Identification of iron- and sulfur-induced genes in Sulfobacillus acidophilus
8
0
0
全文
(2) International Symposium on Extremophiles and Their Applications 2005. the metabolism of iron and sulfur oxidation in a moderately thermophilic acidophile, Sulfobacillus acidophilus. Sulfobacillus acidophilus is an acidophilic, gram-positive eubacterial species that oxidizes ferrous iron, elemental sulfur and mineral sulfides optimally at a temperature of about 50°C [17]. Although this species can grow autotrophically, growth is generally most robust when it uses inorganic energy sources and organic carbon. Two subtractive hybridization experiments were performed, the first comparing cells grown on ferrous iron and yeast extract with cells grown on yeast extract only, the second comparing cells grown on sulfur and sucrose with cells grown on sucrose only.. 2. Materials and Methods 2.1 Bacterial strain and culture conditions Sulfobacillus acidophilus strain NAL (DSM 10332T) was grown at 48°C in flasks shaken at 140 rpm. The medium contained MgSO4.7H2O (0.5 g l-1), (NH4)2SO4 (0.4 g l-1), K2HPO4 (0.2 g l-1) and FeSO4.7H2O (10 mg l-1). The medium was initially adjusted, using sulfuric acid, to pH 1.7 when ferrous iron was the substrate (50 mM FeSO4.7H2O, 0.025% w/v yeast extract), to pH 2.0 for heterotrophic growth on yeast extract (0.025% w/v) or sucrose (2.5 mM) and to pH 2.5 for growth on sulfur (sulfur flowers at 1 g l-1 and 2.5 mM sucrose). Cultures were harvested at mid- to late-exponential growth phase. Cultures used for analysis of genes potentially up-regulated by iron were grown either for several serial cultures in the presence of ferrous iron and yeast extract (iron-adapted culture) or in the presence of yeast extract only before addition of ferrous iron and further incubation (iron-stressed culture). 2.2 RNA isolation and cDNA synthesis RNA was isolated using TRIzol reagent (Invitrogen, Carlsbad USA), according to the manufacturer’s protocol with a modification to improve cell lysis. Frozen cell pellets were resuspended in TRIzol reagent, 1 ml portions were added to Q-Biogene Lysing Matrix E vials and subjected to bead-beating for 40 s at speed 6 in a Hybaid RiboLyser instrument. Contaminating DNA was removed from the RNA by treatment with RNAse-free DNAse. Double-stranded cDNA was synthesized with the Universal RiboClone cDNA synthesis system (Promega, Madison, USA) using random hexanucleotides. First-strand synthesis was carried out for 4 h, and second-strand synthesis was continued overnight. The cDNA was purified using a PCR purification kit (Qiagen, Hilden, Germany). 2.3 RDA Micro-representational difference analysis was carried out as described previously [16]. Between each step, DNA was purified using a Qiagen PCR purification kit. For the first analysis, cells were grown on yeast extract to early exponential phase. The pH was then lowered to 1.65 using sterile dilute sulfuric acid and the culture was divided into two. One half, the tester culture, received ferrous iron to a final concentration of 25 mM. The other half, the driver culture, remained unchanged. Both cultures were incubated for 3 h.. 117.
(3) International Symposium on Extremophiles and Their Applications 2005. For the second analysis, cells grown on sulfur and sucrose were used as the tester culture and cells grown on sucrose only as the driver culture. Difference products were cloned into the pCR2.1-TOPO vector using the TOPO TA cloning kit (Invitrogen, Carlsbad, USA). Clones were analyzed by colony PCR using primers M13for/rev, and PCR products of suitable sizes were sequenced. An inverse PCR approach was used to facilitate sequencing of gene fragments of interest. 2.4 RT-PCR Reverse transcription of RNA used the Promega ImProm-II Reverse Transcription System with random hexanucleotides and 5.0 mM MgCl2. Subsequent PCR used Taq polymerase (Fermentas) and cDNA corresponding to 50 ng of RNA in a final volume of 25 !l. Suitable primers giving products of 200-250 bp size were designed for the obtained sequences. The rpoB gene of S. acidophilus was partially sequenced and used as an external control in the RT-PCR reactions. Design of primers for RT-PCR used the programme Primer3 [18]. Sequences alignments and calculation of neighbour joining trees were performed using ClustalX version 1.81 [19].. 3. Results Sulfobacillus acidophilus used ferrous iron or elemental sulfur as energy sources during growth which was dependent on yeast extract or sucrose as carbon sources. Ferrous ironoxidizing cells were only exposed to iron for a short period (3 h) before RNA extraction. The initial cDNA amplicons and the difference products (DPs) obtained after up to two (ironoxidizing cells) or three (sulfur-oxidizing cells) rounds of subtractive hybridization are shown (Fig. 1). The DP2 (iron) and the DP3 (sulfur) samples were subjected to shot-gun cloning and sequencing. The amino acid sequences deduced from the cDNA restriction fragments derived from the iron-oxidizing cells showed homologies to three subunits of trimeric molybdopterin reductases of the dimethyl sulfoxide (DMSO) reductase family, an ABC-dependent protease, cysteine synthase, cytochrome bd ubiquinol oxidase subunits I and II, and a dihydrolipoamide dehydrogenase component. One sequence with no recognizable nucleotide or amino acid homologies was obtained. The differential expression of a number of these fragments was investigated using comparative reverse transcription PCR. In adddition to a cells exposed to iron for a relatively short time (as for those used in the mRDA analysis), cells adapted to iron over several serial cultures were also examined (Fig. 2). Whereas upregulation of all tested cDNA fragments in the cells exposed to iron for three hours was confirmed, only the oxidoreductase fragments and the protease sequence remained upregulated in iron-adapted cultures when compared to RNA isolated from cultures grown on yeast extract in the absence of iron.. 118.
(4) International Symposium on Extremophiles and Their Applications 2005. Fig. 1 Agarose gels showing the PCR products obtained during mRDA using a) iron/yeast extract- and yeast extract-grown cells and b) sulfur/sucrose- and sucrose-grown cells. M: 100 bp molecular size marker; Amp: initial cDNA amplicon; DP1, 2, 3: difference products obtained after one, two or three rounds of subtractive hybridization.. Fig. 2 Confirmation of differential expression of the cDNA fragments obtained from iron-induced cells using RT-PCR. The left panel shows the analysis of RNA from cells exposed to iron for three hours. The right panel shows results obtained from cells adapted to iron over several serial cultures.. 119.
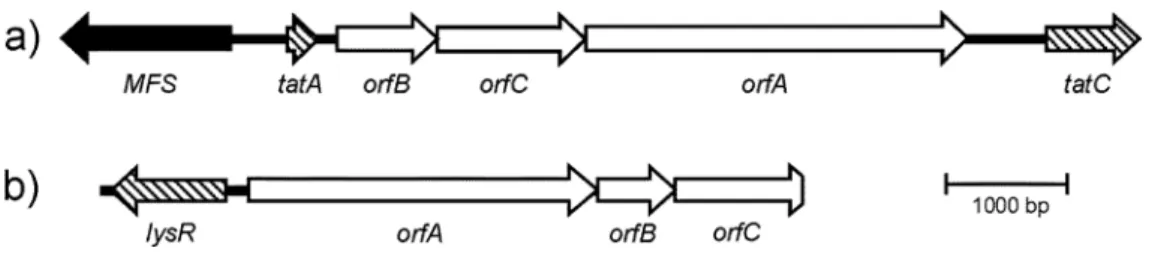
(5) International Symposium on Extremophiles and Their Applications 2005. The sequences of the cDNA fragments derived from the sulfur/sucrose-grown cells showed homologies to genes encoding two different subunits of DMSO-family oxidoreductases, a sulfide dehydrogenase of the sulfide-quinone reductase / flavocytochrome c family, an excisionase and 16S and 23S rRNA. The oxidoreductase genes were tested for differential expression by RT-PCR. Whereas the DMSO-family oxidoreductase genes appeared to be clearly differentially regulated, the sulfide dehydrogenase gene only seemed to be slightly upregulated in sucrose/sulfur- as opposed to sucrose-grown cells (data not shown). The different subunits of the DMSO-reductase family genes obtained in the two experiments formed gene clusters with the order BCA for the iron-induced cells and the order ABC for the sulfur-induced cells. These genes encode the catalytic, molybdopterin-containing subunit (A), an intermediate, Fe-S cluster-containing subunit (B), and the membrane-anchor subunit (C) of trimeric DMSO-family reductases.. Fig. 3 Diagram of the arrangement of the genes encoding the subunits (A, B and C) of DMSO-reductase family proteins obtained with a) iron-induced cells and b) sulfur-induced cells of S. acidophilus.. The deduced amino acid sequence of the iron-induced oxidoreductase subunits indicated a presumptive signal peptide containing a twin arginine motif in the A subunit, suggesting an extracellular localization of the protein. No additional export sequences could be detected in any of the other ORFs. Additionally, the three ORFs obtained with iron-induced cells were framed by ORFs showing homologies to the tatA and tatC genes from the twin arginine translocation (TAT) pathway. An ORF coding for a major facilitator superfamily (MFS) sugar-/phosphate-transporter was found upstream of the tatA gene. RT-PCR experiments indicated that B, C and A are transcribed on a single mRNA and that the tatA and tatC genes seem to be transcribed independently from the BCA message (data not shown). It should be noted here that tatA and tatC are generally situated directly adjacent to each other and that this insertion of ORFs whose products might represent TAT substrates between the tat genes is unusual. No export signals could be identified in any of the sulfur-upregulated DMSO-reductase subunit genes, suggesting that they encoded an intracellular protein. A LysR family type transcriptional regulator was found upstream of ORF A. This is a common arrangement for LysR family trancriptional regulators and genes which are controlled by them. An alignment of the amino acid sequences of the A subunits of different DMSO-reductase family enzymes shows that the subunits loosely cluster according to their substrate specificities. The sequences of the reductases identified within this study cluster together with enzymes performing reductive cleavages of sulfur-sulfur bonds (Fig. 4).. 120.
(6) International Symposium on Extremophiles and Their Applications 2005. Fig. 4 Neighbour joining tree constructed from full amino acid sequences of the catalytic, molybdopterin-containing A subunits of different DMSO-family oxidoreductases. The two sequences obtained in this study are printed in bold.. 4. Discussion The study presented here shows that mRDA could be used to identify differentially expressed genes in Sulfobacillus acidophilus. However, the use of cDNA restriction fragments produced with a single, four base pair-recognition cutter as targets for PCR amplification might lead to a loss of cDNA from genes without suitably situated restriction sites. On the other hand, the production of multiple restriction fragments from long cDNAs might lead to an over-representation of longer transcripts. Nevertheless, this method, as well as differential display PCR [20], should be useful for investigating differential gene expression in the absence of genome sequences. With respect to iron-induced genes, no sequences with a clear involvement in iron oxidation were recovered. Some of the sequences recovered indicated proteins, such as cytochrome bd oxidase and cysteine synthase [21], which could be involved in an oxidative stress response. This is not surprising given the formation of ferric iron during culture growth, which is an oxidant that leads to the formation of reactive oxygen species through the Fenton reaction. A major cell protein induced during iron oxidation by Sulfolobus metallicus was a potential anti-oxidant peroxidase [22]. The sequences obtained with sulfur-induced cells could be related to sulfur metabolism. However, there is no direct evidence for involvement of the putative DMSO-family reductase and sulfide dehydrogenase enzymes in sulfur oxidation by S. acidophilus. Current protein biochemical work involving a major cell protein induced in cells of S. acidophilus grown on. 121.
(7) International Symposium on Extremophiles and Their Applications 2005. sulfur revealed it to be a homologue of an electron-transferring flavoprotein (ETF) [23] (P. Caldwell and P. R. Norris, unpublished work). This heterodimer serves as an electron acceptor for a number of different dehydrogenases and transfers electrons to the main respiratory chain via ETF-ubiquinone oxidoreductase. Interestingly, RT-PCR indicated greater expression of the encoding gene in iron- rather than sulfur-grown cells (data not shown). The comparison of autotrophically-grown iron- and sulfur-oxidizing S. acidophilus by mRDA should facilitate the identification of the genes involved in its lithotrophy, whereas the presence of organic nutrient sources in the experiments presented here could have complicated analysis of the inorganic substrate metabolism.. 5. Acknowledgements The work presented here was partly funded by the EU 6th framework programme integrated project “Biotechnology for Metal bearing materials in Europe” (BioMinE, http://biomine.brgm.fr).. 6. References [1] Rawlings, D.E., Dew, D. and du Plessis, C. (2003) Biomineralization of metal-containing ores and concentrates. Trends Biotechnol 21, 38-44. [2] Appia-Ayme, C., Bengrine, A., Cavazza, C., Giudici-Orticoni, M.T., Bruschi, M., Chippaux, M. and Bonnefoy, V. (1998) Characterization and expression of the co-transcribed cyc1 and cyc2 genes encoding the cytochrome c4 (c552) and a high-molecular-mass cytochrome c from Thiobacillus ferrooxidans ATCC 33020. FEMS Microbiol Lett 167, 171-177. [3] Appia-Ayme, C., Guiliani, N., Ratouchniak, J. and Bonnefoy, V. (1999) Characterization of an operon encoding two c-type cytochromes, an aa(3)-type cytochrome oxidase, and rusticyanin in Thiobacillus ferrooxidans ATCC 33020. Appl Environ Microbiol 65, 4781-4787. [4] Ramirez, P., Guiliani, N., Valenzuela, L., Beard, S. and Jerez, C.A. (2004) Differential protein expression during growth of Acidithiobacillus ferrooxidans on ferrous iron, sulfur compounds, or metal sulfides. Appl Environ Microbiol 70, 4491-4498. [5] Ramirez, P., Toledo, H., Guiliani, N. and Jerez, C.A. (2002) An exported rhodanese-like protein is induced during growth of Acidithiobacillus ferrooxidans in metal sulfides and different sulfur compounds. Appl Environ Microbiol 68, 1837-1845. [6] Yarzabal, A., Appia-Ayme, C., Ratouchniak, J. and Bonnefoy, V. (2004) Regulation of the expression of the Acidithiobacillus ferrooxidans rus operon encoding two cytochromes c, a cytochrome oxidase and rusticyanin. Microbiology 150, 2113-2123. [7] Tyson, G.W., Lo, I., Baker, B.J., Allen, E.E., Hugenholtz, P. and Banfield, J.F. (2005) Genome-directed isolation of the key nitrogen fixer Leptospirillum ferrodiazotrophum sp. nov. from an acidophilic microbial community. Appl Environ Microbiol 71, 6319-6324. [8] Ram, R.J., Verberkmoes, N.C., Thelen, M.P., Tyson, G.W., Baker, B.J., Blake II, R.C., Shah, M., Hettich, R.L. and Banfield, J.F. (2005) Community proteomics of a natural microbial biofilm. Science 308, 19151920. [9] Urich, T., Bandeiras, T.M., Leal, S.S., Rachel, R., Albrecht, T., Zimmermann, P., Scholz, C., Teixeira, M., Gomes, C.M. and Kletzin, A. (2004) The sulphur oxygenase reductase from Acidianus ambivalens is a multimeric protein containing a low-potential mononuclear non-haem iron centre. Biochem J 381, 137146. [10] Muller, F.H., Bandeiras, T.M., Urich, T., Teixeira, M., Gomes, C.M. and Kletzin, A. (2004) Coupling of the pathway of sulphur oxidation to dioxygen reduction: characterization of a novel membrane-bound thiosulphate:quinone oxidoreductase. Mol Microbiol 53, 1147-1160.. 122.
(8) International Symposium on Extremophiles and Their Applications 2005. [11] Kletzin, A., Urich, T., Muller, F., Bandeiras, T.M. and Gomes, C.M. (2004) Dissimilatory oxidation and reduction of elemental sulfur in thermophilic archaea. J Bioenerg Biomembr 36, 77-91. [12] Laska, S., Lottspeich, F. and Kletzin, A. (2003) Membrane-bound hydrogenase and sulfur reductase of the hyperthermophilic and acidophilic archaeon Acidianus ambivalens. Microbiology 149, 2357-2371. [13] Zimmermann, P., Laska, S. and Kletzin, A. (1999) Two modes of sulfite oxidation in the extremely thermophilic and acidophilic archaeon Acidianus ambivalens. Arch Microbiol 172, 76-82. [14] http://www.tigr.org/tdb/mdb/mdbinprogress.html. [15] Tyson, G.W., Chapman, J., Hugenholtz, P., Allen, E.E., Ram, R.J., Richardson, P.M., Solovyev, V.V., Rubin, E.M., Rokhsar, D.S. and Banfield, J.F. (2004) Community structure and metabolism through reconstruction of microbial genomes from the environment. Nature 428, 37-43. [16] Becker, P., Hufnagle, W., Peters, G. and Herrmann, M. (2001) Detection of differential gene expression in biofilm-forming versus planktonic populations of Staphylococcus aureus using micro-representationaldifference analysis. Appl Environ Microbiol 67, 2958-2965. [17] Norris, P.R., Clark, D.A., Owen, J.P. and Waterhouse, S. (1996) Characteristics of Sulfobacillus acidophilus sp. nov. and other moderately thermophilic mineral-sulphide-oxidizing bacteria. Microbiology 142, 775-783. [18] Rozen, S. and Skaletsky, H.J. (2000) Primer3 on the WWW for general users and for biologist programmers. In: Bioinformatics Methods and Protocols: Methods in Molecular Biology (Krawetz, S. and Misener, S., Eds.), pp. 365-386. Humana Press, Totowa, NJ, USA. [19] Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F. and Higgins, D.G. (1997) The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25, 4876-4882. [20] Paulino, L.C., de Mello, M.P. and Ottoboni, L.M. (2002) Differential gene expression in response to copper in Acidithiobacillus ferrooxidans analyzed by RNA arbitrarily primed polymerase chain reaction. Electrophoresis 23, 520-527. [21] Das, A., Silaghi-Dumitrescu, R., Ljungdahl, L.G. and Kurtz, D.M., Jr. (2005) Cytochrome bd oxidase, oxidative stress, and dioxygen tolerance of the strictly anaerobic bacterium Moorella thermoacetica. J Bacteriol 187, 2020-2029. [22] Burton, N.P., Williams, T.D. and Norris, P.R. (1995) A potential anti-oxidant protein in a ferrous ironoxidizing Sulfolobus species. FEMS Microbiol Lett 134, 91-95. [23] Tsai, M.H. and Saier, M.H. (1995) Phylogenetic characterization of the ubiquitous electron transfer flavoprotein families ETF-alpha and ETF-beta. Res Microbiol 146, 397-404.. 123.
(9)
図


関連したドキュメント
On a theoretical side, we state a sound probability space for lucidity and thus for modularity and prove that in this paradigm of lucidity, using a subtractive trade-off and either
Nevertheless, a suitable mix of analytical and numerical techniques allows us to detect a sequence of homoclinic bifurcations—analogous to those occurring in the
The mGoI framework provides token machine semantics of effectful computations, namely computations with algebraic effects, in which effectful λ-terms are translated to transducers..
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
An example of a database state in the lextensive category of finite sets, for the EA sketch of our school data specification is provided by any database which models the
A NOTE ON SUMS OF POWERS WHICH HAVE A FIXED NUMBER OF PRIME FACTORS.. RAFAEL JAKIMCZUK D EPARTMENT OF
In order to use the above radiation induced death rates G u ðtÞ and G q ðtÞ in an ODE model, first consider a cell cycle model for active and quiescent cells without the effects
Thus, in Section 5, we show in Theorem 5.1 that, in case of even dimension d > 2 of a quadric the bundle of endomorphisms of each indecomposable component of the Swan bundle
