Behavioral mechanism of suckling and nipple attachent is important for newborn mammals. The initial suckling is explained to be controlled by locating and grasping nipple induced by olfactory and tactile stimuli(Hall, 1990).Although the role of these stimuli was confirmed in postnatal rat pups and kittens (Larson & Stein, 1984; Singh & Hofer, 1978;
Teicher & Blass, 1977),little is known about how the newborn pups orient and locate the nipple immediately after birth because of the technical difficulty to intentiona11y separate or combine the olfactory and tactile stimuli of the parturient mother and to find the effective behavioral categories in spite of their immature and limited movements.
The relationship between tactile stimulus and oral grasping has been investigated with artificial nipple and intraoral cannula in fetuses and cesarean-delivered rat pups. Periora1
stimulation with artificial nipple as a mimic of mother’s nipple in shape and touch induced mouthing, licking and grasping(Browne, Robinson, & Smotherman, 1994; Robinson, Hoeltzel, Cooke, Umphress, Murrish, & Smotheman, 1992; SmothoIman, Goffman, Petrov, & Varlinskaya, 1997).When bovine light cream as a substitute for mother’s milk was infused through intraoral cannula, it supressed perioral responsiveness(Smother-man & Robinson, 1992),and prior exposure to the artificial nipple increased mouthing in response to the milk substitute(Smotherman, Moore, & Robinson, 1993).The use of those substitutes makes it possible to examine the natural behavioral sequence of suckling following the perioral stimulation with mother’ s nipple. Then the next point is the study of locating nipple before the initial nipple grasping induced by mother’s nipple.
Role of Amniotic Fluid and Mother
,
s Fur in
Approaching and Orienting Mother in Cesarean
Delivered Rat Pups
Noriko KODAMA
Abstract
We investigated the role of amniotic fluid and mother’s ventral fur before the initial nipple grasping in 356 cesarean delivered newborn rats. The odor of amniotic fluid was more effective than that of the fur in activating head extension and up-down movements related to nipple orientation. These movements and mouthing movement were increased by the perioral stimulation with the fur. When the odor of amniodc fluid and the fur and the perioral stimulation with them were combined, more active movements including mouthing and licking were found. These results indicate that the odor of amniotic fluid familiar in prenatal period plays an important role for newborns to orient and locate the mother, and that the perioral stimulation with mother’s ventral fur wet with amniotic fluid activate the nipple probing. This stimulus-response re1ationship would be necessary for the onset of nipple grasping and nipple attachment.
In advance of perioral nipple stimulation, newborn pups receive several olfactory and tactile stimuli of mother’s ventra1 fur with which the olfactory stimuli are attached. The separation of these stimuli from each other is difficult as they are closely related and presented to pups at the same time in natural condition. It is known in the study of tactile stimulus that young rat pups on fake fur probe into the fur at high rate(Ha11, 1990), but the characteristics of its fiber would be different from that of mother’ hair. It is also questionable whether fake fur has the same stimulus effect on responses of newborn pups as mother’s fur. Another consideration would be necessary for examining the stimulus effects on their responses. From this viewpoint, we focused on the role of mother’s fur, made a fur brush from around the ventral area and compared it to fake fur and other fur brushes(Kodama & Smotherman, 1997). The mother’s fur was found to be different from the fake fur in density, the kinds of hairs, and shape(width, 1ength, and the pointed end of a hair).The fur brush from mother was more effective for activating specific head movement than fake fur and paintbrush. These results indicate that the mother’s fur is more adequate for investigating natural responses of newborn pups to mother and the process of nipple locating.
The role of mother’s fur in locating nipple is not clear except the findings that the presence of mother elicited activity connected with nipple searching in postnatal rabbits and rats (Blass & Teicher, 1980; Hall, 1990; Hofer, Shair
& Singh, 1976).The newborn rats before initial suckling attached the anesthetized mother’s nipple washed and then coated with amniotic fluid which is the most familiar stimulus to newborn pups, mother’s saliva, and washed extract from the nipple area.(Teicher & Blass, 1977).As these studies does not separate the effect of fur odor itself from the
other odor and facial stimulation by mother’s fur, it is necessary to separate them for assessing that the fur keeps its specific odor as a source to let the newborn pups approach. Then, assessing the combined effects of odor and fur stimulation is also necessary, because we found the similar effect in our study where the artificial nipple and several stimuli presented at the same time increased the newborn’s responses(Koffman, Petrov, Vallin-skaya, & Smotheman, 1998).
In the process of locating nipple, the mother’ s fur stimulates several body areas of newborn pups, mainly back and face. The back area is usua11y stimulated when the mother carries the pups to the nest and crouches over them in it. In compared to the back area, the face area(nose and perioral areas)is more important for locating nipple because it is frequently stimulated by mother’s ventra1 fur before the onset of initial suckling. Bilateral stimulation of the face area triggers rooting reflex in cesarean delivered mice(Kodama, 1993).The nose area centering the lateral vibrissal pad is the first to become fully sensitive in perinatal period(Narayanan, Fox, & H a m b u r g e r , 1971). F i n e v i b r i s s a e , necessary for vibrissa placing reaction in postnatal period(Fox, 1965),are already grown at birth in rats. When a stiff filament is applied to the lateral vibrissal pad after the perioral stimulation with artificial nipple in rat fetuses, facial wiping which is one of aversive responses increases, while the perioral stimulation with artificial nipple is effective for increasing mouthing and nipple grasping (Browne, et al., 1994).If mother’s ventral fur
itself is applied to those areas instead of artificial tactile stimuli, we could find more natural locating response and the differential effects by the stimulation of several body areas.
One of the problems in measuring pups’ responses to the facial stimulation is their
complexity. Many studies, therefore, mainly describe the kinds of head movement. Once nipple attachment is formed, pups show nosing into the hair of the mother’s ventral fur(Hofer, Shair,& Singh, 1976).They move the head from side to side, scan the surface of the mother with snout and contact with its nipple (Blass & Teicher, 1980; Blass, Shuleikina-Turpaeva, & Luschekin., 1988).The basic components of these movements are head turning and upward-downward movement which originate in prenatal period(Narayanan, et al., 1971).When the tip of the artificial nipple enters the oral cavity, the head is straightened to become aligned with the body trunk in cesarean delivered pups(Smother-man, et al., 1997).These descriptions indicate that the head movement apparently looks very complicated but is consisted of simple b a s i c m o v e m e n t s . I t a l l o w s u s t h e quantitative study assessing the relationship between the stimulated area and the specific head movement. In our preliminary study, head extension and up-down movements increased, and head turning decreased when the perioral area was stimulated by the mother’s fur(Kodama & Smotherman, 1997). The quantitative assay of head movement would be useful for the study of behavioral mechanism of orienting and locating nipple. The present study aimed to investigate the role of the odor of amniotic fluid in head movement(Experiment 1),effects of perioral stimulation with mother’s fur on head movement(Experiment 2),the combined effects of the odor of mother’s fur and the tactile stimulation with the fur(Experiment 3),and the combined effects of the odor of amniotic fluid and the fur and the tactile stimulus of fur on head movement(Experi-ment 4).Cesarean delivered rat pups were tested in these experiments to prevent them from any experience with the odor and tactile stimuli of the mother except amniotic fluid.
General Methods
SubjectsSubjects were 356 cesarean delivered rat pups produced in timed matings of 96 Sprague-Dawley female rats. Adult rats were housed in groups of three females and one male during a 4-day breeding period. Vaginal smears collected daily during this breeding period were used to date conception with the presence of sperm designated as E0. After removal of the breeding male, pregnant females were housed in plastic breeding cages ( 36× 47× 20c m ) u n d e r c o n s t a n t r o o m
temperature(22℃)and 12-hr light-dark cycle (lights on at 0700).Food and water were
freely available. Cesarean Delivery
The pregnant female on E21 was briefly anesthetized with ether and prepared by chemomyelotomy with injection of 100% ethano1(90μl)into the spinal cord between the first and second lumbar vertebrae. This procedure irreversibly blocks neural transmission within the spinal cord at the low thoracic level and eliminates sensation in the lower half of the body. After the chemomye-lotomy, the uterus of pregnant rat in a holding apparatus was exteriorized through a midline laparotomy. The uterus was cut open along the lateral extent of the uterus avoiding uterine blood vessels and points of placental attachment. The amniotic covering was removed, the unbilical cord was cut after its ligation, and the amniotic fluid was absorbed with a paper towel. Eight pups from each female were placed in a plastic cage(16×11 ×5cm)with a moist paper towel for three hours for adaptation. The cage was warmed at 32 ℃ in a incubator. Pups, which skin is pink by well-oxygenated, were selected as subjects. The females were sacrificed by
rapid cervical dislocation. The cesarean delivered pups were euthanized after the experiment. All rats were treated in accordance with guidelines for animal care by National Institutes of Health(PHS Publication 86-23).
Experiment l
Responses to the odor of amniotic fluid after cesarean deliveIy
Newborn pups probe and suckle the mother’ s nipple to which amniotic fluid is attached (Teicher & Blass, 1979).The aim of Experiment
1 was to separate the odor of amniotic fluid from that of other stimuli from mother and to examine the role of the odor of amniotic fluid in head movement related with orienting and locating nipple immediately after birth.
Method
SubjectsSubjects were 128 cesarean delivered pups on E21 produced by 32 female rats. They were assigned to one of four groups: GroupAF, SAL, DW, and NS. The odor of amniotic fluid was presented to AF pups, that of saline to SAL pups, and that of distilled water to DW pups. No stimulus was presented to NS pups.
Procedure
Three hours after delive!y, each pup was placed in a plastic container with rounded corners(bottom dimension 5×5cm)warmed at 32℃ by a heating pad. The amniotic fluid obtained fom the E21 female, saline, and distilled water were soaked into cotton(1cm in diameter)inserted in the round top of stainless wire. The cotton was presented to the pup, 1cm apart from the nose region without contact. A pup was tested for 10 minutes, 5 minutes baseline session and 5
minutes odor presentation session. Its spontaneous body movements were observed in the baseline session, and the responses to the odor in the odor presentation session. All the spontaneous body movements and responses to the odor were videotaped and then recorded in 0.1 second to permit their detailed analysis by reduced-speed playback. Behavioral Recording
A f t e r t h e t e s t , s p o n t a n e o u s b o d y movements and responses to the odor were classified into eight categories. Head turn movement, Head(Turn),is the horizontal or diagonal movement of head relative to the body axis. Head extension and up-down movements, Head(EUD),is the forward movement and/or up-and-down movement of head without its turn. Forelimb movement is the movement of any part of one or both forelimbs. Rearlimb movement is the movement of any part of one or both rearlimbs. Mouth movement is the opening and closing of the mouth. Lick movement is the protrusion of the tongue outside of the oral cavity . Facial wipe is the placement of one or both forepaws against the side of the head with movement of the forepaw(s)in a rostral direction incontact with the face. These seven movements, trunk movement and tail movement provided a measure of Total Activiy. The duration of Total Activity, Head(Turn),Head(EUD),Forelimb, and Rearlilnb, and the frequency of Mouth, Lick, and Facial Wipe were obtained in both sessions. The latency was recorded only in the odor presentation session.
Statistical Analysis
In this experiment, a two-way repeated-measures analysis of variance(ANOVA) with Group and Session was applied in duration and frequency. Post-hoc comparisons were conducted with F test for simple main
effects and Tukey’s HSD test. The latency in odor presentation session was analyzed with a one-way ANOVA with Group and HSD test.
Results and Discussion
The AF pups had the distinct characteristics in Total Activity, Head(Turn),and Head (EUD)as shown in Fig. 1. In duration of
Total Activlty, AF pups were more active in the odor presentation session than SAL, DW, and NS pups. The ANOVA indicated the significant difference of Group(F=3.42, df=3/124, p<.05; HSD test, the AF pups were different from SAL pups, p<.05),Session (F=41.54, df=1/124, p<.01),and Group × Session(F=9.58, df=3/124, p<.01; F test for simple main effects, Session was significant in Group, p<.01, and HSD test, the AF pups were more active in the presentation session than the baseline session, p<.01).It is clear the odor of amniotic fluid activates overall body movements.
Head(Turn)and Head(EUD),assessed s e p a r a t e l y i n t h i s e x p e r i m e n t , w e r e remarkably different from each other. The duration of Head(Turn)decreased in the odor presentation session in AF pups. The ANOVA indicated the significant difference of Session(F=12.34, df=1/124, p<.01)and Group × Session(F=5.56, df=3/124, p<.01; F test for simple main effects, Session was significant in Group, p<.0l, and HSD test, the AF pups were less active in the presentation session than the baseline session, p<.01).In compared to Head
(Turn),duration of Head(EUD)increased in the odor presentation session. The ANOVA indicated the significant difference of Group (F=8.12, df=3/124, p<.01; HSD test, the AF
pups were significantly more active than SAL, DW, and NS pups, ps<.01), Session(F=147.40, df=1/124, p<.01),and Group × Session (F=13.70, df=3/124, p<.01; F test for simple
main effects, Group was significant in the odor presentation session, p<.01, HSD test, the AF pups were more active than SAL, DW, and NS pups, ps<.01).These results show that two kinds of head movement play different roles for responding to the familiar odor, amniotic fluid. The role of head turn movement would be to search and orient the source of odor with its side to side motion. Once the odor is located, head extension and up-down movements will be activated to approach it instead of head turn movement. The decrease of Head(Turn)and increase of Head(EUD) in AF pups in odor presentation session indicate this relationship. It means that the odor of amniotic fluid is the familiar and attractive stimulus for newborn pups.
Pups showed Lick were 4, 4, 6, and 3 in AF, SAL, DW, and NS groups, respectively, in both sessions. Even in the baseline session, they licked their toe, tail and forepaw in the same way as in the odor presentation session. There was no group difference in duration of Forelimb and Rearlimb, and frequency of Mouth. In latency, no group difference was found in any movements. Facial Wipe, regarded as a rejective or aversive response
for perinatal pups(Browne, et al., 1994),was not analyzed because of less than two pups in each group. The experimental situation would not be unpleasant for them.
Experiment 2
Effects of stimulation to body areas with mother's fur on body movements
Mother’s ventral fur, usually wet with amniotic fluid, blood, and saliva at parturition, stimulates the facial and back areas of the newborn pups before the initial nipple grasp response. Nose and perioral areas of the face would be important for nipple searching, as the odor presentation to nose area promotes a c t i v e h e a d m o v e m e n t a s s h o w n i n Experiment l and the stimulation to perioral area with artificial nipple induces nipple grasp response and mouthing(Robinson & Smotheman, 1994).These two areas seem to be sensitive in fetuses(Robinson et al., 1992). Experiment 2 was designed to investigate effects of stimulation to the back, nose, and perioral areas with ventral fur from around the nipple area of mother on body movements of cesarean delivered pups.
Methods
SubjectsA total of 102 cesarean delivered pups from l7 female rats was tested. They were assigned to one of six groups according to the stimulated area(back, nose, and perioral) and fur(dry and wet): Group BD, BW, ND, NW, PD, and PW(17 pups in each group). Fur brush
Fur was obtained from around the nipple area of a E21 donor female(1.5mm ×1.5mm). The root was tied with No.1 silk thread, and inserted into a pipette tip(Eppendorf pipette tip 1-100μl).Then the tip was inserted into a
stainless tube(HypoTube-Type 316 S/S-13 Ga)and cut at the point which inner diameter was 1.2mm to make a round shape of the brush and to bring the area close to the collected area of mother’s fur. The small brush was designed to stimulate the restricted area of pups and to observe their movements in detail.
Procedure
Three hours after delively, each pup was placed in a plastic container wamed at 32℃ by a heating pad in an incubator. It was tested for 6 minutes, 3 minutes baseline session and 3 minutes stimulation session. Its spontaneous body movements were recorded in the baseline session. In the fur stimulation session, responses to the stimulation to back, nose, and perioral areas with a dry or wet brush were recorded. The stimulated areas were the upper half of dorsal body of back area, the viblissal pad of nose area, and the aperture of the mouth of perioral area.
Stimulation involved the gentle placement of the brush in contact with those areas. When the responses were active and vigorous, the placement was stopped temporarily until being quiet, not to stimulate the different area. A wet brush was wet with distilled water wamed at 32℃ . Behavioral recordings were similar to those in Experiment 1.
A three-way reapeated-measures ANOVA with Area, Fur, and Session was applied in duration and frequency. Post-hoc comparisons were conducted with F tests for simple main effect and Tukey’s HSD test. Latency was recorded in the stimulation session and compared with a two-way ANOVA, F tests for simple main effects, and the HSD test. The data was transformed into logarithms when the variance was a function of the mean. A chi-square test was used in analyzing Lick in the stimulation session.
Results and Discussion
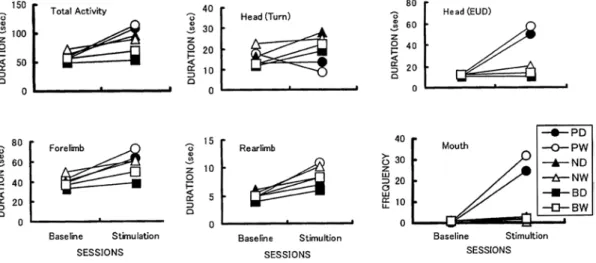
Duration and FrequencyWhen three areas were stimulated with the brush, Total Activity increased more in the perioral area than the other areas(Fig. 2). The ANOVA indicated the significant main effect of Area(F=19.45, df=2/96, p<.01; HSD test, perioral and nose areas were different from back area ps<.01), Fur(F=4.49, df=1/96, p<.05),and Session(F=94.73, df=1/96, p<.01).The interaction effect of Area × Session was also different(F=23.87, df=2/96, p<.01; F test for simple main effects, Area was significant in the fur stimulation session, p<.01, HSD test, three areas were different each other, and the perioral area was the most effective for increasing duration, ps<.01).It is clear that the brush of mother’s ventral fur from around the nipple area has the strong effect on activating body movements when it stimulates the perioral area.
The duration of Head(Turn)increased in back and nose areas but decreased in perioral area in the fur stimulation session. The ANOVA indicated the significant main effect of Area(F=13.10, df=2/96, p<.01; HSD test, perioral and nose areas were different from back area, ps<.01).The interaction effects of
Area × Session(F=5.42, df=2/96, p<.01; F test for simple main effects, Area was significant in the fur stimulation session, p<.01, HSD test, nose area was different fom perioral and back areas, p<.01 and p<.05)and Fur × Session(F=6.72, df=1/96, p<.01; F test for simple main effects, Session was significant in dry fur, p<.01, HSD test, dry fur activates the movement in the fur stimulation session) were found.
In compared to Head(Turn),Head(EUD) was characterized by the increased duration in perioral area. The duration in back area did not increase in the fur stimulation session. The wet fur activated Head(EUD)more than dry fur. The ANOVA indicated the significant main effect of Area(F=61.60, df=2/96, p<.01; HSD test, perioral area activated the movement more than back area, p<.01), Fur(F=3.99, df=1/96, p<.05),and Session(F=180.29, df=1/96, p<.01).The interaction effect of Area × Session was significant(F=105.64, df=2/96, p<.01; F test for simple main effects, Area was significant in the fur stimulation session, p<.05, HSD test, three areas were different each other, and the perioral area was the most effective in increasing duration and the back area was the least, ps<.01).
The duration of Forelimb increased especiaIly in perioral area in fur stimu!ation session. The wet fur activated Forelimb more than the dry fur. The ANOVA indicated the significant main effect of Area(F=8.40, df=2/96, p<.01; HSD test, the perioral and nose areas were different from back area, ps<.01),Fur(F=5.10, df=1/96, p<.05),and Session(F=42.89, df=1/96, p<.01).The interaction effect of Area × Session was signiflcant(F=8.80, df=2/96, p<.01; F test for simple main effects, Area was significant in the fur stimulation session, p<.01, HSD test, perioral and nose areas were more effective in increasing duration, ps<.01).
The duration of of Rearlimb increased in the fur stimulation session. The ANOVA indicated the significant main effect of Session(F=14.50, df=1/96, p<.01).
These findings in duration indicate that perioral area is more sensitive to mother’s fur and that pups inhibit Head(Turn)and activate Head(EUD).The wet fur has some effects of activating Total Activity, Head (EUD)and Forelimb when the perioral area
is stimulated.
The frequency of Mouth extremely increased in perioral area, but not in back and nose arears. The ANOVA indicated the main effect of Area(F=133.52, df=2/96, p<.01; HSD test, perioral area activated the movement more than nose and back areas, ps<.01)and Session(F=160.766, df=1/96, p<.01).The interaction effect of Area × Fur(F=4.49, df=2/96, p<.05; F test for simple main effects, Fur is significant in perioral area, p<.01), Area × Session(F=160.77, df=1/96, p<.01; F test for simple main effects, Area was different in the fur stimulation session, p<.01, HSD test, perioral area was different from nose and back areas, p<.01),and Area × Fur × Session (F=257.26, df=2/96, p<.05; F test for simple
simple main effects, Fur was significant in dry and wet fur in the fur stimulation session,
ps<.01, HSD test, perioral area activated the movement than nose and back areas, ps<.01) were found.
Few pups showed Lick in back and nose areas: one pup and two pups in BD and BW in the fur stimulation session, one pup both in ND and ND in the baseline session. A chi-square test was used to analyze the difference between PD and PW pups only in the stimulation session, because 3 and 8 pups showed Lick in that session and no pup in the basehne session. The significant effect was found between the two groups, χ2(1)=3.841,
p<.05. Facial Wipe was not statistically analyzed because less than three pups in each group showed Facial Wipe.
These results in frequency indicate that perioral stimulation activates Mouth and Lick movements connected with nipple grasping. Before the nipple grasping, newborn pups usually respond to the perioral stimulation with mother’s ventral fur , open the mouth, and lick the fur after delivery. Few Facial Wipe shows that the fur is the different kind of stimulus for the pups from the artificial nipple used in Smotherman, et al.(1997)and that the experimental condition was not unpleasant for the pups because that response is thought to be an aversive one(Browne et a., 1994).
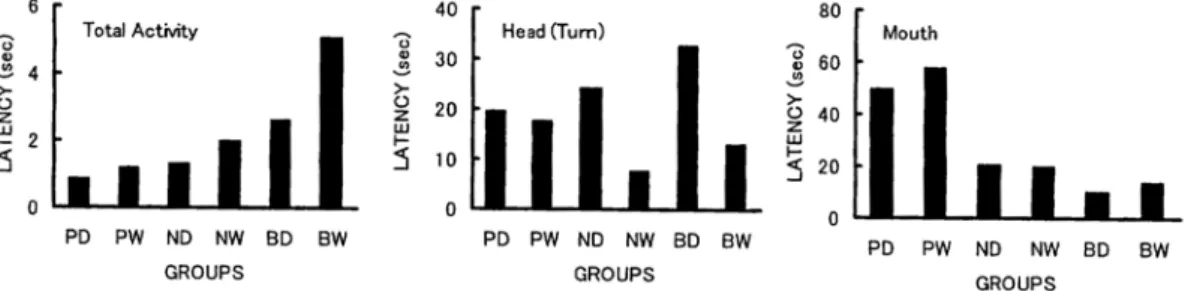
Latency in the fur stimulation session
The latency of Total Activity in nose and perioral areas was shorter than that in back area(Fig. 3).The ANOVA indicated the significant main effect of Area(F=3.76, df=2/96, p<.05; HSD test, the back area had longer latency than nose area, p<.05). In Head(Turn),wet fur had shorter latency than dry fur. The ANOVA indicated the significant main effect of Fur(F=4.49, df=1/96, p<.05).In Head(EUD),perioral and nose areas had shorter latency than back area. The ANOVA indicated the significant
main effect of Area(F=7.40, df=2/96, p<.01; HSD test, the nose and perioral areas had shorter latency than back area, ps<.01). In Mouth, back and nose areas had longer latency and wider variation than perioral area. The ANOVA with transformation into logarithms indicated the significant main effect of Area(F=121.37, df=2/96, p<.01; HSD test, the perioral area had shorter latency than nose and perioral areas, ps<.01).
There was no difference in Forelimb and Rearlimb. The latency of Lick was not analyzed.
These results indicate that perioral stimulation causes rapid response especially in head extension and up-down movements and mouth movement connected with nipple grasping. The head turn movement was occurred faster in wet fur than dry fur. It suggests that wet fur of the mother has a role for permitting the pups orient her ventral area promptly.
Experiment3
Effects of fur presentation and perioral stimulation with fur on body movements In Experiment 2, the perioral stimulation with mother’s fur was effective to activate several movements. The fur has its specific smells of mother such as amniotic fluid, saliva. and milk, which may attract and orient pups to mother after birth. Experiment 3 was conducted to examine the effect of the odor of mother’s fur, and the combined effect of odor
and perioral stimuiation with fur on body movement of cesarean delivered pups.
Methods
A total of 45 cesarean delivered pups from 15 female rats was tested. They were assigned to one of three groups according to t h e c o n d i t i o n ( f u r s t i m u l a t i o n , f u r presentation, and no presentation): Group FS, FP and NP, 15 pups in each group.
Three hours after delively, each pup was placed in a plastic container warmed at 32 ℃ as described in Experiment 2. It was tested for 9 minutes, 3 minutes baseline session, 3 minutes fur presentation session, and 3 minutes fur stimulation session. In FS pups, a dry fur brush was presented close to their nose area in the fur presentation session, and their perioral area was stimulated with it in the fur stimulation session. In FP pups, the dry fur brush was presented in the fur presentation and fur stimulation sessions. In NP pups, their spontaneous movements were observed in three sessions with the fur brush. Behavioral recordings were similar to those in Experiment 1. A two-way repeated-measures ANOVA with Group and Session, F test for simple main effects and HSD test were applied. Latency were recorded in the fur presentation and fur stimulation session. The data was transformed into logarithms when the variance was a function of the mean. For combined effect of fur presentation and perioral stimulation, a two-way
measures ANOVA with Group and Session in duration and frequency and t test in latency were applied FS pups in this experiment and PD pups in Experiment 2.
Results and Discussion
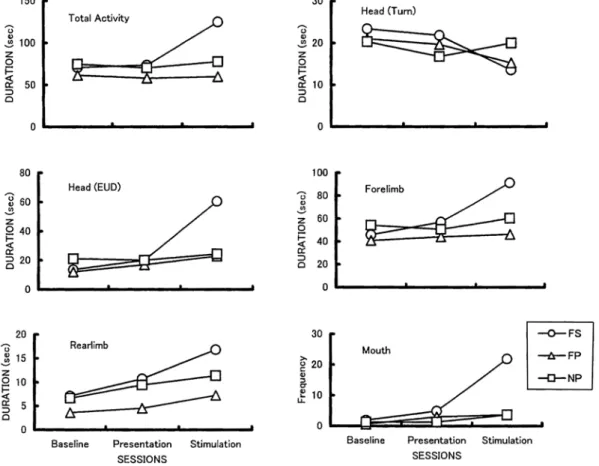
Duration and FrequencyTotal activity was characterized by the increased duration of FS pups in fur stimulation session(Fig. 4).The two-way ANOVA indicated the significant effect of Group(F=4.98, df=2/42, p<.05; HSD test, FS pups was more active than FP pups, p<.01), Session(F=19.69, df=2/84, p<.01; HSD test, pups responded more in the fur stimulation session than the fur presentation and baseline sessions, ps<.01), and Group × Session(F= 15.03, df=4/84, p<.01; F tests for simple main effects, Group was significant in the
stimulation session, p<.01, HSD test, FS pups were more active than FP and NP pups, ps<.01).
There was no significant difference in Head (Turn)which decreased in the presentation
session in FP and FS pups. On the other hand, Head(EUD)increased in the fur presentation session in FS pups. The ANOVA indicated the significant effect of Group (F=7.65, df=2/42, p<.01; HSD test, FS pups
were more active than FP and NP pups, p<.01 and p<.05),Session(F=59.31, df=2/84, p<.01; HSD test, pups responded more in the fur stimulation session than the fur presentation and baseline sessions, ps<.01),and Group × Session(F=27.31, df=4/84, p<.01; F test for simple main effects, Group was significant in the fur stimulation session, p<.01, HSD test, FS pups were more active than FP and NP pups, ps<.01).
The duration of Forelimb increased in FS pups in the fur presentation and stimulation sessions. The ANOVA indicated the significant effect of Session(F=16.82, df=2/84, p<.01; HSD test, pups responded more in the f u r s t i m u l a t i o n s e s s i o n t h a n t h e f u r presentation and baseline sessions, ps<.01) and Group × Session(F=7.92, df=4/84, p<.01; F test for simple main effects, Group was significant in the stimulation session, p<.01, HSD test, FS pups were more active than FP and NP pups, ps<.01).
The duration of Rearlimb increased in the stimulation session in all the groups. The ANOVA indicated the significant effect of Group(F=3.86, df=2/42, p<.05; HSD test, FS pups were more active than FP pups, p<.05) and Session(F=13.21, df=2/84, p<.01; HSD test, pups responded more in the fur stimulation session than the fur presentation and baseline sessions, ps<.01).
In the frequency of Mouth, FS pups
responded more in the fur stimulation session than FP and NP pups. The ANOVA indicated the significant effect of Group(F=13.34, df=2/42, p<.01; HSD test, FS pups were more active than FP and NP pups, ps<.01),Session (F=43.86, df=2/84, p<.01; HSD test, pups
responded more in the fur stimulation session than the fur presentation and baseline sessions, p<.01), and Group × Session(F=22.27, df=4/84, p<.01; F test for simple main effects, Group was significant in the fur stimulation session, HSD test, FS pups were more active than FP and NP pups, ps<.01).The Lick and Facial Wipe were not analyzed because few pups showed these movements.
These results indicate that fur stimulation to the perioral area is the most effective in activating several movements but not fur odor.
Latency in the presentation and stimulation sessions
In Head(EUD),latency of FS pups was the shortest in the fur stimulation session(Fig. 5).The ANOVA indicated the significant effect of Session(F=6.77, df1/42, p<.01)and Group × Session(F=3.38, df=2/42, p<.05; F test for simple main effects, Group was significant in the fur stimulation session, p<.05; HSD test, FS pups had shorter latency than NP pups, p<.05).
In Forelimb and Rearlimb, latency in the fur stimulation session was shorter than that in the fur presentation session in all the groups. The ANOVA indicated the significant effect of Session(F=5.84 and 9.86, dfs=1/42, p<.05 and p<.01).
In Mouth, latency of FS pups in the fur stimulation session was shorter than that of FP and NP pups. The ANOVA indicated the significant effect of Group(F=3.53, df=2/42, p<05; HSD test, FS pups had shorter latency than FP and NP pups, ps<.05),Session (F=13.35, df=1/42, p<.01),and Group × Session(F=5.62, df=2/42, p<.01; F test for simple main effects, Group was significant in the fur stimulation session, p<.01, HSD test, FS pups had shorter latency than FP and NP pups, ps<.01).
The results that FS pups had shorter latency than FP and NP pups indicate the fur stimulation to the perioral area is more effective for rapid response.
Combined effect of the presentation of dry fur odor and the perioral stimulation with dry fur The PD pups in Experiment 2 received the perioral stimulation with dry fur for 3 min like FS pups in this experiment. The difference is that FS pups had 3 min fur presentation session prior to 3 min fur stimulation session. If fur odor and perioral stimulation have the combined effect on body movements, FS pups will be different fom PD pups in the stimulation session. Between FS and PD pups, there seems to be some difference in average
duration of Total Activity(124.7 and 108.7 sec),Head(EUD)(60.4 and 49.5 sec), Forelimb(91.2 and 63.7 sec),and Rearlimb (16.8 and 6.9 sec)in the fur stimulation
session.
A two-way repeated-measures of ANOVA indicated the significant main effect of Session (baseline and stimulation)in Total Activity
and Head(EUD)(F=173.21 and 215.83, df=1/30, ps<.01).In Forelimb, the significant effect was found in Group(F=5.30, df=1/30, p<.05),Session(F=93.51, df=1/30, p<.01), and Group × Session(F=8.23, df=1/30, p<.01; F tests for simple main effect, Group was significant in the stimulation session, p<.01). In Rearlimb, the significant effect was found in Group(F=7.68, df=1/30, p<.01),Session (F=12.41, df=1/30, p<.01),and Group × Session(F=5.63, df=1/30, p<.05; F tests for simple main effect, Group was significant in the stimulation session, p<.01).No significant difference was not found in Head(Turn)and Mouth.
The FS pups seem to have shorter latency than PD pups in Head(EUD)(3.6 and 6.7sec) and Rearlimb(7.0 and 15.1 sec)in the stimulation session. The t test showed a significant difference in Head(EUD)(t=3.58, df=30, p<.01).The other movements were not significant.
These results indicate the combined effect of the presentation of fur odor and the perioral stimulation with fur: the odor presentation prior to the perioral stimulation with dry fur is effective for the activation of Forelimb and Rearlimb and the rapid response of Head (EUD)which are available for approaching
and probing mother’s nipple.
Experiment 4
Combined effects of amniotic fluid and stimulation on body movements in cesarean delivered pups
At parturition, amniotic fluid and mother’s ventral fur are usually combined and stimulate the perioral area of pups before the initial nipple grasping. In Experiment l and 2, the odor of amniotic fluid and the mother’s fur were separately examined and found to be effective for activating several movements. When odor of mother’s fur and its perioral stimulation were combined in Experiment 3, the cumulative effect on several movements was suggested. It indicates the possibility that the odor and taste of amniotic fluid and the perioral stimulation with mother’s fur will be more effective for activating movements when they are combined. Experiment 4 was conducted to examine the combined effects of the odor of amniotic fluid and perioral stimulation with mother’s fur wet with the fluid on body movement of cesarean delivered pups, to approach the natural setting before the initial nipple grasping.
Methods
A total of 81 cesarean delivered pups from 27 female rats was tested. They were assigned to one of three groups according to the condition(fur and amniotic fluid, fur and saline, fur and distilled water): Group FAF, FSAL, and FDW, 27 pups in each group. Three hours after delivery, each pups was placed in a plastic container warmed at 32 ℃ as described in Experiment 2. It was tested for 9 minutes, 3 minutes baseline session, 3 minutes presentation session, and 3 minutes stimulation session. In FAF pups, a fur brush wet with amniotic fluid was presented close to their nose area in the presentation session, and the perioral area was stimulated with it in the stimulation session. In FSAL and FDW pups, a fur brush wet with saline or disti1led water was presented in the presentation session, and the perioral area was stimulated with it in the stimulation session. Behavioral
r e c o r d i n g s w e r e s i m i l a r t o t h o s e i n Experiment l.
A two-way repeated-measures ANOVA with Group and Session, F test for simple main effects, and HSD test were applied in duration and frequency of movements except licking. In Licking, one-way ANOVA was used in the stimulation session because only three pups showed this movement in the baseline and presentation sessions. Facial Wipe was not analyzed because only two pups in FDW and FSAL pups showed this movement in the baseline and stimulation sessions. Latency was recorded in the presentation and stimulation sessions and analyzed by a two-way repeated-measures ANOVA with Group and Session. To assess t h e c o m b i n e d e f f e c t o f t h e f u r o d o r presentation and perinatal stimulation with wet fur and of the odor presentation of amniotic fluid and mother’s fur, a two-way repeated-measures ANOVA with Group and Session in dulation and frequency and t test in latency were applied in FDW or FAF pups in this experiment and PW pups in Experiment 2 or AF pups in Experiment 1.
Results and Discussion
Duration and FrequencyTotal Activity increased in the stimulation session in all the groups(Fig.6).The ANOVA indicated the significant effect of Group(F=5.02, df=2/78, p<.05; HSD test, FAF pups were more active than FSAL pups, p<.01), Session(F=153.23, df=2/156, p<.01; HSD test, pups responded more in the stimulation session than the presentation and baseline sessions, ps<.01),and Group × Session(F=3.12, df=4/156, p<05; F test for simple main effects, Group was significant in the presentation and stimulation sessions, ps<.01, HSD test, FAF pups were more active than FSAL pups in the presentation and
stimulation sessions and than FDW pups in the stimulation session, ps<.01 and p<.05). In Head(Turn),duration of FAF pups decreased more than that of FSAL and FDW pups in the presentation and stimulation sessions. The ANOVA indicated the significant effect of Session(F=14.78, df=2/156, p<.01; HSD test, pups responded less in the presentation and stimulation sessions than the baseline session, ps<.01), and Group × Session(F=2.81, df=4/156, p<.05; F test for simple main effects, FAF pups were significant in Session, p<.01, HSD test, they were less active in the presentation and stimulation sessions than in the baseline session, ps<.01).
Head(EUD),which increased only in FAF pups in the presentation session, increased in all the pups in the stimulation
session. The ANOVA indicated the significant effect of Group(F=16.63, df=2/78, p<.01; HSD test, FAF pups were more active than FSAL and FDW pups, ps<.01),Session(F=243.98, df=2/156, p<01; HSD test, three sessions were different each other and pups in the stimulation session responded the most but those in the baseline session did the least, ps<.01),and Group × Session(F=10.80, df=4/156, p<.01; F test for simple main effects, Group was significant in the presentation and stimulation sessions, ps<01, HSD test, FAF pups were more active than FSAL and FDW pups in both the sessions, ps<.01).
In Forelimb and Rearlimb, duration of all the pups increased in the stimulation session. The ANOVA indicated the significant effect of Session in Forelimb(F=115.60, df=2/156, p<.01; HSD test, pups responded more in the Fig. 6 Mean duration and frequency of body movements in FAF, FSAL, and FDW pups.
stimulation session than the presentation and baseline sessions, ps<.01),and Rearlimb (F=33.02, df=2/156, p<.01; HSD test, pups
responded more in the stimulation session than the presentation and baseline sessions, ps<.01).
In Mouth, frequency of all the pups increased in the stimulation session, and FAF pups were the most active. The ANOVA indicated the significant effect of Group (F=10.18, df=2/78, p<.01; HSD test, FAF pups
were more active than FSAL and FDW pups, ps<.01),Session(F=200.54, df=2/156, p<.01; HSD test, pups responded more in the stimulation session than the presentation and baseline sessions, ps<.01),and Group × Session(F=11.49, df=4/156, p<.01; F test for simple main effects, Group was significant in the stimulatlon session, p<.01, HSD test, FAF pups were more active than FSAL and FDW pups, ps<.01).
In Lick, the average frequency of FAF pups (11.8)was higher than FDW and FSAL pups (4.0 and 5.9, respectively)in the stimulation
session. The one-way ANOVA in the stimulation session indicated the significant effect for Group(F=4.26, df=2/78, p<.05; HSD test, FAF pups were more active than FSAL and FDW pups, ps<.01).
These results indicate that stimulation to the perioral area with amniotic fluid has the obvious combined effect on activating almost all the movements. Higher frequency of Lick in FAF pups would indicates some important role of mother’s fur and amniotic fluid for nipple grasping which is connected with nipple attachment.
Latency in the presentation and stimulation sessions
All the pups had shorter latency in the stimulation session than in the presentation session. The ANOVA indicated the significant effect of Session in Total Activity(F=30.02,
df=1/78, p<.01),Head(Turn)(F=17.27, df=1/78, p<01),Head(EUD)(F=30.00, d f = 1/ 78, p < . 01), F o r e l i m b ( F = 56.77, d f = 1/ 78), p < . 01), R e a r l i m b ( F = 69.66, d f = 1/ 78, p < . 01), a n d M o u t h ( F = 95.55, df=1/78, p<.01).There was no dlfference in Group and Group × Session.
Combined effect of fur odor and perioral stimulation with wet fur
The PW pups in Expehment 2 received the periolal stimulation with wet fur for 3 min like FDW pups in this experiment. The difference is that FDW pups had 3 min fur presentation session prior to 3 min simulation session. .If the odor of fur and distilled water and the perioral stimulation have the combined effect on body movements, FDW pups will be different from PW pups in the stimulation session. Between FDW pups and PW pups, there seems to be some difference in average duration of Head(Turn)(15.2 and 8.3 sec) and Head(EUD)(49.0 and 57.5).
A two-way repeated-measures of ANOVA indicated the significant main effect of Session (baseline and stimulation)in Total Activity,
Head(Turn),Forelimb, Rearlimb, and Mouth(F=127.28, 15.50, 77.35, 40.13, and l67.47, dfs=1/42, ps<01).In Head(EUD), the significant effect was found in Session (F=171.67, df=1/42, p<.01)and Group × Session(F=5.02, df=1/42, p<.05; F test for simple main effect, Group had a significant tendency in the stimulation session, p<.10). The FDW pups seem to have shorter latency than PW pups in Rearlimb(5.6 and l1.9 sec)and longer latency in Mouth(50.3 and 31.8 sec)in the stimulation session. The t test showed a significant tendency in Mouth (t=1.94, df=42, p<.10).
The results indicate that the combined effect of fur odor and perioral stimulation of wet fur exists but is small.
Combined effect of the odor of amniotic fluid and mother’s fur
When the odor of amniotic fluid was presented with cotton for 5 min in Experiment 1, Total Activity and Head(EUD)increased but Head(Turn)decreased. These are in consistent with the results in this experiment where the odor of amniotic fluid was presented with mother’s fur for 3 min. The difference between these experiments was the observation period and the size of material which resulted in the different amount of amniotic fluid leading to its odor density. The average duration of Total Activity, Head (Turn),and Head(EUD)per 3 min was
86.9, 12.7, and 49.5 sec in AF pups, and 80.4, 12.2, and 35.2 sec in FAF pups in this experiment. The t test indicated the significant difference in Head(EUD)(t=2.65, df=57, p<.05).It would be shown that the odor of amniotic fluid and mother’s fur has little combined effect on the activation of pups’ response, and that the odor of amniotic fluid only has a stronge effect on it.
General Discussion
Mother’s ventrum was found to be the most effective for activating upside-down and probing movements in newborn rats several days after birth(Polan & Hofer, 1999).The mother’s fur is one of the skin stimulus inevitably received by the newborns in many mammals before the first nipple grasping, and has its specific smell which contains amniotic fluid, blood, saliva and mother’s ventral skin substances at parturition. In this study, we made the small fur brush of mother’s ventral fur, because it enables us to stimulate the restricted area of pups and to observe their movements in detail. We also attached importance to applying natural material received by the newborns immediately after birth, as fur brush of other animals and
artificial fur were less effective for the activation of movements in cesarean delivered rats(Kodama & Smotherman, 1997).Results in Experiment 2, 3, and 4 indicated that the fur brush caused a lot of changes in body movements, in consistent with our previous study(Kodama & Smotherman, 1997),and that it made it easy to detect several odor and tactile effects with amniotic fluid on body movements. These indicate that the brush is a useful appliance to analyze the effects of several stimuli characterized by the parturient mother on the body movement in newborn pups. Mother’s ventral fur would have an important role to activate the behavior of pups immediately after birth.
We classified head movement into two categories: head turn movement and head extension and up-down movements. Head turn movement has been only described as side-to-side head movement in nipple probing (Blass et al, 1988; Blass & Teicher, 1980; Hall,
1990; L a r s o n & S t e i n , 1984) a n d h e a d extension as straightened movement in oral grasp response(Smotherman, et a1., 1997). These movements were required for quantitatively assessing the role of mother's fur and amniotic fluid in the present study. Results indicated their different features: head turn movement increased when the nose area was stimulated with mother’s fur(Experiment 2)and decreased when the odor of amniotic fluid was presented to pups and their perioral area was stimulated with mother’s fur and amniotic fluid(Experiment 4),while head extension and up-down movements increased when the odor of amniotic fluid was presented to pups(Experiment 1) and when their perioral area was stimulated with mother’s fur (Experlment 2 and 3)and amniotic fluid (Experiment 4).It is clear that head turn
movement and head extension and up-down movements have different functions and should be distinguished from each other. In
natural situation, head turn movement could be more activated when pup’s nose reaches mother’s fur, and would result in probing movement. Head extension and up-down movements would be activated by the odor of amniotic fluid and result in orienting response to nipple when the perioral area is stimulated by the fur.
Amniotic fluid is one of the important stimuli for suckling behavior(Blass & Teicher, 1980),but it is always presented with several stimuli containing mother’s fur at parturition. The present study separated the effect of fur odor from the odor of amniotic f l u i d a n d c o m b i n e d t h e s e o d o r s . I n Experiment 3, fur odor itself had little effect in any movements. It is in contrast to the odor of amniotic fluid which reduced head turn movement and increased total activity and head extension and up-down movements when it was presented with cotton(Experiment 1) or mother’s fur(Experiment 3).Schaal, Marlier, & Soussignan(1995)also found that 2-day-old human newborns preferred and oriented the odor of amniotic fluid. These findings indicate that the odor of amniotic fluid, familiar to fetuses, is one of the attractive stimuli which permit the newborns to orient and locate its source(mother)by specific head movement in mammals.
The other odor stimuli which attract the newborns and elicit nipple attachment are found to be mother’s saliva and nipple extract in 2-day-o1d rats(Singh & Hofer, 1978; Teicher & Blass, 1977),the odor of abdomen of lactating mother in l to 3-day-old rabbits (Courlaud & Schaal, 2000),and the odor of
mother’s breast in 2-day-old human newborns (Makin & Porter, 1989).These stimuli are
not familiar in prenatal period, but would become familiar in the process of nursing after birth. In mammals, the major odor stimulus to attract the newbons immediately after birth would be amniotic fluid familiar in prenatal
period, and then the odor of several stimuli characterized by the lactating mother would become familiar in the process of nursing. The following explanation has been made in relation to fur stimulation in the newborns. When the snout of postnatal rat pups and kittens touches the fur of the anesthetized mother or artificial fur, they move their head from side-to-side, rub their nose in the mother’ s skin and fur, scan them with their snout, locate the nipple with neck flexion and probe the nipple(Blass & Teicher, 1980; Hall,1990; L a r s o n & S t e i n , 1984). A l t h o u g h t h e explanation have not distinguished the effect of dry and wet fur from that of stimulated body areas, it is important to study their relation because the mother’s ventral fur is mostly wet at parturition and lateral or dorsal fur is dry and because the fur stimulates several body areas of newborn pups. In regard to the fur, the wet fur softer than dry fur causes more active head extension and up-down movements and the same level of mouth m o v e m e n t a s d r y f u r ( K o d a m a & Smotherman, 1997).It suggests that each fur has some different effects on body movements. In the present study, we investigated the stimulation effects of dry and wet fur and the differential effects on body movements in Experlment 2. Results indicated that dry fur activated total activity and wet fur head extension and up-down movements and forelimb movement. It is remarkable that the perioral stimulation with wet fur caused lick movement more frequently than that with dry fur. In regard to body areas, the stimulation of back area did not have any prominent effects, but that of nose area was effective on total activity and head turn movement. The perioral stimulation was the most effective in total activity, head extension and up-down movements, mouth movement and lick movement. These findings would give us the following stimulus-behavior sequence. When
face area(nose and perioral)is stimulated with mother’s dry fur, pups move actively at first. Then they turn their head when their nose area is stimulated. It enables them to touch mother’s ventral wet fur with their perioral area, to let their head extend, and to move it up and down. This sequence leads them to opening mouth and licking the fur, and finally to grasping nipple as a natural consequence when their mouth reaches mother’s nipple.
Several stimuli are combined and stimulate the newborns simultaneously when amniotic fluid is attached to the mother’s fur and their perioral area is stimulated. But few studies have investigated the combined effect because of the difficulty of combining several effects intentionally. The present study assessed several combined effects systematically in four experiments. In Experiment 2 and 3, the combined effect of fur odor and perioral stimulation with dry fur was found in activating forelimb and rearlimb movements and eliciting rapid head extension and up-down movements. In Experiment 2 and 4, the combined effect of odor and perioral stimulation with wet fur was found in reducing head extension and up-down movements a little. In Experiment l and 4, there is little combined effect of the odor of amniotic fluid and mother’s fur: the odor of amniotic fluid itself had a strong effect. Experiment 4 was designed to combine the odor of amniotic fluid and mother’s fur and the perioral stimulation with the fur wet with amniotic fluid. Pups showed strong combined effect, which reduced head turn movement, but activated total activity, head extension and up-down movements, mouth movement, and lick movement. It is clear that several combined effects exist and that the combined effect of the odor of amniotic fluid and perioral stimulation with wet fur with the fluid is the most effective for activating newborn’s
movements. Activation of head extention and up-down movements would be more useful for probing mother’s ventral surface in natural situation.
The origin of mouth movement and sucking is in prenatal period in mammals. Fetuses open and close their mouth, move their tongue and lips, and suck when their lips are stimulated with a hair and an artificial nipple (Angulo y González, 1932; Humphrey, 1962;
Smotherman, et a1., 1993)or during their spontaneous body movements(Narayanan, et al., 1971; Smotherman, Richards & Robinson, 1984).Adding to these findings, the present study made it clear that fetuses spontaneously lick their toe, forepaw, and tail. These seem to have common features similar to those of mother’s nipple: sma11, thin at the pointed end, soft and warm. It is assumed the fetuses spontaneously open their mouth, 1ick and grasp several body areas as a preparatory stage of postnatal sucking when those areas touch the mouth of fetuses as a result of body movement. The finding that stimulation of perioral area with artificial nipple elicits nipple grasp response in rat fetuses(Browne, et al., 1994)and thumb sucking in human fetuses (Birnholz, 1988)would cofirm our assumption. Mouth and lick movements were frequently e1icited when the perioral area of pups was stimulated with the small and thin brush to which amniotic fluid was attached. This stimulus-response relationship would be important for the explanation of the onset of nipple attachment as follows. The perioral face area, upper and lower lips, oral cavity, and tongue would not be stimulated after birth but in prenatal period. They are frequently stimulated near term with the activation of fetal body movements. Fetuses open their mouth spontaneously or their perioral area is stimulated with their body areas, forelimb, rearlimb, or tail. After birth, newborn pups receive many novel stimuli
except the odor of amniotic fluid familiar in prenatal period. They orient and approach it with up-down and extention movements of head. Simultaneously, mother approaches, licks, hovers over, and nurses them(Stern, 1989).In these maternal care, pups’ perioral area is stimulated by the mother’s fur with the fluid, and its stimulation elicits the active body movements, especially head, mouth and lick movements. Opening the mouth means that mothe’s fur with amniotic fluid and the other fluid stimulates oral caviry and tongue and that pups taste the familiar amniotic fluid when they lick the fur. It may cause more active mouth and lick movements of pups. When their mouth touches the soft and thin nipple, they would grasp it. The ventral stimulation by pups induces nursing posture and milk ejection of mother(Stern, 1989; Stem, Dix, Bellomo, & Thramann, 1992; Stem & Johnson, 1990).These complex stimulus-response interaction between the mother and her pups would enhance the nipple attachment of pups and promote attachment mechanism between them.
References
Angulo y González, A.W.(1932).The prenatal development of behavior in the albino rat. Journal of Comparative Neurology, 55, 395-442. Becker, L.A., & Smotherman, W.P.(1996).
Manipulation of the dopamine system affects fetal and neonatal responses to an artificial nipple. Developmental Psychobiology, 29, 139-156.
Birnholz, J.C.(1988).On observing the human fetus. In W.P. Smotherman and S.R. Robinson (Eds.),Behavior of the fetus. New Jersey:
Telford Press, pp. 47-60.
Blass, E,M., Shuleikina-Turpaeva, K., & Luschekin, V.(1988).Sensory deteminants of nipple attachment behavior in 2-4-day-old kittens. Developmental Psychobiology, 21, 365-370. Blass, E.M., and Teicher, M.H.(1980).Suckling.
Science, 210, 15-22.
Browne. J.B., Robinson, S.R., & Smotherman, W. P.(1994).Fetal experience with milk or an artificial nipple alters appetitive and aversive responses to perioral cutaneous stimuli. Behavioral Neuroscience, 108, 606-613.
Coureaud, G., & Schaal, B.(2000).Attraction of newborn rabbits to abdominal odors of adult conspecifics differing in sex and physiological state. Developmental Psychobiology, 36, 271-281. Fox, M.W.(1965).The development of reflexes
and neuro-ontogeny of the mouse. Animal Behaviour, 13, 234-241,
Hall, W.G.(1990).The ontogeny of ingestive behavior: Changing control of components in the feeding sequence. In E.M. Stricker(Ed.), Handbook of behavioral neurology, vol 10, Neurobiology of food and fluid intake. New York: Prenum Press, pp.77-123.
Hofer, M.A., Shair, H., & Singh, P.(1996).Evidence that maternal ventral skin substances promote suckling in infant rats. Physiology and Behavior, 17, 131-136.
Humphrey, T.(1962).Some correlations between the appearance of human fetal reflexes and the development of the nervous system. In D.P. Purpura and J.P. Schade(Eds.),Progress in Brain Research: Growth and maturation of the brain , Volume 4. Amsterdam: Elsevier, pp. 93-135.
Kodama, N.(1993).Behavioral development and strain differences in perinatal mice(Mus musculus).Journal of Comparative Psychology, 107, 91-98.
Kodama, N., & Smotherman, W.P.(1997). Mechanism of nipple attachment in rats: A preliminary study of mother’s fur. Memoires of the Faculty of Education Shiga University, 47, 91-102(Japan).
Koffman, D.J., Petrof, E.S., Varlinskaya, E.L., & Smotheman, W.P.(1998).Thermal, olfactory and tactile stimuIi increase oral grasping of an artificial nipple by the newborn rat. Develop-mental Psychobiology, 33, 317-326.
Larson, M.A., & Stein, B.E.(1984).The use of tactile and olfactory cues in neonatal orientation and localization of the nipple. Developmental Psychobiology, 17, 423-436.
Makin, J.W., & Porter, R.H.(1989).Attractiveness of locating females breast odors to neonates. Child Development, 60, 803-810.
(1971).Prenatal development of spontaneous and evoked activity in the rat(Rattus Norvegicus Albinus).Behaviour, 12, 100-134. Polan, H.J., & Hofer, M.A.(1999).Maternally
directed orienting behaviors of newborn rats. Developmental Psychobiology, 34, 269-279. Robinson, S.R., Hoe1tzel, T.C.M., Cooke, K.M.,
Umphress, S.M., Murriish, D.E., & Smotherman, W.P.(1992).Oral capture and grasping of
artificial nipple by rat fetuses. Developmental Psychobiology, 25, 543-555.
Schaa1, B., Mar1ier, L., & Soussignan, R.(1995). Responsiveness to the odour of amniotic fluid in the human neonate. Biology of the Neonate, 67, 397-406.
Singh, P.J., & Hofer, M.A.(1978).Oxytocin reinstates maternal olfactory cues for nipple orientation and attachment in rat pups. Physiology and Behavior, 20, 385-389.
Smotherman, W.P., Arnold, H.M., & Robinson, S. R.(1993).Responses to ecologically relevant stimuli in the rat fetus: Interactive effects of milk and an artificial nipple. Developmental Psychobiology, 26, 359-374.
Smotherman, W.P., Goffman, D., Petrov, E.S., & Varlinskaya, E.L.(1997).Oral grasping of a surrogate nipple by the newborn rat. Develop-mental Psychobiology, 31, 3-17.
Smotherman, W.P., Richards, L.S., & Robinson, S. (1984).Techniques for observing fetal
behavior in utero: A comparison of chemomye-lotomy and spinal transection. Developmental Psychobiology, 17, 661-674.
Stern, J.M.(1989).Maternal behavior: Sensory, hormonal and neural determinants. In F.R. Brush & S. Levine(Eds.),Psychoendocri-nology. New York: Academic Press, pp, 105-226.
Stern, J.M., Dix, L., Bellomo, C., & Thramann, C. (1992).Ventral somatosensory determi-nants
of nursing behavior in Norway rats: Ⅱ . Role of nipple and surrounding sensations. Psycho biology, 20, 71-80.
Stern, J.M., & Johnson, S.K.(1990).Ventral somatosensory determinants of nursing behavior in Norway rats: I. Effects of variations in the quality and quantity of pup stimuli. Physiology and Behavior, 47, 993-1011.
Teicher, M.H, & Blass, E.M.(1977).First suckling response of the newborn albino rat: Th role of olfaction and amniotic fluid. Science, 198,
635-636.
Acknowledgements
This research was performed in 1996-1997 in the laboratory of Dr. W.P. Smotherman, Center for Developmental Psychobiology, Department of Psychology, Binghamton University-SUNY. I am greateful to Dr. Smotherman for giving me his advice and support.