RAGE-dependent NF-κB inflammation processes
in the capsule of frozen shoulders
著者
YANO TOSHIHISA
学位授与機関
Tohoku University
学位授与番号
11301甲第19145号
1
博士論文
RAGE-dependent NF-κB inflammation processes
in the capsule of frozen shoulders
(凍結肩の関節包における最終糖化産物受容体による
NF-κB 反応経路の活性化に関する研究)
東北大学大学院医学系研究科医科学専攻 外科病態学講座整形外科学分野 矢野 利尚
2
Contents
1. Abstract ... 3
2. Introduction ... 5
3. Purpose ... 9
4. Materials and methods ... 10
5. Results ... 18
6. Discussion ... 24
7. Conclusion ... 36
8. References ... 37
9. Figure Legends ...45
10. Table Headings ... 51
11. Acknowledgements ... 52
12. Abbreviations ... 53
3
1. Abstract
Background: Inflammation with fibrosis is one of the primary pathologies of FS.
However, the etiology of frozen shoulder (FS) remains unknown. Advanced glycation end-products (AGEs) cause cross-linking and stabilization of collagen and are reported to increase in FS. The present study aimed to elucidate the pathogenesis of FS by evaluating the receptor of AGE (RAGE)-dependent pathways.
Methods: Tissue samples of the rotator interval (RI), coracohumeral ligament (CHL), and
anterior-inferior glenohumeral ligament (IGHL) were collected from 33 patients with FS, presenting with severe stiffness, and 25 patients with rotator cuff tears (RCT) as controls. Gene expression levels of RAGE, high-mobility group box 1 (HMGB1), Toll-like receptor 2 (TLR2), TLR4, S100 calcium binding protein A1 (S100A1), S100B, nuclear factor-κ B (NF-κB), and cytokines were determined using quantitative real-time polymerase chain reaction. The immunoreactivity of carboxymethyllysine (CML), pentosidine, and RAGE were also evaluated. CML and pentosidine were evaluated using high-performance liquid chromatography (HPLC).
Results: Gene expression levels of RAGE, HMGB1, TLR2, TLR4, S100A1, S100B, and NF-κB were significantly greater in the CHL and IGHL tissue from the FS group
4
HMGB1 in CHL, p = 0.008; in IGHL, p < 0.001; TLR2 in CHL, p = 0.044; in IGHL, p =
0.004; TLR4 in CHL, p = 0.021; in IGHL, p = 0.027; S100A1 in CHL, p = 0.001; in IGHL, p = 0.008; S100B in CHL, p = 0.001; in IGHL, p = 0.012; NF-κB in CHL, p = 0.001; in IGHL, p = 0.035). Further, the immunoreactivity of RAGE and CML was stronger in the CHLs and IGHLs from the FS group compared to those from the RCT group. Pentosidine was found to be weakly immunostained in the CHLs, IGHLs, and RIs from the FS group. HPLC results show that CML levels were significantly greater in the CHLs and IGHLs from the FS group compared to those from the RCT group (CHL, p = 0.011; IGHL, p = 0.008).
Conclusion: The results indicated that in AGEs, CML, rather than pentosidine, exerted a
more pronounced effect on FS pathology. AGEs, HMGB1, and S100 protein are supposed to bring inflammation with fibrosis in FS, by binding to RAGE and activating NF-κB signaling pathways. Hence, suppression of these pathways could provide a potential treatment option for FS.
5
2. Introduction
Frozen shoulder (FS) is characterized by severe pain and restricted active and passive ranges of motion (ROMs) 1. The pain and stiffness associated with FS generally
restrains activities of daily living and working, and disturbs quality of life. The natural course of this disease is considered to be self-limiting 2. Symptoms often last two to three
years, with three classical stages: freezing, frozen, and thawing 3.
Inflammation with fibrosis is one of the primary pathologies of FS. Bunker et al. 4 reported the presence of active fibroblast proliferation and transformation to
myofibroblasts in capsular tissue specimens from patients with FS. Similarly, Hand et al.
5 reported chronic inflammatory changes, with fibroblastic proliferation, in capsular tissue
biopsies from patients with FS. Furthermore, Rodeo et al. 6 reported that adhesive
capsulitis involves both synovial hyperplasia and capsular fibrosis. These studies suggest that chronic inflammation causes proliferative fibrosis in the capsule of FS patients, which plays an important role in the pathogenesis of FS. Additionally, Hagiwara et al. 7
showed that the collagen density and the number of cells significantly increased in FS and the joint capsule became significantly stiffer in scanning acoustic microscopy study, with increased expression of genes associated with inflammation, fibrosis, and chondrogenesis 8. Furthermore, according to the results of shot-gun proteome analysis,
6
the upper and lower portions of the capsule express different pathologies 9. However, the
etiology of FS remains unclear.
Advanced glycation end-products (AGEs), synthesized by non-enzymatic glycation and oxidation in the Maillard reaction, accumulate in various tissues with aging
10,11. Carboxymethyllysine (CML) (Figure 1A) and pentosidine (Figure 1B) are one of the
main AGEs. Saito et al. 12 reported that collagen cross-links can be classified into two
types: 1) lysine hydroxylase and lysyl oxidase-induced links (enzymatic cross-links), and 2) glycation or oxidation-induced cross-links with AGEs (non-enzymatic links). Patients with osteoporosis exhibit significant reductions in enzymatic cross-links and increased AGE cross-cross-links in bone collagen with aging 12. Vaculik et al. 13
reported that serum and bone concentrations of pentosidine are higher in subjects with hip fractures. Additionally, Hwang et al. 14 reported that overexpression of AGEs caused
fibroblastic proliferation in FS. Considering pathological changes associated with FS, AGE cross-links and stabilization of collagen with AGE accumulation may play a potential role in the pathogenesis of FS.
The receptor of AGE (RAGE) is a transmembrane protein with three extracellular domains 15 (Figure 1C). As a non-AGE ligand, high-mobility group box 1
7
1D), has been shown to bind with RAGE 17 (Figure 1E). S100 calcium binding proteins
are a small (10-12 kDa) molecular family with calcium binding motif 18 such as S100A1
and S100B, bind with RAGE to activate calcium-dependent pathways 19 (Figure 1E).
RAGE activates reactive oxygen stress, mitogen-activated protein kinase (MAPK), and extracellular signal-regulated kinase (ERK1/2), which ultimately activates nuclear factor-κ B (NF-factor-κB), in diabetic cardiovascular endothelial cells 20 (Figure 1E). Early growth
response-1 (Egr-1), a transcription factor that is highly associated with cardiovascular complications of type 2 diabetes mellitus (DM) 21,22, is also upregulated in RAGE
signaling-mediated hypoxia (Figure 1E). NF-κB promotes the transcription of proinflammatory mediators, such as intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1) (Figure 1E). NF-κB and Egr-1 activate various inflammatory genes such as tumor necrosis factor alpha (TNFα), interleukin-6 (IL-6), and interleukin-1 beta (IL-1β) 23 (Figure 1E). These factors induce fibroblastic
inflammation and the progression of atherosclerosis 20. RAGE signaling sustains the
transcription of NF-κB, leading to prolonged inflammation and progressive chronic kidney disease 24. In chronic obstructive pulmonary disease, AGEs and HMGB1 have
been shown to increase in long-term smokers, leading to increased oxidative stress, RAGE-dependent signaling, and NF-κB gene expression 25. HMGB1 binds with RAGE,
8
Toll-like receptor 2 (TLR2), and TLR4, and activates NF-κB signaling cascades, leading to the production of cytokines, such as TNFα, IL-6, and IL-1β in macrophages, plasmacytoid dendritic cells, and B cells 26-29. S100A1, one of S100 proteins, binds RAGE
and TLR4, activating NF-κB signaling pathways, resulting in increased secretion of proinflammatory cytokines in endothelial cells and leukocytes 30. The interaction of
S100B with V domain of RAGE activates the transcription of NF-κB to promote neurite outgrowth in concert with amphoterin in the nerve system 31. Additionally, RAGE and
HMGB1 proteins and mRNA expression levels increase in patients with knee osteoarthritis, suggesting that RAGE and HMGB1 are both involved in the development or progression of this condition 32. Thankam et al. 33 reported that RAGE, HMGB1, and
triggering receptors expressed on myeloid cells-1 increase in patients with severe glenohumeral arthritis and rotator cuff injuries. However, no study has investigated the roles of RAGE, HMGB1, S100 protein, and AGEs-RAGE signaling pathway in FS.
9
3. Purpose
I could posit hypotheses as follows. Both joint stiffness and inflammation with fibrosis are the main pathologies in FS. The joint stiffness of FS is supposed to be led by AGE accumulation and AGE cross-links and stabilization of collagen. Inflammation with fibrosis is supposed to be caused by AGEs-RAGE signaling pathways. The purpose of this study was to elucidate AGEs in FS and RAGE-dependent pathways in FS to inform the development of improve treatment options.
10
4. Materials and methods
4.1 Patients and tissue collection
A retrospective case-control study was conducted. The protocols were approved by the institutional review board of Tohoku University (2011-447). To all participants it explained that various products included in samples obtained during arthroscopic surgery were evaluated and participants were free to ask any questions pertaining to the study and their involvement, after which time written informed consent was obtained from all study participants. The inclusion criteria for FS were as follows: (1) a history of a painful stiff shoulder for at least 1 month; (2) restricted passive glenohumeral joint motion, with <100° of forward flexion, <20° of external rotation, and internal rotation (fifth lumbar vertebra or lower); and (3) unremarkable radiological findings 9,34. The exclusion criteria included glenohumeral osteoarthritis, calcific
tendinitis, migrated humeral head, osteonecrosis of the humeral head, a history of traumatic events 9,34 .
The coracohumeral ligament (CHL) originates in the outer space of the coracoid process horizontal limb, and encloses the supraspinatus and subscaplaris muscles 35. Thickening of the CHL and subcoracoid fat triangle of sagittal oblique view
anterior-11
inferior glenohumeral ligament (IGHL) is a hammock-like structure with anchor points on the anterior and posterior glenoid, as well as an attachment to the proximal humeral neck 36. The rotator interval (RI) is a roughly triangular region with interposition of the
coracoid process (the base of the triangle) located between the subscapularis and supraspinatus anteriorly forming the two sides 37 (Figure 2).
Patients with rotator cuff tears (RCTs) without severe ROM restriction, as a model without inflammation in joint capsules, were selected as controls. The inclusion criteria for RCT were as follows: (1) no severe ROM limitations (>140° of forward flexion and >30° of external rotation); and (2) diagnosed by magnetic resonance imaging
9,34. The exclusion criteria included glenohumeral osteoarthritis, calcific tendinitis,
migrated humeral head, and osteonecrosis of the humeral head 9,34. The ROMs in the FS
group patients were significantly smaller than those in the RCT group in forward flexion, lateral elevation, external rotation, internal rotation, 90° abduction with external rotation, 90° abduction with internal rotation, and horizontal flexion (Figure 3A-G).
The surgical indication of FS was that the condition had failed to improve or had deteriorated after 3 months of intensive conservative treatment 9. The surgery was
performed as follows: the patient is placed in the beach-chair position under general anesthesia. A 30° arthroscope is induced throuth a standard portal, and parts of the CHL,
12 IGHL, and RI are collected (Figure 4) 39.
Tissue samples for the quantitative real-time polymerase chain reaction (qRT-PCR) experiments, the immunohistochemistry and the high-performance liquid chromatography (HPLC) analysis of CML were obtained from 14 patients with RCT (5 men and 9 women; mean age, 62.0 years; age range, 49-78 years), and 14 patients with FS (4 men and 10 women; mean age, 56.4 years; age range, 45-68 years) (Table 1). The difference in age distribution between the two groups was marginal but not statistically significant (mean difference: 5.6 years, p = 0.094 analyzed using the Student’s t-test). Samples for the HPLC analysis of pentosidine were obtained from another 19 patients with FS (7 men and 12 women; mean age, 52.6 years; age range, 25-69 years) and 11 patients with RCT (8 men and 3 women; mean age, 62.6 years; age range, 46-79 years) who satisfied the above criteria (mean difference: 10.0 years, p = 0.052 analyzed using the Student’s t-test). There were no bilateral cases.
4.2 Sample preparation, RNA extraction and purification
Three samples were obtained for each patient and for each ligament during arthroscopic surgery. One sample was cut into small pieces and the samples were fixed with 4% paraformaldehyde in 0.1 M phosphate-buffered saline, pH 7.4 supplemented
13
with 18% sucrose for cryoprotection. The paraffin-embedded tissue was cut into 5-μm sections. The hematoxylin and eosin (HE) staining was performed to compare the fibrous changes of ligaments in the FS group with those in the RCT group. Individual fibroblast-like cells were counted on three 200× field (i.e., 20× objective lens and 10× ocular lens; 0.7386 mm2 per field), by a single researcher, in a field we randomly observed 8.
Another sample was immediately placed in a vessel containing 1.5 mL QIAzol (Qiagen, Hilden, Germany) and stored in a liquid nitrogen tank until RNA extraction. The samples were then homogenized with a Polytron (Kinematica AD, Luzern, Switzerland). The total RNA of the homogenate was purified using an RNeasy Fibrous Tissue Mini Kit (Qiagen).
The other sample was preserved and kept semipermanently in a frozen state using a refrigerator at a temperature of -80 °C for high-performance liquid chromatography.
4.3 Quantitative reverse transcription-polymerase chain reaction (qRT-PCR)
Complementary DNA was synthesized using the cloned avian myeloblastosis virus first-strand cDNA synthesis kit (Invitrogen, Carlsbad, CA, USA). Gene expression was evaluated quantitatively by real-time polymerase chain reaction (PCR) using a
14
LightCycler (Roche Diagnostics, Basel, Switzerland). PCR efficiencies and the relative expression levels of RAGE, HMGB1, TLR2, TLR4, S100A1, S100B, NF-κB, Egr-1,
ICAM-1, VCAM-ICAM-1, TNFα, IL-6, and IL-1β, as a function of elongation factor 1-α 1 (EF1α1)
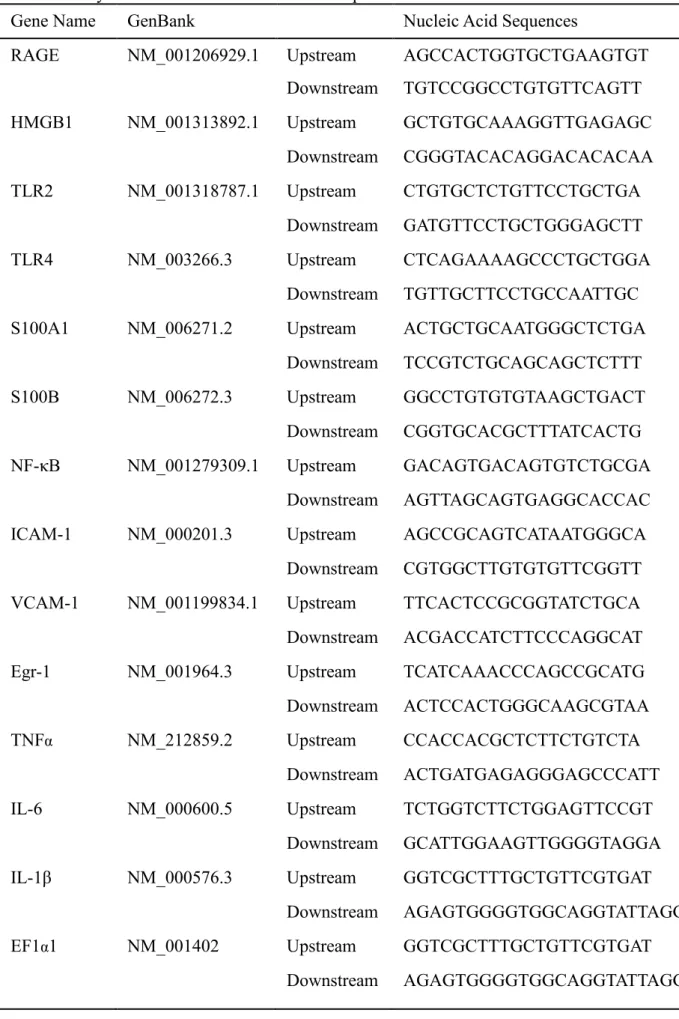
expression, were calculated as previously described 40. The primer sequences for the
expression analysis are provided in Table 2.
4.4 Immunohistochemistry
The paraffin-embedded tissue samples were cut into 5-µm sections for immunohistochemistry. The sections were deparaffinized and immersed in 3.0% hydrogen peroxide for 10 minutes. The slides were incubated with methanol at a temperature of 20 °C for 30 minutes. Endogenous immunoglobulins were blocked by incubation using 10% normal goat serum (Nichirei, Tokyo, Japan) in phosphate-buffered saline. The slides were incubated with antibodies against RAGE (ab3611, 1:100, Abcam, Cambridge, UK), CML (ab27684, 1:500, Abcam), and pentosidine (Wako, Clone No. PEN-12, Tokyo, Japan). The final detection step was carried out using 3,3′-diaminobenzidine tetrahydrochloride (Sigma-Aldrich, St Louis, MO, USA), 0.1 M imidazole, and 0.03% hydrogen peroxide. These slides were counterstained with hematoxylin. For negative controls, normal rabbit immunoglobulin G (IgG) (X0936,
15
Dako, Copenhagen, Denmark) and normal mouse IgG (X0931, Dako) were used as primary antibodies. Individual microvessels immunostained by RAGE were counted on three 200× fields (i.e., 20× objective lens and 10× ocular lens; 0.7386 mm2 per field), by
a single researcher, in a field we randomly observed 8. Immunoreactivity is evaluated as
qualitatively and pathologically positive or negative staining.
4.5 High-performance liquid chromatography (HPLC)
HPLC experiments were outsourced to the Clinical Laboratory Center of Fushimi Pharmaceutical Co. Ltd. HPLC performed according to the manufacturer’s protocols.
Samples were dried in vacuo hydrolyzed by 6N hydrochloric acid (HCl) at 120℃ for 16 hours in sealed test tubes under nitrogen. The HCl was dried in vacuo and solubilized again 250µl of distilled water. The samples after acid hydrolyzation were cleaned by solid phase extraction. The CML-KLH conjugate (100 µl) was dispensed into each well of a micro-titer plate and incubated for 24 hours at 4℃. After washing with PBS containing 0.5 ml/l Tween 20, the wells were blocked with 40% Block Ace (KAC CO., Ltd) at room temperature for 3 hours. Fifty microliters of CML antibody and 50 µl of CML (nippi CO., Ltd, Japan) standard solution or pretreated sample were added to
16
each well and incubated for 1 hour after washing; peroxidase-labeled goat anti-rabbit IgG polyclonal antibodies (DAKO, Denmark) were added and incubated for 1 hour at room temperature. Then, a color development reagent containing 0.5 mg/ml of 3,3)5,5)-tetramethylbenzidine (ScyTek Laboratories, Logan, Utah, USA) was added to each well. The reaction was stopped 10 min later by adding 100µl of TMB stop buffer (ScyTek Laboratories,Logan, Utah, USA). The absorbance was measured within 10 min at 450 nm (main wavelength) and 630 nm (reference wavelength). The standard curve was obtained by measuring standard CML solutions at 0, 0.05, 0.5, 5, and 20.0 µg/ml. 41
Decalcifying bone tissue samples were dried in vacuo and hydrolyzed by 6N HCl at 110 ℃ for 16 hours in sealed test tubes under nitrogen. The HCl was dried in vacuo and solubilized again using 50 µL distilled water. Pentosidine was quantitated based on fluorescence measurement by reverse-phase HPLC using the method described by Odetti et al. 42 The sample in 20 μL was injected into an HPLC system (Shimadzu, Japan) and
separated on a C18 reverse-phase column (Toso, Japan). The effluent was monitored with a fluorescence detector (Shimadzu, Japan) at excitation/emission wavelengths of 335/385 nm. 43,44
17
4.6 Statistical analyses
Differences between the FS and RCT groups were evaluated using the Student’s t-test for age, chi-square test for past medical history, and the Mann-Whitney-U test for qRT-PCR data, cells, and vessels count. Data are expressed as means and 95% confidence intervals (CIs). A p-value <0.05 was considered statistically significant. The statistical software package, SPSS for Windows (version 24.0, SPSS Inc., Chicago, IL, USA), was used for all analyses.
18
5. Results
5.1 HE staining of the CHLs, IGHLs, and RIs
Within the CHLs and IGHLs of the FS participants, the collagen bundles (Figure 5, asterisks) were found to be considerably denser with proliferation of spindle-shaped cells containing sharp or ellipsoidal nuclei, that were mostly fibroblast-like cells (Figure 5, arrows), compared to those from the RCT group (Figure 5A-H). In contrast, the collagen bundles were well-organized in the CHLs and IGHLs from the RCT group (Figure 5C, D, G, and H). However, the number of collagen bundles (Figure 5, asterisks) increased in the RIs from the FS group compared to the RCT group (Figure 5I and J). Proliferation of spindle-shaped cells (Figure 5, arrows) was observed in the RIs from the RCT group (Figure 5K and L).
The number of fibroblast-like cells was significantly higher in the CHLs (368 range; 168-659and IGHLs (446 range; 168-811from the FS group than those from the RCT group in the CHLs (220 range; 90-553 and IGHLs (136 range; 27-258(CHL, p = 0.015; IGHL, p < 0.001) (Figure 6). However, there were no significant differences in the number of fibroblast-like cells in the RIs from the FS group (359 range; 213-781compared to those from the RCT group (340 range; 132-527 (p = 0.968) (Figure 6).
19
5.2 Immunohistochemistry of RAGE and AGEs
RAGE immunoreactivity was more strongly positive in the CHLs and IGHLs from the FS group than those from the RCT group (Figure 7A-H). Photomicrographs of shoulder capsules immunostained for RAGE revealed moderate RAGE immunoreactivity, with brown staining primarily in blood vessels (Figure 7, asterisks) in the CHLs and IGHLs from the FS group (Figure 7A, B, E, and F). RAGE immunostaining was focal positive in the CHLs and IGHLs from the RCT group (Figure 7C, D, G, and H). However, there seemed to be positive immunoreactivity in the RIs from the FS group compared to those from the RCT group (Figure 7I and J), a large number of immunostained blood vessels (Figure 7, asterisks) were observed in the RIs from the RCT group (Figure 7K and L).
The number of immunostained blood vessels was significantly higher in the CHLs (50 range; 9-69 and IGHLs (21 range; 1-66 from the FS group than those in the CHLs (72 range; 26-134 and IGHLs (25 range; 7-50 from the RCT group (CHL, p = 0.001; IGHL, p < 0.001) (Figure 8). However, there were no significant differences in the number of immunostained blood vessels in the RIs (65 range; 58-72from the FS group compared to those from the RCT group (66 range; 60-71 (p = 0.932) (Figure 8).
20
Photomicrographs of shoulder capsules immunostained for CML revealed strongly positive CML immunoreactivity, with brown staining in fibrous tissue, in the CHLs from the FS group (Figure 9A and B). The IGHLs from the FS group were also strongly positive immunostained for CML (Figure 9E and F). The RIs from the FS group and were positive immunostained for CML (Figure 9I and J). In comparison with the samples from the FS group, CML immunostaining was focal positive in the CHLs, IGHLs, and RIs (Figure 9C, D, G, H, K and L).
Photomicrographs of shoulder capsules immunostained for pentosidine revealed almost negative pentosidine immunoreactivity in the CHLs from the FS group (Figure 10A and B). The IGHLs and RIs from the FS group and the CHLs, IGHLs, and RIs from the RCT group had completely negative pentosidine immunostaining (Figure 10C-L).
5.3 Gene expression levels related to RAGE pathways
The gene expression levels of the factors related to the RAGE-dependent pathway, such as RAGE, HMGB1, S100A1, S100B, TLR2, TLR4, NF-κB, ICAM-1,
VCAM-1, TNFα, IL-6 and IL-1β (Figure 11), were significantly greater in the CHLs from the FS group (RAGE; 5.3 HMGB1; 9.3 S100A1; 7.1 , S100B; 4.0
21
, TNFα; 5.2 , IL-6; 4.8 IL-1β; 5.7 ) compared to those from the RCT group (RAGE; 1.0 HMGB1; 1.5 S100A1; 1.2 , S100B; 0.7 TLR2; 1.5 TLR4; 0.9 , NF-κB; 0.9 , ICAM-1; 0.8 , VCAM-1; 1.0 ,
TNFα; 1.2 , IL-6; 1.5 IL-1β; 1.3 ) (RAGE; p = 0.005, HMGB1; p = 0.008
S100A1; p = 0.001, S100B; p = 0.001 TLR2; p = 0.039 TLR4; p = 0.021, NF-κB; p =
0.001, ICAM-1; p = 0.009, VCAM-1; p = 0.006, TNFα; p = 0.016, IL-6; p = 0.044, IL-1β; p = 0.011). However, there were no significant differences in Egr-1 gene expression levels in the CHLs from the FS group (4.9 ) and RCT group (2.0 p = 0.12. (Figure 11H).
The gene expression levels were also significantly greater in the IGHLs (Figure 12) from the FS group (RAGE; 1.1 HMGB1; 1.9 S100A1; 3.3 ,
S100B; 1.5 TLR2; 1.5 TLR4; 1.2 , NF-κB; 1.1 , Egr-1; 1.6
ICAM-1; 1.5 , TNFα; 6.3 , IL-6; 11.4 IL-1β; 6.8 ) compared to those from the RCT group (RAGE; 0.8 HMGB1; 0.5 S100A1; 1.3 ,
S100B; 0.7 TLR2; 0.6 TLR4; 0.7 , NF-κB; 0.7 , Egr-1; 0.5
ICAM-1; 0.7 , TNFα; 2.0 , IL-6; 2.5 IL-1β; 2.4 ) (RAGE; p = 0.016, HMGB1; p < 0.001 S100A1; p = 0.008, S100B; p = 0.012 TLR2; p = 0.004 TLR4; p = 0.027, NF-κB; p = 0.035, Egr-1; p = 0.007, ICAM-1; p = 0.011, TNFα; p = 0.004,
IL-22
6; p < 0.0001, IL-1β; p = 0.001). However, there were no significant differences in VCAM-1 gene expression levels in the IGHLs from the FS group (VCAM-1.5 ) and RCT group (0.9
p = 0.21. (Figure 12J).
The gene expression levels were significantly greater in the RIs (Figure 13) from the FS group (TNFα; 3.1 , IL-6; 3.1 ) compared to those from the RCT group (TNFα; 1.0 , IL-6; 1.0) (TNFα; p = 0.035, IL-6; p = 0.027) (Figure 13K and L). However, there were no significant differences in RAGE HMGB1 S100A1,
S100B TLR2 TLR4, NF-κB, Egr-1ICAM-1, VCAM-1 and IL-1β in the IGHLs from the
FS group (RAGE; 2.3 HMGB1; 1.3 S100A1; 2.2 , S100B; 1.9
TLR2; 1.9 TLR4; 1.2 , NF-κB; 1.3 , Egr-1; 2.0 ICAM-1; 1.5 , VCAM-1; 2.3 IL-1β; 2.0 ) and the RCT group (RAGE; 1.4 HMGB1; 0.9
S100A1; 1.1 , S100B; 1.7 TLR2; 1.1 TLR4; 1.3 , NF-κB; 1.3 , Egr-1; 1.1 ICAM-1; 1.1 , VCAM-1; 1.7 IL-1β; 1.6 RAGE; p = 0.352, HMGB1; p = 0.265 S100A1; p = 0.329, S100B; p = 0.551 TLR2; p = 0.769
TLR4; p = 0.482, NF-κB; p = 0.511, Egr-1; p = 0.220, ICAM-1; p = 0.846; VCAM-1; p =
23
5.4 HPLC of the CML and pentosidine
Expression of the CML level in HPLC was significantly higher in the CHLs
(20.8 μg/g) from the FS group compared to those from the RCT group (15.0 μg/g) (p = 0.011) (Figure 14A). Similarly, the CML level was also significantly higher in the
IGHLs (21.6 μg/g) from the FS group than in those from the RCT group (15.2 μg/g) (p = 0.008) (Figure 14B). However, there were no significant differences in the RIs (20.6 μg/g) from the FS group than in those from the RCT group (19.7 μg/g)p = 0.949. (Figure 14C).
In contrast, there were no significant differences of pentosidine levels in the
CHLs (0.79 μg/g), IGHLs (1.47 μg/g), and RIs (1.13 μg/g) from the
FS group than in the CHLs (1.13 μg/g), IGHLs (1.95 μg/g), and RIs (0.85 μg/g) from the RCT group (CHLs; p = 0.792, IGHLs; p = 0.349, RIs; p = 0.232) (Figure 15)
24
6. Discussion
One of the most important findings of the present study was that CML, rather than pentosidine, accumulated in joint capsules of frozen shoulder, indicationg that AGE accumulation and AGE cross-links and stabilization of collagen are supposed to lead joint stiffness of FS. The other important finding of the present study was that RAGE, HMGB1, and S100 genes and NF-κB signaling pathways were activated in joint capsules of FS, indicating that AGEs-RAGE signaling pathways are supposed to play important roles in the pathogenesis of FS, leading inflammation with fibrosis in joint capsules of FS.
The cause of joint stiffness was classified into 2 components: arthrogenic (bone, cartilage, synovial membrane, capsule, and ligament) and myogenic (muscle, tendon, and fascia) 45. Myogenic components are respond well with rehabilitation,
however, the joint capsule does not. Therefore, surgical intervention, such as arthroscopic pancapsular release, should be considered for recalcitrant cases of FS46,47.
A thickened CHL has been documented as one of the most specific manifestations of FS 48-50. Although a thickened CHL clearly limits the external rotation
of the shoulder joint 48-50, it also severely restricts the ROM in various directions 34, and
release of the entire CHL renders it possible to regain the full ROM in FS 39. The CHL is
25
the greater tuberosity; the other attaches the subscapularis and supraspinatus tendons 35.
Disturbance of the sliding mechanism of these tendons might result in ROM restriction
34. Although data is lacking regarding IGHL effects on the ROM in FS, release of the
IGHL could aid in regaining the ROM. Both ligaments have important roles in the ROM restriction in FS.
Hagiwara et al 8, collagen bundles were dense with less space in the RIs from FS,
on the other hand, the bundles were well-organized in the RIs from RCT. The number of cells were significantly higher in the RIs from the FS group compared to those from the RCT group 8. The results might suggest that the accumulation of AGEs and RAGE, and
the gene expressions of RAGE-dependent pathways in the RIs, as well as CHLs and IGHLs, could be significantly higher in the FS group than in the RCT group. However, there seemed to be more collagen bundles in the RIs from the FS group than those in the RCT group; proliferation of spindle-shaped cells was observed in the RIs from the RCT group in this study. The gene expression levels were not significantly greater in the RIs from the FS group when compared to those from the RCT group. This may be due to as follows; (1) RI capsule wasconsidered as the predominant area of FS 50. Furthermore,
ROM in the shoulder joint decreased with aging and joint capsule including the RI and CHL was one of the main causes of restricted ROM 8. Okuno et al. 51 reported that
26
abnormal vessels around the RI might be a cause of pain, and transcatheter arterial embolization for FS and RCT was a possible treatment option that had failed to improve with conservative treatments. The RI pathology must be a natural aging process, which would causes no significant difference in the RIs in the present study. Additional inflammation could accelerate fibrosis or induce pain in FS.; (2) Another reason was due to individual differences in conservative treatments, tear size, duration of onset, or the aging process of the RCT group 52,53. Huri et al. 54 reported that anterior superior lesion
of rotator cuff tear involved the subscapularis and anterior portion of the supraspinatus tendon and adjacent RI structures. Furthermore, the timing of sample collection relating to the disease stage could affect the results. Conservative treatment was revealed to be effective in 73-80% of cases with full-thickness tears to the rotator cuff 55. Mosca et al. 56
reported that HMGB1 was significantly increased in post-treatment pain-free rotator cuff tendon compared with a healthy shoulder and painful diseased shoulder. The timing of collecting samples from the RI would affect the results from this study.
In the present study, collagen bundle density and the number of cells in the CHLs, IGHLs, and RIs in HE staining from the FS group was apparently higher than in those from the RCT group, which was similar to the previous report 8. Hwang et al. 14
27
hypercellularity, and increased vascularization. In addition, the immunoreactivity of AGEs in the FS capsule was mainly seen around vascular areas (CD34 positive), and immunostaining of AGEs in the RCT group was negative 14. In the present study, the
immunoreactivity of RAGE was also observed around small blood vessels in the CHLs, IGHLs, and RIs in both the FS and RCT groups. The differences might be due to differences of evaluated AGEs, as details regarding the specific AGEs involved were not revealed in the study conducted by Hwang et al. 14. In the present study, the
immunoreactivity of CML was diffuse in the extracellular matrix, as well as small blood vessels, in the CHLs, IGHLs, and RIs from the FS group, suggesting that CML could be the primary cause of RAGE proliferation.
In the present study, CML was more strongly accumulated in the CHLs and IGHLs from patients with FS than in those from patients with RCT. However, pentosidine was not observed in the CHLs and IGHLs from patients with FS or RCT. In contrast, several previous reports have shown increased pentosidine in the joint capsule under various conditions. Lee et al. 57 reported that pentosidine was present in the knee joint
capsule of rat knees that were immobilized for 16 weeks by internal fixation. The difference in results between this previous and the present studies might be due to differences in species (rodent vs human) or FS pathogenesis (immobilization vs various
28
causes). Holte et al. 58 reported an association between shoulder joint stiffness and
pentosidine; AGEs in skin collagen were assessed using punch biopsy and autofluorescence. The difference in results between this previous study and the present study might be due to the difference in the type of samples collected. Pentosidine is a pentose-derived fluorescent cross-link formed between arginine and lysine residues in collagen 59, and its concentration increases gradually with age in human bone collagen 12.
Pentosidine might be associated with age-related changes, therefore there is no significant difference in the accumulation of pentosidine in the joint capsules from FS and RCT in the present study. On the other hand, CML is a glycooxidation product generated by oxidization of glycated modified protein lysine residues by superoxide or hydroxyl radical, as well as a product during oxidation of both carbohydrates and lipids through glyoxal, as one intermediate in the formation of CML from glucose 60. CML is an
indicator of long-term oxidative stress accumulation, such as accumulation in skin collagen with aging, atherosclerosis, diabetes, and Alzheimer disease 60,61.
Cardiovascular injury or diabetic hyperglycemia leads to the storage of AGEs, such as CML, in vascular endothelial cells, and AGEs bind to RAGEs on these cells 62-64,
triggering the activation of NF-κB by interacting with MAPK and ERK1/2 signaling pathways 65,66. NF-κB transmigrates to the nucleus and stimulates the transcription of
29
target genes; endothelial cells subsequently release cytokines, such as TNFα, IL-6, ICAM-1, and VCAM-1 67,68, leading to endothelial damage. In this study, the gene
expression levels of RAGE, NF-κB, and related cytokines were significantly greater in FS capsules than in RCT capsules. This indicates that the RAGE-dependent NF-κB signaling pathway is greatly expressed in the FS capsule. FS incidence is estimated at 3–5% in the general population and up to 20% in those with DM 69. DM coupled with severely
restricted joint motion on the first visit were poor prognostic factors for FS 70. Patients
with DM expressed MAPK, NF-κB, matrix metalloproteinase-3, IL-6, and vascular endothelial growth factor to induce fibrous tissue in the area of the mechanical stress, such as the CHL and long head biceps 71.
Poor posture, including a forward-leaning head, internal rotation of the scapula, and hyper-kyphosis of the thoracic spine, is common in patients with FS. Internal rotation of the scapula due to poor posture creates an ischemic condition in the joint capsule, induced by decreased blood supply via the anterior humeral circumflex artery 72. These
circumstances might lead to angiogenesis, infiltration of inflammatory cells, and the expression of cytokines in the capsule of patients with idiopathic FS 8. Zwang et al. 28
reported that the HMGB1-RAGE/TLR-TNFα pathway is upregulated in renal tissue from hypoxia rat models. Thus, angiogenesis by hypoxia might play an important role in
30
activating the RAGE-dependent NF-κB signaling pathway in neovascularized vessels, leading to inflammation in the FS capsule.
Egr-1 is a major transcription factor expressed in smooth muscle cells, fibroblasts, leukocytes, and endothelial cells, 73 and is highly expressed in response to
hypoxia, inflammation, oxidative stress, growth factors, and vascular injury 73. Hypoxia
stimulates rapid production of AGEs, leading to RAGE signaling pathway and up-regulation of Egr-1 74. Egr-1, with hyperglycemic conditions, promotes proinflammatory
gene expression, such as ICAM-1 75, IL-6, TNFα, 76 77. In the present study, Egr-1 was
more strongly accumulated in the IGHLs in FS. Transforming growth factor beta causes a time- and dose- dependent increase in Egr-1 protein in normal fibroblasts 78. These
reports suggest that IGHL in FS might be under hypoxia, leading that Egr-1 activates progression of fibrosis of IGHL in FS.
Glutathione metabolism, which works as an antioxidant defense process, is downregulated in the IGHLs of patients with FS 9. Our results indicate that Egr-1 was
more strongly accumulated in the IGHLs in FS. The RAGE signaling pathway, which increases reactive oxygen stress and activates NF-κB 20 and Egr-1, is associated with
glutathione metabolism. Moreover, the ICAM-1 gene expression level was significantly higher in both CHLs and IGHLs from patients with FS compared to those from patients
31
with RCT. However, gene expression of VCAM-1 was significantly higher in only the CHLs (not the IGHLs) from patients with FS as opposed to those from patients with RCT. This suggests that the CHL might be under most serious inflammation in FS. FS pathophysiology differs between the upper parts (the RI and middle glenohumeral ligament) and lower parts (the IGHL) of the capsule 9. Differences in blood supply and
tensile stress during shoulder motion could be related to differences between the CHL and IGHL in FS.
RAGE is classified as a pattern recognition receptor (PRR) 79, and binds with
various ligands, such as AGEs, HMGB1, S100 protein, amphoterin, and amyloid-beta peptide 80. PRRs interact with damage-associated molecular patterns (DAMPs), which
initiate inflammation for tissue repair in life-threatening stress 81. HMGB1, which is a
DAMP, binds with RAGE, TLR2, and TLR4, and activates NF-κB signaling cascades, leading to the production of cytokines, such as TNFα, IL-6, and IL-1β in macrophages, plasmacytoid dendritic cells, and B cells 26-29. Endothelial cells stimulated with S100B
activate NF-κB signaling pathways and up-regulate the expression of VCAM-1, with increased TNFα 82. S100B, which is expressed in skeletal muscle 83,84, stimulates
myoblast proliferation and inhibits myogenic differentiation by RAGE-dependent pathways in skeletal muscle cells 84. Vascular injury promotes S100B expression in
32
vascular smooth muscle cells, activating RAGE signaling pathways, increasing NF-κB expression and its nuclear translocation, and promoting vascular remodeling with proliferation of vascular smooth muscle cells 85. The results of the present study could be
regarded as inflammation associated with AGEs-RAGE signaling pathways, rather than general chronic inflammation. AGEs-RAGE signaling pathways as a reaction with DAMPs and PRP, might not be specific to joint capsules of FS. Specific conditions might occur in the FS capsules.
Specific agents have been shown to inhibit AGEs-RAGE signaling pathways. For example, a soluble RAGE (sRAGE), recognized as a splice variant containing all extracellular domains, however, devoid of the transmembrane and intra-cytoplasmic domains, was found to bind RAGE ligands and to effectively prevent the adverse effects associated with RAGE signaling 10. Further, treatment with sRAGE reduced
atherosclerosis in diabetic mice 86. Hence, it was highly expected that both in vitro and in vivo administration of sRAGE would reduce AGE/RAGE-mediated complications, such
as atherosclerosis and insulin resistance 87,88. Wang et al. 89 reported that in patients with
acute coronary syndrome, sRAGE plasma levels increased to assist in the reduction of inflammation during myocardial compensatory protection, suggesting that sRAGE may serve as a predictor of severe coronary heart disease. Moreover, Oh et al. 90 reported that
33
treatment with sRAGE-secreting mesenchymal stem cells prevented neuronal cell death within an RAGE-signaling amyloid β-induced rat brain model, suggesting that it may appreciably improve the effectiveness of cell therapy in Alzheimer disease. In addition, sRAGE may prevent activation of AGEs-RAGE-induced carcinogenesis pathways such as ERK1/2 91. Other RAGE inhibitors have also proven effective. Specifically, FPS-ZM1,
a nontoxic RAGE-specific inhibitor, may elicit neuroprotective effects through attenuating microglial activation, oxidative stress and inflammation by blocking RAGE in rat primary microglia cells 92. Further, Cheng et al. 93 reported that irbesartan, an
angiotensin 2 receptor blocker, may have a protective role in diabetes-related bone damage by blocking the deleterious effects of AGEs/RAGE-mediated oxidative stress. Finally, Wang et al. 94 reported that glycine, the simplest amino acid, suppressed the
AGE/RAGE signaling pathway and inhibited generation of reactive oxygen species in the aorta of streptozotocin-induced diabetic rats and in human umbilical vascular endothelial cells. However, no studies have reported on the role of AGEs-RAGE inhibitors in human shoulder capsules. Nevertheless, these products may serve as key drugs in the suppression of human FS.
AGEs increase in multiple tissues in DM and contribute to end-organ disease and activate RAGE signaling pathways 20-22,95,96. However, the association between AGEs and
34
other metabolic diseases are still unknown. In dyslipidemia, AGEs level was significantly elevated in obesity fatty liver model rats, which also had higher total cholesterol, triglyceride, low-density lipoprotein and high-density lipoprotein levels, along with deteriorated liver function and higher TNF-α and IL-6 levels 97. AGEs affected cholesterol
homeostasis by impairing gene expression on macrophages in chronic injection of galactose-treated mice 98. However, AGEs with dyslipidemia in human have not yet
reported. In hypothyroidism, RAGE revealed a mechanism regulating thyroid hormone secretion in blood samples from patients with type 2 DM-hypothyroidism compared with type 2 DM 99. Significant association of the RAGE system with Hashimoto's thyroiditis
was found only with regard to the prevalence of the -429T>C type of RAGE polymorphism, but not with -374T>A polymorphism 100. In the present study, there were
no significant differences between the two groups in dyslipidemia and hypothyroidism. Further studies are needed to clarify the association between AGEs and metabolism in joint capsules of FS.
The current study has several limitations to acknowledge: (1) the sample size was small; (2) conservative treatment before the visit to our hospital was insufficiently evaluated; (3) normal shoulder capsules were not evaluated; (4) patients in FS group had arthroscopic capsular release after appropriate conservative treatments, so all samples
35
were obtained at the end stage of FS, and mechanisms from its onset to the end stage were not evaluated; (5) there remains the possibility of an influence of natural aging process in the joint capsule; and (6) association between metabolic diseases and FS was not evaluated.
36
7. Conclusion
The results of the present study indicated that in AGEs, CML, rather than pentosidine, exerted a more pronounced effect on FS pathology. AGEs, HMGB1, and S100 protein are supposed to bring inflammation with fibrosis in FS, by binding to RAGE and activating NF-κB signaling pathways. Hence, suppression of these pathways could provide a potential treatment option for FS.
37
8. References
1. Codman EA. Rupture of the Supraspinatus Tendon and Other Lesions in or about the Subacromial Bursa. The Shoulder 1934.
2. Reeves B. The natural history of the frozen shoulder syndrome. Scandinavian journal of rheumatology 1975;4:193-6.
3. Wong CK, Levine WN, Deo K, et al. Natural history of frozen shoulder: fact or fiction? A systematic review. Physiotherapy 2017;103:40-7.
4. Bunker TD, Anthony PP. The pathology of frozen shoulder. A Dupuytren-like disease. The Journal of bone and joint surgery British volume 1995;77:677-83.
5. Hand GC, Athanasou NA, Matthews T, Carr AJ. The pathology of frozen shoulder. The Journal of bone and joint surgery British volume 2007;89:928-32.
6. Rodeo SA, Hannafin JA, Tom J, Warren RF, Wickiewicz TL. Immunolocalization of cytokines and their receptors in adhesive capsulitis of the shoulder. Journal of orthopaedic research : official publication of the Orthopaedic Research Society 1997;15:427-36.
7. Hagiwara Y, Ando A, Chimoto E, Saijo Y, Ohmori-Matsuda K, Itoi E. Changes of articular cartilage after immobilization in a rat knee contracture model. Journal of orthopaedic research : official publication of the Orthopaedic Research Society 2009;27:236-42.
8. Hagiwara Y, Ando A, Onoda Y, et al. Coexistence of fibrotic and chondrogenic process in the capsule of idiopathic frozen shoulders. Osteoarthritis and cartilage 2012;20:241-9.
9. Hagiwara Y, Mori M, Kanazawa K, et al. Comparative proteome analysis of the capsule from patients with frozen shoulder. Journal of shoulder and elbow surgery 2018;27:1770-8. 10. Singh R, Barden A, Mori T, Beilin L. Advanced glycation end-products: a review. Diabetologia 2001;44:129-46.
11. Tessier FJ. The Maillard reaction in the human body. The main discoveries and factors that affect glycation. Pathologie-biologie 2010;58:214-9.
12. Saito M, Marumo K. Collagen cross-links as a determinant of bone quality: a possible explanation for bone fragility in aging, osteoporosis, and diabetes mellitus. Osteoporosis international : a journal established as result of cooperation between the European Foundation for Osteoporosis and the National Osteoporosis Foundation of the USA 2010;21:195-214.
13. Vaculik J, Braun M, Dungl P, Pavelka K, Stepan JJ. Serum and bone pentosidine in patients with low impact hip fractures and in patients with advanced osteoarthritis. BMC musculoskeletal disorders 2016;17:308.
14. Hwang KR, Murrell GA, Millar NL, Bonar F, Lam P, Walton JR. Advanced glycation end products in idiopathic frozen shoulders. Journal of shoulder and elbow surgery 2016;25:981-8. 15. Neeper M, Schmidt AM, Brett J, et al. Cloning and expression of a cell surface receptor
38
for advanced glycosylation end products of proteins. The Journal of biological chemistry 1992;267:14998-5004.
16. Huttunen HJ, Rauvala H. Amphoterin as an extracellular regulator of cell motility: from discovery to disease. Journal of internal medicine 2004;255:351-66.
17. Hori O, Brett J, Slattery T, et al. The receptor for advanced glycation end products (RAGE) is a cellular binding site for amphoterin. Mediation of neurite outgrowth and co-expression of rage and amphoterin in the developing nervous system. The Journal of biological chemistry 1995;270:25752-61.
18. Marenholz I, Heizmann CW, Fritz G. S100 proteins in mouse and man: from evolution to function and pathology (including an update of the nomenclature). Biochemical and biophysical research communications 2004;322:1111-22.
19. Ostendorp T, Leclerc E, Galichet A, et al. Structural and functional insights into RAGE activation by multimeric S100B. The EMBO journal 2007;26:3868-78.
20. Fukami K, Yamagishi S, Okuda S. Role of AGEs-RAGE system in cardiovascular disease. Current pharmaceutical design 2014;20:2395-402.
21. Yan SF, Harja E, Andrassy M, Fujita T, Schmidt AM. Protein kinase C beta/early growth response-1 pathway: a key player in ischemia, atherosclerosis, and restenosis. Journal of the American College of Cardiology 2006;48:A47-55.
22. Vedantham S, Thiagarajan D, Ananthakrishnan R, et al. Aldose reductase drives hyperacetylation of Egr-1 in hyperglycemia and consequent upregulation of proinflammatory and prothrombotic signals. Diabetes 2014;63:761-74.
23. Yao J, Mackman N, Edgington TS, Fan ST. Lipopolysaccharide induction of the tumor necrosis factor-alpha promoter in human monocytic cells. Regulation by Egr-1, c-Jun, and NF-kappaB transcription factors. The Journal of biological chemistry 1997;272:17795-801.
24. Koyama H, Nishizawa Y. AGEs/RAGE in CKD: irreversible metabolic memory road toward CVD? European journal of clinical investigation 2010;40:623-35.
25. Gangemi S, Casciaro M, Trapani G, et al. Association between HMGB1 and COPD: A Systematic Review. Mediators of inflammation 2015;2015:164913.
26. Sha Y, Zmijewski J, Xu Z, Abraham E. HMGB1 develops enhanced proinflammatory activity by binding to cytokines. Journal of immunology (Baltimore, Md : 1950) 2008;180:2531-7. 27. Urbonaviciute V, Furnrohr BG, Meister S, et al. Induction of inflammatory and immune responses by HMGB1-nucleosome complexes: implications for the pathogenesis of SLE. The Journal of experimental medicine 2008;205:3007-18.
28. Zhang C, Dong H, Chen F, Wang Y, Ma J, Wang G. The
HMGB1-RAGE/TLR-TNF-alpha signaling pathway may contribute to kidney injury induced by hypoxia. Experimental and therapeutic medicine 2019;17:17-26.
39
29. Rosenberg JH, Rai V, Dilisio MF, Agrawal DK. Damage-associated molecular patterns in the pathogenesis of osteoarthritis: potentially novel therapeutic targets. Molecular and cellular biochemistry 2017;434:171-9.
30. Hofmann MA, Drury S, Fu C, et al. RAGE mediates a novel proinflammatory axis: a central cell surface receptor for S100/calgranulin polypeptides. Cell 1999;97:889-901.
31. Huttunen HJ, Kuja-Panula J, Sorci G, Agneletti AL, Donato R, Rauvala H. Coregulation of neurite outgrowth and cell survival by amphoterin and S100 proteins through receptor for advanced glycation end products (RAGE) activation. The Journal of biological chemistry 2000;275:40096-105.
32. Sun XH, Liu Y, Han Y, Wang J. Expression and Significance of High-Mobility Group Protein B1 (HMGB1) and the Receptor for Advanced Glycation End-Product (RAGE) in Knee Osteoarthritis. Medical science monitor : international medical journal of experimental and clinical research 2016;22:2105-12.
33. Thankam FG, Dilisio MF, Dietz NE, Agrawal DK. TREM-1, HMGB1 and RAGE in the
Shoulder Tendon: Dual Mechanisms for Inflammation Based on the Coincidence of Glenohumeral Arthritis. PloS one 2016;11:e0165492.
34. Hagiwara Y, Sekiguchi T, Ando A, et al. Effects of Arthroscopic Coracohumeral Ligament Release on Range of Motion for Patients with Frozen Shoulder. The open orthopaedics journal 2018;12:373-9.
35. Arai R, Nimura A, Yamaguchi K, et al. The anatomy of the coracohumeral ligament and its relation to the subscapularis muscle. Journal of shoulder and elbow surgery 2014;23:1575-81. 36. Passanante GJ, Skalski MR, Patel DB, et al. Inferior glenohumeral ligament (IGHL) complex: anatomy, injuries, imaging features, and treatment options. Emergency radiology 2017;24:65-71.
37. Tamborrini G, Moller I, Bong D, et al. The Rotator Interval - A Link Between Anatomy and Ultrasound. Ultrasound international open 2017;3:E107-e16.
38. Kanazawa K, Hagiwara Y, Kawai N, et al. Correlations of coracohumeral ligament and range of motion restriction in patients with recurrent anterior glenohumeral instability evaluated by magnetic resonance arthrography. Journal of shoulder and elbow surgery 2017;26:233-40. 39. Hagiwara Y, Ando A, Kanazawa K, et al. Arthroscopic Coracohumeral Ligament Release for Patients With Frozen Shoulder. Arthroscopy techniques 2018;7:e1-e5.
40. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic acids research 2001;29:e45.
41. Takeuchi M, Makita Z, Yanagisawa K, Kameda Y, Koike T. Detection of noncarboxymethyllysine and carboxymethyllysine advanced glycation end products (AGE) in serum of diabetic patients. Molecular medicine (Cambridge, Mass) 1999;5:393-405.
40
42. Odetti P, Fogarty J, Sell DR, Monnier VM. Chromatographic quantitation of plasma and erythrocyte pentosidine in diabetic and uremic subjects. Diabetes 1992;41:153-9.
43. Takahashi M, Hoshino H, Kushida K, Kawana K, Inoue T. Direct quantification of pentosidine in urine and serum by HPLC with column switching. Clinical chemistry 1996;42:1439-44.
44. Sell DR, Monnier VM. Structure elucidation of a senescence cross-link from human extracellular matrix. Implication of pentoses in the aging process. The Journal of biological chemistry 1989;264:21597-602.
45. Trudel G, Uhthoff HK. Contractures secondary to immobility: is the restriction articular or muscular? An experimental longitudinal study in the rat knee. Archives of physical medicine and rehabilitation 2000;81:6-13.
46. Kelley MJ, McClure PW, Leggin BG. Frozen shoulder: evidence and a proposed model guiding rehabilitation. The Journal of orthopaedic and sports physical therapy 2009;39:135-48. 47. Buchbinder R, Youd JM, Green S, et al. Efficacy and cost-effectiveness of physiotherapy following glenohumeral joint distension for adhesive capsulitis: a randomized trial. Arthritis and rheumatism 2007;57:1027-37.
48. Izumi T, Aoki M, Tanaka Y, et al. Stretching positions for the coracohumeral ligament: Strain measurement during passive motion using fresh/frozen cadaver shoulders. Sports medicine, arthroscopy, rehabilitation, therapy & technology : SMARTT 2011;3:2.
49. Neer CS, 2nd, Satterlee CC, Dalsey RM, Flatow EL. The anatomy and potential effects of contracture of the coracohumeral ligament. Clinical orthopaedics and related research 1992:182-5.
50. Ozaki J, Nakagawa Y, Sakurai G, Tamai S. Recalcitrant chronic adhesive capsulitis of the shoulder. Role of contracture of the coracohumeral ligament and rotator interval in pathogenesis and treatment. The Journal of bone and joint surgery American volume 1989;71:1511-5.
51. Okuno Y, Iwamoto W, Matsumura N, et al. Clinical Outcomes of Transcatheter Arterial Embolization for Adhesive Capsulitis Resistant to Conservative Treatment. Journal of vascular and interventional radiology : JVIR 2017;28:161-7.e1.
52. Clarke GR, Willis LA, Fish WW, Nichols PJ. Preliminary studies in measuring range of motion in normal and painful stiff shoulders. Rheumatology and rehabilitation 1975;14:39-46. 53. Sun Y, Zhang P, Liu S, et al. Intra-articular Steroid Injection for Frozen Shoulder: A Systematic Review and Meta-analysis of Randomized Controlled Trials With Trial Sequential Analysis. The American journal of sports medicine 2017;45:2171-9.
54. Huri G, Kaymakoglu M, Garbis N. Rotator cable and rotator interval: anatomy, biomechanics and clinical importance. EFORT open reviews 2019;4:56-62.
41
orthopaedic science : official journal of the Japanese Orthopaedic Association 2013;18:197-204. 56. Mosca MJ, Carr AJ, Snelling SJB, Wheway K, Watkins B, Dakin SG. Differential expression of alarmins-S100A9, IL-33, HMGB1 and HIF-1alpha in supraspinatus tendinopathy before and after treatment. BMJ open sport & exercise medicine 2017;3:e000225.
57. Lee S, Sakurai T, Ohsako M, Saura R, Hatta H, Atomi Y. Tissue stiffness induced by prolonged immobilization of the rat knee joint and relevance of AGEs (pentosidine). Connective tissue research 2010;51:467-77.
58. Holte KB, Juel NG, Brox JI, et al. Hand, shoulder and back stiffness in long-term type 1 diabetes; cross-sectional association with skin collagen advanced glycation end-products. The Dialong study. Journal of diabetes and its complications 2017;31:1408-14.
59. Dyer DG, Blackledge JA, Thorpe SR, Baynes JW. Formation of pentosidine during nonenzymatic browning of proteins by glucose. Identification of glucose and other carbohydrates as possible precursors of pentosidine in vivo. The Journal of biological chemistry 1991;266:11654-60.
60. Fu MX, Requena JR, Jenkins AJ, Lyons TJ, Baynes JW, Thorpe SR. The advanced glycation end product, Nepsilon-(carboxymethyl)lysine, is a product of both lipid peroxidation and glycoxidation reactions. The Journal of biological chemistry 1996;271:9982-6.
61. Kislinger T, Fu C, Huber B, et al. N(epsilon)-(carboxymethyl)lysine adducts of proteins are ligands for receptor for advanced glycation end products that activate cell signaling pathways and modulate gene expression. The Journal of biological chemistry 1999;274:31740-9.
62. Boelke E, Storck M, Buttenschoen K, Berger D, Hannekum A. Endotoxemia and mediator release during cardiac surgery. Angiology 2000;51:743-9.
63. Mazzini GS, Schaf DV, Oliveira AR, et al. The ischemic rat heart releases S100B. Life sciences 2005;77:882-9.
64. Villegas-Rodriguez ME, Uribarri J, Solorio-Meza SE, et al. The AGE-RAGE Axis and Its Relationship to Markers of Cardiovascular Disease in Newly Diagnosed Diabetic Patients. PloS one 2016;11:e0159175.
65. Ehlermann P, Eggers K, Bierhaus A, et al. Increased proinflammatory endothelial response to S100A8/A9 after preactivation through advanced glycation end products. Cardiovascular diabetology 2006;5:6.
66. Luan ZG, Zhang H, Yang PT, Ma XC, Zhang C, Guo RX. HMGB1 activates nuclear factor-kappaB signaling by RAGE and increases the production of TNF-alpha in human umbilical vein endothelial cells. Immunobiology 2010;215:956-62.
67. Tanaka N, Yonekura H, Yamagishi S, Fujimori H, Yamamoto Y, Yamamoto H. The receptor for advanced glycation end products is induced by the glycation products themselves and tumor necrosis factor-alpha through nuclear factor-kappa B, and by 17beta-estradiol through
Sp-42
1 in human vascular endothelial cells. The Journal of biological chemistry 2000;275:25781-90. 68. Kierdorf K, Fritz G. RAGE regulation and signaling in inflammation and beyond. Journal of leukocyte biology 2013;94:55-68.
69. Thomas SJ, McDougall C, Brown ID, et al. Prevalence of symptoms and signs of shoulder problems in people with diabetes mellitus. Journal of shoulder and elbow surgery 2007;16:748-51. 70. Ando A, Sugaya H, Hagiwara Y, et al. Identification of prognostic factors for the nonoperative treatment of stiff shoulder. International orthopaedics 2013;37:859-64.
71. Kanbe K, Inoue K, Inoue Y, Chen Q. Inducement of mitogen-activated protein kinases in frozen shoulders. Journal of orthopaedic science : official journal of the Japanese Orthopaedic Association 2009;14:56-61.
72. Hagiwara Y, Kanazawa K, Ando A, et al. Blood flow changes of the anterior humeral circumflex artery decrease with the scapula in internal rotation. Knee surgery, sports traumatology, arthroscopy : official journal of the ESSKA 2015;23:1467-72.
73. Silverman ES, Collins T. Pathways of Egr-1-mediated gene transcription in vascular biology. The American journal of pathology 1999;154:665-70.
74. Chang JS, Wendt T, Qu W, et al. Oxygen deprivation triggers upregulation of early growth response-1 by the receptor for advanced glycation end products. Circulation research 2008;102:905-13.
75. Zhang K, Cao J, Dong R, Du J. Early growth response protein 1 promotes restenosis by upregulating intercellular adhesion molecule-1 in vein graft. Oxidative medicine and cellular longevity 2013;2013:432409.
76. Shi L, Kishore R, McMullen MR, Nagy LE. Lipopolysaccharide stimulation of ERK1/2 increases TNF-alpha production via Egr-1. American journal of physiology Cell physiology 2002;282:C1205-11.
77. Pawlinski R, Pedersen B, Kehrle B, et al. Regulation of tissue factor and inflammatory mediators by Egr-1 in a mouse endotoxemia model. Blood 2003;101:3940-7.
78. Bhattacharyya S, Chen SJ, Wu M, et al. Smad-independent transforming growth factor-beta regulation of early growth response-1 and sustained expression in fibrosis: implications for scleroderma. The American journal of pathology 2008;173:1085-99.
79. Chavakis T, Bierhaus A, Al-Fakhri N, et al. The pattern recognition receptor (RAGE) is a counterreceptor for leukocyte integrins: a novel pathway for inflammatory cell recruitment. The Journal of experimental medicine 2003;198:1507-15.
80. Basta G. Receptor for advanced glycation endproducts and atherosclerosis: From basic mechanisms to clinical implications. Atherosclerosis 2008;196:9-21.
81. Venereau E, Ceriotti C, Bianchi ME. DAMPs from Cell Death to New Life. Frontiers in immunology 2015;6:422.
43
82. Valencia JV, Mone M, Koehne C, Rediske J, Hughes TE. Binding of receptor for advanced glycation end products (RAGE) ligands is not sufficient to induce inflammatory signals: lack of activity of endotoxin-free albumin-derived advanced glycation end products. Diabetologia 2004;47:844-52.
83. Arcuri C, Giambanco I, Bianchi R, Donato R. Annexin V, annexin VI, S100A1 and S100B in developing and adult avian skeletal muscles. Neuroscience 2002;109:371-88.
84. Tubaro C, Arcuri C, Giambanco I, Donato R. S100B protein in myoblasts modulates myogenic differentiation via NF-kappaB-dependent inhibition of MyoD expression. Journal of cellular physiology 2010;223:270-82.
85. Cao T, Zhang L, Yao LL, et al. S100B promotes injury-induced vascular remodeling through modulating smooth muscle phenotype. Biochimica et biophysica acta Molecular basis of disease 2017;1863:2772-82.
86. Bucciarelli LG, Wendt T, Qu W, et al. RAGE blockade stabilizes established atherosclerosis in diabetic apolipoprotein E-null mice. Circulation 2002;106:2827-35.
87. Wannamethee SG, Welsh P, Papacosta O, et al. Circulating soluble receptor for advanced glycation end product: Cross-sectional associations with cardiac markers and subclinical vascular disease in older men with and without diabetes. Atherosclerosis 2017;264:36-43.
88. Zaki M, Kamal S, Kholousi S, et al. Serum soluble receptor of advanced glycation end products and risk of metabolic syndrome in Egyptian obese women. EXCLI journal 2017;16:973-80.
89. Wang X, Xu T, Mungun D, et al. The Relationship between Plasma Soluble Receptor for Advanced Glycation End Products and Coronary Artery Disease. Disease markers 2019;2019:4528382.
90. Oh S, Son M, Choi J, Lee S, Byun K. sRAGE prolonged stem cell survival and suppressed RAGE-related inflammatory cell and T lymphocyte accumulations in an Alzheimer's disease model. Biochemical and biophysical research communications 2018;495:807-13.
91. Ahmad S, Khan H, Siddiqui Z, et al. AGEs, RAGEs and s-RAGE; friend or foe for cancer. Seminars in cancer biology 2018;49:44-55.
92. Shen C, Ma Y, Zeng Z, et al. RAGE-Specific Inhibitor FPS-ZM1 Attenuates AGEs-Induced Neuroinflammation and Oxidative Stress in Rat Primary Microglia. Neurochemical research 2017;42:2902-11.
93. Cheng YZ, Yang SL, Wang JY, et al. Irbesartan attenuates advanced glycation end products-mediated damage in diabetes-associated osteoporosis through the AGEs/RAGE pathway. Life sciences 2018;205:184-92.
94. Wang Z, Zhang J, Chen L, Li J, Zhang H, Guo X. Glycine Suppresses AGE/RAGE Signaling Pathway and Subsequent Oxidative Stress by Restoring Glo1 Function in the Aorta of
44
Diabetic Rats and in HUVECs. Oxidative medicine and cellular longevity 2019;2019:4628962. 95. Chen M, Curtis TM, Stitt AW. Advanced glycation end products and diabetic retinopathy. Current medicinal chemistry 2013;20:3234-40.
96. Ahmed N, Thornalley PJ. Advanced glycation endproducts: what is their relevance to diabetic complications? Diabetes, obesity & metabolism 2007;9:233-45.
97. Xiong DD, Zhang M, Li N, Gai JF, Mao L, Li M. Mediation of inflammation, obesity and fatty liver disease by advanced glycation endoproducts. European review for medical and pharmacological sciences 2017;21:5172-8.
98. Kamtchueng Simo O, Ikhlef S, Berrougui H, Khalil A. Advanced glycation end products affect cholesterol homeostasis by impairing ABCA1 expression on macrophages. Canadian journal of physiology and pharmacology 2017;95:977-84.
99. Chen XJ, Gong XH, Jie JP, et al. Receptor for advanced glycation end products reveals a mechanism regulating thyroid hormone secretion through the SIRT1/Nrf2 pathway. Journal of cellular biochemistry 2019;120:4582-98.
100. Giannakou M, Saltiki K, Mantzou E, et al. RAGE polymorphisms and oxidative stress levels in Hashimoto's thyroiditis. European journal of clinical investigation 2017;47:341-7.
45
9. Figure Legends
Figure 1. Molecular formula and AGEs-RAGE signaling pathway
(A) Molecular formula of CML, (B) Molecular formula of pentosidine, (C) Isoform of RAGE, (D) Isoform of HMGB1, (E) AGEs-RAGE signaling pathway
Figure 2. Anatomy of right shoulder
Figure 3. Assessment of ranges of motion in participants
(A) Forward flexion, (B) lateral elevation, (C) external rotation, (D) internal rotation, (E) 90 ° abduction with external rotation, (F)90 ° abduction with internal rotation, and (G) horizontal flexion. All ranges of motion in patients from the FS group were significantly smaller than those from the RCT group.
Figure 4. Surgical findings of shoulder
(A) arthroscopic image in RCT patient (normal control), (B) arthroscopic image of RI in FS patient, (C) arthroscopic image of CHL in FS patient, (D) arthroscopic image of IGHL in FS patient. The joint capsules changed redness and stiffness.
46
Figure 5. Hematoxylin and eosin staining of the CHLs, IGHLs, and RIs
(A) Hematoxylin and eosin (HE) staining of the CHL from the FS group (×100), (B) (×400), (C) the CHL from the RCT group (×100), (D) (×400), (E) the IGHL from the FS group (×100), (F) (×400), (G) the IGHL from the RCT group (×100), (H) (×400), (I) the RI from the FS group (×100), (J) (×400), (K) the RI from the RCT group (×100), (L) (×400). The collagen bundles (asterisk) were considerably denser and contained proliferation of spindle-shaped cells with sharp or ellipsoidal nuclei, resembling fibroblast-like cells (arrows), in the CHLs and the IGHLs from the FS group compared to those from the RCT group. However, the number of collagen bundles (asterisk) increased in the RIs from the FS group than those in the RCT group.
Figure 6. Number of fibroblast-like cells of the CHLs, IGHLs, and RIs
The number of fibroblast-like cells was significantly higher in the CHLs and the IGHLs from the FS group compared to those from the RCT group. However, no statistical differences were observed in the number of fibroblast-like cells in the RIs from the FS group compared to those in the RCT group.