Thesis
Contribution of reactive oxygen species to oleic acid-induced lung injury and preventive effect of Shoseiryu-to
on the lung injury
オレイン酸肺傷害における活性酸素種の関与 および小青竜湯の肺傷害防止効果
Changqing Yang 2003
熊本大学大学院薬学研究科
臨床薬学専攻 病態・生体情報学講座
楊 長青
論 文 要 旨
オレイン酸肺傷害における活性酸素種の関与および小青竜湯の肺傷害防止効果 臨床薬学専攻 病態・生体情報学講座(研究室) 楊 長青
肺血管透過性亢進による重篤な低酸素血症を伴なう急性呼吸窮迫症候群(acute respiratory distress syndrome, ARDS)は、近年の救命医療の進歩にも関わらず、
死亡率が 30%を上回る難治性疾患であり、予防・治療薬の開発が切望されている。オ レイン酸を小動物に静脈内投与すると、ARDS と病態生理学的に類似した肺傷害を惹き 起こすことから、ARDS の病態モデルや病態解明に利用されている。オレイン酸肺傷害 では、ARDS と同様に好中球をはじめとする炎症性細胞、サイトカイン類、接着分子な どを介して、肺血管内皮細胞や肺胞上皮細胞が傷害を受ける。その際、好中球から産 生される活性酸素種(ROSs)が細胞障害に関わることが知られているが、ROSs の発生源 や産生経路についての詳細は不明である。
そこで本研究では、モルモットを用いたオレイン酸肺傷害モデルにおいて、血液ガ ス、肺血管透過性、および肺胞洗浄液(BALF)中の種々の生化学マーカーの変化を指 標として、オレイン酸による酸化ストレスを詳細に検討した。次に、ROSs の発生源お よび産生経路について検討を加えた。また、ROSs は肺血管内皮細胞や肺胞上皮細胞の adenosine triphosphate (ATP)含量を低下させることから、細胞内エネルギーの保 持・補給を目的として、オレイン酸肺傷害に及ぼす細胞膜透過性エネルギー供給体 phosphoenolpyruvate (PEP)の影響を検討した。さらに、気道系に対して抗炎症作用 を示す種々の薬効成分を含む漢方処方、小青竜湯の肺傷害防止効果を検討した。以下 に本研究で得られた知見を要約する。
1. オレイン酸(15 µl/kg)をモルモットに静脈内投与すると、BALF 中の過酸化脂質量、
細胞障害の指標である乳酸脱水素酵素(LDH)活性、肺血管透過性の指標である総 蛋白量などの上昇、細胞内の酸化ストレスの指標である還元型および酸化型グルタ チオンの濃度比(GSH/GSSG 比)の減少などが観察された。Nicotinamide adenine dinucleotide phosphate (NADPH) オ キ シ ダ − ゼ 阻 害 剤 ( diphenyleneiodonium chloride (DPI), 600 µg/kg i.v.)の前処置は、オレイン酸投与による BALF 中の 酸化ストレス指標の変化および低酸素血症を有意に抑制したが、キサンチンオキシ ダ−ゼ阻害剤(allopurinol, 10 mg/kg i.v.)は抑制しなかった。これらの結果か ら、オレイン酸肺傷害において ROSs が重要な役割を果たしていること、および ROSs 産生に NADPH オキシダ−ゼが主として関与していることが示唆された。
2. モルモットにシクロホスファミド(150 mg/kg i.v.)を投与し低顆粒球症を誘発さ せた場合、オレイン酸投与による BALF 中の過酸化脂質量や LDH 活性の増加は有意
に抑制され、低酸素血症も抑制された。これらの結果は、好中球などの炎症性細胞 から産生される ROSs がオレイン酸肺傷害に重要な役割を果たすことを支持するも のである。
3. In vitro 細胞培養系において、肺胞Ⅱ型上皮細胞(A549)はオレイン酸に暴露され ると時間/用量依存的に ROSs を産生したが、その産生は DPI によって有意に抑制さ れた。しかし、オレイン酸による細胞死は DPI によっては抑制されなかった。また、
ウシ肺動脈血管内皮細胞(CPAE)においてもオレイン酸の刺激により ROSs を産生し たが、DPI によって抑制された。これらの結果はオレイン酸肺傷害における ROSs の 発生源として、好中球以外にも肺血管内皮細胞や肺胞上皮細胞が関与することを示 唆している。また、本実験条件下におけるオレイン酸による細胞死の要因として、
ROSs の関与は少なく、オレイン酸の細胞膜に対する直接作用などが推測される。
4. オレイン酸の静脈内投与は、肺組織中 ATP 含量を減少させた。PEP(2, 20, 200 µmol/kg, i.v.)の前処置はオレイン酸投与による低酸素血症を用量依存的に抑制 し、肺血管透過性亢進を抑制した。一方、細胞膜を透過できない ATP(4 µmol/kg, i.v.)を PEP と併用した場合、付加的な肺傷害軽減効果は観察されなかった。これ らの結果から、オレイン酸肺傷害時に肺細胞内エネルギー代謝系の障害が生じてお り、細胞膜透過性エネルギー供給体 PEP などを用いて肺細胞にエネルギーを供給・
補充することにより、急性肺細胞障害を軽減できる可能性が示唆された。
5. 気道系に対する抗炎症作用を示す種々の薬効成分からなる漢方処方、小青竜湯に よるオレイン酸肺傷害の防止効果について検討した。急性実験(オレイン酸(15 µl/kg)投与 3 時間前に小青竜湯(3 g/kg)を単回経口投与)では、小青竜湯はオ レイン酸による低酸素血症を有意に抑制したが、肺血管透過性亢進は抑制できなか った。一方、慢性実験(小青竜湯を 0.75 g,1 日 2 回,2 週間経口投与後、オレイ ン酸(15 µl/kg)投与)では、小青竜湯はオレイン酸による低酸素血症、肺血管透 過性亢進、および BALF 中の酸化ストレス指標の上昇を顕著に抑制した。また、慢 性実験において、小青竜湯はオレイン酸によるトロンボキサン A2の産生促進を抑制 した。
以上の結果から、オレイン酸誘発肺傷害に好中球をはじめ、肺血管内皮細胞や肺胞 上皮細胞から産生される ROSs が深く関与すること、その主な産生経路は NADPH オキ シダ−ゼ依存性であることが明らかとなった。また、肺傷害時に ROSs により細胞内 エネルギー代謝系が障害を受けることから、肺細胞へのエネルギー供給・補充は肺傷 害軽減の一方策になる可能性が示唆された。最後に、小青竜湯がオレイン酸肺傷害を 軽減する基礎的知見は、複雑な 炎症 である ARDS の予防・治療薬の開発において、
多成分系の漢方処方が新たな医薬品資源になりうる可能性を拓くものと考えられる。
Contents
Chapter 1. General introduction
1.1. Acute respiratory distress syndrome (ARDS) and oleic acid-induced lung injury…….1
1.2. Reactive oxygen species in ARDS and oleic acid-induced lung injury………..3
1.3. Endothelial and epithelial injury in ARDS and oleic acid-induced lung injury……….5
1.4. Energy changes in acute lung injury………6
1.5. Objective of this study……….7
Chapter 2. Oxidative stress in early stage of acute lung injury induced with oleic acid….8 2.1. Introduction………...8
2.2. Materials and methods………9
2.2.1. Animal model of lung injury………9
2.2.2. Drugs and chemicals………..10
2.2.3. Experimental procedure………10
2.2.3.1. Oleic acid-induced changes in biomarker in BALF……….10
2.2.3.2. Effects of DPI and allopurinol on partial pressure of oxygen in the artery (PaO2)………...11
2.2.3.3. Determination of biomarkers in BALF………..12
2.2.4. Statistical analysis……….13
2.3 Results………13
2.3.1. Time-dependent effect of oleic acid on lipid peroxidation in BALF……….13
2.3.2. Dose-dependent effect of oleic acid on biomarkers in BALF……….13
2.3.3. Effects of DPI and allopurinol on the lung oxidative changes induced with oleic acid………...15
2.3.4. Effects of DPI and allopurinol on PaO2 in the oleic acid-induced lung injury…….19
2.4. Discussion………20
2.5. Summary……….23
Chapter 3. Contribution of neutrophils on oxidative changes in the acute lung injury induced with oleic acid……….24
3.1. Introduction……….24
3.2. Materials and methods……….25
3.2.1. Materials………..25
3.2.2. Experimental procedure………...26
3.2.2.1. Depletion of granulocyte (PMNs)……….26
3.2.2.2. Blood gas measurement……….26
3.2.2.3. Determination of biomarkers in the BALF………27
3.2.3. Statistical analysis………. 27
3.3. Results………..27
3.3.1. Effect of leukocytopenia on lung oxidative changes induced with oleic acid………27
3.3.2. Effect of leukocytopenia on PaO2 in oleic acid-induced lung injury………27
3.4. Discussion………30
3.5. Summary……….….31
Chapter 4. Possible roles of pulmonary endothelial and epithelial cells in oleic acid-induced lung injury……….32
4.1. Introduction……….…………32
4.2. Materials and Methods……….33
4.2.1 Materials………33
4.2.2. Cell Culture………..33
4.2.3. Measurement of superoxide production in the CPAE or A549 cells induced with oleic acid………33
4.2.4. Cytotoxicity Assays……….…34
4.2.5. Statistical analysis……….35
4.3. Results……….……….35
4.3.1. Superoxide production in CPAE and A549 cells induced with oleic acid………35
4.3.3. Cytotoxicity of oleic acid………36
4.4. Discussion………40
4.5. Summary……….41
Chapter 5. Energy changes in oleic acid-induced lung injury and preventive effect of phosphoenolpyruvate on the lung injury………42
5.1. Introduction………42
5.2. Materials and methods……….43
5.2.1. Materials………..43
5.2.2. Blood gas parameters………43
5.2.3. Pulmonary vascular permeability………..43
5.2.4. Determination of lung ATP content………...44
5.2.5. Statistical analysis……….………45
5.3. Results………..………45
5.3.1. Effects on blood gas parameters………..45
5.3.2. Effects on pulmonary vascular permeability………50
5.4. Discussion………51
5.5. Summary………..54
Chapter 6. Preventive effect of Shoseiryu-to on oleic acid-induced acute lung injury….56
6.1. Introduction……….56
6.2. Materials and methods……….57
6.2.1. Drugs and chemicals………..57
6.2.2. Experimental procedure………58
6.2.3. Measurement of blood gases………59
6.2.4. Determination of vascular permeability………...60
6.2.5. Assessment of oxidant changes in BALF………..60
6.2.6. Determination of TXB2 in the BALF………..60
6.2.7. Statistical analysis……….60
6.3. Results……….……….61
6.3.1. Blood gases………..….…61
6.3.2. Vascular permeability………64
6.3.3. Oxidant changes in BALF……….64
6.3.4. TXB2 level in BALF………64
6.4. Discussion………67
6.5. Summary………..…71
SUMMARY……….72
REFERENCES……….….76
Chapter 1
General introduction
1.1. Acute respiratory distress syndrome (ARDS) and oleic acid-induced acute lung injury The acute respiratory distress syndrome (ARDS) is a complicated syndrome of acute lung inflammation that is characterized by a sequestration of neutrophils in the pulmonary microcirculation and an increase in microvascular permeability (Fig. 1‑1), 1-3) resulting from a variety of predisposing conditions including severe pneumonia, sepsis, massive transfusion and trauma.2) According to the criteria by American-European Consensus Conference,2) ARDS has been defined as severe form of acute lung injury (ALI) (table 1‑1). Mortality rate of ARDS is still high (more than 30%) despite recent advances in intensive care.4) The death often results from impaired gaseous exchange (hypoxemia) and subsequent alveolar fibrosis.5, 6) However, there are only a few drugs available for the prevention and treatment of severe hypoxemia caused by ARDS.
Table 1‑1. Recommended criteria for ALI and ARDS 2)
Timing Oxygenation Chest Radiograph
Plumonary Artery Wedge Pressure
ALI criteria
Acute onset
PaO2/FiO2 ≤ 300 mm Hg (regardless of PEEP level)
Bilateral infiltrates seen on frontal chest radiograph
≤ 18 mm Hg when measured or no clinical evidence of left atrial hypertension ARDS
criteria
Acute onset
PaO2/FiO2 ≤ 200 mm Hg (regardless of PEEP level)
Bilateral infiltrates seen on fronta chest radiograph
≤ 18 mm Hg when measured or no clinical evidence of left atrial hypertension PaO2: partial pressure of oxygen in artery, FiO2: fraction of inspired oxygen,
PEEP: positive end-expiratory pressure.
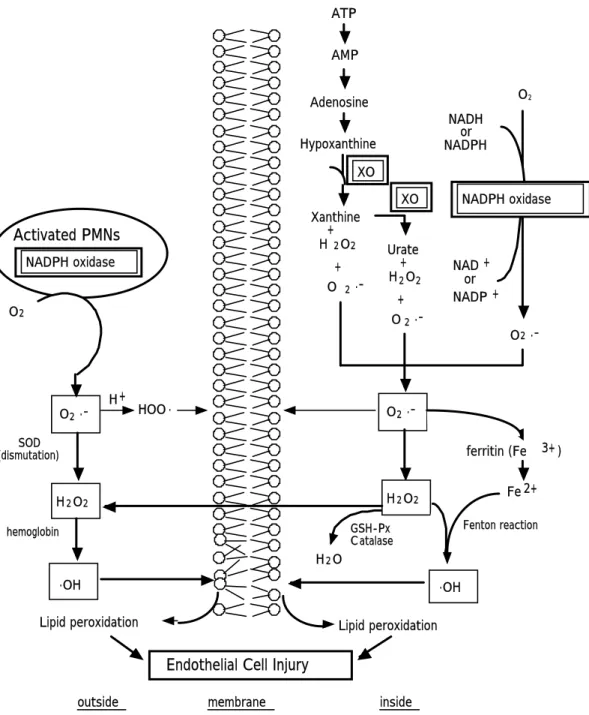
Fig. 1-1. Pathogenesis in the acute lung injury and ARDS
Oleic acid-induced lung injury is a well-characterized animal model of ARDS.7) The lung injury shows many pathophysiological changes that include severe hypoxemia, increased permeability of the alveolar-capillary membrane,8) thrombogenesis,9) hemorrhagic lung edema, decreased lung compliance and increase in pulmonary artery wedge pressure.7, 10) Although the precise pathogenesis of the lung injury is not clear, diverse inflammatory mediators such as thromboxane A2 and prostacyclin,9) phospolipase A211) and oxygen free radicals12) may contribute to the lung injury. These pictures of oleic acid-induced lung injury resemble those in ARDS.1, 3, 12-16)
1.2. Reactive oxygen species in ARDS and oleic acid-induced lung injury
Reactive oxygen species (ROSs) play an important role in ARDS16, 17) as well as several animal models of acute lung injuries.18-22) Free radicals produced by stimulated neutrophils may be one of mediators in pulmonary injury.23, 24) Aggregates of activated leukocytes in the pulmonary microvasculature release oxygen free radicals that are able to attack the polyunsaturated fatty acid chain of membrane lipids, thus initiating a peroxidation process.1) Peroxidation leads to loss of the functional integrity of the cell membranes, culminating in acute increase of alveolar-capillary permeability.
ROSs production systems from activated polymorphonuclear leukocytes (PMNs) and endothelial cells are considered to be three major pathways in acute lung injuries25) (Fig. 1‑2). Sources for ROSs in the lungs are 1) reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase including the phagocytic cell oxidase and nonphagocyte oxidase in the vascular endothelial cells, 2) xanthine dehydrogenase/xanthine oxidase (XDH/XO) and 3) mitochondrial respiration.25) Recently, several papers reported that NADPH oxidase-like enzymes and XO in the lung microvascular endothelial cells were important sources of ROSs generation for lung injuries,26‑29) and that the ROSs-generating pathways could vary from a model to a model.29, 30) For example, endothelial NADPH oxidase is known to participate in the ROSs-generating pathway in ischemia/reperfusion lung injury,27) while xanthine oxidase is involved in the ROSs-generating pathway in anoxia-reoxygenation.29) In addition, van Klaveren et al. reported NADPH oxidase-like enzyme involved in superoxide and
GSH-Px C atalase H2O
O2
.OH .OH
NADH or NADPH
NAD + or NADP +
O2 .-
O2 .-
H2O2
O2 .-
H2O2
H + HOO .
ferritin (Fe 3+ ) Fe 2+
Lipid peroxidation Lipid peroxidation
Endothelial Cell Injury
SOD (dismutation)
hemoglobin Fenton reaction
O2
Xanthine + H 2O2
+ O 2 .-
Urate + H2O2
+ O2 .- ATP
AMP
Adenosine
Hypoxanthine
NADPH oxidase
Activated PMNs NADPH oxidase
XO XO
outside membrane inside
Fig. 1‑2. The proposed mechanism for the generation of lipid peroxidation in the lungs. XO: xanthine oxidase, GSH-Px: glutathione peroxidase.
hydrogen peroxide generation by rat type II cells.
An intravenous administration of oleic acid can cause neutrophil activation.12) Activated neutrophils release superoxides,12) resulting in lung injuries. There are some papers demonstrating that oxidative stresses may contribute to oleic acid-induced lung injury,31 however, little is known the ROSs-generating system in oleic acid-induced lung injury.
1.3. Endothelial and epithelial injury in ARDS and oleic acid-induced lung injury
Two separate barriers form the alveolar-capillary barrier, the microvascular endothelium and the alveolar epithelium (Fig. 1‑1). The acute phase ARDS is characterized by the influx of protein-rich edema fluid into the alveolar spaces as a consequence of increased permeability of the alveolar-capillary barrier.32)
The pulmonary vascular endothelium is particularly susceptible to injury mediated by ROSs derived from circulating inflammatory cells, or produced by pulmonary endothelial cells.33) In the ARDS, an acute inflammatory response is invariably present, in which ROSs are derived from neutrophils, macrophages, and the endothelium.34) All of these cell types have been shown to generate increased amounts of superoxide anion in conditions of oxidative stress, leading to the acute lung injury as well as ARDS.16, 17) Proteolytic enzymes secreted by the neutrophils may also be implicated in injury of the lung microvasculature.35)
The loss of epithelial integrity in acute lung injury and the ARDS has a number of consequences. The loss of epithelial integrity and injury to type II cells disrupt normal epithelial fluid transport, impairing the removal of edema fluid from the alveolar space.36) In addition, injury to type II cells also reduces the production and turnover of surfactant, contributing to the characteristic surfactant abnormalities.37) Abnormalities in the production, composition and function of surfactants probably contribute to alveolar collapse and gas-exchange abnormalities.37) Intravenous administration of oleic acid causes the injury of alveolar epithelial cells,38) leading to abnormal balance of the lung fluid transport and surfactant abnormalities.11, 39)
Although ROSs released from activated neutrophils and other types of cells are
known to cause injury of alveolar type II cells and pulmonary endothelial cells, the precise sequence of the intracellular ROSs remains unclear.
1.4. Energy changes in acute lung injury
H2O2 has been known to be toxic to both alveolar type II cell and endothelial cell in vitro, causing decrease in ATP contents in the cells.40, 41) Oxidant-induced injury of pulmonary microvascular endothelial cells causes a rise in monolayer permeability, which best correlates with a fall in ATP level,42) and the mechanism would be that decreased level of ATP blunted the barrier function of the tight junctions.43) In addition, as ATP is a necessary substrate for cAMP-dependent protein kinase in the alveolar type II cells, reduced cellular ATP content in the alveolar type II cells would presumably favor the dephosphorylated state and thus decrease microsomal cytidyltransferase activity, which may lead to diminished alveolar surfactant.44)
The extent of edema in acute lung injury depends on the balance between restoration of the tight junctions and maintenance of the fluid clearance mechanism. A recent study 36) has shown that the active transport of sodium by the type II cell provides the major driving force for the secondary removal of alveolar fluid. Sodium uptake occurs on the apical surface, partly through amiloride-sensitive and amiloride-insensitive channels. Subsequently, sodium is pumped actively from the basolateral surface into the lung interstitium by Na‑K‑ATPase, and the action of the enzyme needs energy that fueled by hydrolysis of ATP. Therefore, depletion of ATP content in the alveolar type II cell may decrease the enzyme activity, leading to abnormal balance of lung fluid.
Schmeling and co-workers45) demonstrated that acute lung injury after intestinal ischemia-reperfusion caused ATP depletion in the lung tissues, neutrophil sequestration in alveolar capillaries, and endothelial cell injury with increased microvascular permeability, and suggested that activated neutrophils appeared to be involved.
Neutrophils activated by oleic acid can release ROSs such as superoxides, which would decrease ATP level in the lung tissues. The decrease in cellular ATP level would increase pulmonary vascular permeability through blunted barrier function of the tight junctions, eventually leading to hypoxemia.
1.5. Objective of this study
Moriuchi et al. have developed a model of the decrease in PaO2 induced by a low dose (15 µl/kg) of oleic acid for the screening of a drug for hypoxemia with pulmonary vascular permeability.46-48) However, the precise mechanism of the lung injury has not been elucidated.
In recent years, it is reported that ROSs play an important role in the oleic acid-induced lung injury,7) especially neutrophil-mediated superoxides.12) However, detailed investigations have not been carried out on sources for ROS other than neutrophil such as pulmonary endothelial cells and alveolar epithelial cells, and changes in antioxidant systems in the lungs. Therefore, the objectives of this study are to examine 1) oxidative changes in the lung injury induced by oleic acid using bronchoalveolar fluid (BALF) biomarkers, and assess whether or not NADPH oxidase and/or xanthine oxidase contribute to ROSs generation in the lung injury (Chapter 2), 2) effect of PMNs depletion on the oxidative changes and hypoxemia in the lung injury (Chapter 3), 3) whether or not pulmonary artery endothelial cells and/or alveolar epithelial cells generate ROSs by oleic acid, whether or not a possible resource of ROSs released from the cells is an NADPH oxidase-like enzyme, and whether or not the intra-cellular ROSs cause cell death (Chapter 4), 4) changes in ATP content in the lungs injured by the injection of oleic acid, and whether phosphoenolpyruvate, an intermediate substance of glycolysis which can carry an energy into cells, attenuates hypoxemia and pulmonary vascular hyper-permeability induced with oleic acid (Chapter 5), and 5) effect of Shoseiryu-to, a herbal medicine which can affect many inflammatory mediators, on the oleic acid-induced lung injury (Chapter 6).
Chapter 2
Oxidative stress in early stage of acute lung injury induced with oleic acid
2.1. Introduction
Reactive oxygen species (ROSs) play an important role in several models of acute lung injuries induced by ischemia/reperfusion, endotoxin, paraquat, oleic acid and phosgene.18‑22) ROSs production systems from activated polymorphonuclear leukocytes (PMNs) and endothelial cells are considered to be two major pathways in acute lung injuries.25) Sources for ROSs in the lungs are 1) reduced nicotinamide adenine dinucleotide phosphate (NADPH) oxidase including the phagocytic cell oxidase and nonphagocyte oxidase in the vascular endothelial cells, 2) xanthine dehydrogenase/xanthine oxidase (XDH/XO) and 3) mitochondrial respiration.25) Recently, several papers reported that NADPH oxidase-like enzymes and XO in lung microvascular endothelial cells are important sources of ROSs generation for lung injuries,26‑29) and the ROSs-generating pathways can vary from a model to another.29, 30) For example, endothelial NADPH oxidase is known to participate in the ROSs-generating pathway in ischemia/reperfusion lung injury,27) while xanthine oxidase is involved in the ROSs-generating pathway in anoxia-reoxygenation.29)
ROSs generated from cells are scavenged by antioxidant defense systems, including superoxide dismutase (SOD), catalase, glutathione peroxidase, and glutathione reductase, glutathione (GSH), and vitamins A, C, and E. When production of ROSs in tissues exceeds the ability of the antioxidant systems to eliminate them, oxidative stress occurs, which leads to an accumulation of lipid peroxidation products (e.g., malondialdehyde (MDA)), an increase in oxidized glutathione (GSSG) and a decrease in the ratio of GSH/GSSG.30, 49)
Oleic acid-induced lung injury is well known as a model of acute respiratory distress syndrome (ARDS).7) An intravenous administration of oleic acid can produce neutrophil activation12, 50) and activated neutrophils can injure the capillary endothelium
by releasing ROSs12, 51). The precise mechanism of oleic acid-induced lung injury is not elucidated, but one of possible mechanisms for the lung injury is that superoxide anion produced by oleic acid is converted to active oxidant-hydroxyl radical, and the radical may cause direct tissue injuries by lipid peroxidation.31, 52,53) Lipid peroxidation may lead to loss of the functional integrity of the cell membranes, culminating in acute increase in alveolar-capillary permeability. There are some evidences demonstrating that lipid peroxidation may contribute to oleic acid-induced lung injury. 53, 54) Participation of NADPH oxidase, XDH/XO and effects of antioxidant defense system have been reported in other animal models of acute lung injuries,18, 22, 27, 29, 55, 56) but only a few papers demonstrated an involvement of the system in oleic acid-induced lung injury.56) In addition, ROSs-generating pathway in oleic acid-induced lung injury is not elucidated well.
The increase in concentrations of biomarkers in bronchoalveolar lavage fluid (BALF) has been proposed to be one of the sensitive indicators of lung injuries.57, 58) Thus, this chapter deals with oxidative changes in the acute lung injury induced by oleic acid using BALF biomarkers such as thiobarbituric acid reactive substances (TBARS), lactate dehydrogenase (LDH) activity, total proteins, GSH, SOD, and catalase. Furthermore, it is assessed whether or not NADPH oxidase and/or xanthine oxidase contributed to ROSs generation in oleic acid-induced lung injury using diphenyleneiodonium chloride (DPI), a NADPH oxidase inhibitor and allopurinol, a xanthine oxidase inhibitor.
2.2. Materials and methods
This study protocol was approved by the Animal Care and Use Committee of the Kumamoto University. The care and handling of the animals were performed in accordance with National Institutes of Health guidelines of the care and handling of animals.
2.2.1. Animal model of lung injury
To induce acute lung injury, we intravenously injected 15 µl/kg oleic acid without any vehicle using a micro-syringe. Some saline was used to push out oleic acid from the
catheter into the subclavian vein. In our previous study, we used a ventilator to maintain constant ventilation, and 15 µl/kg oleic acid showed similar pattern with those of the spontaneous breathing animals.48) So we used spontaneous breathing guinea pigs in the present study to keep their airways intact.
2.2.2. Drugs and chemicals
Oleic acid was purchased from ICN Biomedicals Inc. (Aurora, Ohio, USA). DPI and allopurinol were purchased from SIGMA Chemical Co. (St. Louis, MO, USA).
2.2.3. Experimental procedure
Hartley strain guinea pigs of either sex were obtained from KBT-Oriental (Tosu, Saga, Japan), and fasted for 24 h before starting experiments. Fifteen µg/kg oleic acid was used in all studies except the experiment to examine dose-dependency of the effect of oleic acid.
2.2.3.1. Oleic acid-induced changes in biomarker in BALF
Animals were randomly assigned to each study groups. Twenty (642 ± 29 g, one saline group and four oleic acid groups) and 40 guinea pigs (491 ± 33 g, one saline group and three oleic acid groups) were used in time-and dose-dependent study of oleic acid on oxidative changes in the markers in BLAF, respectively. Forty guinea pigs (498 ± 38 g, saline, oleic acid, DPI + oleic acid, and allopurinol + oleic acid groups) were used in effects of DPI and allopurinol on oxidative stress induced with oleic acid.
The animals were anesthetized with pentobarbital sodium (25 mg/kg i.p.), and an operation was performed under local anesthesia with 2% procaine to kill pain further and diminish reflective movement to pain. A catheter (1.1 mm outer diameter) was inserted into a unilateral subclavian artery for blood sampling, and the other catheter was inserted into the other unilateral subclavian vein for injection of saline, DPI, allopurinol and oleic acid. Six hundreds µg/kg DPI or 10 mg/kg allopurinol were injected 30 min before the oleic acid injection in the test groups. Oleic acid was injected intravenously without any vehicle using a micro-syringe. A small amount of saline was used to push out
oleic acid from the catheter into the vein. To perform the bronchoalveolar lavage, further administration of pentobarbital sodium (50 mg/kg i.p.) was done 15 min before the bronchoalveolar lavage. Then whole lungs were lavaged twice with 10 ml iced cold saline (the recovery was routinely 90% or greater), and the two BALFs were combined and centrifuged at 400 g for 10 min at 4℃. The aliquots of the supernatant were frozen at -85℃ for the later assay. In the time-dependent study, the animals were underwent bronchoaveolar lavage 0.5, 1, 1.5 or 2 h after the injection of oleic acid, respectively. In the other studies, bronchoaveolar lavage was done 1.5 h after the oleic acid injection.
2.2.3.2. Effects of DPI and allopurinol on partial pressure of oxygen in the artery (PaO2) Thirty guinea pigs (502 ± 44 g) were randomly divided into three groups: the saline + oleic acid, DPI + oleic acid and allopurinol + oleic acid groups. Animals were pre-treated with saline, DPI or allopurinol 30 min before the oleic acid injection, and oleic acid was injected, then blood gases were measured. Partial pressure of oxygen (PaO2), partial pressure of carbon dioxide (PaCO2) and pH in the arterial blood and pH were measured 5 min before the oleic acid injection which was regarded as the value at 0 min, and these parameters were also measured 6, 10, 15, 35, 55 and 75 min after the injection (Fig. 2‑1). To normalize PaO2, the value at time 0 was considered to be 100% and PaO2 at the other times were calculated as follows:
PaO2 (mm/Hg) at a time × 100 PaO2 (%) at a time =
PaO2 (mm/Hg) at time 0
For the measurements of blood gas parameters, 150 µl of blood was collected and analyzed with a blood gas analyzer (ABL300‑Acid‑Base Laboratory, RADIOMETER CO, Tokyo, JAPAN).
Subclavian vein Subclavian artery
Blood sampling
-30 -15 -10 -5 55 75 90
Oleic acid injection
Drug injection Bronchoalveolar lavage
PROTOCOL
Blood sampling time (min) 15 35 10
6 0 Injection of oleic acid
Fig. 2‑1. Arrangement and protocol of blood gas (PaO2) measurements and broncholaveolar lavage.
2.2.3.3. Determination of biomarkers in BALF
The extent of lipid peroxidation was estimated as the concentration of TBARS using the lipid peroxidation test kit from Wako Pure Chemical Industries, LTD (Osaka, JAPAN). LDH and total proteins in BALF were analyzed using a bio-analyzer (Hitachi 7600, Tokyo, JAPAN). The levels of GSH and GSSG were determined using the GSH/GSSG assay kit (BIOXYTECH GSH/GSSG‑412) from OXIS INTERNATIONAL, INC. (Portland, OR, USA). The SOD and catalase activities were determined using the SOD assay kit (BIOXYTECH SOD‑525) from OXIS INTERNATIONAL, INC., and the catalase assay kit (Amplex Red Catalase Assay Kit) from MOLECULAR PROBES, INC.
(Eugene, USA), respectively.
2.2.4. Statistical analysis
Results were expressed as mean±SEM. In the assessment of oxidant changes among the values of the groups, Bartlett’s test was employed to examine the uniform variance of data. Then significant differences between groups were identified after the data were further analyzed by Dunnett's or Tukey's multiple comparison test. A p ≦ 0.05 was considered significant.
2.3 Results
2.3.1. Time-dependent effect of oleic acid on lipid peroxidation in BALF
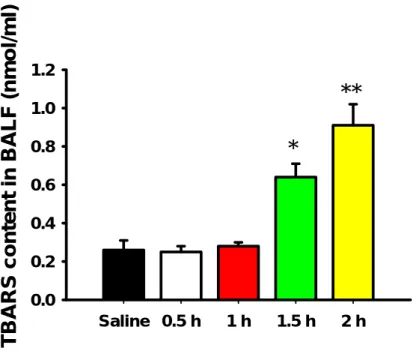
The level of TBARS in BALF was measured as an indicator of lipid peroxidation of the lung tissues. An intravenous injection of oleic acid at a dose of 15 µl/kg to guinea pigs time-dependently increased the level of TBARS in BALF (Fig. 2‑2). The TBARS levels in BALF 1.5 h and 2 h after the oleic acid injection significantly increased to 2.46 (p
< 0.05) and 3.50 (p < 0.01) times the value in the control group, respectively, but the TBARS level in plasma was not significantly different between the saline and oleic acid groups at 1.5 h (Fig. 2‑3).
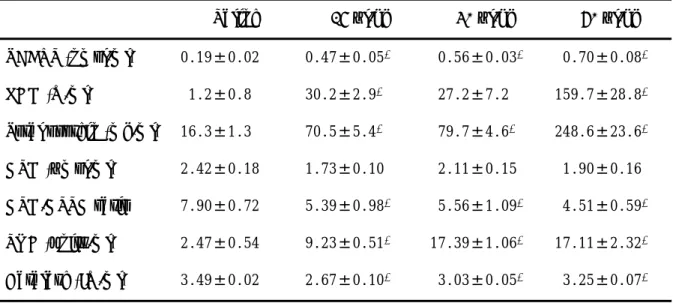
2.3.2. Dose-dependent effect of oleic acid on biomarkers in BALF
Table 2‑1 shows dose-dependent effects of oleic acid on biomarkers in the BALF 1.5 h after the oleic acid injection. The TBARS levels increased to 2.49 (p < 0.01), 2.96 (p <
0.01) and 6.22 (p < 0.01) times the value in the control group after the oleic acid injection at doses of 15, 30 and 60 µl/kg, respectively.
The LDH activity in the BALF served as a marker of alveolar cell injury, and the total protein concentration in the BALF was determined as a marker of alveolar-capillary membrane compromise. In the present study, LDH activity increased to 30.5 (p < 0.01), 22.7 and 133.1 (p < 0.01) times the level in the control group after 15, 30 and 60 µl/kg oleic acid injection, respectively, and total protein level increased to 4.33 (p < 0.05), 4.89
Fig. 2‑2. Time-dependent effects of 15 µl/kg oleic acid on lipid peroxidation in the BALF. *p<0.05, **p<0.01 compared with saline group; Each bar represents mean ± SEM (n = 4).
Saline 0.5 h 1 h 1.5 h 2 h
TBARS content in BALF (nmol/ml)
0.0 0.2 0.4 0.6 0.8 1.0
1.2
**
*
Saline Oleic acid
TBARS contents in plasma (nmol/ml)
0 1 2 3 4 5
Fig. 2‑3. Effects of 15 µl/kg oleic acid on lipid peroxidation in the plasma 1.5 h after oleic acid injection. Each bar represents mean ± SEM (n = 10).
There is not a significant difference (P = 0.12) between the saline and oleic acid groups.
Table 2‑1. Dose-dependent effect of oleic acid on bronchoalveolar lavage fluid biomarkers Saline 15 µl/kg 30 µl/kg 60 µl/kg TBARS (nmol/ml) 0.19±0.02 0.47±0.05* 00.56±0.03* 00.70±0.08* LDH (U/ml) 01.2±0.8 30.2±2.9* 027.2±7.2 159.7±28.8* Total protein (mg/ml) 16.3±1.3 70.5±5.4* 079.7±4.6* 248.6±23.6* GSH (µmol/ml) 2.42±0.18 1.73±0.10 02.11±0.15 01.90±0.16 GSH/GSSG ratio 7.90±0.72 5.39±0.98* 05.56±1.09* 04.51±0.59* SOD (units/ml) 2.47±0.54 9.23±0.51* 17.39±1.06* 17.11±2.32* Catalase (IU/ml) 3.49±0.02 2.67±0.10* 03.03±0.05* 03.25±0.07*
*p <0.05, compared with saline groups. All values are shown as the mean ± SEM (n = 10).
(p < 0.01) and 15.25 (p < 0.01) times the level in the control group after 15, 30 and 60 µl/kg oleic acid injection, respectively.
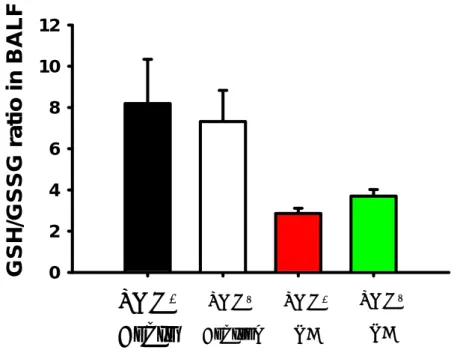
The intravenous injection of oleic acid decreased the GSH level and its ratio to GSSG in the BALF in a dose-dependent manner. The GSH level decreased to 86.7% (p
>0.05), 93.8% (p > 0.05) and 84.4% (p > 0.05) of the value in the control group after the oleic acid injection at doses of 15, 30 and 60 µl/kg, respectively. The GSH/GSSG ratio also decreased to 68.2% (p < 0.01), 70.4% (p < 0.01) and 57.1% (p < 0.01) of the value in the control group after the oleic acid injection at doses of 15, 30 and 60 µl/kg, respectively.
2.3.3 Effects of DPI and allopurinol on the lung oxidative changes induced with oleic acid The TBARS level in the BALF significantly (p < 0.01) increased in the 15 µl/kg oleic acid group compared with the control group. The increase in the TBARS level was significantly inhibited by DPI (p < 0.01), but was still higher (p < 0.01) than that in the control group (Fig. 2‑4). The LDH activity increased to 30.2 (p < 0.01) times that of the control group after the 15 µl/kg oleic acid injection and the increase was inhibited by DPI significantly (p < 0.01, Fig. 2‑5). Total proteins also increased in parallel with the increase in LDH, which was 5.08 (p < 0.01) times the level of the value in the control group after
the 15 µl/kg oleic acid injection, and the increase was also inhibited by DPI significantly (p < 0.05, Fig. 2‑6). However, allopurinol had no inhibitory effects on the increase in LDH and total proteins in the BALF (Fig. 2‑5 and 2‑6).
The decrease in GSH/GSSG ratio in the BALF induced with oleic acid was inhibited by the pretreatment of DPI significantly (Fig. 2‑7). The SOD activity significantly (p < 0.01) increased in the 15 µl/kg oleic acid group compared with that in the control group, and the rise in the SOD activity was significantly inhibited by DPI (p <
0.01), but is still higher (p < 0.05) than that in the control group (Fig. 2‑8). However, allopurinol had no inhibitory effects on such oxidant changes induced with oleic acid (Fig.
2‑7, 2‑8).
Saline OA DPI Allo
TBARS content in BALF (nmol/ml)
0.0 0.1 0.2 0.3 0.4 0.5 0.6
*
* #
*
Fig. 2‑4. Effects of DPI and allopurinol on lipid peroxidation production induced by 15 µl/kg oleic acid in BALF. *p<0.05, compared with saline group; #p
<0.05, compared with oleic acid group; Each bar represents mean ± SEM (n = 10). OA = oleic acid; Allo = allopurinol; DPI = diphenyleneiodonium chloride.
Saline OA DPI Allo
LDH activity in BALF (U/l) 0 10 20 30 40 50 60 70
#
*
Fig. 2‑5. Effects of DPI and allopurinol on LDH activity in BALF induced with 15 µl/kg oleic acid.*p<0.05, compared with saline group; #p<0.05, compared with oleic acid group; Each bar represents mean ± SEM (n
= 10). OA = oleic acid; Allo = allopurinol; DPI = diphenyleneiodonium chloride.
Saline OA DPI Allo
Total protein content in BALF (mg/dl)
0 20 40 60 80 100
#
*
Fig. 2‑6. Effects of DPI and allopurinol on total protein content by 15 µl/kg oleic acid in BALF. *p<0.05, compared with saline group; #p<0.05, compared with oleic acid group; Each bar represents mean ± SEM (n = 10). OA = oleic acid; Allo = allopurinol; DPI = diphenyleneiodonium chloride.
Saline OA DPI Allo
GSH/GSSG ratio
0 2 4 6 8 10 12
*
#
*
Fig. 2‑7. Effects of DPI and allopurinol on changes in GSH/GSSG ratio induced with 15 µl/kg oleic acid in BALF. *p<0.05, compared with saline group;
#p<0.05, compared with oleic acid group; Each bar represents mean ± SEM (n = 10). OA = oleic acid; Allo = allopurinol; DPI = diphenyleneiodonium chloride.
Saline OA DPI Allo
SOD activity in BALF (U/ml)
0 2 4 6 8 10
#
*
Fig. 2‑8. Effects of DPI and Allopurinol on changes in GSH/GSSG ratio induced with 15 µl/kg oleic acid in BALF. *p<0.05, compared with saline group;
#p<0.05, compared with oleic acid group; Each bar represents mean ± SEM (n = 10). OA = oleic acid; Allo = allopurinol; DPI = diphenyleneiodonium chloride.
2.3.4 Effects of DPI and allopurinol on PaO2 in the oleic acid-induced lung injury
Table 2‑2 shows the effects of DPI and allopurinol on PaO2 in the oleic acid-induced lung injury. The pretreatment of DPI significantly inhibited the decreases in PaO2 6, 10, 15 min after the oleic acid injection, while allopurinol had no inhibitory effects on them. On the other hand, in all groups, no significant changes in PaCO2 and pH were observed after the oleic acid injection, compared with the value at 0 min and between the control and test group.
Table 2‑2. Effects of DPI and allopurinol on changes in blood gas parameters in the artery by oleic acid (n = 10)
Time (min) 0 6 10 15 35 55 75
Saline + 15 µl/kg Oleic acid
PaO2 (mmHg) 103.2±3.2 053.3±3.5 048.2±3.6 052.1±4.0 069.8±3.3 080.2±2.7 088.6±3.0 PaCO2 (mmHg) 037.1±0.6 038.1±0.7 038.2±0.8 037.7±0.5 036.1±0.8 036.0±0.4 036.0±0.5 pH 7.381±0.011 7.372±0.011 7.371±0.011 7.382±0.010 7.382±0.012 7.382±0.013 7.389±0.010 600 µg/kg DPI + 15 µl/kg Oleic acid
PaO2 (mmHg) 102.3±4.3 062.9±4.4* 060.6±5.1* 065.2±6.0* 078.0±6.7 085.4±5.5 092.1±4.5 PaCO2 (mmHg) 035.8±0.5 037.2±0.7 037.2±0.8 036.9±1.0 035.2±0.6 035.5±0.5 035.6±0.2 pH 7.402±0.009 7.391±0.008 7.391±0.009 7.395±0.008 7.406±0.009 7.404±0.010 7.399±0.011 10mg/kg Allopurinol + 15 µl/kg Oleic acid
PaO2 (mmHg) 100.2±5.6 052.1±3.0 052.0±3.2 059.3±4.5 075.2±4.6 083.4±6.0 089.6±4.9 PaCO2 (mmHg) 036.1±0.8 037.2±1.0 037.1±1.0 036.0±1.0 035.4±0.9 035.3±1.0 035.2±0.7 pH 7.389±0.011 7.372±0.011 7.379±0.011 7.395±0.012 7.395±0.008 7.391±0.010 7.397±0.009
*p<0.05, compared with oleic acid group at each time; mean ± SEM (n = 10).
2.4. Discussion
We measured the changes in the levels of TBARS, LDH, total proteins, GSH, SOD and catalase in the BALF during the oleic acid-induced lung injury were measured to gain further insight into the mechanism(s) of the lung injury.
Since TBARS including MDA and other lipid peroxides are made from oxidized membrane lipids,59) the level of TBARS in the BALF can be used to assess the oxidative damage to the cell membranes in the lungs.57, 58) Simultaneously, the LDH activity as a marker of alveolar cell injury, and total proteins as a marker of alveolar-capillary membrane compromise were measured. The intravenous injection of oleic acid increased TBARS levels in the BALF in a dose- and time-dependent manner. In addition, the LDH activity and total proteins in the BALF increased with increasing doses of oleic acid.
These results indicate that the increased TBARS level in the BALF reflects enhanced oxidative damage to the cell membranes of alveolar cells and pulmonary endothelial cells, and suggest that the cell injury may at least partially contribute to oleic acid-induced pulmonary hyper-permeability. Zhao and coworkers31) had reported that TBARS level increased in the lung tissues after oleic acid injection would be an important mechanism of the oleic acid-induced lung injury. In contrast, Ward et al.53) had reported that the plasma level of conjugated dienes, used as another index of lipid peroxidation in the oleic acid-injected rats, was not significantly different from that in saline-injected animals.
According to they concluded that oleic acid-induced lung injury was an oxygen radical-independent lung injury. However, in the present study, although there was not significant difference in the plasma TBARS level between the saline- and oleic acid-groups, TBARS level in the BALF was different between them significantly. Taken together, oxidative indices of lipids in the BALF can reflect the changes in the lung tissues in oleic acid-induced acute lung injury, but those in the plasma cannot. Therefore, the conclusion derived by Ward et al. that oleic acid-induced lung injury is oxygen radical-independent lung injury should be reconsidered.
GSH is an intracellular thiol that presents ubiquitously in all tissues and in various body fluids, especially at high levels in the lung tissue and BALF.60, 61) When mammalian cells are exposed to increased oxidative stress, GSH is able to react with and
scavenge H2O2 to form GSSG,62) and then the GSH/GSSG ratio decreases. Therefore, GSH is potentially an important antioxidant, and measurement of the GSH level or determination of the GSH/GSSG ratio is a useful indicator of oxidative stress.49) The present data indicate that the GSH/GSSG ratio in the BALF significantly decreased in response to the oleic acid injection in a dose-dependent manner, demonstrating that the oleic acid injection may produce oxidative stress.
SOD catalyzes the dismutation of superoxide radicals into hydrogen peroxide and oxygen,63) and then the resulting hydrogen peroxide can lead to the production of oxidant-hydroxyl radical through the Fenton reaction, finally oxidant-hydroxyl radical can cause direct tissue injuries by lipid peroxidation.53) In the present study, SOD activity in the BALF was significantly increased by oleic acid dose-dependently. The result indicates that superoxide production is increased as the dose of oleic acid increases, and also indicates that contribution of SOD to oleic acid-induced lung injury may be harmful, because the increase in SOD activity can facilitate production of hydrogen peroxide and finally increases toxic oxidant-hydroxyl radical, which seems to cause increases in TBARS level (Table 2‑1). Therefore, the result further confirms that oxidative stress contribute to the oleic acid-induced lung injury.
In general, catalase activity increases in parallel to SOD activity in the lung injuries such as bleomycin- and ethanol-induced lung injuries.49, 58) In the present study, activities of the two enzymes in the lung tissues were not examined, but the catalase activity decreased and SOD activity increased in the BALF by the oleic acid injection.
One possible explanation is that there might have been overuse of catalase which led to the decrease in catalase activity, if there had been a difference in the relative amount between SOD and catalase in terms of scavenging activity. In addition to their activity in BALF, whether SOD and catalase in the lung tissue are affected by oleic acid or not is intriguing, so study has to be planned to figure out the precise mechanism further.
There are two major sources of the production of superoxide anions. One is NADPH-dependent oxidase, which is not only present in the plasma membrane of phagocytes and macrophages, but also present in pulmonary endothelial cells.28) The other is xanthine oxidase.
The dose of DPI, a NADPH oxidase inhibitor,64) and allopurinol a xanthine oxidase inhibitor65) to inhibit superoxide generation were chosen based on the following reports. Some of in vitro studies reported that DPI at concentrations ranging from 10 to 100 µM exerted the inhibitory effects on superoxide generation from pulmonary microvascular endothelial cells stimulated with paraquat or phorbol ester (PMA),20, 66) and the inhibition of NADPH oxidase activity in neutrophils was complete at DPI concentrations above 10 µM.65) A dose of 600 µg/kg DPI was chosen here because an intravenous administration of the dose would produce about 30 µM of maximum concentration in the blood based on the assumption that the blood volume of guinea pig is about 5.8% of body weight and DPI is only distributed to the blood at first. On the other hand, Hultkvist-Bengtsson and Martensson55) reported that pretreatment of 10 mg/kg allopurinol completely inhibited the elevation in uric acid concentration in the plasma after oleic acid infusion. The result suggests that the oxygen radical producing hypoxanthine/xanthine oxidase (XDH/XO) reaction contribute to oleic acid-induced lung injury, because XDH catalyzes the reactions of purine metabolism to produce uric acid, and XDH can be converted to XO, then XO uses molecular oxygen as its electron carrier producing superoxide. However, in their study they did not assess any oxidant changes.
Recent study using isolated perfused rat lung27) has shown that ROSs production is inhibited by allopurinol (100 µM) in a model of anoxia/reoxygenation lung injury.
Considering above observations, an intravenous dose of 10 mg/kg allopurinol was assumed to produce 1.25 mM of maximum blood concentration in guinea pigs, and the level could be enough to inhibit ROSs production by hypoxanthine/xanthine oxidase. Our results clearly demonstrated that DPI inhibited the oxidative changes in the BALF and the decrease in PaO2 induced with oleic acid, while allopurinol had no inhibitory effects.
Moriuchi et al. has shown that neutrophils activated by oleic acid release oxygen free radicals.12) However, NADPH oxidase from neutrophils is can be one of the most promising ROSs generating enzymes in the oleic acid-induced lung injury. A recent study has demonstrated that NADPH oxidase is responsible for the generation of superoxide in mouse lung microvascular endothelial cells stimulated with PMA, and the superoxide release is inhibited by DPI.66) These results indicate that NADPH oxidases, originated
from lung microvascular endothelial cells as well as from neutrophils, participate in the superoxide generation.
2.5 Summary
1. Oxidative stress would be an important pathogenetic mechanism of the oleic acid-induced lung injury.
2. NADPH oxidase-dependent pathway may contribute largely to the generation of ROSs in the oxidative stress induced with oleic acid.