九州大学学術情報リポジトリ
Kyushu University Institutional Repository
Development of allerge-nspecific immunotherapy of Japanese cedar pollinosis using solid-in-oil nanodispersions
孔, 慶靚
http://hdl.handle.net/2324/4060140
出版情報:九州大学, 2019, 博士(工学), 課程博士 バージョン:
権利関係:
Development of allergen-specific immunotherapy of Japanese cedar pollinosis using solid-in-oil nanodispersions
Qingliang Kong
A thesis submitted in partial fulfillment of the requirements for the degree of
Doctor of Engineering
Department of Applied Chemistry Graduate School of Engineering
KYUSHU UNIVERSITY January, 2020
Contents
Chapter 1 Introduction
1. Japanese cedar pollinosis ... 1
1.1 Pollinosis ... 1
1.2 Mechanism of JCP ... 1
1.3 Trigger factors of JCP ... 2
2. Allergen-specific immunotherapy ... 3
2.1 Present state of AIT ... 4
2.2 Mechanism of AIT ... 4
2.3 Current AIT for JCP ... 5
2.4 Challenges of AIT ... 6
2.5 Novel antigens developed for AIT for JCP ... 7
2.5.1 T cell epitope peptides ... 9
2.5.2 Modified allergens ... 9
2.5.3 DNA vaccines ... 9
2.5.4 Adjuvant formulations ... 10
2.6 Novel administration routes developed for AIT for JCP ... 10
2.6.1 Oral immunotherapy ... 11
2.6.2 Transcutaneous immunotherapy/Epicutaneous immunotherapy ... 12
2.6.3 Intralymphatic immunotherapy ... 12
2.6.4 Intranasal immunotherapy ... 12
3. Nanoparticles developed in AIT for JCP ... 13
3.1 Oligomannose-coated liposomes ... 13
3.2 S/O nanodispersions ... 14
3.3 Applications of S/O nanodispersions ... 15
4. Objectives and outline of the thesis ... 18
Chapter 2 Transcutaneous immunotherapy of pollinosis using solid-in-oil nanodispersions loaded with T cell epitope peptides
1 Introduction ... 27
1.1 Transcutaneous immunotherapy ... 27
1.1.1 Mechanism of TCIT ... 27
1.1.2 Challenge of TCIT ... 28
1.1.3 Strategies developed for TCIT ... 29
1.2 T cell epitope peptide ... 30
1.2.1 Single T cell epitope peptide ... 31
1.2.2 Hybrid T cell epitope peptide ... 31
1.2.3 T cell epitope peptide in present study ... 33
1.3 Objective of this Chapter ... 33
2 Materials and Methods ... 34
2.1 Materials ... 34
2.2 Preparation of S/O nanodispersions containing T cell epitope peptides ... 34
2.3 In vitro skin permeation study ... 35
2.4 Histological observation of YMP back skin treated with S/O nanodispersions... 35
2.5 Permeation pathway study ... 35
2.6 Sensitization and immunotherapy ... 36
2.7 Measurement of serum antibody levels ... 36
2.8 Clinical symptoms of pollen allergy ... 37
2.9 Cytokine assay ... 37
2.10 Statistical analysis ... 37
3 Results and Discussion ... 37
3.1 Characterization of the S/O nanodispersions loaded with T cell epitope peptides ... 37
3.2 In vitro skin permeation study ... 38
3.3 In vivo skin permeation study ... 40
4 Conclusion ... 44
Chapter 3 Transcutaneous delivery of immunomodulating pollen extract- galactomannan conjugate by solid-in-oil nanodispersions for pollinosis
immunotherapy
1. Introduction ... 49
1.1 Allergen modification strategy ... 49
1.2 Pollen allergen-galactomannan conjugate ... 50
1.3 Objective of this Chapter ... 51
2. Materials and Methods ... 51
2.1 Materials ... 51
2.2 Animals ... 52
2.3 Preparation of S/O nanodispersions ... 52
2.4 In vitro analysis of the antigen release ... 52
2.5 In vitro analysis of the skin permeation of the antigen... 53
2.6 In vivo analysis of the skin permeation of antigen ... 53
2.7 Antigen uptake by DC2.4 cells ... 53
2.8 Confocal imaging of DC2.4 cells... 54
2.9 Sensitization ... 54
2.10 TCIT treatment of the PE and PE-GM ... 54
2.11 TCIT treatment of the pollinosis mouse model using PE-GM ... 55
2.12 Measurement of the serum antibody levels ... 55
2.13 Cytokine measurement ... 55
2.14 Effect on pollen-induced nose rubbing ... 56
2.15 Statistical tests ... 56
3. Results and Discussion ... 56
3.1 Preparation and characterization of S/O nanodispersions carrying PE or PE-GM ... 56
3.2 Skin permeation by S/O nanodispersions carrying PE or PE-GM ... 57
3.3 Antigen uptake by DC2.4 cells ... 59
3.4 Comparison of TCIT using S/O nanodispersions carrying PE or PE-GM ... 60
3.5 Comparison of the differences between TCIT and SCIT using PE-GM ... 62
4 Conclusion ... 65
Chapter 4 Solid-in-oil nanodispersions for intranasal vaccination:
Enhancement of mucosal and systemic immune responses
1. Introduction ... 69
1.1 Intranasal vaccination... 69
1.1.1 Nasal immune system ... 70
1.1.2 Marketed intranasal vaccine ... 70
1.2 Nanoparticles for intranasal immunization... 71
1.2.1 Chitosan based nanoparticles ... 71
1.2.2 Polymer based nanoparticles ... 72
1.2.3 Lipid based nanoparticles ... 72
1.2.4 Emulsion ... 73
1.3 Objective of this Chapter ... 73
2 Materials and Methods ... 74
2.1 Materials ... 74
2.2 Preparation of S/O nanodispersions ... 74
2.3 Drug release test ... 75
2.4 In vivo fluorescence imaging ... 75
2.5 In vivo nasal absorption ... 75
2.6 Murine intranasal immunization ... 76
2.7 Measurement of serum and mucosal wash antibody levels ... 76
2.8 Histopathology... 76
2.9 Statistical tests ... 77
3 Results and Discussion ... 77
3.1 Characterization of the S/O nanodispersions in the different oils ... 77
3.2 Nasal absorption of antigen ... 80
3.3 Intranasal immunization and antibody evaluation ... 81
3.4 Histopathology... 83
4 Conclusion ... 84
Chapter 5 Conclusions 1. Summary ... 89
2. Future prospective ... 90
Chapter 1 Introduction
1. Japanese cedar pollinosis
Allergic rhinitis (AR) is an allergic response to specific allergens. It has already become a global public health issue in industrialized countries. Although it is not a serious illness, the allergic symptoms including a runny or stuffy nose, sneezing, itchy, and watery eyes, greatly affect the quality of life in persons of all ages, such as work productivity, learning and sleeping.
Moreover, several studies reported that AR often precedes the bronchial asthma [1]. The allergens contain pollen, dust, latex, animal dander even certain food and plant. Among them, pollen is the most common allergen responsible for the development of AR in Japan.
1.1 Pollinosis
Pollen, derived from tree, grass and weed, is the major allergen of AR named pollinosis or hay fever in the world [2]. It differs regionally: grass-induced pollinosis is the most common pollen allergy in Europe; birch pollen is a major source of pollinosis in Northern Europe and North America; cypress-induced pollinosis is reported worldwide; Japanese cedar (Cryptomeria japonica, JC) induced pollinosis is the most popular AR in Japan.
JC pollinosis (JCP), is a type I hypersensitivity, which occurs when the immune system overreacts to cedar pollen (Fig. 1-1). More
than 26.5% of people in Japan suffers from it caused by repetitive exposure to the cedar pollen, therefore, it was considered as a part of Japanese national disease [3]. The prevalence of it has increased more than other ARs, and still continues to increase.
During the cedar pollen season between February and April up to 3 months, many patients experience severe allergic symptoms. Therefore, development of an effective treatment for JCP is necessary.
1.2 Mechanism of JCP
JCP is a typical type I hypersensitivity, which is a classical Immunoglobulin-E (IgE)- Fig. 1-1 The blooming Japanese cedar tree [3].
Reprinted from J. Allergy Clin. Immunol. Vol. 133, Takechiyo et al., Present state of Japanese cedar pollinosis: The national affliction, 632-639, Copyright 2014, with permission from Elsevier.
mediated disease. The development of JCP occurs in two steps, sensitization to cedar pollen and development of immediate and late symptoms [2,4–9]. Sensitization is the first step of JCP.
Initial contact with tiny amounts of allergen (cedar pollen) at mucosal surfaces may favor allergen uptake by antigen presenting cells (APCs). The APCs then display the allergen fragments on their cell surfaces to induce the differentiation and/or activation of T helper cells type 2 (Th2) from the native T cells (Th0) in regional lymph nodes. Then pro-inflammatory cytokines such as interleukin-4 (IL-4) and IL-13 are produced to activate B cells secreting antibodies IgE specific for the allergen. T cell activation in the presence of IL-4 increases differentiation into Th2 cells from Th0 cells. As a result, JCP patients produce cedar pollen allergen-specific IgE antibodies (Fig. 1-2A). IgE antibodies attach to the IgE receptor FcɛRI (high-affinity receptor for IgE) on the surface of effector cells, such as mast cells and basophils. Subsequent repeated allergen contact causes the crosslinking of effector-cell-bound IgE, trigger the degranulation of these cells and the release of active mediators, such as histamine and leukotrienes (Fig. 1-2B). Th2 cytokine IL-5 induces tissue eosinophilia and the release of inflammatory mediators from eosinophils. Through these procedures, the allergic symptoms of allergy are induced. Moreover, sensitization induces the production of IgE memory B cells. When the allergen invades body again, IgE memory B cells are boosted after the allergen contact to produce allergen-specific IgE antibodies.
1.3 Trigger factors of JCP
Environmental and genetic factors have been known to influence development of JCP.
Japanese cedar trees planted and covered nearly 18% of the total land area of Japan, according to the Forestry Agency of Japan [3]. Although pollen counts vary significantly from year to year due to the weather conditions change, however, the total pollen counts from 1995 to 2013 have been significantly increased compared to those in the period from 1965 to 1994 [3], related to the prevalence of JCP increased recent years. In addition, overlapping genetic factors influence the development of JCP in the study of familial clustering and intra-individual clustering [10].
Fig. 1-2 Mechanism of Japanese cedar pollinosis. Suppressive activities and inductive activities are indicated by red and grey arrows, respectively. APC, antigen presenting cell;
FcɛRI, IgE receptor; IFN, interferon; IL, interleukin; Th0, naïve T; Th1, T helper type 1; Th2, T helper type 2.
2. Allergen-specific immunotherapy
Allergen avoidance, pharmacotherapy and allergen-specific immunotherapy (AIT) are the main options for JCP patients. However, complete avoidance for JC pollen is impossible by wearing a mask and glasses. Most patients received the pharmacotherapy including oral or intranasal administrations of antihistamines and intranasal administrations of corticosteroid aerosol to relieve the allergic symptoms. Different from pharmacotherapy, AIT is the only
curative treatment for JCP.
2.1 Present state of AIT
AIT can change the natural course of type 1 allergic disease, therefore, it is considered as the only curative intervention. The repeated and increased doses of allergens are administrated to patients, aiming to induce the antigen-specific immune tolerance and reduce the reliance on pharmacotherapy. The first study of AIT for hay fever was conducted in 1911 [11]. With the development of the technology, the efficiency and safety of AIT for allergic diseases have improved. Currently there are two major forms of AIT for JCP used in the clinical practice, consisting of the subcutaneous immunotherapy (SCIT) and the sublingual immunotherapy (SLIT) treated with the cedar pollen extract (PE) allergen.
2.2 Mechanism of AIT
The mechanism of AIT for treating JCP is still unclear due to the complexity of pollen allergen, however, extensive clinical and preclinical studies give a big picture for the mechanism of AIT [2,5–9]. A shift from allergic Th2 immunity to Th1 immunity and a generation of regulatory T (Treg) cells are considered as the major reason for the suppression of Th2 immunity (Fig. 1-3). The levels of cytokine IL-4, IL-5 and IL-13 are decreased, leading to the decreases of effector cells (mast cells, basophils and eosinophils). IgG1, IgG4, and IgA antibodies are increased after AIT under the influence of interferon-γ (IFN-γ), IL-10, and transforming growth factor-β (TGF-β). These changes of antibodies and cytokines inhibit IgE binding and competitively inhibit allergen binding and IgE complex formation.
Fig.1-3 Mechanisms of allergen-specific immunotherapy. Suppressive activities and inductive activities are indicated by red and grey arrows, respectively. Breg, regulatory B;
APC, antigen presenting cell; IFN, interferon; IL, interleukin; Th0, naïve T; Th1, T helper type 1; Th2, T helper type 2; Treg, regulatory T.
The parameters for the evaluation to AIT are introduced in brief (Table 1-1) [6,8]. Serum antibody IgE, which released following the exposure to allergen and causing the degranulation of effector cells, is the most relevant parameter in AIT. The decreases in total IgE and allergen- specific IgE levels are reported in almost all the researches of AIT. The competition ability of antibody IgG, IgG4 and IgA for IgE is well-known, which can block allergen-IgE complex formation. The allergen-specific IgG1 and IgG2a are used to evaluate the balance of Th2/Th1 immunity. Th1, Th2 and Treg type cytokines are used to evaluate Th1, Th2 immunity and Treg cells, respectively. Allergen-specific Treg cells are reported to induce the immune tolerance during AIT, and regulatory B (Breg) cells have the capacity to inhibit T cell-mediated inflammatory responses and induce allergen-specific IgG and IgA antibodies.
Table 1-1 Summary of the parameters for the evaluation to AIT
Domains of biomarker Parameters
Antibodies IgE (total IgE and allergen-specific IgE), IgG (IgG1, IgG2a and IgG4)
IgA
Cytokines and chemokines Th2 type: IL-4, IL-5, IL-9, IL-13, IL-19, IL-21 Th1 type: IFN-γ, IL-12
Treg type: IL-10, TGF-β
Cellular parameters Treg cells, Breg cells
2.3 Current AIT for JCP
JC PE is commonly used to treat JCP as the antigen. Cry j is a registered allergen derived from the genus Cryptomeria and the species japonica [11]. Two major allergens of JC PE, Cry j 1 and Cry j 2, have been isolated and characterized [12]. Cry j 1 (molecular weight (MW) of 45-51 kDa), as the first allergen isolated from JC PE, is a basic glycoprotein homologous of pectate lyase protein. Cry j 2 (MW of 37-45 kDa), the second allergen isolated from JC PE, is a protein homologous of polygalacturonase. The detail of Cry j 1 and Cry j 2 has been introduced in the previous literature [11]. New allergens from JC PE have been reported, such as Cry j 3 and Cry j 4, however, allergens Cry j 1 and Cry j 2 still affect more than 90% of JCP patients [13].
SCIT treated with allergen JC PE, as the most traditional form of AIT, has been introduced in the clinical treatments for over 100 years [14]. A crude PE has been available since 1969 and the standard PE for SCIT became available in 2000. To induce the long-term tolerance, SCIT is commonly taken up to 3 to 5 years from a low dose of PE in an initial up-dosing phase and gradually increased until a high dose in a maintenance phase (Fig. 1-4). In the initial phase,
increasing does are treated to build the tolerance and evaluate the sensitivity of the patients to doses. Then the maximum doses tested are administrated in the maintenance phase. SCIT is more effective than pharmacotherapy in clinical trials and real-life studies.
Fig.1-4 Current allergen-specific immunotherapy used in real-life studies.
SLIT is introduced as a safe and simple alternative to SCIT, because of the convenient home- based therapy and no injections. The World Health Organization (WHO) position paper mentioned SLIT as a possible alternative to SCIT in 1998 [1]. The SLIT treatment for JCP has already been approved by regulatory authorities in Japan since 2014. A sublingual drop named
“Cedartolen®” and a sublingual tablet named “Cedarcure®” derived from PE (Torii Pharmaceutical Co., Ltd., Tokyo) were approved and became available on the market in 2014 and 2018, respectively (Fig. 1-5). Efficacy and safety of SLIT for JCP patients is summarized in this literature [1]. Similar with SCIT, SLIT is also commonly taken up to 3 to 5 years.
Fig.1-5 Sublingual drop named “Cedartolen®” and tablet named “Cedarcure®” (https://www.alg-immunotherapy.jp/index.php).
2.4 Challenges of current AIT
SCIT and SLIT of JC PE are effective for the treatment of JCP, however, low compliance
and persistence for patients were reported in clinical trials and real-life studies [15]. The allergen PE and the administration routes are considered as the major reasons.
Although whole JC PE is used as the antigen molecule in SCIT and SLIT, there are still some concerns about its safety [7]. The complexity of whole PE has a risk of side effects caused by its cross-linking ability with allergen-specific IgE antibodies bound to the surface of effector cells. SCIT was reported the severe allergic side effects even death after injection of JC PE.
Therefore, a novel antigen, which can avoid such side effects of allergens and improve the efficiency to shorten the duration of AIT, is required.
Another challenge is related to the administration route. Patients received SCIT need to endure the pain from the weekly injection in the long therapeutic period. Although no severe allergic side effects caused by SLIT were reported, some side effects including itching and swelling of the mouth and tongue, headache, and ear pruritus were reported [1]. These side effects may cause dose limitations and the treatment duration up to 3–5 years. The cost of a long therapeutic period may also limit the broad applicability of SCIT and SLIT. Therefore, a safe, simple and cost-less administration route of AIT for JCP, is required.
2.5 Novel antigens developed for AIT of JCP
Through the understanding of AIT, the most important aspects of effective AIT for JCP are avoiding side effects of the antigen and improving the vaccine efficiency to shorten the duration of AIT. To realize these, researchers focus on the crucial aspect of AIT: the antigens. The safe antigen to avoid cross-linking with an allergen-specific IgE antibody is necessary. Moreover, modifying the allergen, such as targeting of immune cells or enhancing the uptake of antigens by immune cells by the attachment of polysaccharides or polymer to allergens, or suppressing Th2 immunity by the attachment of adjuvant to allergens, is expected to shorten the duration of AIT.
Several antigens have been developed in AIT for JCP, including T cell epitope peptides, modified allergens, DNA vaccines and adjuvant formulations, are summarized in Table 1-2.
Most researches are still at the preclinic stage; however, oral immunotherapy (OIT) of transgenic rice seeds has been at the clinical phase I stage [16], and OIT of Cry j 1- galactomannan conjugate [17–20] and intramuscular immunotherapy of LAMP-based DNA vaccine have been at the clinical phase II stage [21,22].
Table 1-2. Novel antigens used for the immunotherapy of Japanese cedar pollinosis
Antigen Allergen Route Mechanism References
T cell epitope
peptide p246-259 of Cry j 2 Oral Suppression of Th1 and
Th2 responses [23,24]
Integrated peptide from three
epitopes Oral
Inhibition of lymph node cell proliferation to Cry j
1/2 [25]
Cry-consensus from five or
six epitopes SC Induction of IgG2a
Th1 deviation [26,27]
Rice seed containing peptides
from fourteenth epitopes Oral - [28]
Rice seed containing 7Crp
from seven epitopes Oral Inhibition of T-cell proliferative response to
Cry j 1 [29]
Chicken egg containing 7Crp
from seven epitopes Oral Suppression of Th2
responses [30]
7Crp from seven epitopes SL Induction of IL-10–
producing Tr1 cells [31]
7CrpR from seven epitopes T - [32]
Seven epitope mixture T Suppression of Th1 and
Th2 responses [33]
Modified allergens
Cry j 1-glactomannan
conjugate Oral Induction of IgG4 and IL-
10 [17–20]
PE-galactomannan conjugate T Suppression of Th2 responses
Th1 deviation [34]
Polyethylene glycol modified
Cry j 1/2 SC
Induction of IgG and Th1 responses
Suppression of Th2 responses
[35]
Rice seed containing fusion
protein from Cry j 1/2 Oral Suppression of Th1 and
Th2 responses [36,37]
DNA vaccine Vaccine encoded Cry j 1 gene IM Induction of Th1
responses [38]
Vaccine encoded T cell epitope peptide (p247-258)
from Cry j 2 IM Induction of Th1
responses [39]
Cry j 1-LAMP and Cry j 2-
LAMP IM Induction of IgG
Th1 deviation [21,22]
Adjuvant formulation
Cry j 2 T cell epitope peptide-
CpG conjugate SC
Suppression of IL-4 and IL-5
Suppression of Th2 responses
[40]
Cry j 1-CpG conjugate SC Induction of IgG2a
Th1 deviation [41]
Recombinant protein concluding T cell epitope
peptide and cholera toxin B - - [42]
Oligomannose-coated
liposomes carrying Cry j 1 ID
Induction of IFN-γ and suppression of IL-4/IFN-γ and IL-5/IFN-γ ratios Th1 deviation
[43]
7CrpR from seven epitopes
combined with CpG T Th1 deviation [44]
ID, intradermal; IM, intramuscular; SC, subcutaneous; SL, sublingual; T, transcutaneous.
2.5.1 T cell epitope peptides
The use of T cell epitope peptides derived from pollen allergens was proposed to reduce the side effects of whole PE [45]. T cell epitope peptides are short antigen-derived molecules recognized by T cells. Because they do not cross-link with allergen-specific IgE bound to the surface of mast cells and basophils, treatment using T cell epitope peptides do not pose a potential risk of side effects. Another advantage is considered that the ratio of epitope peptides could be adjusted for an individual therapy in the future. Several dominant T cell determinants have already identified as major T cell epitopes [26,46]. The applications of T cell epitope peptides in AIT of JCP will be discussed in Chapter 2.
2.5.2 Modified allergens
Although AIT using T cell epitope peptides seems to be safe and effective, it is not applicable to all JCP patients. Because different genetic backgrounds in humans respond differently to different T cell epitope peptides, genetic differences in the T cell receptors cause an insufficient of T cell epitopes in some JCP patients [47]. The modified allergens including whole T cell epitopes information are effective to all JCP patients. The applications of modified allergens in AIT of JCP will be discussed in Chapter 3.
2.5.3 DNA vaccines
DNA vaccine is a novel approach for the prevention and treatment of JCP. The antigen gene encoded by bacterial plasmid vectors induces the long-lasting expression of a specific antigen, causing a long-term persistence of the immune response [48]. Unlike T cell epitope peptides and modified allergens, however, DNA vaccines have not been studied extensively may be due to its complexity. Toda et al. developed a DNA vaccine that encoded the Cry j 1 gene. The IgE antibody level induced by allergen Cry j 1 was suppressed in mice treated intramuscular injection with the DNA vaccine, because intramuscular administration of DNA vaccine effectively induced the antigen-specific Th1 type responses [38]. In a later report, Toda et al.
designed a new DNA vaccine that encoded a gene of T cell epitope peptide (p247-258) from Cry j 2. Anti-Cry j 2 IgE level was reduced after administration of this DNA vaccine, but IgG2a responses were enhanced markedly, suggesting the potential to alleviate JCP allergic reactions [39]. Recently, Su et al. (Immunomic Therapeutics, Inc. USA) found that a fusion of lysosomal- associated membrane protein-1 (LAMP-1) with an antigen in DNA plasmids could enhance antigen immunogenicity. Su et al. developed two LAMP-based DNA vaccines to treat JCP, Cry j 1-LAMP and Cry j 2-LAMP, that encoded the allergen Cry j 1 and Cry j 2, respectively [21].
The reduced titers of IgG1 and IgE, and improved titers of IgG2a were observed in the Cry j 1- LAMP and Cry j 2-LAMP plasmids immunized mice, suggesting that Th1 response was skewed from the Th2 allergic response. Safety and long-term immune responses of Cry J 2-LAMP DNA vaccine for JCP were evaluated in a later report [22]. This Phase IA and IB clinical trial suggested that the Cry J 2-LAMP DNA vaccine is safe and immunologically effective in JCP treatments. Astellas Pharma Inc. now is leading a Phase II clinical studies of Cry j-LAMP DNA vaccine (ASP-4070) in Japan.
2.5.4 Adjuvant formulations
JCP is a typical Th2 induced allergy, therefore, formulation or conjugation of a Th1- enhancement or T-cell tolerance adjuvant with the allergen is a strategy to improve the efficiency of AIT. CpG oligodeoxynucleotide (ODN) is an adjuvant for the Th1 immune response, therefore, it is expected to be useful in the development of AIT for JCP [48]. Suzuki et al. firstly conjugated the CpG to the Cry j 2 T cell epitope peptide [40]. Cry j 2-specific IgE, IL-4, and IL-5 were decreased significantly in mice administrated with CpG-Cry j 2 peptide conjugate compared to other groups. Moreover, the CpG-Cry j 2 peptide conjugate could significantly attenuate nasal symptoms compared with a physical mixture of CpG and Cry j 2 peptide. In a later report, allergen Cry j 1 was conjugated with CpG. It showed a decrease of anti-Cry j 1 IgE response and increase of anti-Cry j 1 IgG2a in a JCP mouse model, suggesting that CpG switched on Th1 immunity through the upregulation of IgG2a [41]. Another adjuvant, cholera toxin B subunit, is used as a carrier molecule to induce mucosal immune responses and T-cell tolerance to antigens. Hoang et al. fused cholera toxin B subunit with the T cell epitope peptides from Cry j 1 and Cry j 2 to become a recombinant protein successfully [42]. The recombinant protein was antigenic against anti-Cry j 1 and anti-Cry j 2 antibodies, however, the adjuvant effect of cholera toxin B was not discussed in this study. To the best of our knowledge, no adjuvant formulations are used in the clinical trials of AIT for JCP, may be due to a concern of the safety of the adjuvant.
2.6 Novel administration routes developed for AIT of JCP
As the alternative methods of SCIT and SLIT, several administration routes developed for the AIT of JCP including OIT and transcutaneous immunotherapy (TCIT)/ epicutaneous immunotherapy (EPIT) were investigated [5,49]. Moreover, intralymphatic immunotherapy (ILIT) and intranasal immunotherapy (INIT) are succeed in the AIT of AR caused by grass pollen and dermatophagoides. Although ILIT and INIT are not studied in AIT for JCP, we present them here due to the similarity in AR caused by different allergens. Table 1-3
summarized the advantages, disadvantages, mechanisms and represent researches in the different administration routes.
Table 1-3 Different administration routes of AIT
Administration
routes Advantages Disadvantages Mechanisms Represent research
OIT Possible of
relatively large amount of allergen
Antigen
degradation by enzymatic
degradation
Targeting immune cells in the gut associated lymphoid tissue
Administration of Galactomannan-Cry j 1 conjugate [17–20], transgenic rice [28,50,51],
transgenic egg [30]
and T cell epitope peptide [23,24] for AIT of JCP
TCIT/EPIT
1. Needle-free, non-invasive and self-administration
2. High
immunological functions of skin 3. Avoidance of side effects
Barrier of stratum corneum
against extraneous molecules
Targeting antigen presenting cells in the skin
AIT of milk allergy [52], peanut allergy [52,53], grass pollen allergy [54–57] and JCP [32–34,44].
ILIT
1. Smaller allergen does and less
numbers of
injection
2. Avoidance of side effects
Requirement for ultrasound-guided administration
Direct
administration of allergen into lymph nodes
AIT of grass pollen allergy [58–61]
INIT Needle-free, non-
invasive and self- administration
Adverse local nasal side effects
Targeting immune cells in nasal-associated lymphoid tissues
AIT of
dermatophagoides- induced allergic rhinitis [62,63]
AIT, allergen-specific immunotherapy; EPIT, epicutaneous immunotherapy; ILIT, intralymphatic immunotherapy; INIT, intranasal immunotherapy; OIT, oral immunotherapy; TCIT, transcutaneous immunotherapy.
2.6.1 Oral immunotherapy
OIT is commonly used for treating food allergies, however, several research of OIT for JCP have been studied. Oral administration allows relatively large amount of allergen deliver to immune cells in the gut associated lymphoid tissue, aiming to induce the effective immune tolerance. The represent researches of OIT for JCP are oral administration of galactomannan-
Cry j 1 conjugate [17–20], transgenic rice [28,50,51], transgenic egg [30] and T cell epitope peptide [23,24]. The antigen degradation by enzymatic reactions is considered as the disadvantage of OIT.
2.6.2 Transcutaneous immunotherapy/Epicutaneous immunotherapy
TCIT/EPIT attracts much attention not only as an alternative method of SCIT and SLIT to improve the compliance and persistence, but also due to their immunological functions of the skin. Abundant APCs including Langerhans cells (LCs) and dendritic cells (DCs) are reported in the skin, and they can induce an effective immune response. By targeting LCs and DCs, TCIT/EPIT can reduce both local and systemic adverse effects. The background of TCIT/EPIT and technologies developed for TCIT will be introduced in Chapter 2. Clinical efficacy and safety of EPIT for a food allergy (e.g. milk and peanut allergy) were investigated in several clinical trials [52,64]. The efficacy of EPIT in tree-pollen Bet v 1 induced allergic asthma and the efficacy of TCIT in grass induced allergic rhinoconjunctivitis were investigated [54–57].
However, few research studies focused on the TCIT of JCP. Therefore, we focus on the TCIT of JCP in Chapter 2 and 3.
2.6.3 Intralymphatic immunotherapy
ILIT is developed as an alternative of SCIT. Allergen is directly administrated into lymph nodes to increase the efficacy of antigen presenting, enhance the immunogenicity of antigen, and thus achieve the benefits of AIT rapidly. Due to its high efficacy, ILIT by administrating smaller allergen does and less numbers of injection could achieve beatifical results. Moreover, ILIT also reduces the side effects of AIT because of the less exposure of allergens to the effector cells. It has been proven as a safe and effective AIT for grass pollen allergy [58–61]. However, ultrasound-guided administration is necessary for ILIT and makes implement in practice difficultly. Moreover, patients may be difficult to accept this unique administration route.
2.6.4 Intranasal immunotherapy
INIT has recently emerged, because it is needle-free, non-invasive and could be used for AIT without the need for a medical professional. The nasal mucosa is easily accessible and highly vascularized, and has abundant immune cells with the potential to induce achieve the benefits of AIT. In addition, INIT is reportedly able to induce both systemic and mucosal immune responses. The efficacy of INIT for dermatophagoides-induced AR was investigated [62,63]. However, the study of INIT used for AIT of JCP was not reported to date. Therefore, the potential of S/O nanodispersions in INIT will be discussed in Chapter 4.
3. Nanoparticles developed in AIT for JCP
Besides the antigens and the administration routes developed for AIT of JCP, a variety of particulate delivery systems have also been developed. Controlled release of antigen can be achieved by nanoparticles (NPs) for avoiding the side effects caused by the burst release of an antigen. The particulate nature decides its immunopotentiating effect through facilitating uptake and transport of antigens to lymphoid organs. In addition, the modification of NPs surface by carbohydrates or adjuvants can also target APCs as well as modulate the immune response.
Chitosan NPs, polymer NPs and virus-like NPs are employed for AIT and summarized in the literature [65,66].
Poly(D,L-lactic-co-glycolic) acid (PLGA) NPs/microspheres are the represent vehicles and have been developed for the treatment of birch pollen induced pollinosis by OIT and SLIT [67–
70]. PLGA particles triggered Th1 immune reposes in mice [71], therefore, they are expected to be useful in the treatment of type 1 allergic diseases. The immunomodulatory effect of targeting enterocytes or M cells was realized through birch PE encapsulated PLGA NPs modification with wheat germ agglutinin [67] and Aleuria alantia lectin [68], respectively.
However, few researches studied the effect of NPs in the AIT of JCP, apart from the oligomannose-coated liposomes (OMLs) and solid-in-oil (S/O) nanodispersions. Therefore, OMLs system developed in AIT of JCP is presented firstly. Then, S/O nanodispersions system is introduced as another antigen delivery system. Finally, the applications of S/O nanodispersions in AIT are presented, especially for AIT for JCP.
3.1 Oligomannose-coated liposomes
OMLs are prepared from the liposomes coated with neoglycolipids constructed from mannotriose and dipalmitoylphosphatidylethanolamine (Man3-DPPE). After administration of OMLs, they are taken up rapidly by macrophages and accumulated in lymphoid tissues [72].
Moreover, a tumor immunity study demonstrated that OMLs induced a robust Th1 immune response, with a significant increase in IFN-γ production and a decrease in cytokine IL-4 production [72]. Ishii et al. evaluated the anti-allergic effect of JCP with OMLs carrying Cry j 1 and found that OMLs loaded with Cry j 1 inhibit the IgE antibody level and the ratio of IL- 5/IFN-γ [43]. This inhibition might be caused by a shift from a Th2 immune response to a Th1 immune response. Therefore, OMLs carrying Cry j 1 have a potential as the immunotherapeutic agents to treat JCP. However, only total IgE antibody levels and the ratio of IL-5/IFN-γ were evaluated in allergen Cry j 1-senstized mice, the most important parameter of AIT in JCP, allergen Cry j 1-specific IgE was not evaluated in present study. Moreover, the detail of other
Th1 and Th2 cytokines were not discussed expect IL-5 and IFN-γ cytokines.
3.2 S/O nanodispersions
S/O nanodispersions are oil-based dispersions of nano-sized particles with an antigen formed by coating it with hydrophobic surfactant molecules. The antigen-surfactant complex, derived from the removal of water and organic solvent from water-in-oil (W/O) emulsions by lyophilization, is dispersed in an oil vehicle to form S/O nanodispersions (Fig. 1-6) [73].
Compared with conventional W/O emulsion, S/O nanodispersions showed a high antigen encapsulation efficiency up to 99.5% and high stability for more than 3 months [73,74].
The main applications of S/O nanodispersions for AIT are TCIT. Aided by the hydrophobic surfactant and the oil vehicle, S/O nanodispersions can overcome the barrier of SC and deliver the antigen into the skin. The main mechanism for S/O nanodispersions to permeate the skin was discussed by using confocal microscopic observation and computational dynamics simulation, suggesting that dissociation of surfactants happened after S/O nanodispersions contracted the lipid membrane in SC, causing hydrophobic surfactants removed from the antigens in SC and only hydrophilic antigens can permeate into epidermis and dermis [75–78].
Fig.1-6 Preparation and application methods of S/O nanodispersions [73].
3.3 Applications of S/O nanodispersions Tahara et al. firstly reported the transcutaneous protein delivery using S/O nanodispersions carrying fluorescein isothiocyanate (FITC)-labeled insulin (Mw 6 kDa), enhanced green fluorescent protein (EGFP, Mw 27 kDa) and horseradish peroxidase (HRP; Mw 40 kDa) [75]. The permeation of all proteins into the skin was increased by using S/O nanodispersions compared with the aqueous solution (Fig. 1- 7). The penetrated EGFP and HRP exhibited green fluorescence and catalytic activity, respectively, suggesting that these proteins can permeate into the skin in a functional form. Later, Tahara et al.
developed a transcutaneous immunization method by using S/O nanodispersions carrying ovalbumin (OVA) [79]. A high
level of OVA-specific IgG was observed in the mouse treated with S/O nanodispersions. This result showed the potential of S/O nanodispersions used in TCIT.
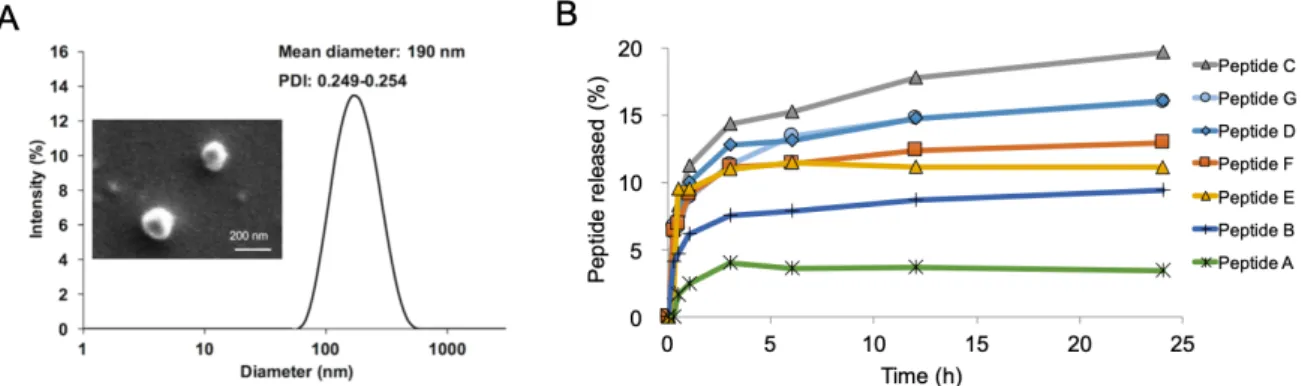
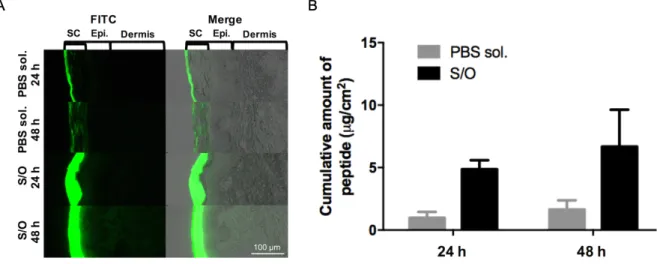
Kitaoka et al. firstly investigated the potential of TCIT for JCP by using S/O nanodispersions carrying the hybrid peptide [32]. To improve the poor solubility of 7Crp, small linkers (triarginine) were introduced between the epitopes in 7Crp (Fig. 1-8A). This new hybrid peptide named 7CrpR was produced by an Escherichia coli system. The spherical nanoparticle of S/O loaded with 7CrpR was observed in microscope images and a mean diameter of 230 nm was measured. In a skin permeation study, fluorescence Cy3-labeled 7CrpR was observed in the SC layer and even epidermis of mouse auricle treated with S/O nanodispersions. However, 7CrpR was observed only on the topmost layer of SC of mouse auricle after treated with a PBS solution.
Pollinosis mouse models were prepared, then treated with 7CrpR dispersed in S/O nanodispersions or the PBS solution. Antibody IgE levels are suppressed in mice following S/O nanodispersions carrying 7CrpR compared to that of PBS groups (Fig. 1-8B, C), especially a significant decrease in allergen Cry j 1-specific IgE level was observed. These results suggested the potential of TCIT for JCP by using S/O nanodispersions carrying 7CrpR.
Fig.1-7 Fluorescence microscopy of the pig skin sections treated with S/O nanodispersions and aqueous solution containing FITC-labeled insulin observed through a 10× objective (scale bar: 200 µm) [75]. Reprinted from J. Control. Release Vol. 131, Tahara et al., A solid-in-oil nanodispersion for transcutaneous protein delivery, 14-18, Copyright 2008, with permission from Elsevier.
Fig.1-8 (A) Amino acid sequences of 7CrpR. (B) Serum total IgE and (C) Cry j 1-specific IgE levels in the mice after treatment of S/O nanodispersions loaded with 7crpR [32].
Reprinted from AAPS PharmSciTech Vol. 16, Kitaoka et al., Transcutaneous Peptide Immunotherapy of Japanese Cedar Pollinosis Using Solid-in-Oil Nanodispersion Technology, 1418–1424, Copyright 2015, with permission from Springer Nature.
The addition of Th1 promoting adjuvant, which switches naïve T cell immunity to Th1 immunity for suppressing Th2 immunity, was introduced into S/O nanodispersions system.
Kitaoka et al. prepared S/O nanodispersions loaded with OVA and CpG ODN 1585 [80]. CpG- ODN is a short synthetic DNA containing unmethylated CpG motifs, which can be recognized by Toll-like receptor 9 to induce a Th1 immunity. The mean IgG1/IgG2a ratio was decreased by co-encapsulation of CpG ODN 1585 into S/O nanodispersions, suggesting the immune balance was skewed from Th2 immunity to Th1 immunity. After investigated the efficiency of Th1 promoting adjuvant in immunity, Kitaoka et al. evaluated the efficiency of another adjuvant R848 in TCIT for JCP [44]. R848 (resiquimod), which is an imidazoquinoline-like molecule that interacts with TLR7/8 receptors, was added into the oil vehicle (R848out) or inside the nanoparticle (R848in) of the S/O system as an immumodulating material. Both R848out S/O and R848in S/O showed an approximate diameter of 265 nm. 7CrpR and R848 could gradually release from both R848out S/O and R848in S/O, however, higher release efficiency of R848 was observed from R848out S/O. Therefore, R848out S/O was chosen to use in TCIT of the pollinosis mouse models. Compared with the PBS solution, the reduction of total IgE, Cry j 1-
specific IgE and the production of the ratio of Cry j 1-specific IgG2a to IgE were observed by administration of S/O nanodispersions, suggesting the allergy was alleviated and Th1 immunity was improved, especially by the addition of R848 into S/O nanodispersions (Fig. 1-9).
Therefore, TCIT using S/O nanodispersions carrying 7CrpR and R848 is considered shifting the Th1 and Th2 immune balance toward the Th1 immunity.
Fig.1-9 Serum antibody levels in the pollinosis model mice after TCIT of S/O nanodispersions carrying 7CrpR with or without R848. The serum levels of (A) total IgE, (B) Cry j 1-specific IgE and (C) Cryj1-specific IgG2a antibodies were measured by ELISA.
(D) The ratios of Cry j 1-specific IgG2a to Cry j 1-specific IgE were calculated [44].
4. Objectives and outline of the thesis
The main objective of the present thesis is to develop and evaluate a safe, simple and effective AIT by using S/O nanodispersions loaded with the novel antigens, and thus, achieving the benefits of AIT rapidly and safely.
The main content of this thesis is consisting of the following chapters (Table 1-4):
Table 1-4 The outline content of present thesis
Chapter Formulation Antigen Administration
route 2 S/O nanodispersions Mixture of T cell epitope peptides TCIT 3 S/O nanodispersions Cedar pollen extract-
galactomannan conjugate TCIT
4 S/O nanodispersions Ovalbumin INIT
In Chapter 2, the TCIT for JCP by using S/O nanodispersions carrying T cell epitope peptides was developed and investigated. The use of T cell epitope peptides derived from pollen allergen was proposed to reduce the side effects of whole PE, without reducing the immunoregulatory activity. Because they do not cross-link with allergen-specific IgE bound to the surface of mast cells, treatment with T cell epitope peptides do not pose a potential risk of side effects. Although the hybrid peptide 7CrpR has already introduced in the S/O system, it has a relatively high molecular mass and the transcutaneous delivery of 7CrpR might be difficult due to the barrier function of the skin. Separated T cell epitope peptides are short and more easily permeate the skin.
In Chapter 3, the TCIT for JCP was improved by using S/O nanodispersions loaded with a JC PE-galactomannan conjugate. Although TCIT treated with T cell epitope peptides seems to be safe and effective, it is not applicable to all JCP patients due to the different genetic backgrounds in humans. Genetic differences in the T cell receptors may result in minor T cell epitope in some JCP patients. The modified allergen PE-galactomannan conjugate, including whole T cell epitopes information, is considered to be effective to all JCP patients.
In Chapter 4, a novel INIT was proposed by using S/O nanodispersions. INIT, as a non- invasive immunization approach, can induce both systemic and mucosal immune responses.
Ovalbumin was used as the model antigen. The efficacy and safety of INIT by using S/O nanodispersions were investigated. INIT using S/O nanodispersions is considered as another relatively simpler method of AIT for JCP in the future.
In Chapter 5, the main contents of this thesis were summarized and future perspective regarding the AIT for JCP was also discussed.
References
1. Masuyama, K.; Matsuoka, T.; Kamijo, A. Current status of sublingual immunotherapy for allergic rhinitis in Japan. Allergol. Int. 2018, 67, 320–325.
2. Gunawardana, N.C.; Durham, S.R. New approaches to allergen immunotherapy. Ann.
Allergy, Asthma Immunol. 2018, 121, 293–305.
3. Yamada, T.; Saito, H.; Fujieda, S. Present state of Japanese cedar pollinosis: The national affliction. J. Allergy Clin. Immunol. 2014, 133, 632-639.
4. Valenta, R. The future of antigen-specific immunotherapy of allergy. Nat. Rev. Immunol.
2002, 2, 446–453.
5. Su, Y.; Romeu-Bonilla, E.; Heiland, T. Next generation immunotherapy for tree pollen allergies. Hum. Vaccin. Immunother. 2017, 13, 2402–2415.
6. Shamji, M.H.; Durham, S.R. Mechanisms of allergen immunotherapy for inhaled allergens and predictive biomarkers. J. Allergy Clin. Immunol. 2017, 140, 1485–1498.
7. Prickett, S.R.; Rolland, J.M.; O’Hehir, R.E. Immunoregulatory T cell epitope peptides:
The new frontier in allergy therapy. Clin. Exp. Allergy 2015, 45, 1015–1026.
8. Larché, M.; Akdis, C.A.; Valenta, R. Immunological mechanisms of allergen-specific immunotherapy. Nat. Rev. Immunol. 2006, 6, 761–771.
9. Głobińska, A.; Boonpiyathad, T.; Satitsuksanoa, P.; Kleuskens, M.; van de Veen, W.;
Sokolowska, M.; Akdis, M. Mechanisms of allergen-specific immunotherapy: Diverse mechanisms of immune tolerance to allergens. Ann. Allergy, Asthma Immunol. 2018, 121, 306–312.
10. Nakamura, S.; Tsunoda, S.; Sakaida, H.; Masuda, S.; Said, A.S.; Takeuchi, K. Analysis of factors associated with cedar pollen sensitization and development of pollinosis in a young Japanese adult population. Allergol. Int. 2019, 68, 39–45.
11. Fujimura, T.; Kawamoto, S. Spectrum of allergens for Japanese cedar pollinosis and impact of component-resolved diagnosis on allergen-specific immunotherapy. Allergol.
Int. 2015, 64, 312–320.
12. Yasueda, H.; Yui, Y.; Shimizu, T.; Shida, T. Isolation and partial characterization of the major allergen from Japanese cedar (Cryptomeria japonica) pollen. J. Allergy Clin.
Immunol. 1983, 71, 77–86.
13. Sone, T.; Morikubo, K.; Miyahara, M.; Shimizu, K.; Tsunoo, H.; Kino, K.; Alerts, E. T Cell Epitopes in Japanese Cedar (Cryptomeria japonica) Pollen Allergens: Choice of Major T Cell Epitopes in Cry j 1 and Cry j 2 Toward Design of the Peptide-Based Immunotherapeutics for the Management of Japanese Cedar Pollinosis. J. Immunol.
1998, 161, 448–457.
14. Noon, L. PROPHYLACTIC INOCULATION AGAINST HAY FEVER. Lancet 1911, 177, 1572–1573.
15. Kiel, M.A.; Röder, E.; Gerth van Wijk, R.; Al, M.J.; Hop, W.C.J.; Rutten-van Mölken, M.P.M.H. Real-life compliance and persistence among users of subcutaneous and sublingual allergen immunotherapy. J. Allergy Clin. Immunol. 2013, 132, 353-360.e2.
16. Takaishi, S.; Saito, S.; Kamada, M.; Otori, N.; Kojima, H.; Ozawa, K.; Takaiwa, F.
Evaluation of basophil activation caused by transgenic rice seeds expressing whole T cell epitopes of the major Japanese cedar pollen allergens. Clin. Transl. Allergy 2019, 9, 1–8.
17. Murakami, D.; Kubo, K.; Sawatsubashi, M.; Kikkawa, S.; Ejima, M.; Saito, A.; Kato, A.; Komune, S. Phase I/II study of oral immunotherapy with Cry j1-galactomannan conjugate for Japanese cedar pollinosis. Auris Nasus Larynx 2014, 41, 350–358.
18. Murakami, D.; Sawatsubashi, M.; Kikkawa, S.; Ejima, M.; Saito, A.; Kato, A.; Komune, S. Safety and efficacy of a new regimen of short-term oral immunotherapy with Cry j 1- galactomannan conjugate for Japanese cedar pollinosis: A prospective, randomized, open-label study. Allergol. Int. 2015, 64, 161–168.
19. Murakami, D.; Sawatsubashi, M.; Kikkawa, S.; Ejima, M.; Saito, A.; Kato, A.; Komune, S. Effect of short-term oral immunotherapy with Cry j1-galactomannan conjugate on quality of life in Japanese cedar pollinosis patients: A prospective, randomized, open- label study. Auris Nasus Larynx 2016, 43, 50–55.
20. Murakami, D.; Sawatsubashi, M.; Omori, H.; Saito, A.; Kato, A.; Komune, S.;
Nakagawa, T. Safety and efficacy of short-term oral immunotherapy with Cry j 1- galactomannan conjugate for Japanese cedar pollinosis: a randomized controlled trial.
Sci. Rep. 2017, 7, 46142.
21. Su, Y.; Connolly, M.; Marketon, A.; Heiland, T. CryJ-LAMP DNA Vaccines for Japanese Red Cedar Allergy Induce Robust Th1-Type Immune Responses in Murine Model. J. Immunol. Res. 2016, 2016, 1-15.
22. Su, Y.; Romeu-Bonilla, E.; Anagnostou, A.; Fitz-Patrick, D.; Hearl, W.; Heiland, T.
Safety and long-term immunological effects of CryJ2-LAMP plasmid vaccine in Japanese red cedar atopic subjects: A phase I study. Hum. Vaccin. Immunother. 2017, 13, 2804–2813.
23. Hirahara, K.; Saito, S.; Serizawa, N.; Sasaki, R.; Sakaguchi, M.; Inouye, S.; Taniguchi, Y.; Kaminogawa, S.; Shiraishi, A. Oral administration of a dominant T-cell determinant peptide inhibits allergen-specific TH1 and TH2 cell responses in Cry j 2-primed mice. J.
Allergy Clin. Immunol. 1998, 102, 961–967.
24. Murasugi, T.; Nakagami, Y.; Yoshitomi, T.; Hirahara, K.; Yamashita, M.; Taniguchi, Y.; Sakaguchi, M.; Ito, K. Oral administration of a T cell epitope inhibits symptoms and reactions of allergic rhinitis in Japanese cedar pollen allergen-sensitized mice. Eur. J.
Pharmacol. 2005, 510, 143–148.
25. Yoshitomi, T.; Hirahara, K.; Kawaguchi, J.; Serizawa, N.; Taniguchi, Y.; Saito, S.;
Sakaguchi, M.; Inouye, S.; Shiraishi, A. Three T-cell determinants of Cry j 1 and Cry j 2, the major Japanese cedar pollen antigens, retain their immunogenicity and tolerogenicity in a linked peptide. Immunology 2002, 107, 517–522.
26. Toshio Sone, Keiko Morikubo, Michinori Miyahara, Naoki Komiyama, Kimiko Shimizu, H.T. and K.K. T Cell Epitopes in Japanese Cedar (Cryptomeria japonica) Pollen Allergens: Choice of Major T Cell Epitopes in Cry j 1 and Cry j 2 Toward Design of the Peptide-Based Immunotherapeutics for the Management of Japanese Cedar Pollinosis. J.
Immunol. 1998, 161, 448–457.
27. Tsunematsu, M.; Yamaji, T.; Kozutsumi, D.; Murakami, R.; Kimura, S.; Kino, K. Effect of Cry-consensus peptide, a novel recombinant peptide for immunotherapy of Japanese cedar pollinosis, on an experimental allergic rhinitis model in B10.S mice. Allergol. Int.
2007, 56, 465–72.
28. Takaiwa, F.; Yang, L. Development of a rice-based peptide vaccine for Japanese cedar and cypress pollen allergies. Transgenic Res. 2014, 23, 573–584.
29. Takagi, H.; Saito, S.; Yang, L.; Nagasaka, S.; Nishizawa, N.; Takaiwa, F. Oral immunotherapy against a pollen allergy using a seed-based peptide vaccine. Plant Biotechnol. J. 2005, 3, 521–533.
30. Kawabe, Y.; Hayashida, Y.; Numata, K.; Harada, S.; Hayashida, Y.; Ito, A.; Kamihira, M. Oral Immunotherapy for Pollen Allergy Using T-Cell Epitope-Containing Egg White Derived from Genetically Manipulated Chickens. PLoS One 2012, 7, e48512.
31. Yamanaka, K. ichi; Yuta, A.; Kakeda, M.; Sasaki, R.; Kitagawa, H.; Gabazza, E.C.;
Okubo, K.; Kurokawa, I.; Mizutani, H. Induction of IL-10-producing regulatory T cells with TCR diversity by epitope-specific immunotherapy in pollinosis. J. Allergy Clin.
Immunol. 2009, 124, 842-845.
32. Kitaoka, M.; Shin, Y.; Kamiya, N.; Kawabe, Y.; Kamihira, M.; Goto, M. Transcutaneous Peptide Immunotherapy of Japanese Cedar Pollinosis Using Solid-in-Oil Nanodispersion Technology. AAPS PharmSciTech 2015, 16, 1418–1424.
33. Kong, Q.; Kitaoka, M.; Wakabayashi, R.; Kamiya, N.; Goto, M. Transcutaneous immunotherapy of pollinosis using solid-in-oil nanodispersions loaded with T cell
epitope peptides. Int. J. Pharm. 2017, 529, 401–409.
34. Kong, Q.; Higasijima, K.; Wakabayashi, R.; Tahara, Y.; Kitaoka, M.; Obayashi, H.; Hou, Y.; Kamiya, N.; Goto, M. Transcutaneous Delivery of Immunomodulating Pollen Extract-Galactomannan Conjugate by Solid-in-Oil Nanodispersions for Pollinosis Immunotherapy. Pharmaceutics 2019, 11, 563.
35. Fujimura, T.; Fujinami, K.; Ishikawa, R.; Tateno, M.; Tahara, Y.; Okumura, Y.; Ohta, H.; Miyazaki, H.; Taniguchi, M. Recombinant Fusion Allergens, Cry j 1 and Cry j 2 from Japanese Cedar Pollen, Conjugated with Polyethylene Glycol Potentiate the Attenuation of Cry j 1-Specific IgE Production in Cry j 1-Sensitized Mice and Japanese Cedar Pollen Allergen-Sensitized Monkey. Int. Arch. Allergy Immunol. 2015, 168, 32–43.
36. Wakasa, Y.; Takagi, H.; Hirose, S.; Yang, L.; Saeki, M.; Nishimura, T.; Kaminuma, O.;
Hiroi, T.; Takaiwa, F. Oral immunotherapy with transgenic rice seed containing destructed Japanese cedar pollen allergens, Cry j 1 and Cry j 2, against Japanese cedar pollinosis. Plant Biotechnol. J. 2013, 11, 66–76.
37. Wakasa, Y.; Takagi, H.; Watanabe, N.; Kitamura, N.; Fujiwara, Y.; Ogo, Y.; Hayashi, S.; Yang, L.; Ohta, M.; Tin, W.W.T.; et al. Concentrated protein body product derived from rice endosperm as an oral tolerogen for allergen-specific immunotherapy-a new mucosal vaccine formulation against Japanese cedar pollen allergy. PLoS One 2015, 10, 1–22.
38. Toda, M.; Sato, H.; Takebe, Y.; Taniguchi, Y.; Saito, S.; Inouye, S.; Takemori, T.;
Sakaguchi, M. Inhibition of immunoglobulin E response to Japanese cedar pollen allergen (Cry j 1) in mice by DNA immunization: Different outcomes dependent on the plasmid DNA inoculation method. Immunology 2000, 99, 179–186.
39. Toda, M.; Kasai, M.; Hosokawa, H.; Nakano, N.; Taniguchi, Y.; Inouye, S.;
Kaminogawa, S.; Takemori, T.; Sakaguchi, M. DNA vaccine using invariant chain gene for delivery of CD4+T cell epitope peptide derived from Japanese cedar pollen allergen inhibits allergen-specific IgE response. Eur. J. Immunol. 2002, 32, 1631–1639.
40. Suzuki, M.; Ohta, N.; Min, W.P.; Matsumoto, T.; Min, R.; Zhang, X.; Toida, K.;
Murakami, S. Immunotherapy with CpG DNA conjugated with T-cell epitope peptide of an allergenic Cry j 2 protein is useful for control of allergic conditions in mice. Int.
Immunopharmacol. 2007, 7, 46–54.
41. Kaburaki, Y.; Fujimura, T.; Kurata, K.; Masuda, K.; Toda, M.; Yasueda, H.; Chida, K.;
Kawarai, S.; Sakaguchi, M. Induction of Th1 immune responses to Japanese cedar pollen allergen (Cry j 1) in mice immunized with Cry j 1 conjugated with CpG oligodeoxynucleotide. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 157–161.
42. Hoang, V. Van; Zou, Y.; Kurata, K.; Enomoto, K. Expression of recombinant T-cell epitopes of major Japanese cedar pollen allergens fused with cholera toxin B subunit in Escherichia coli. Protein Expr. Purif. 2015, 109, 62–69.
43. Ishii, M.; Koyama, A.; Iseki, H.; Narumi, H.; Yokoyama, N.; Kojima, N. Anti-allergic potential of oligomannose-coated liposome-entrapped Cry j 1 as immunotherapy for Japanese cedar pollinosis in mice. Int. Immunopharmacol. 2010, 10, 1041–1046.
44. Kitaoka, M.; Naritomi, A.; Kawabe, Y.; Kamihira, M.; Kamiya, N.; Goto, M.
Transcutaneous pollinosis immunotherapy using a solid-in-oil nanodispersion system carrying T cell epitope peptide and R848. Bioeng. Transl. Med. 2017, 2, 102–108.
45. O’Hehir, R.E.; Prickett, S.R.; Rolland, J.M. T Cell Epitope Peptide Therapy for Allergic Diseases. Curr. Allergy Asthma Rep. 2016, 16, 1–9.
46. Saito S, Hirahara K, Kawaguchi J, Serizawa N, Hino K, T.Y. Identification of T cell determinants in Cry j 1 and Cry j 2 of size suitable for immunotherapy against Japanese cedar pollinosis. Annu Rep Sankyo Res Lab 2000, 49–58.
47. Immonen, A.; Farci, S.; Taivainen, A.; Partanen, J.; Pouvelle-Moratille, S.; Närvänen, A.; Kinnunen, T.; Saarelainen, S.; Rytkönen-Nissinen, M.; Maillere, B.; et al. T Cell Epitope-Containing Peptides of the Major Dog Allergen Can f 1 as Candidates for Allergen Immunotherapy. J. Immunol. 2005, 175, 3614–3620.
48. Sakaguchi, M.; Hirahara, K.; Fujimura, T.; Toda, M. Approaches to immunotherapies for Japanese cedar pollinosis. Auris Nasus Larynx 2011, 38, 431–438.
49. Ricketti, P.A.; Alandijani, S.; Lin, C.H.; Casale, T.B. Investigational new drugs for allergic rhinitis. Expert Opin. Investig. Drugs 2017, 26, 279–292.
50. Takaishi, S.; Saito, S.; Endo, T.; Asaka, D.; Wakasa, Y.; Takagi, H.; Ozawa, K.; Takaiwa, F.; Otori, N.; Kojima, H. T-cell activation by transgenic rice seeds expressing the genetically modified Japanese cedar pollen allergens. Immunology 2019, 158, 94–103.
51. Takagi, H.; Hiroi, T.; Yang, L.; Tada, Y.; Yuki, Y.; Takamura, K.; Ishimitsu, R.;
Kawauchi, H.; Kiyono, H.; Takaiwa, F. A rice-based edible vaccine expressing multiple T cell epitopes induces oral tolerance for inhibition of Th2-mediated IgE responses. Proc.
Natl. Acad. Sci. U. S. A. 2005, 102, 17525–17530.
52. Mondoulet, L.; Dioszeghy, V.; Thébault, C.; Benhamou, P.H.; Dupont, C. Epicutaneous immunotherapy for food allergy as a novel pathway for oral tolerance induction.
Immunotherapy 2015, 7, 1293–1305.
53. Langlois, A.; Graham, F.; Bégin, P. Epicutaneous peanut patch device for the treatment of peanut allergy. Expert Rev. Clin. Immunol. 2019, 15, 449–460.
54. Cabauatan, C.R.; Campana, R.; Niespodziana, K.; Reinisch, C.; Lundberg, U.; Meinke,
A.; Henning, R.; Neubauer, A.; Valenta, R. Heat-labile Escherichia coli toxin enhances the induction of allergen-specific IgG antibodies in epicutaneous patch vaccination.
Allergy Eur. J. Allergy Clin. Immunol. 2017, 72, 164–168.
55. Siebeneicher, S.; Reuter, S.; Wangorsch, A.; Krause, M.; Foetisch, K.; Heinz, A.; Naito, S.; Reuter, A.; Taube, C.; Vieths, S.; et al. Epicutaneous immunotherapy with a hypoallergenic Bet v 1 suppresses allergic asthma in a murine model. Allergy Eur. J.
Allergy Clin. Immunol. 2015, 70, 1559–1568.
56. Siebeneicher, S.; Reuter, S.; Krause, M.; Wangorsch, A.; Maxeiner, J.; Wolfheimer, S.;
Schülke, S.; Naito, S.; Heinz, A.; Taube, C.; et al. Epicutaneous immune modulation with Bet v 1 plus R848 suppresses allergic asthma in a murine model. Allergy Eur. J.
Allergy Clin. Immunol. 2014, 69, 328–337.
57. Agostinis, F.; Forti, S.; Di Berardino, F. Grass transcutaneous immunotherapy in children with seasonal rhinoconjunctivitis. Allergy 2010, 65, 410–411.
58. Witten, M.; Malling, H.J.; Blom, L.; Poulsen, B.C.; Poulsen, L.K. Is intralymphatic immunotherapy ready for clinical use in patients with grass pollen allergy? J. Allergy Clin. Immunol. 2013, 132.
59. Hylander, T.; Latif, L.; Petersson-Westin, U.; Cardell, L.O. Intralymphatic allergen- specific immunotherapy: An effective and safe alternative treatment route for pollen- induced allergic rhinitis. J. Allergy Clin. Immunol. 2013, 131, 412–420.
60. Hylander, T.; Larsson, O.; Petersson-Westin, U.; Eriksson, M.; Kumlien Georén, S.;
Winqvist, O.; Cardell, L.O. Intralymphatic immunotherapy of pollen-induced rhinoconjunctivitis: A double-blind placebo-controlled trial. Respir. Res. 2016, 17, 1–9.
61. Senti, G.; Prinz Vavricka, B.M.; Erdmann, I.; Diaz, M.I.; Markus, R.; McCormack, S.J.;
Simard, J.J.; Wüthrich, B.; Crameri, R.; Graf, N.; et al. Intralymphatic allergen administration renders specific immunotherapy faster and safer: A randomized controlled trial. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 17908–17912.
62. Tsai, J.J.; Liao, E.C.; Tsai, F.H.; Hsieh, C.C.; Lee, M.F. The effect of local nasal immunotherapy in allergic rhinitis: Using strips of the allergen dermatophagoides pteronyssinus. J. Asthma 2009, 46, 165–170.
63. Marcucci, F.; Sensi, L.G.; Caffarelli, C.; Cavagni, G.; Bernardini, R.; Tiri, A.; Riva, G.;
Novembre, E. Low-dose local nasal immunotherapy in children with perennial allergic rhinitis due to Dermatophagoides. Allergy Eur. J. Allergy Clin. Immunol. 2002, 57, 23–
28.
64. Langlois, A.; Graham, F.; Bégin, P. Epicutaneous peanut patch device for the treatment of peanut allergy. Expert Rev. Clin. Immunol. 2019, 15, 449–460.
65. Scheiblhofer, S.; Machado, Y.; Feinle, A.; Thalhamer, J.; Hüsing, N.; Weiss, R. Potential of nanoparticles for allergen-specific immunotherapy–use of silica nanoparticles as vaccination platform. Expert Opin. Drug Deliv. 2016, 13, 1777–1788.
66. Pohlit, H.; Bellinghausen, I.; Frey, H.; Saloga, J. Recent advances in the use of nanoparticles for allergen-specific immunotherapy. Allergy Eur. J. Allergy Clin.
Immunol. 2017, 72, 1461–1474.
67. Walter, F.; Schöll, I.; Untersmayr, E.; Ellinger, A.; Boltz-Nitulescu, G.; Scheiner, O.;
Gabor, F.; Jensen-Jarolim, E. Functionalisation of allergen-loaded microspheres with wheat germ agglutinin for targeting enterocytes. Biochem. Biophys. Res. Commun. 2004, 315, 281–287.
68. Roth-Walter, F.; Schöll, I.; Untersmayr, E.; Fuchs, R.; Boltz-Nitulescu, G.; Weissenböck, A.; Scheiner, O.; Gabor, F.; Jensen-Jarolim, E. M cell targeting with Aleuria aurantia lectin as a novel approach for oral allergen immunotherapy. J. Allergy Clin. Immunol.
2004, 114, 1362–1368.
69. Schöll, I.; Kopp, T.; Bohle, B.; Jensen-Jarolim, E. Biodegradable PLGA Particles for Improved Systemic and Mucosal Treatment of Type I Allergy. Immunol. Allergy Clin.
North Am. 2006, 26, 349–364.
70. Schöll, I.; Weissenböck, A.; Förster-Waldl, E.; Untersmayr, E.; Walter, F.; Willheim, M.; Boltz-Nitulescu, G.; Scheiner, O.; Gabor, F.; Jensen-Jarolim, E. Allergen-loaded biodegradable poly(D,L-lactic-co-glycolic) acid nanoparticles down-regulate an ongoing Th2 response in the BALB/c mouse model. Clin. Exp. Allergy 2004, 34, 315–
321.
71. Batanero, E.; Barral, P.; Villalba, M.; Rodríguez, R. Encapsulation of Ole e 1 in biodegradable microparticles induces Th1 response in mice: A potential vaccine for allergy. J. Control. Release 2003, 92, 395–398.
72. Kojima, N.; Biao, L.; Nakayama, T.; Ishii, M.; Ikehara, Y.; Tsujimura, K.
Oligomannose-coated liposomes as a therapeutic antigen-delivery and an adjuvant vehicle for induction of in vivo tumor immunity. J. Control. Release 2008, 129, 26–32.
73. Kitaoka, M.; Wakabayashi, R.; Kamiya, N.; Goto, M. Solid-in-oil nanodispersions for transdermal drug delivery systems. Biotechnol. J. 2016, 11, 1375–1385.
74. Wakabayashi, R.; Sakuragi, M.; Kozaka, S.; Tahara, Y.; Kamiya, N.; Goto, M. Solid-in- Oil Peptide Nanocarriers for Transcutaneous Cancer Vaccine Delivery against Melanoma. Mol. Pharm. 2018, 15, 955–961.
75. Tahara, Y.; Honda, S.; Kamiya, N.; Piao, H.; Hirata, A.; Hayakawa, E.; Fujii, T.; Goto, M. A solid-in-oil nanodispersion for transcutaneous protein delivery. J. Control. Release


![Fig. 2-1 Immune system of the skin [10]. Ag, antigen; dDCs, dermal dendritic cells; LC, Langerhans cells; SC, stratum corneum](https://thumb-ap.123doks.com/thumbv2/123deta/9810440.1885958/35.892.163.740.142.592/immune-system-antigen-dermal-dendritic-langerhans-stratum-corneum.webp)
![Fig. 2-2 The strategies developed for the enhancement of TCIT [15]. Reprinted from Vaccine Vol](https://thumb-ap.123doks.com/thumbv2/123deta/9810440.1885958/36.892.119.789.402.568/fig-strategies-developed-enhancement-tcit-reprinted-vaccine-vol.webp)
![Fig. 2-4 A) Seven major T cell epitopes of Cry j 1 and Cry j 2 in human patients with Japanese cedar pollinosis [40]](https://thumb-ap.123doks.com/thumbv2/123deta/9810440.1885958/39.892.208.681.581.976/seven-major-epitopes-human-patients-japanese-cedar-pollinosis.webp)