分子動力学計算によるミトコンドリア外膜受容体 Tom20 の プレ配列認識機構の解析
物理学専攻 宗行研究室 修士二年 小室 靖明
研究主題と用いた手法
多様で複雑な生命現象の素過程となる蛋白質と基質一分子の相互作用は,これまでさまざまな実験・理 論的手法により,生物学・化学・物理学の垣根を越え活発に研究されてきた.特に,基質の結合に伴う蛋 白質の構造変化には数多くの報告があり,基質の認識過程を説明するモデルが複数提唱されている.この ように従来は,蛋白質に焦点が当てられてきた一方で,「基質一分子」の構造変化や結合過程についての知 見は,実験・理論の両面とも未だ十分に得られていなく,発展途上にある.そこで,認識過程において基 質一分子のダイナミクスが主役となる,ミトコンドリアへの輸送蛋白質に付加されたプレ配列と,その受 容体蛋白質Tom20との複合体をターゲットに選び(図1),認識過程を分子シミュレーションを用いて解 析した.
図 1 ミトコンドリア外膜受容体Tom20とプレ配列の結晶構造と認識モデル [1, 2]
(上段)3種類の結晶構造.プレ配列(赤,オレンジ,緑),Tom20(グレー),疎水サイト(シアン).(中 段)3種類の結晶構造でそれぞれTom20と結合しているロイシン残基.(下段)“シーソー認識モデル”の模 式図.緑の円柱はαヘリックス構造のプレ配列を表す.赤の残基はロイシン 1,緑の残基はロイシン 4,青 の残基はロイシン 5 を表し,コンセンサス配列の 3 つの疎水性残基に対応する.これらは,Tom20の 2 つの疎水性サイト(青い円)によって認識される:サイト 1 の構成残基(Ile74, Leu106, Leu110),サイ ト 2 の構成残基(Val109, Thr113).この 3 つの結合様式は早い平衡状態にあり,ロイシン 4 は支点の働 きをする.他の結合様式も複数含め,この 3 つの結合様式と交換が早い平衡状態にあると考えられる.
研究成果
図2 レプリカ交換によるプレ配列の主成分空間における自由エネルギー地形とX線結晶構造との比較
(左)プレ配列の主成分空間における自由エネルギー地形.4 つの構造はクラスター分析によって得られ た各クラスターの代表構造.(右上)主成分空間での4つのクラスターの自由エネルギー障壁.(右下)各 クラスターとX線結晶構造とのRMSD値.値が小さいほどずれが小さく,近しい構造を示している.
図3 Tom20とプレ配列の接触原子数と各クラスターと結晶構造の対応
(左)Tom20(疎水基)とプレ配列(ロイシン残基)との距離が4.5 Å以内のペアを接触原子としてカウン トした.(右)図2の結果と合わせたクラスターと結晶構造の対応.
本研究の意義
Tom20-プレ配列複合体の認識機構の提唱 1 基質の素早い結合と解離の必要性
Tom20はミトコンドリア外膜に存在する膜透過装置 TOM40の 1 つのサブユニットである.そのため
Tom20はプレ配列を認識した後,Tom22やTom5を介してTom40が作るチャネルへと輸送蛋白質を引き 渡す必要がある(図 3).このため,Tom20 とプレ配列の結合は適度に弱くなければならない(解離定数 Kd = μMオーダー).もし結合が強すぎると解離が抑制されて前駆体蛋白質の輸送が最初の段階で滞って しまう.すなわち,Tom20とプレ配列の相互作用では,素早い結合と解離の両方が重要であると考えられ る.
図3 ミトコンドリア外膜受容体TOM40複合体中のTom20とプレ配列
2 Tom20-プレ配列相互作用のフラストレーション
プレ配列にはコンセンサス配列として疎水性残基であるロイシンが3 つあり,これがTom20の疎水サ イトと結合する.しかし,3つの結晶構造(A-pose, M-pose, Y-pose)ではプレ配列のロイシンがすべて同 時に結合するのではなく,その一部しか認識されない不完全な結合をしている(図1).これらのことから,
Tom20の疎水サイトはプレ配列の3つのロイシン残基を同時に受け入れるには狭く,フラストレーション
が生じると考えられる.そのためTom20は3つのロイシン残基を同時に受け入れるのではなく,一度に2 つのロイシン残基のみを受け入れることで安定な結合を実現している.そしてプレ配列が熱揺らぎによっ て複数の結合様式を辿ることで,総体として3つの疎水性残基を認識している.
3 基質のダイナミクスが主役となる蛋白質との認識機構
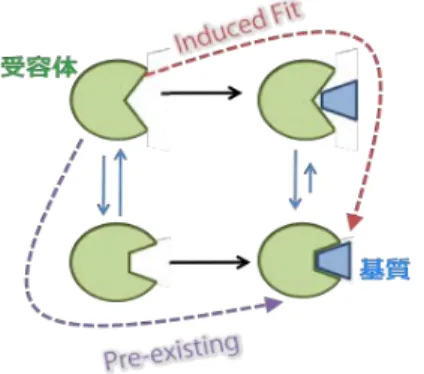
上で述べたように,Tom20による認識ではプレ配列の速い結合と解離の両方が重要である.これは一般 に,薬剤分子が標的蛋白質に強く結合することによって,その蛋白質の機能を阻害または促進する働きと は対照的な相互作用である(図4).
図4 蛋白質と基質の従来の相互作用モデル
赤い経路がinduced-fitモデル,紫の経路がpre-existingモデル.緑が蛋白質を表し,青い台形が基質を表 す.induced-fitでは基質の結合に伴って,蛋白質が形を変え複合体に適した形に変形する.pre-existing モデルでは,複合体に適した構造は熱揺らぎよって基質結合以前に存在し,そこに基質が結合することで
複合体が形成される.
Tom20はこのような認識過程を実現するために,コンセンサス配列の3つの疎水性残基を3つの疎水サイ
トで結合するような強い認識ではなく,部分的に結合する不完全な結合様式の速い交換で達成している.
図5 Tom20と結合した状態でプレ配列が感じる構造的なフラストレーションと本研究によって明らかに
なった結合様式の自由エネルギー障壁
従来,蛋白質と基質の相互作用では蛋白質にその焦点が当てられ,基質の結合に応答する構造変化が論 じられてきた.しかし,本研究で明らかになった基質のダイナミクスが主役となる認識過程は,Tom20と プレ配列に限らず広く生体環境に存在すると思われる.このような認識機構をもつ蛋白質と基質では,多 くの場合でその解離定数が大きく複合体の構造決定が難しい.従って実例が少ないのが現状であるが,標 的膜上の受容体とシグナル配列など,広く一般に適応されうる認識機構であると考えられる.
1. Saitoh T, Igura M, Obita T, et al., The EMBO journal. 2007;26 (22):4777-87.
2. Saitoh T, Igura M, Miyazaki Y, et. al., Biochemistry. 2011;50 (24):5487-5496.
![T andT HybridMeasurementsusingFastSpin-echoSequences 高速スピンエコー法を用いたT ・T 同時計測MRI [論文]](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)