doi 10.2323/jgam.2020.07.002
2021 Applied Microbiology, Molecular and Cellular Biosciences Research Foundation
Full Paper
*Corresponding author: Naotaka Tanaka, Department of Applied Biological Science, Faculty of Agriculture, Kagawa University, 2393 Ikenobe, Miki-cho, Kagawa 761-0795, Japan.
Tel: +81 87-891-3115 Fax: +81 87-891-3021 E-mail: [email protected]
None of the authors of this manuscript has any financial or personal relationship with other people or organizations that could inappropriately influence their work.
Introduction
Glycosylation is a major covalent modification that modulates the structure and function of membrane and other secreted proteins in eukaryotes. In the budding yeast
Saccharomyces cerevisiae, elongation of N-linked
oligosaccharides occurs by extension of an α 1,6-linked mannose backbone onto which mannose residues are at-tached (Ballou and Ballou, 1995). Many glycosylation-defective mutants have been isolated in S. cerevisiae, and genetic selections have resulted in the identification of a large number of genes required for glycosylation (Herscovics and Orlean, 1993). Molecular characteriza-tion of the products of these genes has revealed a number of biochemical activities involved in this process. In con-trast, the synthesis of the N-linked oligosaccharides of S.
pombe is not well understood. Glycoproteins of S. pombe
contain a large amount of galactose in addition to mannose (Manners and Meyer, 1977; Moreno et al., 1991), indicat-ing that S. pombe is equipped with mechanisms for pro-tein galactosylation similar to animal cells. Gemmill and Trimble (1996) demonstrated that the outer chain struc-ture of galactomannan has an α-1,2-linked galactose or pyruvylated galactose attached to a poly-α-1,6-linked mannose backbone, and that the pyruvylated galactose epitope bears remarkable structural resemblance to the
N-The fission yeast gmn2
+gene encodes an ERD1 homologue of
Saccharomyces cerevisiae required for protein glycosylation and
retention of luminal endoplasmic reticulum proteins
(Received March 18, 2020; Accepted July 27, 2020; J-STAGE Advance publication date: February 3, 2021)
Naotaka Tanaka,1,∗ Akinari Kagami,1 Keisuke Hirai,1 Shotaro Suzuki,1 Shiori Matsuura,1 Takamasa Fukunaga,2 Mitsuaki Tabuchi,1 and Kaoru Takegawa2
1 Department of Applied Biological Science, Faculty of Agriculture, Kagawa University,
2393 Ikenobe, Miki-cho, Kagawa 761-0795, Japan
2 Department of Bioscience and Biotechnology, Faculty of Agriculture, Kyushu University,
744 Motooka, Nishi-ku, Fukuoka 819-0395, Japan
The gmn2 mutant of Schizosaccharomyces pombe has previously been shown to exhibit defects in pro-tein glycosylation of N-linked oligosaccharides (Ballou, L. and Ballou, CE., Proc. Natl. Acad. Sci.
USA , 92, 2790–2794 (1995)). Like most
glycosylation-defective mutants, the S. pombe gmn2 mutant was found to be sensitive to hygromycin B, an aminoglycoside antibiotic. As a result of complementation analysis, the gmn2+ gene was
found to be a single open reading frame that en-codes a polypeptide of 373 amino acids consisting of multiple membrane-spanning regions. The Gmn2 protein shares sequence similarity with
Kluyveromyces lactis and Saccharomyces cerevisiae
Erd1 proteins, which are required for retention of luminal endoplasmic reticulum (ER) proteins. Al-though disruption of the gmn2+ gene is not lethal,
the secreted glycoprotein showed a significant glycosylation defect with destabilization of the glycosyltransferase responsible for N-glycan elon-gation. It was also shown that a significant amount of BiP was missorted to the cell surface according to ADEL receptor destabilization. Fluorescent microscopy revealed that the functional Gmn2-EGFP fusion protein is mainly localized in the Golgi membrane.
These results indicate that the Gmn2 protein is required for protein glycosylation and for reten-tion of ER-resident proteins in S. pombe cells.
Key Words: ERD1; glycosylation;
acetylneuraminic acid-linked galactose epitope found in mammalian cells. A major galactosyltransferase has been purified (Chappell and Warren, 1989), and the correspond-ing gene cloned and identified as gma12+ (Chappell et al., 1994). Analysis of the biosynthetic pathway of protein galactosylation has revealed that galactosyltransferase exhibits genetic redundancy (Chappell et al., 1994; Umeda et al., 2000).
Several glycosylation-defective mutants have been iso-lated independently in S. pombe (Ballou and Ballou, 1995; Ballou et al., 1994; Huang and Snider, 1995; Takegawa et al., 1996; Umeda et al., 2000). Ballou et al. (1994) screened for glycosylation-defective mutants that fail to adhere to QAE-Sephadex, and isolated three bearing recessive mu-tations (gmn1-3). One of these mutants, gmn3, was dem-onstrated to have a mutational lesion in the gms1+ gene (Tanaka and Takegawa, 2001). gms1+/gmn3+ encodes a UDP-galactose transporter and cell-surface glycoproteins of gms1∆ cells completely lack galactose residues due to a defect in the ability to transport UDP-galactose from the cytosol to the lumen of the Golgi apparatus (Tabuchi et al., 1997; Takegawa et al., 1996; Tanaka and Takegawa, 2001; Tanaka et al., 2001). Characterization of gmn3/gms1 mutants showed that galactose residues are not essential for growth of S. pombe cells. However, protein galactosylation was found to be required for maintenance of normal cell shape, sexual agglutination, tolerance to-ward various drugs, and non-sexual flocculation (Tanaka et al., 1999, 2001).
Another gmn mutant, gmn2, has been described to ag-glutinate weakly with Bandeiraea simplicifolia lectin, which is specific for terminal α-galactopyranosyl units and which reacts strongly with wild type S. pombe cells (Ballou and Ballou, 1995). The glycosylation-defective phenotype of the gmn2 mutant has since been shown not to be linked to the gms1+ locus. In order to better understand the role of the gmn2+ gene product in protein glycosylation, the
gmn2+ gene was isolated from an S. pombe genomic li-brary. Here we report that the gmn2+ gene product is highly homologous to S. cerevisiae Erd1p, which is involved in oligosaccharide synthesis and retention of ER-resident proteins.
Materials and Methods
Strains, media, and genetic methods. Escherichia coli
XL1-blue (Agilent, Santa Clara, CA) was used for all clon-ing procedures. Wild type S. pombe strains TP4-5A
(h-leu1 ura4-D18 ade6-M210) and TP4-1D (h+ leu1 his2 ura4
ade6-M216) were obtained from Dr. T. Toda (Hiroshima
Univ., Japan). LBP12-5B (h+ gmn2) was kindly provided by C. Ballou (Ballou and Ballou, 1995). NTD-1 (h+ gms1∆
leu1 his2 ura4 ade6-M216) has been described previously
(Tabuchi et al., 1997). Standard rich medium (YES) and synthetic minimal medium (MM) for S. pombe cells were used as described elsewhere (Moreno et al., 1991). A pre-viously described electroporation method (Suga and Hatakeyama, 2001) was used for S. pombe gene disrup-tion and a modified lithium acetate method (Morita and Takegawa, 2004) was used for plasmid transformation. General genetic methods have been described previously (Alfa et al., 1993).
Vectors and plasmid constructs. E. coli plasmids
pBluescript KS (+) and (–) were obtained from Stratagene. The S. pombe vector pAL-KS (+) and pAU-KS (+) were a gift from Dr. C. Shimoda (Osaka City Univ., JP), and pJK210 for integration at ura4+ was kindly provided by Dr. J. Keeney (Johns Hopkins University, USA) (Keeney and Boeke, 1994). S. pombe ura4+ was kindly provided by Dr. P. Russell (Scripps Institute, USA). Plasmid DNA was propagated in E. coli XL1-blue and prepared using Flexi-Prep Kit (GE Healthcare, Chicago, IL) or FastGene Plasmid Mini Kit (NIPPON Genetics, JP). An S. pombe genomic library (pAL-SK (+)) was kindly provided by Dr. K. Okazaki (Kazusa DNA Research Institute, JP).
To tag the C-termini of Mnn9p and Anp1p with enhanced green fluorescent protein (EGFP), the fragments contain-ing the open readcontain-ing frames of mnn9+ and anp1+ genes along with their respective promoter regions were ampli-fied from the S. pombe chromosomal DNA by PCR using PrimeSTAR HS DNA polymerase (Takara) and the fol-lowing gene-specific primers containing added restriction sites (underlined): mnn9+, 5 ′-GTTTTCTCGAG-ACGGAATGACTTAACGCGG-3′ (XhoI) and 5′-GTTTT-GGATCCTCATTATGATGGAACACTA-3′ (BamHI);
anp1+, 5′-GTTTTGTCGACCCAAAGACCGTA-TATGGG-3′ (SalI) and 5′-GTTTTGGATCCTCA-ACATGATCGTCTGCAT-3′ (BamHI); och1+, 5′TA C C G G G C C C C C C C C T C G A G G 5′TAT C G C A A -GTTTTA-3′ (XhoI) and 5′-TAGAATCGCACTCGAG-CAAAACACAGGTGGGCGCT-3′ (XhoI); gmh3+, 5′-
GTTTTGTCGACCGCTAGTCCATTTATAAGAATACG-3′ (XhoI) and 5
′-GTTTTCTGCAG-ATGAACCCTCTCACGATATTC-3′ (PstI); erd2+, 5′-GTTTTCTCGAGTTCCTCCTGTCTTGATTCTTC-3′ (XhoI) and 5′-GTTTTGGATCCTTGAACAGT-TCTACAGGGAAAAAAG-3′ (BamHI); gmn2+, 5 ′-GTTTTCTCGAGGATGGCGATGTAAGTTATGATG-3′ (XhoI) and 5′-GTTTTGGATCCTTA-TCTGGATGTATTGTCGGGATG-3′ (BamHI). Each PCR products were digested with the appropriate restriction enzymes and the digested fragments were cloned into the pAL-EGFP (mnn9+, anp1+, gmh3+) or pAU-EGFP (och1+) or pJK210-EGFP (erd2+, gmn2+) plasmids digested with the same enzymes.
Disruption of the gmn2+ gene. The gmn2+ locus was dis-rupted in wild type S. pombe TP4-1D by replacing an in-ternal gmn2+ gene fragment with the S. pombe ura4+ gene to produce strain AKD10. A 0.6-kb XhoI-HindIII fragment was eliminated from the cloned gmn2+ open reading frame and replaced with a 1.6-kb ura4+ cassette (Grimm et al., 1988). A linearized DNA fragment carrying this disrupted
gmn2+ gene was used to transform a wild type haploid strain, and ura4+ transformants were selected. To confirm that the gmn2+ gene had been disrupted, ura4+ transformants were analyzed by Southern blot to verify correct integration of the deletion constructs.
Detection of S. pombe invertase activity. Invertase
activ-ity was detected by staining as described (Gabriel and Wang, 1969). Briefly, yeast strains were grown to the mid-log phase in YES at 30°C and a 5-ml aliquot was centri-fuged, washed with water, and resuspended in 5 ml of YES
gmn2 encodes an Erd1 69 containing 0.05% glucose. Cells were cultured for 3 h at
30°C, centrifuged, and resuspended in 50 mM Tris-HCl (pH 7.0), 1 mM PMSF, 1 mM dithiothreitol, 2% SDS and 10% glycerol. Acid-washed glass beads were added to the cell suspension, and the cells were disrupted by 4 cycles of vortex mixing for 30 s (total 2 min) followed by hold-ing on ice for 30 s. After centrifugation at 15,000 × g for 10 min at 4°C, the supernatant was resolved by 4% nondenaturing polyacrylamide gel electrophoresis (Laemmli, 1970). The gel was then transferred to a solu-tion of 0.1 M sucrose, 0.1 M sodium acetate (pH 5.0) at 37°C for 3 h, was washed with water, placed in 0.1% 2,3,5-triphenyltetrazolium chloride in 0.5 M NaOH, and boiled to detect red bands.
Acid phosphatase staining. Glycosylation of acid
phos-phatase was analyzed as described (Huang and Snider, 1995), with slight modifications. Cells were grown in 5 ml of MM medium to mid-log phase at 30°C. To induce acid phosphatase, cells were centrifuged, washed, and resuspended in 5 ml of phosphate-free MM media, and incubated for 12 h at 30°C. Cells were then collected by centrifugation, washed once in 62.5 mM Tris-HCl (pH 6.8), and suspended in 240 µl lysis buffer (62.5 mM Tris-HCl (pH 6.8), 1 mM EDTA, 2 mM phenylmethylsulphonyl fluo-ride, 0.1 mM dithiothreitol, and 10% glycerol, pH 6.8). Cell lysates were prepared using 0.5-mm glass beads in a Mini BeadBeater (BioSpec Products). The lysates were recovered and centrifuged at 15,000 × g for 10 min at 4°C, and the supernatant was loaded onto a 6% native poly-acrylamide gel. Electrophoresis and staining of the acid phosphatase activity were performed as described by Schweingruber et al. (1986).
Lectin staining. Horseradish peroxidase-conjugated
pea-nut lectin (HRP-PNA) was used to stain the cell surface as described (Takegawa et al., 1996). Briefly, fission yeast cells were incubated on YPD plates for 4 days at 28°C. The colonies were transferred onto filter paper by replica plating, and the filters were incubated in 10 mM Tris-HCl buffer (pH 7.4), 0.15 M NaCl, and 0.05% Tween 20 at room temperature for 1 h, followed by the addition of per-oxidase-conjugated lectin at room temperature for 1 h with continuous shaking. The peroxidase reaction was per-formed by incubating the filter paper in 0.03% 3,3-diaminobenzidine and 0.003% H2O2 in phosphate-buff-ered saline until spots were visible.
Brefeldin A treatment. Brefeldin A (Sigma, St. Louis)
was stored at 4°C as a 10 mg/ml stock solution in ethanol. Fission yeast cells expressing a GFP fusion protein were grown to mid-log phase in an appropriate medium, to which BFA was added to a final concentration of 100 µg/ ml. Cells were then grown for 45 min prior to microscopic observation.
Fluorescence microscopy. Fission yeast cells expressing
an EGFP fusion protein were grown to mid-log phase in an appropriate medium. EGFP localization was monitored by direct fluorescence. Cells were observed using an Ol-ympus BX-50 fluorescence microscope (OlOl-ympus, Tokyo, Japan), and images were captured with a Sensys Cooled CCD camera using a MetaMorph (Roper Scientific, San Diego, CA).
Western immunoblotting. Yeast cells expressing
GFP-tagged proteins were collected and resuspended in 1 ml of PBS. After the addition of trichloroacetic acid (TCA) to a final concentration of 10%, cells were incubated on ice for 15 min and precipitated by centrifugation. TCA-treated yeast cells were washed twice with 1 ml of ice-cold ac-etone and dried. Total proteins were extracted from the dried cells by beating cells with glass beads (two pulses of 5 min each at 65°C) in Urea-SDS cracking buffer (6 M urea, 1% SDS, 50 mM Tris-HCl, pH 7.5). The total pro-tein concentration of the cell extracts was determined us-ing a micro-BCA protein assay kit (Thermo Scientific). Then, 30 µg of total protein was subjected to immunoblotting analysis, which was performed using af-finity-purified rabbit anti-GFP antiserum (our laboratory stock) or rabbit anti-β Tubulin antibody (BioAcademia, 63-160) and horseradish peroxidase-conjugated anti-rab-bit IgG serum (Cell Signaling Technologies). Signals were detected using Immunostar LD Western blotting detection reagent (FUJIFILM Wako Pure Chemical).
Total secreted BiP was analyzed by immunoblotting using a dot-blot format (Dean, 1994). Wild type and gmn2∆ cells were grown on solid media overlaid with a nitrocel-lulose membrane. During this period, secreted proteins bound to the filters. Filters were removed from the plates and adhering cells were removed from them by washing the filters in a phosphate-buffered saline. The filters were then processed for standard western blotting with antibod-ies against endogenous BiP (Pidoux and Armstrong, 1992), kindly provided by Dr. J. Armstrong (Univ. of Sussex, UK). 1
H NMR spectroscopy. S. pombe were cultured in YES
medium at room temperature and harvested at an early stationary phase. Galactomannans were extracted from the cell pellets as follows. First, the galactomannoproteins were extracted from the cell pellets as described before (Peat and Rees, 1961) by autoclaving each pellet at 125°C for 90 min in a citrate buffer (20 mM citrate-NaOH, pH 7.0) followed by precipitation of glycoproteins using methanol. These precipitates were resuspended in milliQ-water. Deuterium-exchanged polysaccharide-alditol was measured on JEOL JNM GX-400 spectrometer operated at 400 MHz in the Fourier transform mode at 25°C as de-scribed (Takegawa et al., 1996). Chemical shifts were ex-pressed in ppm relative to internal acetone at 2.225 ppm.
Results
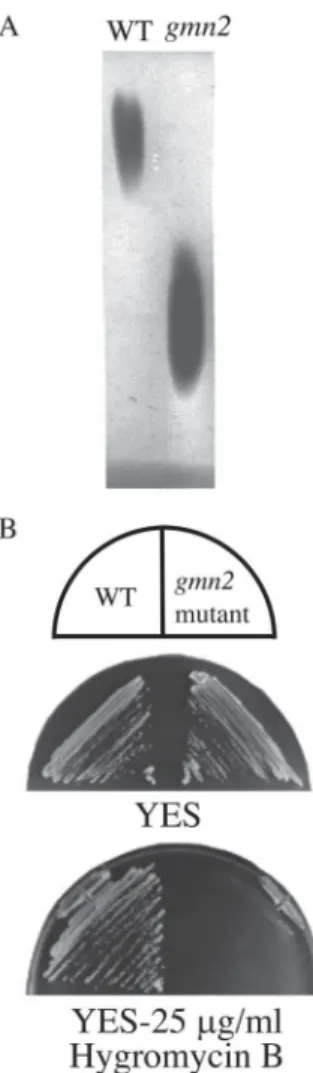
gmn2 mutants are defective in protein glycosylation
The fission yeast gmn2 mutant originally isolated had a reduced amount of cell surface galactose (Ballou and Ballou, 1995; Ballou et al., 1994). To confirm glycosylation of secreted glycoproteins in gmn2 mutants, staining for invertase activity was performed on a nondenaturing polyacrylamide gel (Fig. 1A). S. pombe invertase, encoded by inv1+, has been cloned and sequenced (Tanaka et al., 1998), and is highly glycosylated (67% carbohydrate (Moreno et al., 1985)) with 16 poten-tial N-glycosylation sites (Tanaka et al., 1998). This glyco-protein is a useful marker glyco-protein whose electrophoretic mobility on native gel depends on the size and number of
gmn2 mutant migrated faster than that isolated from wild
type cells (Fig. 1A), while endoglycosidase-treated inver-tase from both mutant and wild type had the same mobil-ity (data not shown). This result suggests that glycosylation of invertase in gmn2 cells is reduced relative to wild type. In S. cerevisiae, a number of glycosylation mutants (alg,
vrg, mnn and others) are sensitive to aminoglycosides such
as hygromycin B, a therapeutically important class of an-tibiotics (Dean, 1995). These glycosylation mutants ex-hibit a severe glycosylation defect at an early-stage of the Golgi glycosylation pathway. The fission yeast gmn2 mu-tant exhibited significant sensitivity to hygromycin B (Fig. 1B). Other fission yeast glycosylation-defective mutants such as gms1∆ and och1∆ also exhibit sensitivity to hygromycin B (Tabuchi et al., 1997; Tanaka et al., 2001; Yoko-o et al., 2001).
Isolation of gmn2+ gene
The extreme sensitivity of the gmn2 mutant to hygromycin B was used as a selection to clone the gmn2 gene. gmn2 cells were transformed with a pAL-SK (+)
(LEU2)-based S. pombe genomic library, and among ap-proximately 20,000 Leu+ transformants obtained, three clones exhibited wild type resistant to hygromycin B. The three complementing plasmids were found to contain two open reading frames (ORF) based on a S. pombe genomic database analysis (Fig. 2A). The 3.6-kb fragment contain-ing SPAPB21F2.02, which is an S. cerevisiae DOP1 homo-logue involved in cell shape and cell size control (Pascon and Miller, 2000), did not complement the hygromycin B sensitivity of the gmn2 mutant. In contrast, the 2.3-kb frag-ment containing SPAC227.01c, which is an S. cerevisiae
ERD1 (ER Retention Defective) homologue involved in
retention of ER-resident proteins, complemented the hygromycin B sensitivity (Fig. 2A). The SPAC227.01c ORF was predicted to encode a protein of 373 amino ac-ids with 6 potential N-glycosylation sites. Transformation of the gmn2 mutant with the cloned gene not only restored wild type tolerance to hygromycin B, but also comple-mented the glycosylation defects (data not shown). There-fore, we designated this gene sgm2+ for suppressor of gmn2 mutation.
A BLAST search revealed a significant degree of simi-larity to K. lactis Erd1 (23.8% identity (Dean, 1994)) and to S. cerevisiae Erd1 (23.8% identity (Hardwick et al., 1990)) (Fig. 2B). Although the amino acid sequence of Sgm2p had a relatively low overall level of similarity with
K. lactis and S. cerevisiae Erd1 proteins, the hydropathic
profiles and secondary structures were found to be highly conserved (Fig. 2C). The hydropathic profile suggests that Sgm2p encodes an intrinsic membrane protein with be-tween five (predicted by the SOSUI system, Hirokawa et al. (1998)), and eight (TMpred program, Hofmann and Stoffel (1993)) putative transmembrane domains (Fig. 2C). These results strongly suggest that sgm2+ encodes an Erd1-like S. pombe protein.
To confirm that sgm2+ is identical to gmn2+, we first sequenced the sgm2+ locus in the gmn2 mutant as follows. A 1.3-kb fragment containing sgm2+ was amplified by PCR from genomic DNA isolated from gmn2 mutant cells. To avoid possible PCR artifacts, at least three complete sgm2+ alleles were sequenced independently. The mutant sgm2+ allele was found to have undergone a single base pair sub-stitution that resulted in the replacement of TAC (Tyr195) with a stop codon, TAA. Next, ura4+ was integrated adja-cent to sgm2+ in wild type (TP4-1D). A heterozygous dip-loid generated by crossing the sgm2+ ura4+ integrant to a
gmn2 ura4 mutant was sporulated and subjected to
ran-dom spore analysis. Uracil auxotrophy and hygromycin B sensitivity were found to be completely linked among spore progeny, i.e., all ura auxotrophs were sensitive to hygromycin B, indicating that gmn2+ is identical to sgm2+ (data not shown).
Phenotypic analysis of gmn2∆∆∆∆∆ cells
In S. cerevisiae, the ERD1 gene product is required for correct localization of soluble proteins that normally re-side in the ER (Hardwick et al., 1990). Cells lacking ERD1 secrete resident ER proteins and exhibit defects in the processing of glycoproteins (Hardwick et al., 1990). To examine the physiological role of the Gmn2 protein, the
gmn2+ gene was disrupted by insertion of ura4+ in a hap-Fig. 1. Glycosylation defect and hygromycin B sensitivity of the gmn2
mutant.
A. Activity stain for invertase. Cells from wild type (left) and the gmn2 mutant (right) induced for invertase expression were homogenized with glass beads and the extracts were resolved by electrophoresis in 4% polyacrylamide. The gel was developed with triphenyltetrazolium chlo-ride. B. Wild type (WT) and gmn2 mutant cells were streaked onto YES, and YES containing 25 µg/ml hygromycin B plates, and incubated for 3 to 6 days at 30°C.
gmn2 encodes an Erd1 71
loid strain. Three independent ura4+ transformants were
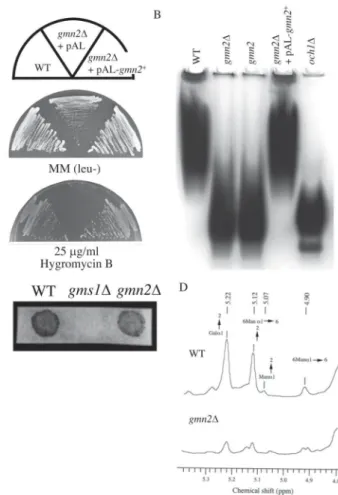
subjected to Southern analysis and were found to contain one copy each of the disrupted gmn2 gene (data not shown). The gmn2∆ cells were found to be viable despite growing slightly slower than the wild type (Fig. 3A MM (leu-)) and exhibited the same phenotypes as those of the original gmn2 mutant. The gmn2∆ cells were highly sen-sitive to hygromycin B, being unable to grow on YES plates containing 25 µg/ml of the drug (Fig. 3A).
Trans-formation of the gmn2∆ strain with plasmid pAL-gmn2+
complemented this phenotype (Fig. 3A).
To investigate the effect of disrupting gmn2+ on protein
glycosylation, the electrophoretic mobility of acid phos-phatase, another recognized glycoprotein in S. pombe, was analyzed. Cell extracts were loaded onto a native poly-acrylamide gel, electrophoresed, and stained (Fig. 3B). The mobility of acid phosphatase secreted from gmn2∆ cells was found to be faster than that of the enzyme isolated from wild type cells, but almost the same as that of acid phosphatase from the original gmn2 mutant. To analyze the extent of the glycosylation defect, the mobility of the acid phosphatase was compared with that from an och1∆ strain which is deficient in the α-1,6-mannosyltransferase activity involved in initiation of mannose outer chain elon-gation of N-linked oligosaccharides (Yoko-o et al., 2001). The mobility of the acid phosphatase from gmn2∆ cells was found to be slower than that of och1∆ cells, suggest-ing that elongation of N-linked carbohydrate chains was Fig. 2. Sequence analysis of the sgm2+ gene.
A. Restriction map of the isolated genomic 4.97 kb SpeI fragment, and complementation analysis of different subclones. B. Alignment of the amino acid sequence of the Sgm2 protein (SpSgm2p), K. lactis Erd1 (KlErd1p), and S. cerevisiae Erd1 (ScErd1p). Identical amino acids in at least two of the three sequences (including Sgm2p) are highlighted. C. The algorithm of Kyte and Doolittle (1982) was used to plot hy-dropathy profiles of Sgm2p, K. lactis Erd1p, and S. cerevisiae Erd1p, with a window of 11 amino acids. Hydrophobic portions of the proteins are above the line. Boxes indicate the positions of putative membrane spanning segments. Open boxes: analysis based on TMpred (Hofmann and Stoffel 1993); hatched boxes: analysis based on SOSUI (Hirokawa et al., 1998).
Fig. 3. Phenotypic analysis of gmn2∆ cells.
A. Wild type (WT), gmn2∆ with empty vector (gmn2∆ + pAL), or gmn2∆ transformed with pAL-gmn2+ (gmn2∆ + pAL-gmn2+) were cultured on YES plates with or without 25 µg/ml hygromycin B, and incubated for 3 to 6 days at 30°C. B. Activity stain for acid phosphatase. Lysates from wild type (WT), gmn2∆ (gmn2∆), gmn2 mutant (gmn2), gmn2∆ transformed with pAL-gmn2+ (gmn2∆ + pAL-gmn2+), or och1∆ (och1∆) induced for acid phosphatase expression were subjected to gel electro-phoresis and stained for invertase activity. C. Horseradish peroxidase-conjugated peanut lectin (HRP-PNA) staining reveal agglutination with PNA. D. 1H NMR spectroscopy of gmn2∆ glycans. The purified N-linked galactomannans from the wild type (WT) and gmn2∆ were analyzed by one-dimensional 1H NMR spectroscopy. The peaks were normalized with respect to that of acetone.
reduced in gmn2∆ cells relative to the wild type, but not as severely as in och1∆ cells (Fig. 3B).
Analysis of galactosylation of cell surface proteins in the gmn2∆ disruptant by staining with HRP-PNA, which specifically recognizes galactose residues (Tanaka et al., 2001), indicated that this process was not impaired (Fig. 3C).
To confirm the glycan structure of gmn2∆ cells, N-linked galactomannans were extracted and analyzed by 1H
NMR spectroscopy (Fig. 3D). The NMR spectrum of the wild type polysaccharide had H-l signals with intensities at 4.91, 5.07, 5.12, and 5.22 ppm, which was consistent with those of the oligosaccharides containing galactose from S. pombe (Ballou et al., 1994). A signal at 4.90 ppm indicates an α-1,6-linked unsubstituted mannose
back-bone; at 5.07 ppm, a terminal mannose linked α-1,2; at 5.12 ppm, a substituted mannose main chain at position 2 and at 5.22 ppm, mainly terminal galactose linked α-1,2 as side chains. Therefore, the N-glycan of wild type cells have an α-1,6-mannose backbone with side chains con-sisting of mainly α-1,2-linked galactose residues, and small amounts of α-1,2-linked mannose residues. In con-trast, the NMR spectrum of gmn2∆ N-glycan is quite dif-ferent from that of wild type (Fig. 3D). The anomeric pro-ton region of the spectrum showed a significant decrease with the peak of the main chain substituted with side chains, although the galactose side chain at 5.22 ppm was present. These results revealed that gmn2∆ cells have a remarkable defect in the elongation of the N-linked galactomannan backbone.
Fig. 4. Gmn2p is involved in stable localization of glycosyltransferases responsible for backbone biosynthesis of N-glycan.
A. Intracellular localization of Och1-EGFP in wild type (WT) and gmn2∆. S. pombe wild type and gmn2∆ cells expressing pAL-Och1-EGFP were cultured in minimal medium without leucine for 18 h, and then the cells were examined using an Olympus BX-50 fluorescence microscope. pAU-Och1-EGFP (B) or pAL-Mnn9-EGFP (C: upper) or pAL-Anp1-EGFP (C: lower) or pAL-Gmh3-EGFP (D) was expressed in wild type (WT) and
gmn2∆, respectively. The prepared solubilized crude proteins (30 µg) were separated on 10% SDS-PAGE, and they were immunoblotted with Anti-EGFP or Anti-β Tubulin antibody.
gmn2 encodes an Erd1 73
Gmn2p is involved in stable localization of glycosyltransferases responsible for backbone biosynthe-sis of N-glycan
In S. cerevisiae, it is shown that gene disruption of ERD1 severely affects the localization of Och1p which initiates polymannose outer chain elongation of N-glycan in cis-Golgi, but not Mnn9p which mediates elongation of the
N-glycan mannan backbone (Okamoto et al., 2008). In S. pombe, Och1p has also been identified in fission yeast
(Yoko-o et al., 2001), and recently we have also identi-fied that Mnn9p and Anp1p are involved in mannan elon-gation and that they associate with each other to form a
complex (Ohashi et al., 2020, in press). Therefore, we at-tempted to confirm whether the localization of these glycosyltransferases was affected in gmn2∆ cells (Fig. 4). As expected, Och1-EGFP expressed in the wild type strain showed strong fluorescence as a typical Golgi-like dots, but faint fluorescent dots were confirmed in gmn2∆ cells (Fig. 4A). Similarly, the signal of Och1-EGFP was weaker than that of the wild type strain in Western blot-ting analysis (Fig. 4B). Furthermore, unlike S. cerevisiae described above, weak expression of Mnn9 and Anp1 was also confirmed (Fig. 4C). On the other hand, it was also revealed that the expression of Gmh3-EGFP, which is a α-1,2-galactosyltransferase (Ohashi et al., 2011; Yoko-o et al., 1998), was equivalent to that of the wild type in
gmn2∆ cells (Fig. 4D). These results suggest that the Gmn2p is required for the stable localization of the mannosyltransferases involved in the initial N-glycan elon-gation, but not in the stability of the late-acting galactosyltransferase.
Soluble ER proteins are secreted in gmn2∆∆∆∆∆ cells BiP is a member of a class of resident ER proteins, which are soluble in the lumen. The localization of BiP is medi-ated by a C-terminal tetrapeptide (HDEL) signal that is both necessary and sufficient for ER retention in S.
cerevisiae (Munro and Pelham, 1987). The S. pombe BiP
gene has been cloned from (bip1+; SPAC22A12.15c) and
the C-terminal tetrapeptide ADEL of S. pombe BiP has Fig. 6. Intracellular localization of Gmn2-EGFP in wild type cells.
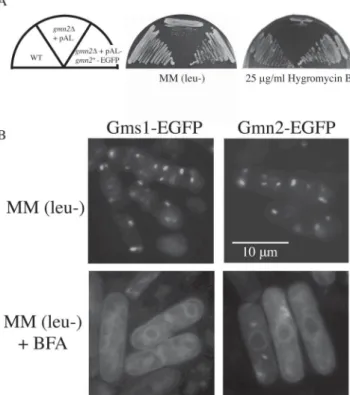
A. Cells were streaked onto YES plates with, or without, 25 µg/ml hygromycin B, and incubated for 3 to 6 days at 30°C. B. Intracellular localization of Gms1-EGFP (left lane) or Gmn2-EGFP (right lane) was visualized by fluorescence microscopy in wild type cells. Wild type cells expressing Gms1-EGFP or Gmn2-EGFP fusion protein were grown to mid-log phase in appropriate medium (upper panel), and BFA was added at a final concentration of 100 µg/ml. Cells were grown for 45 min prior to observation (bottom panel).
Fig. 5. Immunological dot-blot analysis of BiP secretion from the gmn2∆ strain.
A. Cells were grown on solid media overlaid with a nitrocellulose mem-brane to allow adherence of secreted BiP. BiP was detected by immunoblotting using anti-BiP antibodies as a probe (lower panel). B. Intracellular localization of Erd2-EGFP integrated into wild type (WT) or gmn2∆ strain was visualized by fluorescent microscopy. C. These cells were cultured until the logarithmic growth phase, and the expres-sion of Erd2-EGFP was confirmed by immunoblotting with Anti-EGFP or Anti-β Tubulin antibody.
been shown to be necessary for retention in the ER (Pidoux and Armstrong, 1992).
In S. cerevisiae, cells lacking the ERD1 gene secrete the endogenous ER-resident protein, BiP (Hardwick et al., 1990). In order to determine whether the Gmn2 protein is required for retention of luminal ER proteins in S. pombe, BiP secretion was examined in the wild type, and in the
gmn2∆ cells by immunoblotting (Fig. 5A). As expected, cell surface-associated BiP was not detected in wild type cells. In contrast, gmn2∆ cells missorted and secreted a significant amount of BiP to the cell surface. These re-sults indicate that Gmn2p is required for normal retention of a luminal ER protein in S. pombe cells.
Analogous to the stability of glycosyltransferases (Fig. 4), we analyzed whether the localization and expression of Erd2p, a receptor for BiP (ADEL), was unstable in
gmn2∆ cells (Figs. 5B and C). As a result, it was con-firmed that Erd2-EGFP expressed in the wild type strain showed ER and Golgi-like localization, but that the fluo-rescence intensity was significantly reduced in gmn2∆ cells, and the expression was also decreased.
These results suggest that Gmn2p is also involved in the stable localization of Erd2p for Bip receptor and indi-rectly contributes to the retention of Bip between the ER and the Golgi apparatus.
Intracellular localization of Gmn2-EGFP fusion protein
The intracellular localization of Erd1 in S. cerevisiae and K. lactis has not been well established, although the
gmn2+ product appears to be an integral membrane pro-tein as described above (Fig. 2A). To determine the intra-cellular localization of Gmn2 in S. pombe cells, an en-hanced green fluorescent protein (EGFP) was fused with the carboxyl terminus of Gmn2 under the control of the
gmn2+ promoter (Fig. 6). To confirm that the Gmn2-EGFP fusion protein was functional in gmn2∆ cells, transformants were cultured on YES plates containing 25 µg/ml hygromycin B at 30°C. Because the Gmn2-EGFP fusion protein restored normal tolerance towards hygromycin B, it was concluded that the Gmn2-EGFP fu-sion protein was functional in S. pombe cells (Fig. 6A). By fluorescent microscopy, cells expressing the Gmn2-EGFP fusion construct appeared to have multiple green spots per cell (Fig. 6B). This intracellular staining pattern is similar to that exhibited by Golgi-localized α-1,2-galactosyltransferase (Gma12p) and the UDP-galactose transporter protein, Gms1p (Brazer et al., 2000; Chappell et al., 1994; Tanaka and Takegawa, 2001).
To further characterize the localization of Gmn2p to the Golgi membrane, the effect of brefeldin A (BFA) was ex-amined, which causes recycling of the Golgi apparatus to the ER in many organisms including fission yeast (Klausner et al., 1992; Lippincott-Schwartz et al., 1989; Pelham, 1991; Turi et al., 1994). Both wild type and gmn2∆ cells expressing Gms1-EGFP or the Gmn2-EGFP were grown at 30°C, and then treated with BFA for 90 min. The Gmn2-EGFP protein was recycled back into the ER just as Gms1-EGFP, indicating that Gmn2-EGFP localized mostly to the Golgi membranes (Fig. 6B). After the wash-ing of cells and resuspension in normal growth medium without BFA, Gmn2-EGFP was found to relocalize to the
Golgi from the ER (data not shown). These results sug-gest that the Gmn2 protein functions at the Golgi mem-brane where it is required for normal glycosylation of se-creted glycoproteins and for retention of ER proteins.
Discussion
In this report, the fission yeast gmn2+ gene product was shown to be functionally equivalent to the S. cerevisiae Erd1 protein. Budding yeast erd mutants were originally isolated based on the defective retention of an HDEL-con-taining invertase fusion protein (Pelham et al., 1988). Mutants with a strong phenotype fell into two complementation groups and the corresponding genes have since been cloned. ERD1 encodes a 43-kd membrane pro-tein that is not required for normal growth or secretion of invertase (Hardwick et al., 1990). Hardwick et al. (1990) suggested that the budding yeast Erd1p is required for the maintenance of the structural integrity or appropriate aque-ous environment of the Golgi. However, the precise func-tions of Erd1p in S. cerevisiae remain unknown. The ERD1 homolog, also present as XPR1 in humans, is reported only as a human cell-surface receptor for xenotropic and poly-tropic murine leukemia viruses (Battini et al., 1999) and is presumed to be functionally completely different from yeast ERD1. The ERD1 homologues from various filamentous fungi are available in the genomic database, but few reports have been published regarding functional analysis.
The present study found that a functional Gmn2-GFP fusion protein localized to the Golgi membrane, and relo-cated to the ER membrane after treatment with BFA (Fig. 6C). To our knowledge, this is the first report that the prod-uct of a yeast ERD1 homologue localizes to and functions at the yeast Golgi membrane. Although the exact com-partment containing Gmn2p within the Golgi stacked cis-terna has not been determined, Gmn2p appears be local-ized to the cis-compartment of the Golgi apparatus, pos-sibly to regulate the cis-Golgi environment, intra-Golgi transport, or both.
Based on the mobility of acid phosphatase and on cell surface galactose content, the gmn2∆ cells exhibited a se-vere defect in elongation of N-linked polymannose back-bones (Fig. 3B), but not in cell surface protein galactosylation (Fig. 3C). 1H NMR analysis also revealed that the N-glycan of gmn2∆ cells had a significantly shorter α-1,6-mannose backbone, but the α-1,2-galactose side chain were modified according to the length of the back-bone (Fig. 3D). It has been known that the α1,2- and α1,3-galactose residues is also modified in the Man9GlcNAc2 core structure derived from the och1∆ strain in which the elongation of the outer chain in the cis-Golgi is completely stopped (Ohashi et al., 2009). This means that the intact galactosyltransferase located in the med-to late-Golgi layer will modify the galactose chain. Indeed, in gmn2∆ cells, glycosyltransferases that function in the early glycosylation pathway are unstable in both localization and expression; however, stable expression of galactosyltransferase resulted in normal galactosylation of the cell surface (Figs. 4 and 3C). From these results, it is highly possible that Gmn2p is specifically involved in the
gmn2 encodes an Erd1 75 stability of the cis-Golgi layer structure.
In the Golgi of S. cerevisiae, glycosylation occurs solely by the addition of mannose and several mannosyltransferases have been identified. Two hetero-oligomeric complexes that contain Mnn9p as a common subunit, Mnn9p-Van1p (M-Pol I) and Mnn9p-Anp1p-Hoc1p-Mnn10p-Mnn11p (M-Pol II), transfer either α-1,6-or α-1,2-mannose to acceptor N-glycosyl groups (Jungmann and Munro, 1998). Interestingly, in S.
cerevisiae, the localization of Och1p to the Golgi
appara-tus is severely affected by the deletion of ERD1 while the localization of Mnn9p is not appreciably affected (Okamoto et al., 2008). Recently we have reported that Mnn9p and Anp1p are involved in mannan elongation and that they associate with each other to form a complex in
S. pombe (Ohashi et al., 2020 (in press)). However, S. pombe lacks ScVan1 and ScHoc1 protein homologs. Thus,
it is not clear whether S. pombe has distinct M-Pol I and II-like protein complexes. At present, no interaction be-tween Gmn2p and Mnn9p or Anp1p has been detected due to the difficulty of solubilizing membrane proteins. In the future, we should analyze whether gmn2∆ cells impair the interaction between Mnn9p and Anp1p.
One glycosylation mutant in S. pombe, the ryh1 null mutant, also secretes an underglycosylated invertase (Hengst et al., 1990). The ryh1+ gene was originally iso-lated as a homologue of S. cerevisiae YPT1 encoding a GTP-binding protein involved in intracellular protein transport (Hengst et al., 1990). The Ryh1 protein is 73% identical to human Rab6, which regulates transport be-tween the early and late Golgi compartment (Martinez et al., 1994, 1997). Interestingly, the mobility of invertase from gmn2∆ cells on native PAGE was faster than that of invertase from ryh1∆ cells (Tanaka et al. unpubl.), indi-cating that the glycosylation defect of the gmn2∆ strain is more severe than that of the ryh1∆ strain. This is consist-ent with the Golgi defect in gmn2∆ influencing an up-stream component of Ryh1p that is involved in regulation of intracellular Golgi transport. Gmn2p may also be re-quired to maintain normal vesicular transport within the Golgi.
Loss of Gmn2p causes S. pombe cells to missort the ER-resident protein, BiP (Fig. 5A). Recycling of the fission yeast BiP, ADEL-containing protein from the Golgi to the ER is deficient in gmn2∆ cells. In S. cerevisiae, BiP is transported to the Golgi from the ER and is then selec-tively retrieved by the ER via the Golgi-localized receptor, Erd2 (Lewis et al., 1990; Semenza et al., 1990). The bud-ding yeast Erd2 protein encodes the HDEL receptor and is essential for growth (Semenza et al., 1990). ERD2 homo-logues have been identified in a wide variety of other spe-cies including plants (Lee et al., 1993) and humans (Hsu et al., 1992; Lewis et al., 1990). Because the conservation of ER-retention signals between budding (HDEL) and fis-sion (ADEL) yeasts suggests that S. pombe should also possess Erd2 receptor-like proteins, the complete S. pombe genome database was searched for genes encoding ERD2 homologues. This search revealed the presence of one gene (SPBP8B7.22) on S. pombe chromosome II, suggesting that BiP interacts with Erd2p in the Golgi cis-compart-ment which is then recycled back to the ER in S. pombe
cells. Interestingly, genome-wide screen of disruptants shows that the erd2∆ strain is not lethal (Calvo et al., 2009). In the gmn2∆ strain, a large fraction of BiP that is trans-ported to the cis-Golgi compartment might bind poorly to Erd2p due to an impaired cis-Golgi function (Figs. 5B and C).
We are currently examining the retention stability of cis-Golgi compartment-resident proteins, such as the mannosyltransferase complexes and the Erd2 homologue in the gmn2∆ strain. Further analysis of gmn2+ and the function of its product should further the understanding of the cis-Golgi compartment relative to glycosylation and sorting of ER luminal proteins.
Acknowledgments
We thank Drs. Clinton Ballou, Takashi Toda, Chikashi Shimoda, and John Armstrong for providing the S. pombe strains, antibodies, and plasmids. We acknowledge the technical expertise of the DNA core fa-cility of the Gene Research Center, Kagawa University. This work was supported by JSPS KAKENHI Grant Numbers JP16K07669, and by the New Energy and Industrial Technology Development Organization (to K.T.).
References
Alfa, C., Fantes, P., Hyams, J., McLoed, M., and Warbrick, E. (1993) Experiments with fission yeast: a laboratory course manual. Cold Spring Harb. Lab. Press.
Ballou, C. E., Ballou, L., and Ball, G. (1994) Schizosaccharomyces
pombe glycosylation mutant with altered cell surface properties. Proc. Natl. Acad. Sci. USA, 91, 9327–9331.
Ballou, L. and Ballou, C. (1995) Schizosaccharomyces pombe mutants that are defective in glycoprotein galactosylation. Proc. Natl. Acad.
Sci. USA, 92, 2790–2794.
Battini, J. L., Rasko, J. E., and Miller, A. D. (1999) A human cell-sur-face receptor for xenotropic and polytropic murine leukemia vi-ruses: possible role in G protein-coupled signal transduction. Proc.
Natl. Acad. Sci. USA, 96, 1385–1390.
Brazer, S. C., Williams, H. P., Chappell, T. G., and Cande, W. Z. (2000) A fission yeast kinesin affects Golgi membrane recycling. Yeast,
16, 149–166.
Calvo, I. A., Gabrielli, N., Iglesias-Baena, I., García-Santamarina, S., Hoe, K.-L. et al. (2009) Genome-wide screen of genes required for caffeine tolerance in fission yeast. PLoS One, 4, e6619.
Chappell, T. G. and Warren, G. (1989) A Galactosyltransferase from the Fission Yeast. J. Cell Biol., 109, 2693-2702.
Chappell, T. G., Hajibagheri, M. A., Ayscough, K., Pierce, M., and Warren, G. (1994) Localization of an α 1,2 galactosyltransferase activity to the Golgi apparatus of Schizosaccharomyces pombe. Mol.
Biol. Cell, 5, 519–528.
Dean, N. (1994) Cloning and DNA sequence of a Kluyveromyces lactis
ERD1 homologue. Yeast, 10, 1117–1124.
Dean, N. (1995) Yeast glycosylation mutants are sensitive to aminoglycosides. Proc. Natl. Acad. Sci. USA, 92, 1287–1291. Gabriel, O. and Wang, S. F. (1969) Determination of enzymatic activity
in polyacrylamide gels. I. Enzymes catalyzing the conversion of nonreducing substrates to reducing products. Anal. Biochem., 27, 545–554.
Gemmill, T. R. and Trimble, R. B. (1996) Schizosaccharomyces pombe produces novel pyruvate-containing N-linked oligosaccharides. J.
Biol. Chem., 271, 25945–25949.
Grimm, C., Kohli, J., Murray, J., and Maundrell, K. (1988) Genetic engineering of Schizosaccharomyces pombe: a system for gene dis-ruption and replacement using the ura4 gene as a selectable marker.
Mol. Gen. Genet., 215, 81–86.
Hardwick, K. G., Lewis, M. J., Semenza, J., Dean, N., and Pelham, H. R. (1990) ERD1, a yeast gene required for the retention of luminal endoplasmic reticulum proteins, affects glycoprotein processing in the Golgi apparatus. EMBO J., 9, 623–630.
Hengst, L., Lehmeier, T., and Gallwitz, D. (1990) The ryh1 gene in the fission yeast Schizosaccharomyces pombe encoding a GTP-bind-ing protein related to ras, rho and ypt: structure, expression and identification of its human homologue. EMBO J., 9, 1949–1955. Herscovics, A. and Orlean, P. (1993) Glycoprotein biosynthesis in yeast.
FASEB J., 7, 540–550.
Hirokawa, T., Boon-Chieng, S., and Mitaku, S. (1998) SOSUI: classifi-cation and secondary structure prediction system for membrane proteins. Bioinformatics, 14, 378–379.
Hofmann, K. and Stoffel, W. (1993) TMbase-a database of membrane spanning proteins segments. Biol. Chem. Hoppe. Seyler., 347, 166. Hsu, V. W., Shah, N., and Klausner, R. D. (1992) A brefeldin A-like phenotype is induced by the overexpression of a human ERD-2-like protein, ELP-1. Cell, 69, 625–635.
Huang, K. M. and Snider, M. D. (1995) Isolation of protein glycosylation mutants in the fission yeast Schizosaccharomyces pombe. Mol. Biol.
Cell, 6, 485–496.
Jungmann, J. and Munro, S. (1998) Multi-protein complexes in the cis G o l g i o f S a c c h a ro m y c e s c e re v i s i a e w i t h a l p h a 1 , 6 -mannosyltransferase activity. EMBO J., 17, 423–434.
Keeney, J. B. and Boeke, J. D. (1994) Efficient Targeted Integration at
leul-32 and ura4-294 in Schizosaccharomyces pombe. Genetics, 136,
849–856.
Klausner, R. D., Donaldson, J. G., and Lippincott-Schwartz, J. (1992) Brefeldin A: insights into the control of membrane traffic and or-ganelle structure. J. Cell Biol., 116, 1071–1080.
Kyte, J. and Doolittle (1982) A simple method for displaying the hy-dropathic character of a protein. J. Mol. Biol., 157, 105–132. Laemmli, U. K. (1970) Cleavage of structural proteins during the
as-sembly of the head of bacteriophage T4. Nature, 227, 680–685. Lee, H. I., Gal, S., Newman, T. C., and Raikhel, N. V. (1993) The
Arabidopsis endoplasmic reticulum retention receptor functions in yeast. Proc. Natl. Acad. Sci. USA, 90, 11433–11437.
Lewis, M. J., Sweet, D. J., and Pelham, H. R. (1990) The ERD2 gene determines the specificity of the luminal ER protein retention sys-tem. Cell, 61, 1359–1363.
Lippincott-Schwartz, J., Yuan, L. C., Bonifacino, J. S., and Klausner, R. D. (1989) Rapid redistribution of Golgi proteins into the ER in cells treated with brefeldin A: evidence for membrane cycling from Golgi to ER. Cell, 56, 801–813.
Manners, J. D. and Meyer, T. M. (1977) The molecular structures of some glucans from the cell walls of Schizosaccharomyces pombe.
Carbohydr. Res., 57, 189–203.
Martinez, O., Schmidt, A., Salamero, J., Hoflack, B., Roa, M. et al. (1994) The small GTP-binding protein rab6 functions in intra-Golgi transport. J. Cell Biol., 127, 1575–1588.
Martinez, O., Antony, C., Pehau-Arnaudet, G., Berger, E. G., Salamero, J. et al. (1997) GTP-bound forms of rab6 induce the redistribution of Golgi proteins into the endoplasmic reticulum. Proc. Natl. Acad.
Sci. USA, 94, 1828–1833.
Moreno, S., Ruiz, T., Sanchez, Y., Villanueva, J. R., and Rodriguez, L. (1985) Subcellular localization and glycoprotein nature of the in-vertase from the fission yeast Schizosaccharomyces pombe. Arch.
Microbiol., 142, 370–374.
Moreno, S., Klar, A., and Nurse, P. (1991) Molecular genetic analysis of fission yeast Schizosaccharomyces pombe. Methods Enzym., 194, 795–823.
Morita, T. and Takegawa, K. (2004) A simple and efficient procedure for transformation of Schizosaccharomyces pombe. Yeast, 21, 613– 617.
Munro, S. and Pelham, H. R. (1987) A C-terminal signal prevents se-cretion of luminal ER proteins. Cell, 48, 899–907.
Ohashi, T., Ikeda, Y., Tanaka, N., Nakakita, S., Natsuka, S. et al. (2009) The och1 mutant of Schizosaccharomyces pombe produces galactosylated core structures of N-linked oligosaccharides. Biosci.
Biotechnol. Biochem., 73, 407–414.
Ohashi, T., Nakakita, S., Sumiyoshi, W., Yamada, N., Ikeda, Y. et al. (2011) Structural analysis of α 1,3-linked galactose-containing oligosaccharides in Schizosaccharomyces pombe mutants harboring single and multiple α-galactosyltransferase genes disruptions.
Glycobiology, 21, 340–351.
Ohashi, T., Tanaka, T., Tanaka, N., and Takegawa, K. (2020) SpMnn9p and SpAnp1p form a protein complex involved in mannan synthe-sis in the fission yeast Schizosaccharomyces pombe. J. Biosci.
Bioeng. (in press).
Okamoto, M., Yoko-o, T., Miyakawa, T., and Jigami, Y. (2008) The cy-toplasmic region of α-1,6-mannosyltransferase Mnn9p is crucial for retrograde transport from the Golgi apparatus to the endoplas-mic reticulum in Saccharomyces cerevisiae. Eukaryot. Cell, 7, 310– 318.
Pascon, R. C. and Miller, B. L. (2000) Morphogenesis in Aspergillus
nidulans requires Dopey (DopA), a member of a novel family of
leucine zipper-like proteins conserved from yeast to humans. Mol.
Microbiol., 36, 1250–1264.
Peat, S. and Rees, D. A. (1961) Carbohydrase and sulphatase activities of Porphyra umbilicalis. Biochem. J., 79, 7–12.
Pelham, H. R. (1991) Multiple targets for brefeldin A. Cell, 67, 449– 451.
Pelham, H. R., Hardwick, K. G., and Lewis, M. J. (1988) Sorting of soluble ER proteins in yeast. EMBO J., 7, 1757–1762.
Pidoux, A. L. and Armstrong, J. (1992) Analysis of the BiP gene and identification of an ER retention signal in Schizosaccharomyces
pombe. EMBO J., 11, 1583–1591.
Schweingruber, M. E., Fluri, R., Maundrell, K., Schweingruber, A. M., and Dumermuth, E. (1986) Identification and characterization of thiamin repressible acid phosphatase in yeast. J. Biol. Chem., 261, 15877–15882.
Semenza, J. C., Hardwick, K. G., Dean, N., and Pelham, H. R. (1990)
ERD2, a yeast gene required for the receptor-mediated retrieval of
luminal ER proteins from the secretory pathway. Cell, 61, 1349– 1357.
Suga, M. and Hatakeyama, T. (2001) High efficiency transformation of
Schizosaccharomyces pombe pretreated with thiol compounds by
electroporation. Yeast, 18, 1015–1021.
Tabuchi, M., Tanaka, N., Iwahara, S., and Takegawa, K. (1997) The
Schizosaccharomyces pombe gms1+ gene encodes an UDP-galac-tose transporter homologue required for protein galactosylation.
Biochem. Biophys. Res. Commun., 232, 121–125.
Takegawa, K., Tanaka, N., Tabuchi, M., and Iwahara, S. (1996) Isola-tion and characterizaIsola-tion of a glycosylaIsola-tion mutant from
Schizosaccharomyces pombe. Biosci. Biotechnol. Biochem., 60,
1156–1159.
Tanaka, N. and Takegawa, K. (2001) Functional characterization of Gms1p/UDP-galactose transporter in Schizosaccharomyces pombe.
Yeast, 18, 745–757.
Tanaka, N., Ohuchi, N., Mukai, Y., Osaka, Y., Ohtani, Y. et al. (1998) Isolation and characterization of an invertase and its repressor genes from Schizosaccharomyces pombe. Biochem. Biophys. Res.
Commun., 245, 246–253.
Tanaka, N., Awai, A., Bhuiyan, M. S., Fujita, K., Fukui, H. et al. (1999) Cell surface galactosylation is essential for nonsexual flocculation in Schizosaccharomyces pombe. J. Bacteriol., 181, 1356–1359. Tanaka, N., Konomi, M., Osumi, M., and Takegawa, K. (2001)
Charac-terization of a Schizosaccharomyces pombe mutant deficient in UDP-galactose transport activity. Yeast, 18, 903–914.
Turi, T. G., Webster, P., and Rose, J. K. (1994) Brefeldin A sensitivity and resistance in Schizosaccharomyces pombe. Isolation of multi-ple genes conferring resistance. J. Biol. Chem., 269, 24229–24236. Umeda, K., Yoko-o, T., Nakayama, K. I., Suzuki, T. O., and Jigami, Y. (2000) Schizosaccharomyces pombe gmd3+/alg11+ is a functional homologue of Saccharomyces cerevisiae ALG11 which is involved in N-linked oligosaccharide synthesis. Yeast, 16, 1261–1271. Yoko-o, T., Roy, S. K., and Jigami, Y. (1998) Differences in in vivo
acceptor specificity of two galactosyltransferases, the gmh3+ and
gma12+ gene products from Schizosaccharomyces pombe. Eur. J.
Biochem., 257, 630–637.
Yoko-o, T., Tsukahara, K., Watanabe, T., Hata-Sugi, N., Yoshimatsu, K. et al. (2001) Schizosaccharomyces pombe och1+ encodes α-1,6-mannosyltransferase that is involved in outer chain elongation of