Expression of a Novel Stress-inducible Protein, Sestrin 2, in Rat 1
Glomerular Parietal Epithelial Cells 2
3
Hiroko Hamatani1, Keiju Hiromura1, Toru Sakairi1, Satoshi Takahashi1, 4
Mitsuharu Watanabe1, Akito Maeshima1, Takamoto Ohse2, Jeffery W. Pippin3, 5
Stuart J. Shankland3, and Yoshihisa Nojima1 6
7
1Department of Medicine and Clinical Science, Gunma University Graduate School of Medicine, 8
Maebashi, Japan. 2Division of Nephrology and Endocrinology, University of Tokyo School of 9
Medicine, Tokyo, Japan. 3Division of Nephrology, University of Washington, Seattle, Washington 10
11 12 13
Running head: Sestrin 2 in Glomerular Parietal Epithelial Cells 14
15 16
Corresponding author: Keiju Hiromura, Department of Medicine and Clinical Science, Gunma 17
University Graduate School of Medicine, 3-39-22 Showa, Maebashi, Gunma 371-8511, Japan.
18
TEL; +81-27-220-8166, FAX; +81-27-220-8173, E-Mail; [email protected] 19
20 21 22
ABSTRACT 23
Sestrin 2, initially identified as a p53 target protein, accumulates in cells exposed to stress and 24
inhibits mammalian target of rapamycin (mTOR) signaling. In normal rat kidneys, sestrin 2 was 25
selectively expressed in the PECs, identified by the marker PGP9.5. In adriamycin nephropathy, 26
sestrin 2 expression decreased in PECs on day 14, together with increased expression of 27
phosphorylated S6 ribosomal protein (P-S6RP), a downstream target of mTOR. Sestrin 2 28
expression was markedly decreased on day 42, coinciding with glomerulosclerosis and severe 29
periglomerular fibrosis. In puromycin aminonucleoside nephropathy, decreased sestrin 2 30
expression, increased P-S6RP expression, and periglomerular fibrosis were observed on day 9, 31
when massive proteinuria developed. These changes were transient and nearly normalized by 32
day 28. In crescentic glomerulonephritis, sestrin 2 expression was not detected in cellular 33
crescents, whereas P-S6RP increased. In conditionally immortalized cultured PECs, the forced 34
downregulation of sestrin 2 by shRNA resulted in increased expression of P-S6RP, and 35
increased apoptosis. These data suggest that sestrin 2 is involved in PEC homeostasis by 36
regulating the activity of mTOR. In addition, sestrin 2 could be a novel marker of PECs, and 37
decreased expression of sestrin 2 might be a marker of PEC injury.
38
39
KEY WORDS: sestrin-2, glomerular parietal epithelial cells, mTOR 40
41
Introduction 42
Sestrin 2 is a stress-inducible protein, initially identified as a hypoxia-responsive gene 43
product (7). Sestrin 2 accumulates within cells exposed to stress, and plays an important role in 44
suppressing the production of reactive oxygen species (ROS) and protecting cells from oxidative 45
damage (6). In addition, sestrin 2 inhibits mammalian target of rapamycin (mTOR) signaling 46
through a redox-independent mechanism, by activating 5'-adenosine monophosphate-activated 47
protein kinase (AMPK) and phosphorylating tuberous sclerosis protein 2 (TSC2) (5). Although 48
sestrin 2-knockout mice are reported to be fully viable and to not display any gross 49
developmental abnormalities (28), a recent study showed that deletion of sestrin 2 exacerbates 50
obesity-induced mTOR activation, glucose intolerance, insulin resistance, and hepatosteatosis 51
(18).
52
Target of rapamycin (TOR) is a Ser/Thr kinase that was originally identified in yeast mutants 53
resistant to the effect of rapamycin (29). Subsequently, mammalian TOR (mTOR) was cloned in 54
mammalian cells (29). mTOR constitutes a part of two distinct multiprotein complexes, TOR 55
complex 1 (TORC1), which is sensitive to rapamycin, and TORC2, which is not sensitive to 56
rapamycin (4, 29). Activated TORC1 directly phosphorylates two proteins, p70 ribosomal protein 57
S6 kinase (p70S6K) and 4E-binding protein 1 (4E-BP1), which stimulate ribosome biogenesis 58
and translation to increase the cell mass (13). In turn, phosphorylated p70S6K phosphorylates 59
S6 ribosomal protein (S6RP), which also stimulates translation (29).
60
Persistent mTOR activation is associated with diverse diseases including cancer, allograft 61
rejection, autoimmune disorders, cardiovascular diseases and metabolic disorders (29). Studies 62
in animals are underway to identify the roles of mTOR signaling in the pathogenesis of kidney 63
diseases such as glomerular diseases, polycystic kidney, and renal cancer (13). Genetic deletion 64
of TORC1 in mouse podocytes induces proteinuria and progressive glomerulosclerosis, whereas 65
genetic reduction of podocyte-specific TORC1 in diabetic animals suppresses the development 66
of diabetic nephropathy (10, 14).
67
Among resident cells in the glomeruli, glomerular parietal epithelial cells (PECs) remain 68
poorly understood (22). Recent studies have shown that PECs are dynamic and constantly 69
responsive to cues within the glomeruli (22). In juvenile mice, PECs have been shown to migrate 70
to become podocytes (1). PECs also contribute to the development of the sclerotic lesions in 71
focal segmental glomerulosclerosis. (26). In addition, PECs are also reported to function as a 72
second barrier of the glomerular filtrate, with their tight junctions to prevent filtered protein from 73
escaping into the extraglomerular space (21).
74
In this study, we first demonstrated that sestrin 2 was predominantly expressed in normal rat 75
PECs. We further evaluated the expression of sestrin 2 and mTOR signaling in normal and 76
diseased kidneys and attempted to determine the role of sestrin 2 and the association between 77
sestrin 2 and mTOR signaling by using conditionally immortalized cultured PECs 78
79 80
Methods 81
Animals and experimental protocol 82
All rats were purchased from Charles River Japan (Kanagawa, Japan) and fed a standard 83
diet and given water ad libitum. Six-week-old male Wistar rats were used to investigate the 84
normal kidneys. For induction of adriamycin (ADR) nephropathy, 6-week-old male Wistar rats 85
were administered a single injection of 7.5 mg/kg of ADR (doxorubicin hydrochloride;
86
Sigma-Aldrich St. Louis, MO) via the tail vein; on days 0, 8, 14 and 42 after the injection, the rats 87
were euthanized by injection of pentobarbital sodium (Kyoritsu Pharmaceutical, Tokyo, Japan) 88
and kidney samples were harvested. For induction of puromycin aminonucleoside (PAN) 89
nephropathy, 6-week-old male Wistar rats were administered a single injection of 100 mg/kg of 90
PAN (Sigma-Aldrich, St. Louis, MO) via the tail vein, as described previously (27); on days 0, 9, 91
and 28 after injection, the rats were sacrificed. For induction of crescentic glomerulonephritis, 92
7-week-old male Wistar Kyoto (WKY/NCrlCrlj) rats were administered a single injection of a 93
nephritogenic monoclonal antibody at 80 μg/body (clone a84; Iwai Chemicals Company, Tokyo, 94
Japan) via the tail vein; On day 10 after the induction, kidney samples were harvested.
95
All animal experiments were carried out in accordance with the institute of Experimental 96
Animal Research of Gunma University and were handled using protocols approved by the 97
Animal Care Committee of Gunma University.
98 99
Measurement of urinary protein excretion 100
Twenty-four-hour urine was collected under normal conditions and each week after the 101
induction of nephritis using a metabolic cage. The urinary protein concentration was determined 102
using a Bio-Rad protein assay kit (Nippon Bio-Rad Laboratories, Tokyo, Japan).
103 104
Primary antibodies 105
The primary antibodies used in this study were as follows; rabbit polyclonal anti-sestrin 2 106
antibody (ProteinTech Group, Chicago, IL), mouse monoclonal anti-PGP9.5 antibody (clone 107
13C4; Gene Tex, Irvine, CA), mouse monoclonal anti-β actin antibody (Santa Cruz Biotechnology, 108
Santa Cruz, CA), rabbit monoclonal anti-phospho-S6 ribosomal protein (P-S6RP) antibody (Ser 109
235/236; Cell signaling Technology, Beverly, MA), mouse monoclonal anti–α-smooth muscle 110
actin (α-SMA) antibody (clone 1A4; Sigma-Aldrich), rabbit anti-PAX2 antibody (Invitrogen 111
Corporation, Camarillo, CA), rabbit monoclonal anti-phospho-p70 S6 kinase antibody (Thr 389;
112
Cell signaling), and rabbit monoclonal anti-phospho-4E-BP1 antibody (Thr 37/46; Cell signaling).
113
114
Immunohistochemistry 115
Kidneys were fixed in formalin and embedded in paraffin. Four-micrometer sections were 116
stained with periodic acid-Schiff (PAS). For the immunohistochemical analysis, 4-µm sections 117
were deparaffinized and rehydrated. Antigens were retrieved by microwaving at 500 W for 10 118
min in 10 mmol/l citric acid. Endogenous peroxidase activity was blocked with periodic acid 119
(Nichirei, Tokyo, Japan) for 45 sec. The sections were incubated with the primary antibodies at 120
4ºC overnight, followed by incubation with the biotinylated secondary antibody (Vector 121
Laboratories, Burlingame, CA) for 30 min at room temperature and with horseradish 122
peroxidase-avidin (Vector Laboratories) for 30 min at room temperature. Color development was 123
performed using diaminobenzidine tetrahydrochloride solution (Nichirei). Sections were 124
counterstained with methyl green or PAS.
125 126
Quantification by immunohistochemistry 127
The glomerular damage was quantified by grading the severity of the glomerulosclerosis 128
and mesangial expansion on PAS-stained sections, on a scale of 0 to 4 (0, no lesion; 1, 0 to 129
25%; 2, 25 to 50%; 3, 50 to 75%; 4, 75 to 100%) as described previously (11). Fifty glomeruli 130
from each rat were evaluated, and the average score was calculated. Sestrin 2 expression in the 131
PECs and periglomerular α-SMA expression were quantified on a scale of 0 to 4 (0, no lesion; 1, 132
0 to 25%; 2, 25 to 50%; 3, 50 to 75%; 4, 75 to 100% around the Bowman’s capsule) in each 133
glomerulus. The average score of 50 glomeruli per section was obtained. The number of 134
PAX2-positive cells attached to the Bowman’s capsule was counted in 50 glomeruli per section.
135
The percent area of P-S6RP-positive PECs per glomerulus was assessed using Photoshop CS6 136
(Adobe, San Jose, CA), as follows. First, the total number of pixels corresponding to the 137
glomerulus was counted in the captured image of each glomerulus. After removing the 138
glomerular tuft, the number of pixels showing P-S6RP positivity (brown area) within the 139
Bowman’s capsule was counted. The percent P-S6RP-positive area was calculated using the 140
formula; number of pixels showing P-S6RP positivity / total number of pixels corresponding to the 141
glomerulus x 100 (%). The average percentage of 50 glomeruli per section was calculated.
142 143
Western blot analysis 144
Western blot analysis was performed as described elsewhere, with some modifications (27).
145
In brief, protein was extracted from the cultured PECs using RIPA lysis buffer (1 × TBS, 1%
146
Nonidet P-40, 0.5% sodium deoxycholate, 0.1% SDS, 0.004% sodium azide, Santa Cruz) 147
containing PMSF, protease inhibitor cocktail and sodium orthovanadate (Santa Cruz). The 148
protein concentration was determined by the BCA protein assay (Pierce, Rockford, IL) in 149
accordance with the manufacturer’s directions. Ten μg of the protein extract was separated on 150
4-20% precast polyacrylamide gels (Nippon Bio-Rad) and transferred to a polyvinyl difluoride 151
membrane (Immobilon-P; Millipore, Bedford, MA). After blocking with 2% bovine serum albumin 152
to reduce nonspecific antibody binding, the membrane was incubated with primary antibodies 153
overnight at 4ºC, washed with Tris-buffered saline (20mM Tris-HCl, 150mM NaCl, and 0.1%
154
Tween 20), and incubated with an alkaline phosphatase-conjugated anti-rabbit IgG or 155
anti-mouse IgG antibody (Promega, Madison, WI) at room temperature for 2 hours. After further 156
washing, detection of the bound antibody was performed using chromogen 157
5-bromo-4-chloro-3-indolyl phosphate/nitroblue tetrazolium (Sigma-Aldrich).
158 159
Cell culture 160
Conditionally immortalized mouse PECs were generated as previously described (23). Cells 161
were cultured on a 100-mm type-I collagen-coated culture dish (Iwaki, Tokyo, Japan) in RPMI 162
1640 containing penicillin, streptomycin, amphotericin B, and 2% fetal bovine serum. Cells were 163
propagated at 33ºC with 5 U/ml recombinant mouse interferon-γ (Millipore, Billerica, MA) and 164
differentiated at 37ºC in the absence of interferon-γ for 10 days.
165 166
Sestrin 2 knockdown with small hairpin RNA (shRNA) 167
Sestrin 2 silencing was performed using shRNA by a previously described method (24). In 168
brief, pLKO.1-puro lentiviral plasmids encoding shRNA for mouse sestrin 2 or nontarget control 169
(NTC), together with a puromycin resistance gene, were purchased from Thermo Scientific Open 170
Biosystems (Huntsville, AL). HIV-1-based lentiviral particles were generated in HEK293FT cells 171
(American Type Culture Collection, Manassas, VA) by co-transfection of shRNA or NTC shRNA 172
with pCMV R8.91 and pCMV-VSV-G using FuGENE6 reagent (Roche Diagnostics, Indianapolis, 173
IN). Cultured PECs were infected with the lentiviral particles in the presence of 8 μg/ml polybrene 174
for 5h at 33ºC. The effective viral titer was confirmed by observation of the cell survival in the 175
presence of 3 μg/ml puromycin (Invivogen, San Diego, CA). Five shRNA clones were tested for 176
their ability to silence sestrin 2 mRNA, and the best-performing clone was selected.
177 178
Measurements of apoptosis 179
Apoptosis was measured by Hoechst 33342 (Dojindo Molecular Technologies, Tokyo, 180
Japan) staining and by careful morphological analysis, as previously reported (12). The apoptotic 181
cells were counted in six randomly selected fields, and expressed as a percentage in triplicate. In 182
addition, the degree of apoptosis was also measured by measurement of the caspase-3 activity 183
using the APOPCYTO caspase colorimetric assay kit (Medical & Biological Laboratories, Nagoya, 184
Japan) in accordance with the manufacturer’s instructions.
185
186
Statistical analyses 187
Data are expressed as mean ± standard error of the mean (SEM). Differences between two 188
groups were compared using the two-tailed t test. Comparisons of multiple groups were 189
performed by ANOVA and if the ANOVA revealed significance, Tukey’s test was applied, using 190
IBM SPSS statistics 21 (IBM SPSS, Tokyo, Japan). P values < 0.05 were considered as 191
indicative of statistically significance.
192 193 194
Results 195
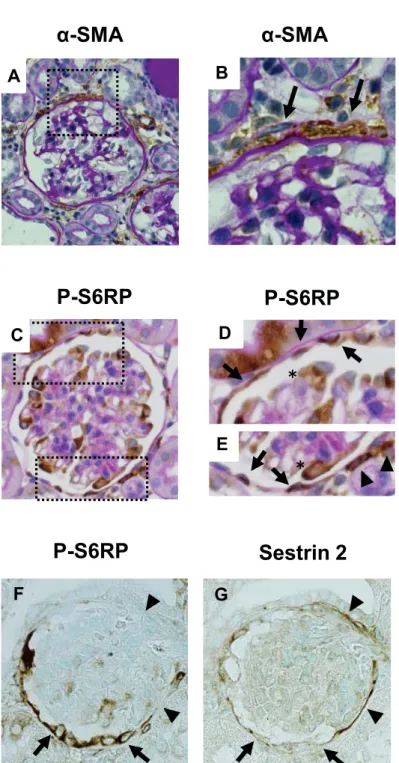
Expression of sestrin 2 in the normal rat kidney 196
Immunohistochemical staining of the normal rat kidney revealed selective expression of 197
sestrin 2 along the Bowman’s capsule (Fig.1, A and B). The staining pattern of sestrin 2 was 198
similar to that of PGP9.5 (Fig.1C), a well-known marker of PECs (25), indicating that sestrin 2 199
was predominantly expressed in the PECs. Staining was absent elsewhere in the kidney, and not 200
detected when the primary antibody was removed as a negative control (data not shown).
201 202
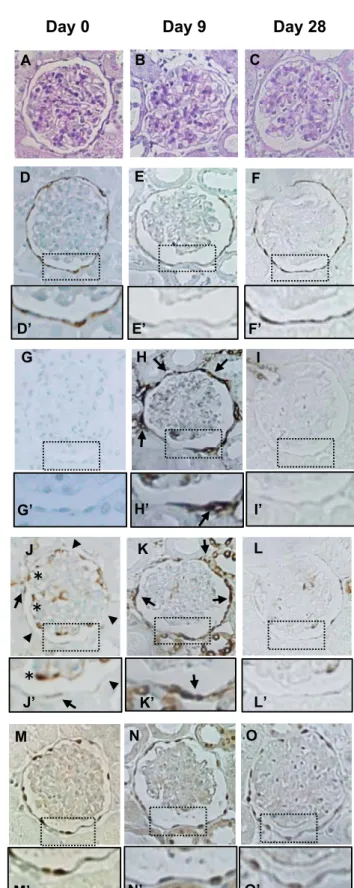
Sestrin 2 staining decreases in ADR nephropathy 203
To determine the sestrin 2 expression in PECs in proteinuric disease conditions, the ADR 204
model of focal segmental glomerulosclerosis was induced in rats. As shown in Figure 2A, urinary 205
protein excretion increased progressively after ADR injection. Severe glomerulosclerosis was 206
observed in PAS-stained sections of the kidney on day 42 (Fig. 2B and Fig. 3A-D). Quantitation 207
shows that sestrin 2 staining in PECs was unchanged until day 8, but decreased on day 14, with 208
markedly decreased staining on day 42 (Fig. 2C and Fig. 3E-H). In contrast, α-SMA staining, 209
which was observed only in the afferent or efferent arterioles on day 8 after ADR injection, was 210
detected along Bowman’s capsule on day 14, with marked increase of its expression on day 42 211
(Fig. 2D and Fig. 3I-L). However, the α-SMA-positive cells were not PECs, because they were 212
located outside the Bowman’s capsule, as shown in Figure 4A and 4B.
213 214
S6RP staining increases in ADR nephropathy 215
Because sestrin 2 is considered a negative regulator of the mTOR pathway (5), the 216
phosphorylation levels of S6RP, a direct phosphorylation target of mTOR, was examined using 217
anti-phospho S6RP antibody. P-S6RP staining in PECs was barely detected in control rats. In 218
contrast, P-S6RP staining increased significantly by day 14 (Fig. 2E, Fig. 3M-P). As shown in 219
Figure 4C and 4D, the P-S6RP-positive cells were found along Bowman’s capsule, in contrast to 220
the α-SMA-positive cells (Fig. 4A and 4B). In addition, examination of serial sections of the 221
kidney in ADR nephropathy revealed a reciprocal relationship between the expression of sestrin 222
2 and that of P-S6RP; i.e., PECs which showed increased P-S6RP expression showed 223
decreased sestrin 2 expression, whereas PECs with sustained sestrin 2 expression did not show 224
P-S6RP expression (Fig. 4F and 4G). The number of PECs in ADR nephropathy was examined 225
by PAX2 staining, because PAX2 was expressed in the nucleus of PECs (2), allowing to count 226
the number of PECs. The number of PAX2-positive cells decreased slightly with the progression 227
of ADR nephropathy (Fig. 2F and Fig.3 Q-T).
228 229
Sestrin 2 expression in PAN nephropathy 230
The expression of sestrin 2 was examined in PAN nephropathy, a model of minimal-change 231
nephrotic syndrome. As shown in Figure 5A, urinary protein excretion was markedly increased 232
on day 9 of PAN nephropathy, but was almost normal by day 28. Although glomerulosclerosis in 233
PAN nephropathy, this was much less severe as compared to ADR nephropathy (Fig. 5B and Fig.
234
6A-C). Sestrin 2 staining was moderately decreased on day 9 but increased again on day 28 (Fig.
235
5C and Fig. 6D-F). In contrast to sestrin 2 expression, the number of α-SMA-positive cells 236
outside the Bowman’s capsules was increased on day 9, but decreased again by day 28 (Fig. 5D 237
and Fig. 6G-I). The reciprocal relationship between sestrin 2 and P-S6RP was also observed in 238
PAN nephropathy (Fig. 5C, E and Fig. 6J-L), where P-S6RP increased on day 9, when sestrin 2 239
was reduced. The number of PAX2-positive cells was slightly increased on day 28 (Fig. 5F and 240
6M-Q).
241 242
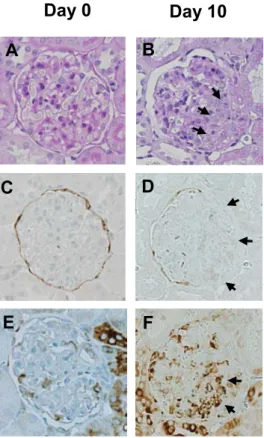
Sestrin 2 expression in crescentic glomerulonephritis 243
PECs proliferate to form crescents in crescentic glomerulonephritis (22). To determine the 244
sestrin 2 expression in crescents, WKY rats were injected with anti-GBM antibody. Massive 245
proteinuria was observed at day 10 (4.0 ± 3.8 vs. 229.3 ± 27.5 mg/day, day 0 vs. day 10, P <
246
0.01). As shown in Figure 7, sestrin 2 staining was not detected in the cellular crescents, but was 247
detected in cells lining Bowman’s capsule not participating in the crescent. In contrast, P-S6RP 248
staining increased in a subpopulation of cells within the crescents.
249 250
Downregulation of sestrin 2 by shRNA increased the activity of mTOR 251
To examine the role of sestrin 2 in PECs, conditionally immortalized cultured mouse PECs 252
were silenced for sestrin 2 using specific shRNA. Reduced expression of sestrin 2 protein was 253
confirmed by western blot analysis compared to PECs transfected with NTC (Fig, 8A and 8B).
254
Sestrin 2-silinced PECs cultured under growth-restrictive conditions for 10 days had increased 255
levels of phosphorylated 4E-BP1, p70S6K and S6RP, which are direct downstream targets of 256
mTOR (Fig. 8A and 8C).
257
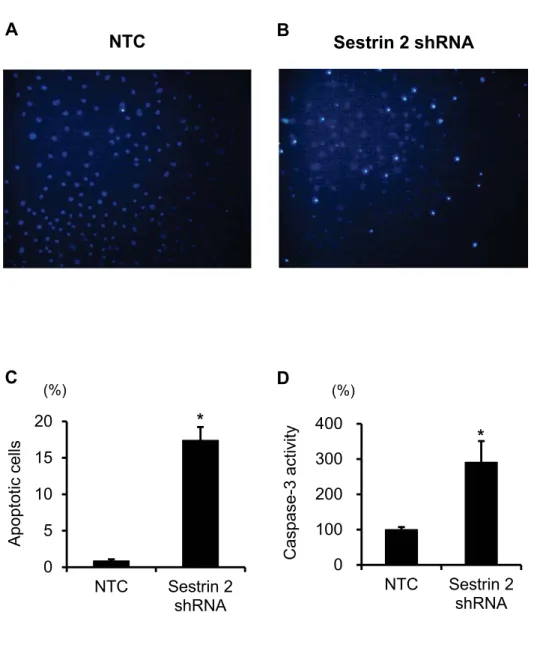
To determine the biological role of sestrin 2 in cultured PECs, apoptosis was measured. In 258
transfected PECs with reduced levels of sestrin 2- apoptosis was increased measured by 259
Hoechst 33342 staining and caspace-3 activity (Fig.9).
260 261 262
Discussion 263
Recent studies have shown that many renal cell types express genes unique to that cell, 264
which may serve not only as specific ‘markers’ for identification, but are also likely serve 265
cell-type-specific functions (22). Examples in podocytes include nephrin, podocin, NEPH1, 266
GLEPP-1, podocalyxin and synaptopodin (20). In PECs, unique proteins include PAX2, PAX8, 267
the tight junction proteins claudin-1 or occludin, ubiquitin-related protein PGP9.5, and the 268
intermediate filament protein cytokeratin (2, 21, 23, 25). The first major finding of this study was 269
that sestrin 2 was predominantly expressed in the glomerular PECs in normal rats. Sestrin 2 is a 270
novel p53 target protein that is known to accumulate in cells exposed to stress (17). Just recently, 271
increased expression of sestrin 2 was reported in renal proximal tubules in a model of renal 272
ischemia-reperfusion injury (15). In addition, upregulation of sestrin 2 expression was shown in 273
cultured renal tubular cells (NRK-52E) exposed to oxidative stress (15). Taken together, the 274
results of the current study show that in contrast to other cell types where sestrin 2 expression is 275
related to stressors, sestrin 2 is constitutively expressed in normal PECs.
276
The expression of sestrin 2 in PECs was examined in three experimental models of 277
nephrotic syndrome. The second major finding was that sestrin 2 staining in PECs was closely 278
linked with proteinuria. In both the ADR and PAN models of podocyte injury characterized by 279
proteinuria, sestrin 2-staining PECs was decreased, and this coincided with the presence of 280
proteinuria. This was further highlighted by transient proteinuria in PAN being accompanied by a 281
transient decline in sestrin 2 staining. Finally, in the anti-GBM model, reduced sestrin staining 282
accompanied the proteinuria, but the decrease was selectively in the proliferating cells of the 283
glomerular crescents. Taken together, although the models used where diseases of podocytes or 284
the GBM, changes to sestrin 2 occurred in PECs, where it is normally expressed.
285
Because sestrin 2 is known to inhibit mTOR signaling by activating AMPK and 286
phosphorylation of TSC2 (5), we also examined the downstream phosphorylation targets of 287
mTOR, with an emphasis on P-S6RP. The third major finding of our study was the paradoxical 288
expression of decreased expression of sestrin 2 with increased expression of P-S6RP during the 289
periods of heavy proteinuria in both the ADR nephropathy and PAN nephropathy models. This 290
data suggests that sestrin 2 regulates the activity of mTOR in the PECs in both health and 291
disease conditions.
292
Interestingly, increased periglomerular fibrosis, which was demonstrated by the 293
accumulation of α-SMA-positive cells around the Bowman’s capsule was observed when the 294
PECs showed decreased sestrin 2 and increased P-S6RP expression. In addition, 295
periglomerular fibrosis was reversible in the PAN nephropathy model when the proteinuria 296
resolved and expression of sestrin 2 was restored in the PECs. PECs have tight junctions and 297
are considered to serve as a permeability barrier (21). Therefore, we hypothesize that heavy 298
proteinuria induces PEC injury, which in turn results in the disruption of their tight junctions that 299
allows leakage of protein into the extraglomerular space, followed by the development of 300
periglomerular fibrosis.
301
In contrast to the reversible changes of sestrin 2 expression and periglomerular fibrosis in 302
the PAN nephropathy model, which is a model of transient nephrotic syndrome, progressive 303
decrease of sestrin 2 expression was observed in the ADR nephropathy model, which is a model 304
of progressive nephrotic syndrome. In the ADR nephropathy model, the severe decrease of 305
sestrin 2 expression in the PECs was also associated with a decrease in the number of PECs, 306
severe periglomerular fibrosis and increased glomerular sclerosis, suggesting that prolonged 307
PEC injury was closely linked to the glomerular and periglomerular injury. Although we could not 308
detect any apoptotic cells within the Bowman’s space by TUNEL staining (data not shown) on 309
day 42 of ADR nephropathy, we suppose that the reduction in the number of PEC was due to 310
apoptosis or detachment of the PECs under the condition of reduced sestrin 2 expression.
311
Because apoptotic cells are likely detached and washed away in the urinary ultrafiltrate, there 312
may only be a small window for the detection of apoptotic PECs in vivo (8). Previous in vitro 313
studies have shown that silencing of sestrin 2 increased apoptosis, including of the renal tubular 314
cells (3, 6, 15, 19). The fourth major finding of our study was that silencing of sestrin 2 in cultured 315
PECs increased phosphorylation of the downstream targets of TORC1 (4E-BP1, p70S6K and 316
S6RP) and induced apoptosis. Because immature PECs cultured under growth-permissive 317
conditions stop proliferating and differentiate under growth-restrictive conditions, we supposed 318
that the dysregulation of mTOR activity was associated with the apoptosis of PECs in vitro.
319
However, the precise mechanisms underlying the role of sestrin 2 in PEC survival need to be 320
determined in the future.
321
The above data show that decreased expression of sestrin 2 was associated with a 322
decrease in the number of PECs in the ADR nephropathy model and cultured PECs. Meanwhile, 323
we observed that, in the crescentic glomerulonephritis model, in which PECs are considered to 324
proliferate to form crescents, sestrin 2 expression was decreased in the cells within the crescent, 325
whereas P-S6RP expression was highly increased in these cells. Considering the role of mTOR 326
in cell growth and proliferation (9, 29), the increased activity of mTOR in these cells seems 327
explicable. Recently, Kurayama et al. reported increased expression of P-S6RP in the PECs 328
before apparent crescent formation and further increase in the expression in the crescentic 329
lesions in rat anti-GBM nephritis (16). In addition, they studied the effects of the mTOR inhibitor, 330
everolimus, in this crescent model. Interestingly, early treatment with the mTOR inhibitor led to 331
increased cellular necrosis of the PECs, whereas later treatment reduced glomerular crescent 332
formation. These findings demonstrate the complex role of the mTOR pathway in the PECs.
333
Taken together, Figure 10 shows our working hypothesis about sestrin 2 expression and 334
mTOR activity in the PECs in normal and diseased kidneys based on the present data. In the 335
normal rat kidney, sestrin 2 is predominantly expressed in the PECs and minimal mTOR activity 336
is detected. These conditions may be required for maintenance of the homeostasis of PECs. In 337
injured PECs, decreased expression of sestrin 2 and increased activity of mTOR are observed.
338
In the PAN nephropathy or ADR nephropathy model, injured PECs lose their barrier function and 339
the leakage of protein around the Bowman’s space causes periglomerular fibrosis. In addition, 340
sustained injury of the PECs in the ADR nephropathy model leads to apoptosis and/or 341
detachment of PECs, which in turn causes glomerulosclerosis. In contrast, in anti-GBM nephritis, 342
decreased expression of sestrin 2 and increased activity of mTOR are associated with 343
proliferation of the PECs to form crescents. Although the reason why decreased sestrin 2 344
expression and increased mTOR activity are associated with different outcomes in the PECs still 345
remains unclear after this study, our data show that sestrin 2 could be a novel marker of PECs 346
and that decreased expression of sestrin 2 might be a marker of PEC injury.
347 348
GRANTS 349
This research was funded by grants from the Ministry of Education, Culture, Sports, Science 350
and Technology of Japan.
351 352
DISCLOSURES 353
None of the authors have any competing interests to declare.
354 355
REFERENCES 356
1. Appel D, Kershaw DB, Smeets B, Yuan G, Fuss A, Frye B, Elger M, Kriz W, Floege J, 357
Moeller MJ. Recruitment of podocytes from glomerular parietal epithelial cells. J Am Soc 358
Nephrol 20: 333-343, 2009.
359
2. Bariety J, Mandet C, Hill GS, Bruneval P. Parietal podocytes in normal human glomeruli.
360
J Am Soc Nephrol 17: 2770-2780, 2006.
361
3. Ben-Sahra I, Dirat B, Laurent K, Puissant A, Auberger P, Budanov A, Tanti JF, Bost F.
362
Sestrin2 integrates Akt and mTOR signaling to protect cells against energetic 363
stress-induced death. Cell Death Differ 20: 611-619, 2013.
364
4. Budanov AV. Stress-responsive sestrins link p53 with redox regulation and mammalian 365
target of rapamycin signaling. Antioxid Redox Signal 15: 1679-1690, 2011.
366
5. Budanov AV, Karin M. p53 target genes sestrin1 and sestrin2 connect genotoxic stress 367
and mTOR signaling. Cell 134: 451-460, 2008.
368
6. Budanov AV, Sablina AA, Feinstein E, Koonin EV, Chumakov PM. Regeneration of 369
peroxiredoxins by p53-regulated sestrins, homologs of bacterial AhpD. Science 304:
370
596-600, 2004.
371
7. Budanov AV, Shoshani T, Faerman A, Zelin E, Kamer I, Kalinski H, Gorodin S, 372
Fishman A, Chajut A, Einat P, Skaliter R, Gudkov AV, Chumakov PM, Feinstein E.
373
Identification of a novel stress-responsive gene Hi95 involved in regulation of cell viability.
374
Oncogene 21: 6017-6031, 2002.
375
8. Chang AM, Ohse T, Krofft RD, Wu JS, Eddy AA, Pippin JW, Shankland SJ.
376
Albumin-induced apoptosis of glomerular parietal epithelial cells is modulated by 377
extracellular signal-regulated kinase 1/2. Nephrol Dial Transplant 27: 1330-1343, 2012.
378
9. Dowling RJ, Topisirovic I, Fonseca BD, Sonenberg N. Dissecting the role of mTOR:
379
lessons from mTOR inhibitors. Biochimica et biophysica acta 1804: 433-439, 2010.
380
10. G del M, Hartleben B, Herbach N, Liu S, Zschiedrich S, Lu S, Debreczeni-Mór A, 381
Lindenmeyer MT, Rastaldi MP, Hartleben G, Wiech T, Fornoni A, Nelson RG, Kretzler 382
M, Wanke R, Pavenstädt H, Kerjaschki D, Cohen CD, Hall MN, Rüegg MA, Inoki K, 383
Walz G, Huber TB. Role of mTOR in podocyte function and diabetic nephropathy in 384
humans and mice. J Clin Invest 121: 2197-2209, 2011.
385
11. Hiramatsu N, Hiromura K, Shigehara T, Kuroiwa T, Ideura H, Sakurai N, Takeuchi S, 386
Tomioka M, Ikeuchi H, Kaneko Y, Ueki K, Kopp JB, Nojima Y. Angiotensin II type 1 387
receptor blockade inhibits the development and progression of HIV-associated nephropathy 388
in a mouse model. J Am Soc Nephrol 18: 515-527, 2007.
389
12. Hiromura K, Pippin JW, Blonski MJ, Roberts JM, Shankland SJ. The subcellular 390
localization of cyclin dependent kinase 2 determines the fate of mesangial cells: role in 391
apoptosis and proliferation. Oncogene 21: 1750-1758, 2002.
392
13. Huber TB, Walz G, Kuehn EW. mTOR and rapamycin in the kidney: signaling and 393
therapeutic implications beyond immunosuppression. Kidney Int 79: 502-511, 2011.
394
14. Inoki K, Mori H, Wang J, Suzuki T, Hong S, Yoshida S, Blattner SM, Ikenoue T, Ruegg 395
MA, Hall MN, Kwiatkowski DJ, Rastaldi MP, Huber TB, Kretzler M, Holzman LB, 396
Wiggins RC, Guan KL. mTORC1 activation in podocytes is a critical step in the 397
development of diabetic nephropathy in mice. J Clin Invest 121: 2181-2196, 2011.
398
15. Ishihara M, Urushido M, Hamada K, Matsumoto T, Shimamura Y, Ogata K, Inoue K, 399
Taniguchi Y, Horino T, Fujieda M, Fujimoto S, Terada Y. Sestrin-2 and BNIP3 regulate 400
autophagy and mitophagy in renal tubular cells in acute kidney injury. Am J Physiol Renal 401
Physiol 305: F495-509, 2013.
402
16. Kurayama R, Ito N, Nishibori Y, Fukuhara D, Akimoto Y, Higashihara E, Ishigaki Y, Sai 403
Y, Miyamoto K, Endou H, Kanai Y, Yan K. Role of amino acid transporter LAT2 in the 404
activation of mTORC1 pathway and the pathogenesis of crescentic glomerulonephritis. Lab 405
Invest 91: 992-1006, 2011.
406
17. Lee JH, Budanov AV, Park EJ, Birse R, Kim TE, Perkins GA, Ocorr K, Ellisman MH, 407
Bodmer R, Bier E, Karin M. Sestrin as a feedback inhibitor of TOR that prevents 408
age-related pathologies. Science 327: 1223-1228, 2010.
409
18. Lee JH, Budanov AV, Talukdar S, Park EJ, Park HL, Park HW, Bandyopadhyay G, Li N, 410
Aghajan M, Jang I, Wolfe AM, Perkins GA, Ellisman MH, Bier E, Scadeng M, Foretz M, 411
Viollet B, Olefsky J, Karin M. Maintenance of metabolic homeostasis by Sestrin2 and 412
Sestrin3. Cell Metab 16: 311-321, 2012.
413
19. Liu SY, Lee YJ, Lee TC. Association of platelet-derived growth factor receptor beta 414
accumulation with increased oxidative stress and cellular injury in sestrin 2 silenced human 415
glioblastoma cells. FEBS Lett 585: 1853-1858, 2011.
416
20. Mundel P, Shankland SJ. Podocyte biology and response to injury. J Am Soc Nephrol 13:
417
3005-3015, 2002.
418
21. Ohse T, Chang AM, Pippin JW, Jarad G, Hudkins KL, Alpers CE, Miner JH, Shankland 419
SJ. A new function for parietal epithelial cells: a second glomerular barrier. Am J Physiol 420
Renal Physiol 297: F1566-1574, 2009.
421
22. Ohse T, Pippin JW, Chang AM, Krofft RD, Miner JH, Vaughan MR, Shankland SJ. The 422
enigmatic parietal epithelial cell is finally getting noticed: a review. Kidney Int 76: 1225-1238, 423
2009.
424
23. Ohse T, Pippin JW, Vaughan MR, Brinkkoetter PT, Krofft RD, Shankland SJ.
425
Establishment of conditionally immortalized mouse glomerular parietal epithelial cells in 426
culture. J Am Soc Nephrol 19: 1879-1890, 2008.
427
24. Sakairi T, Abe Y, Kopp JB. TGF-beta1 reduces Wilms' tumor suppressor gene expression 428
in podocytes. Nephrol Dial Transplant 26: 2746-2752, 2011.
429
25. Shirato I, Asanuma K, Takeda Y, Hayashi K, Tomino Y. Protein gene product 9.5 is 430
selectively localized in parietal epithelial cells of Bowman's capsule in the rat kidney. J Am 431
Soc Nephrol 11: 2381-2386, 2000.
432
26. Smeets B, Kuppe C, Sicking EM, Fuss A, Jirak P, van Kuppevelt TH, Endlich K, 433
Wetzels JF, Grone HJ, Floege J, Moeller MJ. Parietal epithelial cells participate in the 434
formation of sclerotic lesions in focal segmental glomerulosclerosis. J Am Soc Nephrol 22:
435
1262-1274, 2011.
436
27. Takeuchi S, Hiromura K, Tomioka M, Takahashi S, Sakairi T, Maeshima A, Kaneko Y, 437
Kuroiwa T, Nojima Y. The immunosuppressive drug mizoribine directly prevents podocyte 438
injury in puromycin aminonucleoside nephrosis. Nephron Exp Nephrol 116: e3-10, 2010.
439
28. Wempe F, De-Zolt S, Koli K, Bangsow T, Parajuli N, Dumitrascu R, Sterner-Kock A, 440
Weissmann N, Keski-Oja J, von Melchner H. Inactivation of sestrin 2 induces TGF-beta 441
signaling and partially rescues pulmonary emphysema in a mouse model of COPD. Dis 442
Model Mech 3: 246-253, 2010.
443
29. Wullschleger S, Loewith R, Hall MN. TOR signaling in growth and metabolism. Cell 124:
444
471-484, 2006.
445
446 447 448
FIGURE LEGENDS 449
Fig. 1. Immunohistochemical staining of sestrin 2 in the normal rat kidney. A and B:
450
Sestrin 2 was selectively expressed in the PECs. C: PGP9.5, a known marker of PECs was also 451
principally expressed in the PECs. Original magnification: A, x100; B and C, x400.
452 453
Fig.2. Changes of the urinary protein excretion, glomerulosclerosis and expression of 454
each of the proteins in the PECs in the rat model of ADR nephropathy.
455
Rats were injected with ADR and the urinary protein excretion and kidney sections were 456
examined on days 0, 8, 14 and 42 (n = 6, at each time-point). A: Urinary protein excretion (UP).
457
B: Glomerulosclerosis (GS). C: Sestrin 2. D: α-SMA. E: P-S6RP. F: PAX2. The degree of 458
glomerulosclerosis (B) was estimated on the PAS-stained kidney sections as described in the 459
methods section. The degrees of sestrin 2 (C), α-SMA (D), P-S6RP (E), PAX2 (F) expression 460
were estimated on the immunohistochemically stained kidney sections using each specific 461
antibody as described in the methods section. * p < 0.01 vs. day 0. # p < 0.01 vs. day 8. ## p <
462
0.05 vs. day 8. § p < 0.01 vs. day 14.
463 464
Fig.3. Immunohistochemical staining for each protein in the PECs during ADR 465
nephropathy.
466
Rats were injected with ADR and the kidney sections were examined on day 0 (A, E, I, M, Q), 467
day 8 (B, F, J, N, R), day 14 (C, G, K, O, S), and day 42 (D, H, L, P, T). A-D: PAS staining.
468
Severe glomerulosclerosis was observed on day 42 (D). E-H: Sestrin 2. Sestrin 2 expression 469
decreased on day 14 (G and G’) with an even more marked decrease observed on day 42 (H 470
and H’). I-L: α-SMA. α-SMA expression was not detected on day 0 (I and I’) or day 8 (J and J’), 471
except in the small arteries (J: arrow). Expression of α-SMA was detected around the basement 472
membrane of the Bowman’s capsule on day 14 (K and K’: arrows) and markedly increased on 473
day 42 (P and P’: arrows). M-P: P-S6RP. Along the Bowman’s capsule, weak P-S6RP 474
expression was detected in some cells (M: arrows), but most cells were negative for P-S6RP 475
expression on day 0 (M and M’: arrow heads). Within the glomerular tufts, some cells showed 476
strong staining for P-S6RP (M and M’: asterisks). These cells were identified as podocytes 477
based on their localization. Strong expression of P-S6RP was observed in the cells along the 478
Bowman’s capsule on day 14 (O and O’: arrows) and day 42 (P and P’). Q-T: PAX2. PAX2 was 479
detected in the cells along the Bowman’s capsule, in a nuclear localization. The number of 480
PAX2-positive cells was mildly decreased on day 42 (T and T’). Original magnification: x400.
481 482
Fig.4. Immunohistochemical stainings for α-SMA, P-S6RP and sestrin 2 in a rat model of 483
ADR nephropathy.
484
A and B: α-SMA immunostaining counterstained with PAS staining on day 42 in the model of 485
ADR nephropathy. α-SMA-positive cells localized outside the Bowman’s capsule (B: arrow). C-E:
486
P-S6RP immunostaining counterstained with PAS staining on day 14 of ADR nephropathy.
487
P-S6RP-positive cells mainly localized within the Bowman’s capsule (D and E: arrow). A few 488
P-R6RP-positive cells were also detected outside the Bowman’s capsule (E: arrowheads). F:
489
P-S6RP. G: Sestrin 2. Serial sections of P-S6RP (F) and sestrin 2 (G) on day 14 of ADR 490
nephropathy showed a reciprocal relationship between sestrin 2 and P-S6RP expressions. PECs 491
which showed strong P-S6RP expression (F: arrows) showed faint sestrin 2 expression (G:
492
arrows). In contrast, PECs which showed no P-S6RP expression (F: arrowheads) showed 493
sustained sestrin 2 expression (G: arrowheads). Original magnification: x400.
494 495
Fig.5. Changes of the urinary protein excretion, glomerulosclerosis and expression of 496
each of the proteins in the PECs of the rat model of PAN nephropathy.
497
Rats were injected with PAN and the urinary protein excretion and kidney sections were 498
examined on days 0, 9 and 28 (n = 6, at each time-point). A: Urinary protein excretion (UP). B:
499
Glomerulosclerosis (GS) C: Sestrin 2. D: α-smooth muscle actin (α-SMA). E: P-S6RP. F: PAX2.
500
The degree of glomerulosclerosis (B) was estimated on the PAS-stained kidney sections as 501
described in the methods section. The degrees of sestrin 2 (C), α-SMA (D), P-S6RP (E), PAX2 502
(F) expressions were estimated on the immunohistochemically stained kidney sections using 503
each specific antibody as described in the methods section. * p < 0.01 vs. day 0. ** p < 0.05 vs.
504
day 0. # p < 0.01 vs. day 9.
505 506
Fig.6. Immunohistochemical staining for each protein in the PECs in a rat model of PAN 507
nephropathy.
508
Rats were injected with PAN and the kidney sections were examined on day 0 (A, D, G, J, M), 509
day 9 (B,E, H, K, N), and day 28 (C, F, I, L, O). A-C: PAS staining. D-F: Sestrin-2. Sestrin 2 510
expression decreased on day 9 (E and E’), but was restored on day 28 (F and F’). G-I: α-SMA.
511
Strong expression of α-SMA was detected around the basement membrane of the Bowman’s 512
capsule on day 9 (H and H’: arrows), but disappeared on day 28 (I and I’). J-L: P-S6RP.
513
P-S6RP was detected weakly in some cells along the Bowman’s capsule, (J and J’: arrows), but 514
most cells were negative for P-S6RP expression on day 0 (J and J’’: arrowheads). Strong 515
expression of P-S6RP was observed in the cells along the Bowman’s capsule on day 9 (K and 516
K’: arrows), but the expression almost disappeared on day 28. Within the glomerular tufts, some 517
cells showed strong staining for P-S6RP (J and J’: asterisks). M-O: PAX2. PAX2 was detected in 518
the cells along the Bowman’s capsule in a nuclear localization. Original magnification: x400.
519 520
Fig.7. Immunohistochemical stainings for sestrin-2 and P-S6RP in anti-GBM 521
antibody-induced glomerulonephritis. WKY rats were injected with anti-GBM antibody and 522
the kidney sections were examined on day 0 (A, C, E; serial sections) and day 10 (B, D, F; serial 523
sections). A and B: Periodic acid-Schiff (PAS) staining. Cellular crescents were observed on day 524
10 (B: arrows). C and D: Sestrin-2. Sestrin-2 expression was not detected in the area of the 525
crescent formation (D: arrows). E and F: P-S6RP. Strong P-S6RP expression was detected 526
within the crescents (F: arrows). Original magnification: x400.
527
528
Fig.8. Effect of sestrin 2 downregulation on the expression of mammalian target of 529
rapamycin (mTOR) in conditionally immortalized cultured PECs. Conditionally immortalized 530
cultured PECs were transfected with shRNA targeting sestrin 2 or non-target control (NTC).
531
Transfected PECs were then cultured under growth-restrictive conditions for 10 days and the 532
proteins were extracted for western-blot analysis. A: Western-blot analysis for sestrin 2 and the 533
downstream targets of mTOR. Sestrin 2 expression was reduced in the PECs transfected with 534
shRNA targeting sestrin 2 as compared with that in the PECs transfected with NTC.
535
Phosphorylation of the downstream targets of mTOR: P-4E-BP1, P-p70S6K and P-S6RP 536
expressions were also increased in the PECs transfected with shRNA targeting sestrin 2 as 537
compared with that in the PECs transfected with NTC shRNA. B: Relative expression of sestrin 2 538
protein detected by western-blot analysis. As compared with that in the PECs transfected with 539
NTC, a significant reduction of sestrin 2 protein expression was observed in the PECs 540
transfected with sestrin 2 shRNA. * p < 0.01. B: Relative expression of the proteins downstream 541
of phosphorylated mTOR. The expressions of P-4E-BP-1, P-p70S6K and P-S6RP were 542
significantly increased in the PECs transfected with sestrin 2 shRNA as compared with that in the 543
PECs transfected with NTC. ** p < 0.05.
544 545
Fig.9. Effect of sestrin 2 downregulation on cellular apoptosis in conditionally 546
immortalized cultured PECs. Conditionally immortalized cultured PECs were transfected with 547
shRNA targeting sestrin 2 or non-target control (NTC). Transfected PECs were then cultured 548
under growth-restrictive conditions for 10 days and the degree of apoptosis was determined.
549
A-C: Hoechst 33342 staining in cultured PECs. A: PECs transfected with NTC showed faint 550
apoptosis. B: Increased apoptosis was observed in the PECs transfected with sestrin 2 shRNA.
551
C: Apoptosis as determined by Hoechst 33342 staining was significantly increased in the PECs 552
transfected with sestrin 2 shRNA than in the PECs transfected with NTC. * p< 0.01. D: The 553
degree of apoptosis as determined by caspase-3 staining was also significantly increased in the 554
PECs transfected with sestrin 2 shRNA as compared with that in the cells transfected with NTC. * 555
p< 0.01.
556 557
Fig.10. Schema. Sestrin 2 and activity of mTOR in normal and diseased PECs. The details 558
are described in the text.
559 560 561
Figure 1
B
Sestrin 2 PGP9.5
C
0 200 400 600 800
d0 d8 d14 d42
0 1 2
d0 d8 d14 d42
GS (score
0 1 2 3
d0 d8 d14 d42
0 1 2 3 4
d0 d8 d14 d42
-SMA (score)UP (mg/day Sestrin 2(scor
0 0.2 0.4 0.6 0.8 1
d0 d8 d14 d42
P-S6RP (%) PAX2 (cells/gl)
Figure 2
D E F
0 5 10 15
d0 d8 d14 d42
##
*
*
*
# i*
#*
#i
*
###
*
# iPAS
Sestrin 2
Figure 3
E F G H
E’ F’ G’ H’
PAX 2 -SMA
P-S6RP
I J K L
I’ J’ K’ L’
M N O P
M’ N’ O’ P’
Q R S T
Q’ R’ S’ T’
䠆 䠆
䠆 䠆
Figure 4
Sestrin 2 P-S6RP
F G
P-S6RP P-S6RP
䠆
䠆
C D
E
0 1
d0 d9 d28
0 5 10 15
d0 d9 d28
GS (score
UP (mg/day Sestrin 2(sco
*
*
0 0.5 1
d0 d9 d28
P-S6RP (%)
* **
Figure 5
D E F
0 1 2 3
d0 d9 d28
-SMA (score)
*
0 1 2 3 4
d0 d9 d28
PAX2 (cells/gl)
#
#
#
# 0
200 400
d0 d9 d28
Figure 6
Sestrin 2
-SMA
PAX 2 P-S6RP
D E F
D’ E’ F’
G H I
G’ H’ I’
J K L
J’ K’ L’
M N O
M’ N’ O’
䠆 䠆
䠆
Figure 7
PAS
Sestrin 2
P-S6RP
C D
E F
0 2 4 6 8 10 12
P-4E-BP1 P-p70S6K P-S6RP Sestrin 2 shRNA Sestrin 2
P-p70S6K P-4E-BP1
P-S6RP -actin
Figure 8
0.0 0.2 0.4 0.6 0.8 1.0 1.2
NTC Sestrin 2 shRNA
*
Relative expression of sestrin 2protein
B
NTC
Relative expression
**
** **
C
Figure 9
*
(%)
0 5 10 15 20
NTC Sestrin 2 shRNA
0 100 200 300 400
NTC Sestrin 2 shRNA
*
Apoptotic cells Caspase-3 activity
C D
(%)Figure 10
Proliferation Loss of
barrier function
Apoptosis and/or detachment
mTOR mTOR
Maintenance of homeostasis
Periglomerular fibrosis
Glomerulo- sclerosis
Crescent formation