TheJapanese Journalq广Psγthonontic Science 20!3
,
Vo],
32,
No、
1,
79−
87講演論文
『
知 覚科 学
』
へよ
う
こそ
一
ヒト
心理物 理 学
と
動 物神経 行 動学
と の
統合
一
力 丸 裕
同 志 社 大 学 生 命 医 科 学 部・
生命医科学研究科,
同志社 大学二 =一
ロ セ ンシ ング・
バ イオナビゲー
シ ョ ン研 究セ ンター
“ !!Welcome
to
sensory
science
−
lntegration
between
human
psychophysics
andanimal
neurobehaviology一
Hiroshi
RIQuIMARoux
Fa‘ulty 〔)
fLife
and Medicat S‘iences/Graduate School ofMedicalScien
‘es,
Doshisha
University
,
Neurosensing andBionavigation Research Center
.
Doshisha U海γθ厂5砂The
present
paper
describes common auditory systems frQm humans tobats
for
effective listening.
First,
the noise−
vocoded speech sounds and auditory induction are explained to consider reaレtime modification of neuronal signal processing,
Second, in order to discuss localized functions in the brain
,
the biosonar system ofb
劉ts andpit
−
falls!problems are introduced as examples.
Final1}〜fbr better understanding of ef 罫ective and precise listening,
usage ofstabilized carrier waves in the bat biosonar System and strategies fi〕r sophist 正⊂ated signal detections injammed
situa ロon are discussed after intreducing small wireless telemetry microphone system (Telemike).
Key words ;noise
−
vocoded speech sounds,
auditory induction,
brain
plasticity
, Telemike,
Doppler−
shift compen−
sation
,
echo arnplitude compensation,
carrier waves,
echo interference reductionは じ め に 知 覚につ い て解 明 する場合
,
一
般 的に行 動 学 的・
心 理 学的なアプロー
チ と神 経 生 理 学 的なアプロー
チ が あ る。
そ れ ぞ れの アプロー
チ に ヒ トを用い る方 法と ヒ ト以 外の 動 物を用い る方 法が あ る。 本 論 説で は,
ヒ ト と動 物と の 共 通 点に着 目 し,
都 合 良 く神 経 ネッ トワー
ク を創設す る 仕組み と,
機 能 局 在の仕 組みと,
効 率よく聴き取る仕組 みについ て,
そ れ ぞ れの アプロー
チ を用いて議 論 する、
1.
瞬 時にネッ トワー
クを改変
し都合
よ く知 覚 す る 仕 組 み 1.
1 脳によ る リア ル タ イ ム音声情報 創 成:劣 化 雑 音 音 声の知 覚音声の強弱を表 す 振 幅 包 絡 を残 し
,
音 声 信 号は音声信Correspollding address :Faculty of
Life
and Medical Sciences!Graduate
School of Medical Sciences,
Doshisha University,
Neurosensillg and Bionavigation Research Center
,
DQshishaUniversity
,
1−
3 Tatara Miyakodani,
Kyotanabe,
Kyoto 610−
D321
,
Iapan
,
E−
mail:hrikimar@ mail.
doshisha
.
ac.
jp
号と同じ周波数帯域を もつ雑 音で置 き換 えた音 を 劣 化 雑 音 音 声 (Noise
−
vocoded speech sound,
NVSS )と呼ん でいる (Figure 1)。
は じめて NVSS を聴い たとき に は,
音 声と し ての了解 度は ほ とんどないが,
NVSS を文 字化し た もの を黙 読後に同一
のNVSS を聴 くと, 了 解 度の高い音 声と して聴き取る こ とがで き る。 こ の よ う な訓 練 を繰 り返 す と, 文 字 情 報に よ る フ ィー
ドバ ッ ク な しで,
NVSS で構 成さ れ た文 章を了 解 する こ と が 短 期 間で可能にな る (RiquimarQux,
2006)。
NVSS に は基 本 周 波 数 成 分が 存 在 しない に もか かわらず,
ピッ チアクセ ン ト を知 覚 するこ と が できる。
換 言 すると,
基 本 周 波数の上 昇・
下降が知 覚さ れ るので あ る。 今 回 使 用し たの は,
12か ら13モー
ラか らな る短 文で ある。
音 声 信号の全体の帯 域を4000Hz まで に制 限 し,
バ ン ドパ ス・
フ ィル ター
によっ て 0か ら600Hz,
600か ら1500Hz,
1500か ら210e Hz.
2100 か ら4eOO Hz の4 帯 域に分 割 し,
そ れ ぞ れの帯 域の振 幅 包 絡を抽 出 する。
一
方,
0か ら4000・Hzの帯 域を持つ 白色 雑 音を音 声 信 号 と 同 じ周波 数 帯 域の バ ン ドパ ス・
フィ ル ター
に通して,
4 つ の帯域の白色 雑 音を 用意す る。
帯 域ごと に白色 雑 音と80 基礎心 理 学 研究 第32 巻 第1号 4000
で
(
N ヱ 属 O § コ す O 」 」 060 』 02 0.
3 0.
4 Time(s) o.
58萋
oaξ
一
〇.
5一
・
1 0 0』 02 0.
3 0、
4 T o(s) 4000(
N ヱ X§
弓2
」 篇me (s> 0 ℃ 書3
竃 繹 く fime(s>Figure 1
.
Comparisonbetween
an original speech sound“
hashi”
(chopsticks )and a synthesized noise
−
vocoded speechsound (NVSS )from the originaL Origical speech sound (top)and 4
−band
NVSS (botton1
).
Left:Temporal change infrequency structure (sound
−
spectrogram ).
Right:Amplitude change in time (from Rlqu正maroux,
2006).
振 幅 包 絡 を掛け合 わせ, そ れ ぞ れ の帯 域に対応する振 幅 変 調 雑 音を作 成す る。 最後に,
4帯域の振 幅 変 調 雑 音 を 加 算 して,
NVSS が完 成する。
fMRI (fun・ti・n・l m ・gn
・ti・・es ・n ・n ・e im・gi・g,
黼 旨的 磁 気 共 鳴 画 像 )を用いた計 測の結果,
通常の音 声を聴取 す る際に は左 右の上側 頭回だ けが賦活さ れ る が,
劣化雑 音 音 声を聴取す ると上側頭 回に加えて,
島皮 質,
補 足 運 動 野,
前 帯 状同な どの領 域 も賦 活され る よ うで あ る (橘・
力 丸,
2005)。
す な わ ち、
通常の音声 聴取時に賦活さ れ る領域以外の複数の領域を動員して,
雑 音を音 声に変 換 す る作 業がな されるのである。
1.
2 脳に よる リ アル タ イ ム修 復:連 続聴 錯 覚 連 続し た音 楽や音 声を部 分 的に消 去し,
消 去した部 分 に広 帯 域 雑 音を挿 入する と,
欠 損し た情 報が脳に よ り創 成・
修 復さ れ,
連 続し た音楽や音 声として聴 取さ れ,
連 続 聴 錯 覚 として 良 く知ら れ てい る。
初め て聴 く曲で も,
楽 器 演 奏で も,
歌 曲で も都合良く修 復 さ れ る。 連続 聴 錯 覚は,
げっ 歯動物の スナネズミを 用い た行 動 学 的 実 験で も 示 さ れ.
連 続 音と して知 覚さ れる (Kobayasi,
Usami,
& Riquimaroux,
2012)。 こ の機 能に よ り,
環 境 雑 音に よ り 音響 的には途 切れ途 切れとなっ た信 号 音 を連 続 音と し て 知覚す るこ とがで きる。
雑 音の音 圧レ ベ ル が 高いほど連 続音とな りや すい。
しか し,
雑 音の周 波 数 帯 域に信 号 音 が含まれていない と,
連 続 音には な ら ない。 これ らの特 徴は,
ヒ トで も動物で も共 通 してい る(Figure 2)。
2.
脳 内 機 能 局 在の仕 組み 2.
1 コウモリを 用いた 研 究 コウモ リ は,
超 音 波 音 声を用い て生物ソ ナー
と呼ばれ る精 度の高い計 測 を行っ て い る。
生 物ソ ナー
を行 うコ ウ モ リは,
放 射 する超 音 波 音 声 (パ ル ス)に よっ て 2 種類 に大 別さ れ る。
周波数定 常部 分 (constant frequency,
CF) の後に短い下降周波数変調 部 分 (frequency modulated,
FM ) を もつ パ ル ス を用い るCF−
FM コ ウモ リと下降周波 数 変 調音 をパ ル ス として用い るFM コウモ リで あ る(力丸・
菅,
1990,
Figure 3)。
こ こ で は,
CF−
FM コ ウモ リの特 徴につ い て だ け述べ,
FM コ ウモ リにつ いて の説明は別の機 会 に譲る。
CF−
FM コ ウモ リの パル ス は,
ヒ トと同 様に声 帯 振 動 に より生成さ れ, 周 波 数の構造は倍 音 (調波)構 造を し てい る。
音 源で は基本周波 数が最強で高次 倍 音になるに つれてパ ワー
が盛 衰する が,
放射さ れ るパ ル ス に おい ては
,
第2倍 音が最強で あ る (Figure 4,
Hartley& Suthers,
1990)
.
一
般 的に,
こ の特徴はCF−
FM コ ウモ リ に共 通で あ る。力丸lr 知覚科学』へ ようこそ
一
ヒ ト心 理物理学と動物 神経 行動学と の統 合一
81 A 1、
0 86
4
2 0 0 0 0 9Eo の =
a
の ohoO O.
0尋 十 十
S1 S4糶 鶲 ■
B 1,
0 2 田 o 塁a
部 hoO 8(
◎ 4 2 0 0 0 0 2° 3° 4° 5° 6° 7°一一一
■
NeiSe intenSity(dB SPL} No.
Go Go S5 O.
0 2可
0
1
2
」
コ ω ω Φ」
ユ η 匚 コ O 切 Φ ≧ 駑一
〇 匡 C 1、
O 86
4
2 α α α α o 駕
」
Φ 切8
ユ の o 」 oO 「τ「「π
0,
0 St喜
S2 蠶m−
mam一
S3 †No notch Around tone Above tone Notch condition
Hgure 2
.
Auditory induction in Mongolian gerbils.
A:Effects of the sound pressure
leve
艮on the auditory in−
duction
.
The higher the level of noise inserted in silentintervals
,
thehigher
the occurrence ofthe auditory in−
duction (perceived continuity )(Sl)
.
When the ent 童reinterrupted signal sound was masked by no正se
,
no au−
ditory induction was generated(no continuity )
.
(S4).
B:Animals showed Go responses f‘)r continuous s 童
g−
nals
,
while No−go
responses for interrupted signals,
C:When the
frequency
range of noiseburst
containedthe signal frequency
,
continuity was indnced (Sl,
S3).
However
,
if the rloise did not covered the signal fre−
quency
,
continuity was not perceived(S2) (fromKobayasi et aL,2012)
,
CF−
FM コウモ リ の生 物ソ ナー
の神 経 機 構に関 する基 礎 的な解説は,
拙 著を参 考に して頂き たい (力 丸・
菅,
1gge)。 CF−
FM コ ウモ リを代表 す る 種の ヒゲコウモ リ(Pteronotusparnelii
parnelii
)で は,
聴 覚 野の機 能 局 在 地 図が神経生 理学 的アプロー
チ に より早 くか ら作 成され ていた が (Suga,
1984),
行動学 的ア ブn一
チ に よ る局所の機 能 確認は な かっ た。
機 能 局 在 地 図に関 する行 動 学 的 確 認は,
オペ ラン ト条 件付けした ヒゲコ ウモ リの聴 覚 野に抑 制 性 神 経 伝 達 物 質 GABA−
A の作動薬ム シ モ ル (muscimol )を局 所 投 与 する弁 別実験で初めて確認さ れ た (Riqutmaroux,
Gaioni,
& Suga,
1991,
1992).
2.
Ll 匕ゲコ ウモ リ の生物ソナー
音と聴 覚 野 CF−
FM コ ウモ リの代 表で あ るヒゲコ ウモ リ の放 射パ oo 50 00 甜 21
1
[
N 工 X】
》 O 匚 Φコ
σ O」
Lo 50 00 50[
N エ ヱ一
》 O 匚 Φ コ σ Φ」
LO15105 司 mem 苅 0 52
1
0
1
2
3 Φ
」
D の 09」
α U 匚 コ O ω Φ 冫 ; 偲一
Φ 江 B’
丶.
吐
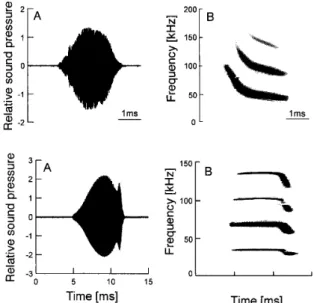
墨 Time[ms ]Figure 3
,
Tn)es ofbiosonar
pulse ofbats
.
Tbp:Pulse ofFM
bat
(lapanese
house
bat
).
Bottom:Pulse of CF−
FMbat
(Taiwanese
leaf
−
nosedbat
).
A;Temporal change inamplitude
.
B:Temporal change in fr巳quenGy structure.
一
60dS o le 20 ” mt{ms)
Figure 4
,
Frequency characteristics of emi はed CF−
FMpulse
(ωp
)and CF−
FMpulse
atglottis
(bottom).
Le良:Relative power spectra for CF Gomponents
.
Rヱght:Temporal frequenoy structure (sound spectrogram )
.
The second harrnonics are the mest intense for em 孟tted
pulses
,
while the fundamental frequency componentsare
the
strongest atthe
glottis(modifiedfrom
Hartley&Suthers
,
1990).
ル ス は,
周波 数一
定 (CF,
constant frequency)成分と後続 の周 波 数 変 調 (FM,
frequency modulated )成 分か ら なる倍 音構 造を し てい る。 パル スの音源で は基本周波数成 分が 最 強で あるが,
放 射パ ル ス中で は,
第2倍 音 成 分が最 強 である、
距 離に応 じた遅 延 時 間 (エ コー
遅 延 )の後に戻っ て来るエ コー
の周 波 数は,
速 度に応じ た ドップ ラー ・
シ フ トに より上 昇す る (Figure 5 右)。 彼らの聴 覚 野は,
神 経 生 理学的 研究の結果,
聴覚野の機 能地図が報 告さ れ て い る(Figure 5左 )。
その うちの幾つ か を紹 介 する。82 基礎心理学 研 究 第32巻 第1号
2
.
1.
2一
次 聴 覚 野とDSCF 領 域ヒゲコ ウモ リの
一
次聴覚 野は前 後に広 が り,
後方か ら 前 方に かけて低 周 波 数か ら高 周 波 数に変 化 する周 波 数 軸 (トノ トピー
)が存 在す る。
ほぼ円形の DSCF (Doppler−
shifted constant frequen⊂y
,
ドッ プ ラー・
シ フ ト し た定常周波数) 領域は
,一
次聴覚野の一
部で,一
次聴覚野の中 央 部に位 置 する (Figure 5左 )。
ドップラー ・
シフ ト した 約 60kHz か ら約 63kHz
の範囲のエ コー
の第 2 倍音に鋭く 反応するニ ュー
ロ ンが集結し て い る。 円形領域の中心部 は約60kHz に反 応し, 円周 部は約63kHz
に良 く反 応す る ように周 波 数 軸が放 射 状に配 置さ れてい る。
換 言 する と,
同 じ 周波 数に同 調 す るニュー
ロ ンが同心円 状に配 列 されて い る。 60kHz よ り も低い周 波 数 はDSCF 領 域 よ り 尾側,
63kHz よ り も高い周波 数はDSCF 領 域よ り吻 側に 表示さ れてい る (力 丸・
菅,
1990)。 2,
L3 FM−
FM 領 域FM
−
FM 領域は,
DSCF 領 域の背 側に位置 し (Figure 5 左),
パ ル スのFM 基本周波数成分 (FMI )とエ コー
のFM 第2,
3,
4倍 音 成 分 (PMI,
FM2,
FM3 )の特定のエ コー
遅 延 で の組み合わせ に鋭 く反 応 するニ ュー
ロン が集結 して いる (組み合わせ感 受 性ニ ュー
ロ ン)。
前 方か ら後 方に向かっ て エ コー
の遅 延 軸 (O.
4’
−
18 ms )が存 在す る。
この エ コー
遅 延は3m 以内の距離 軸に相 当する (力丸・
菅,
1990)。 2.
L4 前帯状回の運 動地図前 帯 状回 (ACg )を電気 刺激す ると ヒゲコ ウモ リ はエ コ
ー
ロ ケー
シ ョ ン用の パ ル スを放 射すると い う。 さ ら に,
刺 激 する部 位に よっ て,
放 射パ ル スの CF周 波 数が 異 な り,
周 波 数 地 図 (運 動 地 図)が存 在 す ると報 告 され た (Gooler& σNeil,
1987)。
しか し,
Riquimarouxらの実 験に より,
ACg を削 除 して もパ ル ス 放 射に影 響せず,
ドッ プ ラ
ー ・
シフ ト補 償 も消 失し ない こ と が 報告さ れた。 す な わ ち
,
周波 数地図が存 在し て もACg に よっ て,
放射パ ル スの周波数がコ ン トロ
ー
ル さ れ てい る わけで は ない こ と が 判っ た (Riquimaroux , Gaieni,& Suga,1989)。
こ の よ う に
,
トポグ ラフ ィ ッ ク な地 図が存在 して も,
そ の地図は必ずしも特 定の場 所が特定の機能を持つ 機能地 図と は限 らない の である。
そ の部 位の神 経 活 動 を止め,
機 能が消 失 することを確 認 する必 要がある。
2.
1.
5DSCF 領 域とFM−
FM 領 域の機 能局在に関す る 行 動 学 的 研 究 ヒゲ コウモ リのDSCF 領 域とFM−
FM 領 域 と呼 ばれ る 領域には,
前述し た ように,
そ れ ぞ れエ コー
の第 2 倍音 の地 図とエ コー
遅 延 時 間の地 図が存 在 する。
エ コー
の第 2 倍音の周波数 弁別訓練とエ コー
遅延 時間の弁別訓 練を Go−
Nogo タス ク で行っ た ヒゲコ ウモ リの聴 覚 野 内のこ れ ら2 領域の神 経活動を抑 制性 伝達物 質 GABA−
A の作動 薬で あるム シ モル を脳 表か ら投 与して止め る実 験 を行っ た (Riquimaroux et al.
,
1991,
1992)。
ム シ モ ル をDSCF 領 域に投 与し て神 経 活 動を 阻害す る と,
微 細な 周波 数 弁 別 能 (△flf
=
0.
05 kHz161.
OkHz=
0.
008)は な くな る が,
粗大 な周 波 数 弁 別 (△f/f=
2kHz16LO kHz=
O.
03)は影 響を受 けない こ とが判明し た。 ま た,
隣接し たFM−
FM 領域に 投 与して も,
周波数弁 別能には影 響を全 く 与えない。
ム シ モ ル をFM−
FM 領 域に投 与して神 経 活 動を阻 害す る F 120 ge 主 60 よ 30 o Pulse EchoI
、
「
_ .
L
.
「
「
一
.
.
層
H’“
5i
’;
一 一
X
丶
FM・ H3ず
芥
陣
・ H ・噺
丶
FM2 DS Hr戸
…
\ FM1畷一
一
レ Delay − 一 〇 10 20 30 msFigure 5
.
Biosonar sounds used by the mustached bat,
one of GF−
FM bats(right),
and neurophysiologically ft)und subareasin the auditory cortex (lefの
.
Right:Solid lines showpulses
,
dotted
lines
indicate echoes.
Pulses and echoes are harmonicallystructured
.
From the 且rst through 4thharmonics
areilustrated
(H1−
H4 ).
Echofrequency
ishigher
than pulse frequencアbecause ofthe Doppler
−
shif ヒ(DS).
Echo shows adelay
due
to distance to targets(Delay).
A pulse alld an echo consist of aconstant 丘equency component (CF)and a frequency modulated component (FM )
.
The second harmonic is the most in−
tense
.
Left:Left side view of entirebrain
and the magnified auditory cortical area,
The
round−
shaped DSCF area islocated
力 丸:『知 覚 科 学』 へ よ うこそ
一
ヒ ト心 理 物 理 学と動 物 神 経 行 動 学と の統 合一
83 と,
微細 な 時 間弁 別 能 (△t!t=
1 ms /4 ms=
0.
25)はな くな るが,
粗 大な時 間 弁 別 (△t!t=
36 ms14 ms=
9.
0)は影 響を 受け ない。
さ ら に, 隣接し たDSCF 領域に投 与して も,
時間弁別 能に は全く影響はない こ とも判っ た (Figure 6)。
2.
2 機 能 局 在 研 究の落とし穴一
次 聴 覚野の機 能を解明 す る た め に,
一
次 聴 覚野に削 除 や損 傷 を 与え,
術 前と術 後の行 動 比 較に よ り,
その機 能 を検 討 する方 法が行 わ れてきた。一
次 聴 覚 野に は トノ ト ピー
(周波数の場所 表示)が存在す る。一
次聴 覚 野に 周 波 数 弁 別 機 能があるか否か を行 動 学 的に試 験 するた め に,
周 波 数 弁 別 訓 練を受け た動 物を用い て,
術 後に 周波 数 弁別能を テス トする試み が多く行わ れ て き た。 例 え ば,
1kHz と2kHz の弁 別 訓 練をし た動物 (例え ばネコ ) では,一
次 聴 覚 野を削 除して も弁 別 能は消 失し ない と報 告さ れ てい る。 この事実を もっ て,
一
次 聴 覚 野は周 波 数 弁 別の機 能はない と結 論づけて い る研 究 者が多い。
はた して, こ の論 理は正しい のであろうか。
ムシモル を局 所 的に投 与し て神 経 活 動を阻 害 する と,
微 細な周 波 数 弁 別 能 (△ftf=
O.
05 kHz16LO kHz= 0.
008)はな くなるが,
粗 大 な 周 波 数 弁 別 (△flf=
2 kHzt61.
0 kHz=
0.
03)は影 響 を 受 け ない。
前 出の例で説 明す る と,
1kHz と1.
008kHz
の弁 別に は一
次聴 覚野が必 要である が,
lkHz と1.
03kHz
の 弁 別に は、一
次 聴 覚 野は不 要とい うこ と になる。
し た が っ て, 粗大な弁 別は一
次 聴 覚 野で はな く, 皮 質 下で実 行さ れ てい る と考え ら れ る。 周波数の表 現と周 波 数の弁 別とは,
区 別 す るべ きで あ る。 2.
3 非侵襲的方法によ る機能局 在 計測の問 題 点 前 述の動 物実験で の機 能 局 在で注 意を喚 起 した こ と は,
ヒ ト の機 能 局 在 研 究にも あて は まる。
す なわち,
特 定の刺 激に よっ て,
脳内の特定部位が賦活さ れ てい る こ とがfMRI な どで 示さ れ たとして も機能局 在を証 明し た こ と に はな らない。 当 該 部 位の機 能が 出血 や 梗 塞で阻 害 されたり,
手 術に よっ て削 除さ れ た後に,
機 能が失 われ るこ とが確 認さ れ れ ば,
機 能 局 在が確定する。 てんか ん 患 者の右 側Heschl回 を 削 除 する と時 間ピッ チの弁 別が 困 難に な る とい う術 前・
術 後の比 較 実 験は,
説 得 力が あ る (Zat。rre,
1988)。 3.
効 率 よ く聴 き取る仕 組み 3.
1 テ レマ イク 搭載による飛行中の生物ソ ナー
パ ル ス とエ コー
の音響 計測 コ ウモ リ は,
飛 行 中に超 音 波パ ル ス を放 射 し,
戻っ て くるエ コー
を聴い て,
獲 物や障 害 物につ い て の情 報 を得 る。 飛 行 中のコ ウモ リが どの ような音を聴いてい るかを 計 測 するには,
コウモ リにマ イクロ フ ォン を搭 載 しなけ ればな ら ない。
私の グルー
プは,
1990年 代の末にコウモ リに搭 載可 能な小 型 軽量ワイ ヤレ ス の超音 波マ イ クロ フ ォ ン (Telemike,
テ レマイク)を開 発し,
コ ウモ リか ら放 射さ れ る生物ソ ナー
用パ ル ス と戻っ て来るエ コー
を 頭 上で記 録す る研 究を開始し た (Riquimaroux & Watanabe,
Σ 吟 差 羽 =8
仰 O 二 8 ヒ 8 9 ち E一
9,
= LOm 」00 80 60 40 100828°
2
璽
6・ τempo旧
ldiscrimination Pre O 3 6 9 24 40
网
2010 P陶 036924 100 80 60 40 10Q 80 60 40 BFrequency tiisoriminarion Pfe O 3 6 9窿
2り ヱ12005
世 Pre O 3 6 9 24阿o口 「safler mu3c ■moI
Figure 6
.
Behaviorally con血rmedfunctions
of DSCF and FM−
FM areas.
Muscimol,
agonist of inhibitory neuretransmitterGABA
.
A,
was applied to DSCF (bottom)or FM−
FM areas (top).
Scoresfrom
temporal (left
)and frequency discrimina−
tion tests (right)before and after the drug application are shown
、
The ordinate represents percent correct,
while the84 基 礎 心 理 学 研 究 第32巻 第1号
B
蕁響驚Tel
.
,
鷺Figure 7
.
Horseshoe bat and Telemike.
A:Ahorseshoe bat flying in the flight chamben B:Ahorseshoebat
with 丁demike
mounted
,
which weighs about O.
5 g including an antenna and a battery.
C:Telemike.
Prequency characteristics are roughlyflat
betwee
20 and lOO kHz.
2000)
。
開発 当初の テレ マ イク は約3g であっ た た め,
比 較 的 大 型の テ ラソカ グ ラコ ウモ リ に搭 載し た。
そ の後の 改 良で重 量が lg 未満とな り,
体 車5g程 度の小 型の コ ウ モ リ にも搭 載 可 能となっ た (Figure 7)。 テレマ イク の出 現で,
そ れ ま で わか ら なか っ た多 くの事 実が明 らか に なっ た。 その 中か ら,
い くつ か の事 柄につ い て論 説す るv 3.
2 ドップラー ・
シ フ ト補償とエ コー
振 幅 補 償の目的CF
−
PM コ ウモ リで あ るヒゲコ ウモ リ (Pteronotusparnelii
parnelii
),
キク ガ シ ラコ ウモ リ (Rhinotophu∫.
ferrumequinum
niρpon),
テラ ソ カグラコ ウモ リ(Hipposideros terasensis)の放 射パ ル ス は倍 音 構 造 を な して お り
,
声 道 特 性に よ り第2 倍音が最 強で あ る。 音源に近い声 道内で計測す ると勿
論
,
基 本 周 波 数が最 強であ る(Figure 2)。 CF−
FM コ ウモ リ で は,
ドップラー・
シフ ト補 償 (Doppler−
shift compensation )とい う興 味 深い 行 動が知ら れてい る
。
彼らは,
大 きな反 射 物か ら戻っ て来るエ コー
の第2倍 音の周 波 数が一
定 と なる よ うに,
相対速 度に応じて放 射す るパル ス の周波 数 を変 化 させ るの である。 エ コー
の周 波 数は飛 行 速 度に よっ て ドップ ラー ・
シ フ トす るの で ある が,
パ ル ス の周 波数を調節し て速度に よ る 周波数の シフ トを な く し,
常 にエ コー
の 周波数を一
定に保つ ので あ る。
これ ま で は,
テレ マ イク が なか っ たの で,
研 究に は大 き な ブ ランコ が よ く使わ れ た。
ブランコE
でもコ ウモ リはパ ル スを放 射 する。 コ ウモ リ を乗せ たブランヌ を揺らすと,
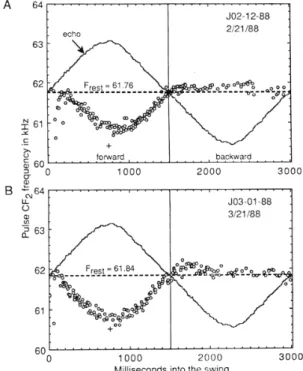
ブラン コ A 64 63 62 echoE0
1 0 4 ビ N エ エ ≡ む 5コ
σ
よ £ o 墜 コ 匹「
63 B緯
響
D
十 torvva:
d JD2.
12・
882 !21/88 鍔 謡蟾
蹴 贈 b臼ckward 1000 2000 30eo 62 61一
一
罵
60 0 JO3・
01・
883 /2VB8゜
醜
球 転 ・o 1000 20DO
MillisecondS intO the SWIng
30eo
Figure 8
,
Doppler−
shift compensation for echo CF2 丘e−
quellcy measured
from
mustachedbats
on a pendu−
lum
.
Left half:Forward swing,
Right half:backward
swing
.
Every individual (A,
B)shows Doppler−
shiftcompensation only for the forward swings but not fbr
the
backward
swings.
Open cir⊂les represent pulse CF2丘equencies measured
,
Solid hnes indi¢ate veloc 孟ty ofpendulum
.
Frest illustrates CF2 frequency of pulses at力 丸:Ii知 覚 科 学 へ よ う こ そ
一
ヒ ト心 理物理学と動物 神経 行動 学と の統 合一
85B
冖
70.
OlM69.
5 よ8
69.
。 雪 σ68.
59 哩 68・
Oo67.
56 ア.
O EGhoes夢 噸”
{ 鴨’
η
ノ
. 隔 .二
llぎ
_ ..宀 ’一
一
一
一
.
.
.
1−.
.
.
.
−
L−
.
.
.
L.
.一
.
L−
.
一
.
一
一
.
亅 5 0 Target distance[m]C
GO 冨 o 且_
t・
10 巳 CO.
20 ゆ3 −
3。 錏一
4a−
50・
60 5 Pulses■
, 幽 ■圏 .■ ■●■
v.、駈
’
「
L
品
ご
・ ’ 学噴
鸞
●
句
.
『
1_
.
、
1− 、
.
.
_
L_
._
.
_
[.
.
.
_、
亅 4 3 2 1 0 Target distance{m】Figure 9
.
Dopper−
shift (A,
B)alld e⊂h{, amplitude c〔〕111pelコsations (C)〔,f a flying horseshoc bat shown through Tdemke.
Frequencies(B)and amplitudes of echoes (C)are kept almost ⊂onstmt
,
A:Abscissa shows Ume,
whilc ordinatc indicatesfrequency
.
B:The dot亡ed 【ine illustrates pulse CF2 丘equency at rest (from Hiryu el aL、
2008).
68
.
0主
M 67.
5 玄867
.
o 雪866
.
5 虚f66 .
oo 65.
5 BatA;;
∵
∵
7
ど
蟻
礎
醗
es 疎 譲 袰.
○ 剛 wa 飼.
壼冊
睡・
ce 十liag 攤 出 〔e 帥 m 日ted) ρ ωse 69.
5黛 邑 69
・
o 台868
・
5言
・8.
・ し Lpa 6ア.
50 67.
0¶
1.
5 4.
0 心.
5 T董rne to captureIs
璽 3.
o BatB 熱 隷 灘 鶏鷲
灘嚇 ・‘ ・ ・ …。
謁
騨
岬 嬲_・羈.
藤
.
;繍tP
■ 1「衄 w魍潟 睡9楓w滷随 鑼 mothOS 豊1 旭d 繭
一
2.
0一
1.
5_
1.
0_
O.
5 rlme 牝o captureIs
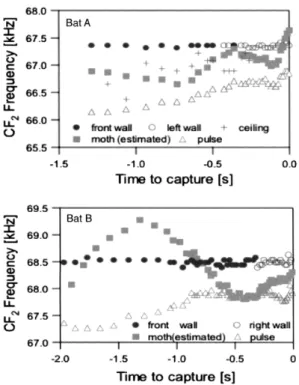
肇 0Figure lO
.
Pulse and e⊂ho
frequencies
ofh {〕rseshc)cbats
durlng flight f6r capturing a
flying
moth recorded viaonboard Telemike
.
Tbp panel:Bat A firs.
t冂ew towardthe front wall
,
then turned left丘)r capturing a m 〔}th.
Bottom pane】:Bat B directed toward the front wall
,
then turned right to
get
a moth.
Data indicate that batsconduct Doppler
−
shifL compensation fbr walls locatedhl the
night
directien
,
but
notfor
targeted nloths(
from
Mantani et al.
,
2012),
の速 度はIE弦 波 状に変 化す る の で
,
止[ni
の壁か ら のエコ
ー
の周波数は ドッ ブ ラー ・
シ フ ト し て IE弦 波状に変化 す る。
こ の ドッ プラー ・
シ フ ト を キャ ン セル す る よ う に,
コウモ リ は放 射パ ル スの周 波 数 をL
ドさせ るの であ る(Figure 8,
Gaioni,
Riquimaroux,
&Suga,
1990)。
た だ し,
プ ランコ がター
ゲッ トか ら遠ざか り,
後 方に振れ るとき に は,
周 波 数 補 償は行わなL 飛 行「ti
の CF−
FM コ ウモ リの頭1
ニテレ マ イク でパ ル ス とエ コー
の周波 数を計 測す る とブラン コ11
と同 じ よ う にエ コー
の第2倍1
’
1
: の周波 数 が一
・
・
定にな る よ うにパ ル ス の周波 数をk
トさせ る こ とが わか っ た (Figure gA、
B,
Hiryu,
Shiori,
Hosekawa,
Riquima−
roux,
&Watanabe,
2008)o 以前は,
ドッ フ ラー ・
シフ ト補 償は,
獲 物と の相 対 速 度を計測す る一.
r
一
段と考え ら れて いた。 し か し,
テ レマ イ クtOf)Fl
い た研 究で,
こ の考え方は覆さ れ た。
コウモ リ は,
搬 送 波の周 波 数を安 定さ せ る た め に大き な壁の よ う な物 体か らの強いエ コー
に対し て ドップ ラー・
シ フ ト補 償を行い,
獲 物に対しては ドップ ラー ・
シ フ ト補 償 を 行 わ ない こ と が 判IVjし た (Figure 10,
Mantani et aL,
2012)。 コ ウモ リの獲 物 とな る昆 虫のサ イズは小 さい の で,
獲 物 か ら のエ コー
は四方に散 乱し,
コ ウモ リに到達す るエ コー
の振llllllは極め て小さ く な り,
信頼性のあ る信1}には な りに くい。
一
方,
周 波 数の固 定さ れ た持 続 時 間の長い 強 烈なCFパ ル ス は,
遠 方の 大き な 反射 物 体か ら な ら聴 き取 り1.
1∫能なエ コー
と して戻っ て くる と疹え ら れ る。
人 き な壁や地面か ら は,
周 波 数 が 定 常で振1幅の人 きい安定86 基 礎心 理学 研 究 第32巻 第1号 し たエ コ
ー
がコ ウモ リに戻っ て来る。
し た がっ て,
大 き な 壁 や 地 面とコ ウモ リの間に獲 物が存 在 す れ ば,
エ コー
の周 波 数 と振 幅が獲 物の羽 ばたきによっ て変 化 するの で, 周波 数が定 常で振幅の大きい安定し たエ コー
は,
搬 送 波の役割を果た す こと が わ か る。 距離の離れ た獲物か らの直 接エ コ.
一
は聴き取る こ と が で き な くて も,
安定し た搬 送 波と し て のエ コー
を利 用す れば,
獲 物の羽 ばた き 情 報 (平 均 周期 26.
5ms )を得るこ とが可能に なる。 獲 物の羽ば た き情 報の載っ たエ コー
か ら搬 送 波の周 波 数 を 取 り除けば,
獲 物の羽 ばた き情報だ け が残る。 こ れ が,
コ ウモ リが ドップラー ・
シ フ ト補 償を用いる理由と考え て いる。
実は,
私が テレ マ イ クを実 験に導 入し た最 大の 理 由の ひとつ はこ の仮 説 を確 認す る た めで あっ た。
さ ら に,
ドッ プラー ・
シ フ ト補償は時分割 処 理で実 行 さ れて いるこ ともわか っ て きた。 す なわち,
複 数の壁が 存 在す るときに は,
前 方と側 方の壁に対して,
時 分 割で 交互 に ドップラー ・
シ フ ト補 償を実行 して い るの で あ る。
正 面の壁に対する速 度成 分は大きい の で,
ドップ ラー ・
シ フ ト補 償 量も大き く,
側 方の壁に対 する速 度 成 分は比 較 的 小 さいの で,
ドップラー ・
シフ ト補償量 も小 さい。
こ の結 果,
パ ル ス の周 波 数は ジ グ ザ グになる の で ある。 大 き な 壁に近づ くとエ コー
の 音 圧レ ベ ルが上 昇する。
コウモ リ は壁か ら戻るエ コー
の音圧レベ ルが一
淀 になる ように放射パ ル スの音圧レ ベ ル を変 化 させ る (Figure 9C,
Hiryu et aL,
2008)。
こ れ をエ コー
振 幅 補 償とよん で い る。
ドップラー ・
シ フ ト補 償とエ コー
振 幅 補 償を組み合わ せ ると,
戻っ て来るエ コー
の周 波 数も振 幅も一
定となり,
エ コー
に含ま れ る信 号 成分を抽 出する のが,
非 常に簡 単 にな ると考 え られる。
神 経 系の演 算 速 度は極め て遅い の で,
リアル タ イム 処理 を す る仕 事の負荷を軽 減する必要 がある。 こ の目 的のために,
ドップラー ・
シ フ ト補 償と エ コー
振 幅 補 償は有 効で あろう。
3.
3 混 信に対 する対 策・
戦 略 弱いエ コー
か ら信 号を抽 出する際に,
エ コー
の重な り に よ る混 信は大き な問 題となる。 混 信に は2種 類 ある。
自分 自身のエ コー
同志が重 なっ て しま う場 合と自分のエ コー
と近 く を 飛 ぶ他の コ ウモ リのパ ル スやエ コー
が重な る場合で ある。 樹木の密 度の高い樹 林などで は,
1発の パ ル ス に対して多 くのエ コー
が数 珠つ なが りとなっ て 戻っ て く るので,
次に放射されたパ ル ス に対 するエ コー
が重なっ て し ま う。 自分のパ ル ス に対 するエ コー
で ある の で,
重 なっ て し まうと対 象 物につ い て の情報が得 られ な く な り,
大 変 都 合が 悪い こ と に な る。 自分 自身のエ コー
が 重 な る 場 合,
FM コ ウモ リの実 験 結 果で は,
エ コー
が重 な ら ない よ うにパ ル ス の終 端 周 波 数 を ず ら して エ コー
の重 な りを 防 ぐ行 動 を実 行 するこ と が記 録されてい る (Hiryu
,
Bates,
Simmons,
& Riquimaroux,
2010)。 複数 の CF−
FM コ ウモ リが洞窟 内の ように狭い空 間を同 時に 飛 行 する場 合,
他の コ ウモ リのパ ル ス やエ コー
が時 間 的 に も周 波 数 的に も重なっ て し ま う。 こ の よ う な混信 状況 下で も,
彼らは壁に衝 突する こ とも な く,
他個体と の衝 突 もなく, ス ムー
ズに飛行 する。 自分のエ コー
だけを抽 出す る に はどの ような手 段を用い るのであ ろ うか。 常識 的に推 測する と,
自分のエ コー
と他のコ ウ モ リのパ ル ス やエ コー
が重な ら ない よ う に自分のパ ル スの周 波 数を シ フ トす る手 法が考え ら れる。
テ レマ イクを搭 載し た2 頭 の コ ウモ リを 同 時に飛 行 させ ると,
興 味 深い行 動 をと る こと が分か っ た。
彼 らは,
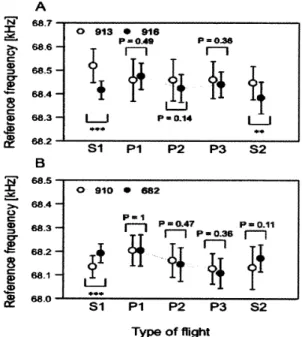
相 手の周 波 数と自 分の周 波 数 の間 隔 を 広 げて重 な ら ない よ うにす るの で は な く,
逆に 互い の周 波 数の接 近させ る の で ある。 相 千が周 波 数 を 下 げる と,
自 分も下 げる の で ある (Figure l l,
Furusawa,
Hiryu, KQbayasi
,
&Riquimaroux,
2012)。
この逆 説的行 動に より
,
どの ようにエ コー
情報を抽 出する か を解明 する A7 β 石 鴻 溜 2 68 63 册 68 68 68冖
N護
ぎ
。 コ冨
§
鬟
罍 匡 B β 4 3 2 」 ρ 銘 鮑 舶 68 68 68冖
翌
= 。§
聾
§
叢
匡 S帽 Pl P2 P3 S2 $l PI P2 P3 S2 Type of 竈igh象Figure l L Paradoxical strateg γ
for
jammed
situations.
Comparison of echo CF2 frequencies between solo
flights and paired flights
.
A:Bat#913and Bat#,16 are
paired
.
B:Bat#910and Bat#682 are paired.
First,
solo且ights were conducted (
S1
),
thenpaired
flights
wereexam [ned three times (P1
,
P2,
P3 ),
and solo fhghtswere tested once again (S2)
.
Data indi¢ate that batstend to shift their echo frequencies closer for paired
flights that those
for
solo flights.
* *P
<0.
Ol,
** *P〈
力 丸:『知 覚 科 学』へ よ うこそ
一
ヒ ト心 理 物 理 学と動 物 神 経 行 動 学と の統 合一
87 の は,
大 変興味深い研究対 象である。 複数の コウモ リの パ ル スやエ コー
の周 波 数が接 近する と ウナ リ を生 ずる。
ヒ トの心 理 物 理 (心 理 音 響 ) 分 野で は, マ ス ク す る音 (マ ス カー
)に ウ ナ リ が存 在する と信 号が検 知し や す く な る 現象が知ら れて い る。 コ ウモ リがエ コー
の周 波 数を 敢 えて接 近 させ るの は,
ウナ リを 利 用 して信 号 検 知 を容 易に す ること が 目的か も しれ ない。 両耳性マ ス キングレベ ル 差 (Binaural masking level difference)
,
連 続 聴 錯 覚,
さらに は音 脈 分 凝 (Auditory streaming ) もコ ウモ リの信 号抽 出に関連して い る可能性がある (力 丸
,
1994)。 ヒ トの心 理音響学の研究成 果がコ ウモ リの生 物ソナー
の処 理 機 構に役 立ち,
今 後の通 信 技 術へ の有 効 応 用も考え ら れ る の で, 今後の発 展が待ち遠しい 。 ま と め・
中枢 神経系は,
劣化 雑音音声 知覚や連続 聴錯覚に代表 さ れ るよ うに,
都 合 良 く刺 激 を創 り変 える仕 組みを 備 え,
迅 速な リ アル タ イ ム処理を実行し て知覚してい る。・
脳 内特定部位の機能局在を確認する に は,
特定刺 激に 対 する反 応 を 測 定 する だ けで は な く,
特 定 部 位の活 動 が失われた場 合に機 能が消 失 することを 確 認 する必 要 がある。
・
コ ウモ リは,
強い安定し たエ コー
に信 号を載せ て運び (搬 送 波 ),
信 号 抽 出を して い る と考 えられる。
これ は, ラジ オ電 波の検 波 方 式に似て い る。
・
コ ウモ リは混 信 対 策と し て,
搬 送 波と な るエ コー
の 周 波 数を離 して 混 信を回 避 する の で は な く,
彑い のエ コー
の周 波 数 を接 近 させ る こ と によっ て,
ノ イ ズ・
リ ダ クシ ョ ンを実 行し,
信 号 抽 出を有 利に してい ると考 えられ る。 引用文 献Furusawa
,
Y,
Hiryu,
S.
,
Kobayasi,
K,
,
& Riquimaroux,
H.
(2012)
.
Convergence of reference frequen¢ies by multipleCF
−
FM bats (Rhinolophusferrumequinum
nippon )during
paired
且ights evaluated with onboard microphones・
Journal
ofComparative Physiotogy A,198,683
−
693.
Gaioni
,
SJ.
,
Riquimaroux,
H,
,
& Suga,
N.
(1990).
BiosQnarbehavior of mustached bats swung on a pendulum prior to
cortical ablation
.
ノournalOfNeuroph
グsiolo9γ,
64,
1801−
1817.
Gooler
,
D.
M,
,
&O’
Neill,
W E (1987).
Topographic represen−
tation of vocal frequency denlonstmted bア microstimula
−
Uon of anterior cubgulate cortex in the e¢holocating bat
,
Pteronotus Parne〃i ParnelJi
.
10urnal
of Compa 厂ativePhγsiologアA
,
161,
283−
294.
Hartle}〜D
.
1.
,
8【Suthers,
R.
A.
(1990).
Sonar pulse rad 正ationand filtering in the mustached
bat,
Pteronotusparnellii
rubiginosus
.
Thelournal
q
广the Acoustical SocietJ/ qプAmerica
,
87,
2756−
2772.
Hiryu, S
.
, Shiori、
Y.
、
Hosokawa,
T.
,
Riquimaroux,
H.
,
8(Wata−
nabe
,
Y (2008).
On−
board telemetry of emitted soundsfrom free
−
flying bats:Compensation for velo ¢ity and dis−
tance stabilizes echo frequency and amplitude
,
Journal
ofComparative Ph)ノsiolog
γA
,
194.
841−
85LHiryu
,
S.
,
Bates,
M.
,
Simmons.
J.
,
& Riquimaroux,
H.
(2010),
FM echolocating
bats
shift frequencies to avoid broadcast−
eChO ambiguity in CIUtter
、
PrOCeedingS
〔ゾ the NatiOnalAごademy 【ゾScience∫ ()
f
the United States(ゾAtnerica,
107,
7048−
7053.
Kobayasi
,
K.
,
Usami,
A.
,
&Riquimaroux,
H.
(2012).
BehaVior−
al evidence
for
auditory induction in a species of rodent :Mongolian
gerbil
(Meriones unguiculatus ).
TheJournal
〔’f
the Acoustical Socieり10fAmerica
,
132,
4063−
4068.
Mantani
,
S.
,
Hir四,
S.
,
Fulioka,
E.
,
Matsuta,
N.
,
Riquimaroux,
H
.
,
& Wata皿abe,
Y (2012),
Echolocatien behavior of the
Japanese
horseshQebat
in pursuit offluttering
prey.
Journal
〔〜プComparative Physiology A
,
198,
741−
751.
Riquimaroux
,
H.
,
Gaioni
,
S.
J
.
,
&Suga,
N ,
(1989).
Effects of
bilateral
ablation of auditory andtor cingulate cortices onbat echolDcation behavior
.
Associationfor
Research inOtolaryngoiogy
,
12,
28.
力 丸 裕
・
菅 乃 武 男 (1990),
コ ウモ リの生 物ソナー
の神 経 機 構
科 学
,
60,
802−
811.
(Riquimar 。ux,
H.
,
& Suga,
N
.
(1990))Riquimaroux, H
.
, Gaioni, S.
J.
,8(Suga, N.
(1991).
Cortical¢omputatienal maps control the auditory perception
・
Science
,
251,
565−
568.
Riquimaroux
,
H,
,
Gaioni,
S.
J
.
,
&Suga,
N.
(1992).
lnactivatienof the DSCF area of the auditory cortex with muscimol
disrupts
frequency
discrimination in the mustached bat,
Journal
()fNeurophysietogy
,
68,
1613−
1623.
力 丸 裕 (1994)
.
音 響・
聴 覚 系の生 理 学 伊 藤 正 男(編 ) 岩 波 講 座 「認 知 科 学」 第3巻 「視 覚と聴 覚」
岩 波 書 店
pp
.
129−
179.
(Riquimaroux,
H.
(1994))Riquimaroux
,
H.
,
& Watanabe,
Y (2000).
Characteristics of
bat
sonar sounds recordedby
a telemetry system aロd
a fb【ed
ground
micrephene,
ProceedingsOf
WESTPRAC VII,
233−
238.
Riquimaroux
,
H.
(2006).
Perception of noise−
vocoded speechsounds :Sentences
,
words,
accents and melodies.
AcousticalScience【隻Technolog7
,
27♪
325−
33LSuga
,
N.
(1983).
The extent to which biosonar information isrepresented in the bat auditory cortex of the mustached
bat.
In G
.
M.
Edelman,
W.
E.
Gall,
& W.
M.
Cowan (Eds.
) Dア襯 而 σ帥 cα50f Neocortical Function,
New York:John
Wiley&
Sons,
pp.
315−
373.
橘 亮輔
・
力 丸 裕 (2005).
劣化 雑 音音声 知覚に関す る脳 内 活 動:functional MRI に よ る研 究 日本 音 響 学会 聴 覚研究 会 資 料
,
35,
67−
72.
(Tachibana,
R.
,
& Riqui−
maroux
,
H.
(2005))Zatorre
,
R.
(1988).
Pitch perception of complex tones and hu−
man temporal