, O., M. Black, W. Hoeh, R. A. Lutz and R. Vrijenhoek. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol., 3: 294–299.
Galtier, N., M. Gouy and C. Gautiel. 1996. SEAVIEW and PHYLOWIN: two graphic tools for sequence alignment and molecular phylogeny. Comput. Appl. Biosci., 12: 543–548.
Gouy, M., S. Guindon and O. Gascuel. 2010. Seaview version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol., 27: 221–224.
Henmi, Y., K. Eguchi, R. Inui, J. Nakajima, N. Onikura and G. Itani. 2018. Field survey and resin casting of Gymnogobius macrognathos spawning nests in the Tatara River, Fukuoka Prefecture, Japan. Ichthyol. Res., 65: 168–171. 逸見泰久・伊谷 行・岩崎敬二・西川輝昭・佐藤 正典・佐藤慎一・多留聖典・藤田喜久・福田 宏・ 久保弘文・木村妙子・木村昭一・前之園唯史・ 松 原 史・ 長 井 隆・ 成 瀬 貫・ 西 栄 二 郎・ 大澤正幸・鈴木孝男・和田恵次・渡部哲也・山 西良平・山下博由・柳 研介.2014.日本の干潟 における絶滅の危機にある動物ベントスの現状 と課題.日本ベントス学会誌,69: 1–17. 乾 隆帝.2015.エドハゼ.環境省自然環境局野 生生物課希少種保全推進室(編),レッドデータ ブック 2014―日本の絶滅のおそれのある野生生 物 4 汽水・淡水魚類.pp. 320–321.ぎょうせ い,東京. 乾 隆帝・小山彰彦.2014.本州・四国・九州の 河 口 干 潟 に 生 息 す る ハ ゼ 類 . 魚 類 学 雑 誌,61: 105–109.
Inui, R., A. Koyama and Y. Akamatsu. 2018. Abiotic and biotic factors influence the habitat use of four species of Gymnogobius (Gobiidae) in riverine estuaries in the Seto
Inland Sea. Ichthyol. Res., 65: 1–11.
Jordan, D. S. and J. O. Snyder. 1901. A review of the gobioid fishes of Japan, with descriptions of twenty-one new species. Proc. U.S. Nat. Mus., 24: 33–132.
Jordan, D. S., S. Tanaka and J. O. Snyder. 1913. A catalogue of the fishes of Japan. J. Coll. Sci., Tokyo Imper. Univ., 33: 1–497.
Kinoshita, K. 2002. Burrow structure of the mud shrimp Upogebia major (Decapoda: Thalassinidea: Upogebiidae). J. Crust. Biol., 22: 474–480.
Kinoshita, K., G. Itani and T. Uchino. 2010. Burrow morphology and associated animals of the mud shrimp Upogebia yokoyai (Crustacea: Thalassinidea: Upogebiidae). J. Mar. Biol. Assoc. U.K., 90: 947–952.
Knebelsberger, T. and R. Thiel. 2014. Identification of gobies (Teleostei: Perciformes: Gobiidae) from the North and Baltic Seas combining morphological analysis and DNA barcoding. Zool. J. Linn. Soc., 172: 831–845. Koyama, A., R. Inui, H. Iyooka, Y. Akamatsu and N.
Onikura. 2016. Habitat suitability of eight threatened gobies inhabiting tidal flats in temperate estuaries: model developments in the estuary of the Kuma River in Kyushu Island, Japan. Ichthyol. Res., 63: 307–314. Koyama, A., R. Inui, K. Umemura, M. Wakabayashi, K.
Kanno and N. Onikura. 2017. The first record of the spawning nest of Gymnogobius cylindricus and Gymnogobius macrognathos. Ichthyol. Res., 64: 261–263. Matsui, S., K. Nakayama, Y. Kai and Y. Yamashita. 2012. Genetic divergence among three morphs of Acentrogobius pflaumii (Gobiidae) around Japan and their identification using multiplex haplotype-specific PCR of mitochondrial DNA. Ichthyol. Res., 59: 216– 222.
Stevenson, D. E., 2002. Systematics and distribution of fishes of the Asian goby genera Chaenogobius and Gymnogobius (Osteichthyes: Perciformes: Gobiidae), with the description of a new species. Species Divers., 7: 251–312. 鈴木孝男・木村昭一・木村妙子・森 敬介・多留 聖典.2013. 干潟ベントスフィールド図鑑.特定 非営利活動法人 日本国際湿地保全連合,東京, 257pp. 鈴木寿之・増田 修.1993. 兵庫県で再発見された キセルハゼと分布上興味あるハゼ科魚類 4 種. I.O.P. Diving News, 4: 2–6.
Tanaka, S. 1908. Notes on a collection of fishes made by Prof. Ijima in the southern parts of Sakhalin. Ann. Zool. Jpn., 6: 235–254.
2018 年 11 月 5 日発行
記録 ・ 調査報告 Note
1〒 305–0053 茨城県つくば市小野川 4 番地 1 2〒 270–1194 千葉県我孫子市我孫子 1646 電力中央研究所 環境科学研究所 (2018 年 1 月 29 日受付;2018 年 7 月 24 日改訂;2018 年 7 月 25 日受理;2018 年 10 月 2 日 J–STAGE 早期公開) キーワード:アユ , なわばり , 防衛 , 垂直距離 , 水平距離 Japanese Journal of Ichthyology © The Ichthyological Society of Japan 2018Haruo Honda* and Ryosuke Yamamoto. 2018. Height and horizontal distance of territorial attacks by ayu (Plecoglossus altivelis altivelis) were observed in aquaria. Japan. J. Ichthyol., 65(2): 205–209. DOI: 10.11369/jji.18-003.
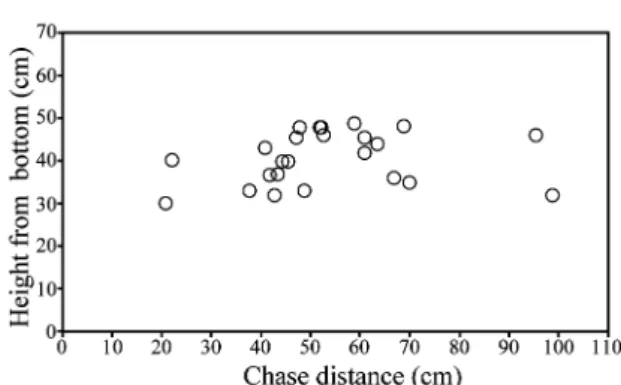
Abstract The height and horizontal distance of territorial attacks by ayu (Plecoglossus
altivelis altivelis) were observed in aquaria. Territory holders responded towards intruders when approached to within 30–40 cm, chasing them for 40–100 cm. Such chases ceased ca. 30–40 cm from the bottom (in 50–100 cm water depth), indicating that territorial defense limits in water depths greater than 50 cm were about 30–40 cm (vertical distance) and 40–100 cm (horizontal distance).
*Corresponding author: Environmental Science Research Laboratory, Central Research Institute of Electric Power Industry, 4–1 Onogawa, Tsukuba, Ibaraki, Japan (e-mail: [email protected])
ア
ユ Plecoglossus altivelis altivelis は,日本の河 川中流域を代表する魚として親しまれ,漁 業・遊漁や養殖の対象としても重要であることか ら,多くの研究がなされている.また,河川流量 の保持など,河川環境の保全を図る点でも指標的 な魚種の一つとなっている. よく知られているように,初夏から夏にかけて の成長期には,アユは,瀬の岩盤や礫の河床にな わばりを作り,付着藻類を摂餌する(例えば,宮 地 ほ か,1952; 川 那 部,1957; 水 野・ 川 那 部, 1957;立原・木村,1988;Iguchi and Hino, 1996). そ し て, 近 年, 亜 種 の リ ュ ウ キ ュ ウ ア ユ P. altivelis ryukyuensis の 摂 餌 な わ ば り に つ い て も, 本州産のアユと同様に機能していることが明らか にされている(Awata et al., 2011).しかし,これ までのアユなわばりの研究では,なわばりは平面 図的に観察されたものが多く,河床底との関係は 十分明らかになっていない.また,水槽でアユの なわばり行動を側面から観察すると,なわばり個 体は侵入者を水底付近から追いかけた後,水底か らほぼ一定の垂直距離で侵入者を解放することが 観察されるが,このようなアユのなわばり行動の 報告は,ほとんど見ることができない.本研究で は,アユのなわばり行動を側面から観察し,なわ ばり個体の侵入者を追撃する距離,および攻撃を 止めた時の底からの垂直距離について明らかにす ることができたので,ここに報告する. 材料および方法 供試魚 アユは,千葉県水産総合研究センター 内水面研究所が生産した人工種苗を手賀沼漁業協 同組合(千葉県柏市)が自然日長下で育成したも のを 2007 年および 2009 年の 6 月から 8 月に購入 した.購入後,水中ポンプを用い,水流を起こし た水量 2000 l の循環濾過水槽 (室内設置)に収容水槽で観察したアユなわばりの垂直距離と水平距離
本田晴朗
1・山本亮介
2本田晴朗・山本亮介 206 し,市販のアユ用配合飼料を与え,水温 22–26˚C で 2 週間の予備飼育を行った後,観察に用いた. 供試アユの体長(SL)は 13.3–20.5 cm であった. 実験水槽 観察にはアクリル窓付 FRP 水槽(アー ス社),濾過槽,温度調節装置からなる循環濾過 シ ス テ ム を 2 系 統 用 い た(Fig. 1). 水 槽 1 は, 2.5×0.5×0.7 m 水槽 (最大観察可能水深 0.6 m)に 水中ポンプを用い,水槽底面に成長期のアユの選 好流速 30–70 cm/S(本田・山本,2006) を発生さ せた.また,なわばり形成の対象物として,水槽 底に 25×25×5 cm のコンクリ-トブロック 3 個を 配置した.この水槽 1 は,水深 30–60 cm の行動 観察に用いた(Fig. 2).水槽 2 は 1.0×0.5×1.1 m 水 槽(最大観察可能水深 1.0 m)で,水深 30–100 cm での行動観察に用いた.水槽 2 は長さが 1 m と短 いため,水槽底面での流速は 10–20 cm/S と選好 流速より遅い速度での観察となった. 観察手順 最大水深とした水槽に 5 個体のアユ を収容後,3 日間飼育し,1–2 個体のアユになわ ばりを作らせた.そして,なわばり行動を側面か ら 観 察 し た. な わ ば り 行 動 は, ビ デ オ カ メ ラ (SONY HDR-CX12)で撮影し,記録は,ビデオ編 集 ソ フ ト(COREL VideoStudio12) を 用 い, ビ デ オの再生速度を遅くして,なわばり個体が侵入ア ユを追いかけた水平距離と放した点の垂直距離を モニター画面に物差をあてて測定した.また,水 深との関係について記録した後,約 5 日間は最大 水深で飼育し,行動を随時撮影記録した.さらに, 水槽 1 になわばりが 2 個形成された場合は,なわ ばり個体 2 個体の行動を観察記録した.観察は 2007 年 8–9 月 お よ び,2009 年 6–10 月 に 行 っ た. 実験室の光条件は 14L:10D とし,実験馴致中およ び実験中は無給餌としたが,アユは水槽に付着し た藻類を接餌した.観察は 8:30–13:00 に行った. 結 果 なわばりの形成過程 水槽 1 におけるなわばり 形成までの一連の行動は,次の様であった.アユ を水槽に入れると,5 個体のアユは水槽下流部の 流 速 35–45 cm/S の 所(Fig. 2) に 群 れ を 作 っ た. 次に,最大体長個体が 2 枚のコンクリートブロッ
Fig. 2. Typical ayu territories in experimental tank. Upper: side view; lower: plane view. Solid arrows indicate attack dash of ayu territory holder and neighbor. Dotted arrows indicate foraging routes. In plane view, dotted and dashed lines indicate extent of feeding territories.
クの上やその付近を占拠しだした(なわばり個体 1).続いて,2 番目の個体が最上流部に設置した コンクリートブロックを占拠した(なわばり個体 2).残りの 3 個体は水面近くに群れとなった.こ れらの行動は 24 から 48 時間で終了した.なお, 全個体とも夜間は水槽 1 の底付近に留まっており, 早朝の午前 5 時に照明がつくと,2–3 時間後に最 大体長個体が 2 枚のコンクリートブロックを対象 にしたなわばりを形成することを確認した. なわばりアユの攻撃開始距離 2007,2009 年 のデータ総計 51 回の攻撃行動の記録から,なわ ばり個体が侵入者に示した攻撃ありをプラス,攻 撃なしをマイナスとして各々の頻度を Fig. 3 に示 した.侵入個体は,やや上方から底に向かって侵 入してくるが,なわばり個体に発見され,逃避の ために向きを変え始めた時のなわばり個体との距 離が 40 cm 以内になると攻撃された.また 2 個体 の距離が 40 cm になると攻撃されない場合も観察 された.距離が 45 cm の場合,侵入者が向きを変 え始めると攻撃はされなかった.同一水槽になわ ばり個体が 2 個体出現した場合は,両者の距離が 10 cm で攻撃が起こり,15–25 cm では攻撃のある 場合とない場合が観察された. な わ ば り ア ユ の 追 撃 距 離 2009 年 6 月 26 日 10:00,7 月 3 日 9:00, お よ び 7 月 28 日 9:30 か ら それぞれ 10 分間,追撃行動(attacking dash)につ いて観察し,追撃行動が認められた 4 個体(ここ では,個体名を TO-A から TO-D と定義)の追撃 開始・終了位置を調べた結果を Fig. 4 に示す.7 月 3 日の観察では,追撃行動は,侵入者がなわば り個体に 30–40 cm 近づくと開始され , 約 0.7–1.0 m/S の速度で追いかけることを確認した.この観 察では,なわばりが 2 個形成され,なわばり個体 1(TO-A) は 7 回, 隣のなわばり個体 2(TO-B) は 1 回の攻撃を侵入者に行った. なわばり個体 の攻撃開始と終了位置から形成されたなわばりの 面積は,コンクリートブロックの大きさ(25×25 cm)と水槽の幅(0.5 m)から換算して 1.0 m2と 見積もられた.また,隣のなわばり個体 2 のなわ ばりは 0.4 m2であった.なわばり個体 1 が摂餌 ルートを遊泳する際は,水槽底から 17.5 ± 9.1 cm (平均 ± 標準偏差,n = 60,1 秒ごとの位置)の位 置で波状の遊泳行動を示し,水槽底から 0–5 cm の位置で付着藻類を食んだ. 7 月 28 日の観察では, なわばり個体 1(TO-C)が 1.1 m2のなわばりを作り, 16 回の追撃行動が観察された.なわばり個体 2 は 0.4 m2 のなわばりを水槽の下流コーナー部分に作っ たが,10 分間に攻撃は観察されなかった.なわば り個体 1 の遊泳行動は,水槽底から 15.6 ± 10.2 cm の位置で観察された.最初に観察した 6 月 26 日 の例では,なわばり個体(TO-D)は 0.7 m2のな わばりを水槽ほぼ中央部に作ったが,観察された
Fig. 4. Observed attack responses of four ayu territory holders in experimental tank (Side view 250 ×60 cm).
Fig. 3. Distance between neighboring ayu territory holders (closed bars) and intruders (open bars).