九州大学学術情報リポジトリ
Kyushu University Institutional Repository
カイコ卵および後発生器官におけるアルドラーゼア イソザイムおよびその関連酵素の動態
長岡, 純治
九州大学農学研究科農学専攻
https://doi.org/10.11501/3106907
出版情報:Kyushu University, 1995, 博士(農学), 課程博士 バージョン:
権利関係:
Chapter IV
Distribution of some enzymes related to aldolase in post-embryonic organs
INTRODUCTION
The previous chapter showed that the S and F aldolase isozymes are expressed in an organ-specific manner in larval, pupal and adult organs. The author therefore attempted to resolve wheth
er the distribution of some other enzymes change in relation to aldolase. The principle of the study is the same as that described in INTRODUCTION for Chapter II. Hexokinase was omitted from the list of enzymes to be elucidated, since emphasis was laid on the fructose metabolism.
MATERIALS AND METHODS
Animals
Eggs of a commercial hybrid race, Gunpo x Shugyoku, obtained after oviposition, were chilled for a long period of time followed by the hot HCl-treated as described in Chapter III. The hatched larvae were reared on an artificial diet (see Chapter I) at 25°C. Larval, pupal and adult organs were dissected according to the methods as described in Chapter III.
Determination of enzyme activities
The extraction of organs and the determination of enzyme activities were done as described in Chapter II. For aldolase, PFK, SDH and iCDH, 10 mg organs were homogenized in 500 Jll ice cold PKM. For G6PDH, 5 mg organs were homogenized similarly.
The homogenates were centrifuged and the supernatants were used for enzyme assays. The protein content was determined by the method of Bradford (1976) using a Bio-Rad protein assay kit with bovine serum albumin as a standard.
RESULTS
Enzyme activities in larval, pupal and adult fat bodies and muscles The enzyme activities were assayed using extracts from the fat bodies and muscles (the latter included the epidermal cells and cuticles) at different stages during post-embryonic development.
The results normalized by the respective protein contents in the specimens used for the enzyme assays are listed in Table 10. The fat bodies contained high aldolase activity at the fifth larval instar, but only weak aldolase activity thereafter. This confirms the data shown in Chapter III. The muscle aldolase activity was high, although fluctuating among the three stages tested. PFK of the fat bodies and muscles were at similar activity levels in the larvae and pupae but gave high activities in the adults. The NAD-SDH and NADP-SDH activities were high in the larval fat bodies at the fifth instar and at the spinning stage, but gradually decreased until
72
-.J w
TABLE 10. Several enzyme activities in the fat bodies and muscles of larvae, pupae and adults.
ALD NAD- NADP-
PFK G6PDH
FBP F1P SDH
Fat body
5L4 64.0 +12.4 3.69 + 0.62 68 +28 44.6 + 34.0 3.59 + 0.44 57.5 + 7.7 PP2 2.3 + 0.8 0.14 + 0.01 69 +11 29.3 + 1.8 1.27 + 1.25 20.5 + 12.0 P4 0.9 + 0.2 0.07 + 0.05 77 + 3 2.5 + 1.1 0.47 + 0.43 12.8 + 8.5 AO 2.1 + 1.0 0.19 + 0.10 102 +42 1.2 + 0.2 0.16 + 0.00 13.2 + 5.4
Muscle*
5L4 49.8 + 10.3 1.56 + 0.03 48 + 0 1.8 + 1.0 0.46 + 0.14 17.0 + 5.3 P4 17.2 + 8.2 0.60 + 0.03 61 + 0 0.7 + 0.5 0.49 + 0.16 16.5 + 1.0 AO 27.1 +11.3 0.52 + 0.19 107 +35 0.5 + 0.3 0.21 + 0.15 10.1 + 7.4
*Containing cuticles and epidermal cells.
ALD denotes aldolase. For other abbreviations see Chapter II. Enzyme activity was expressed in mU/
mg protein of the enzyme solution. Each value represents the mean of five separate samples with ±S.E. SL4, PP2, P4 and AO denote day-4 larvae at the 5th instar, day-2 larvae after the onset of spinning, day-4 pupae and newly emerged adults, respectively. Adult muscles were from the thorax parts. Gunpo x Shugyoku was used.
TABLE 11. NAD-iCDH and NADP-iCDH activities in the fat bodies and muscles of larvae, pupae and adults.
NAD- NADP-
iCDH Fat body
5L4 0.78 +0.07 274 + 68
PP2 0.80 +0.00 162 + 102
P4 0.23 +0.04 90 + 74
AO 0.17 +0.10 89 + 41
Muscle*
5L4 2.66 +0.82 64 + 3
P4 3.00 +0.93 47 + 4
AO 5.66 +0.40 87 + 4
*Containing cuticles and epidermal cells.
Values are mU/mg protein mean +SEM for n = 5. See Table 10 for other comments.
74
emergence. The G6PDH activity was much higher in the larval fat bodies than in the pupal and adult fat bodies. The latter were at comparable levels to those in the muscles. The level of NAD-iCDH activity in the fat bodies was less than those of the muscles. NADP
iCDH was consistently high in activity in particular larval fat bodies (Table 11).
DISCUSSION
As shown in Chapter
III(Table 9), the PBP/FlP activity ratio of aldolase in the fat body was significantly lower than that of the muscle fractions (about IO to
20for the former whereas about
30to
50
for the latter). Thus fat body aldolase has a high affinity for PIP than muscle aldolase. The results in the present chapter indicated that the larval fat bodies were rich in most of the enzyme activities tested (the NAD-iCDH activity was low but detectable). The finding that NAD-SDH activity was present in the larval fat bodies is supported by the fact that mRNA for this enzyme was localized here (Niimi
et al.,I993). The author concludes that the metabolism of PIP and other carbohydrates is essentially active in the fat bodies.
Probably, the so-called polyol pathway involving PIP metabolism functions to avoid the undesirable accumulation of glucose in the fat body, which may be converted via sorbitol and fructose into PIP, and the latter in turn becomes a substrate of aldolase. In verte
brates, the polyol pathway is known to operate in those organs that
are insulin insensitive (e.g. the lens, peripheral nerves and renal
glomoruli), where the SDH activity rises adapting diabetic increase in the glucose concentration (Jeffery and Jomvall,
1988).
On the other hand, the muscles (plus epidermal cells and cuticles) gave a higher FBP/F1P activity ratio (about
30).
This fact, together with the poorer activities for both types of SDH than those detected in the larval fat bodies, suggests that the F1P utility might be much more limited in the muscles compared to the larval fat bodies. Instead, the marked PFK and NAD-iCDH activities in the muscle fractions may mirror energy metabolism of the muscles, in particular for the wing muscle in the adults. Increasing activities of iCDH at the period of adult muscle differentiation were alsoreported previously (Miake et al.,
1976).
The smaller activity of aldolase in the larval fat bodies at the spinning stage was in contrast to the large value at the feeding stage (sudden fall from
64
to 2.3, see Fig.14
and Table10).
However, the activity of PFK in the fat bodies did not change largely andremained higher than aldolase throughout the pupal-adult
development. In the silkworm fat bodies, aldolase may regulate the rate of glycolysis and may be responsible for the metabolic
switchover, related to the dramatic fall of trehalose synthase activity in the fat bodies during metamorphosis (Hirano and Yamashita,
1983).
Aldolase is also involved in the transition of carbon flow from carbohydrate to fat catabolism in the locust flight muscle (Storey,1980).
76
SUMMARY
The larval and adult muscles possessed highly FBP/FlP
activity ratio, suggesting that FlP utilization is less effective. This is agreement with the fact that the NADP-SDH and NAD-SDH
activities were low. The PFK, G6PDH, NAD-iCDH and NADP
iCDH activities were high, indicating that the overall carbohydrate metabolism is at an augmented state in the muscles.
Chapter V
Partial purification and characterization of aldolase isozymes
INTRODUCTION
Study of enzymatic properties using crude extracts indicated the occurrence of two aldolase isozymes in B. mori, one acting primarily on FBP and predominantly be distributed in neonates and the muscles, and the other not only acting on FBP but also FlP, and is distributed in younger eggs and the fat bodies. Some organs exhibited three additional bands between the S and F bands upon zymogram. These may be heterohybrids. To confirm the identity of isozymes and the formation of heterohybrids, purification and characterization of S and F isozymes are tried in this chapter. Also the author tried to detect the genomic aldolase sequence region of B. mori by Southern hybridization using an authentic class I aldolase probe, (a eDNA clone for human aldolase B).
MATERIALS AND METHODS
Chemicals
Phosphocellulose (P-cellulose) was purchased from Serva Feinbiocheica (Heidelberg, Germany). Sephadex G-200 Superfine (Pharmacia LKB Biotechnology, Uppsala, Sweden) was used for gel
78
filtration. Silver staining kit, cesium chloride (CsC12), polyvinyl
pyrolidone, sodium pyrophosphate, salmon sperm DNA, ethidium bromide and bovine serum albumin were obtained from Wako Pure Chemical Industries (Osaka, Japan). Size marker proteins (No. 17-
0445-01),
Ampholines and Ficoll400
were purchased from Pharmacia LKB Biotechnology (Uppsala, Sweden). Purified rabbit muscle aldolase A, proteinase K and biotilated dUTP were obtained from Boehringer (Mannheim, Germany). A DNA labeling kit was supplied from Takara (Osaka, Japan). A detection kit for biotilated deoxynucleotide was produced by Sumitomokinzoku (Osaka, Japan). GeneScreen membrane was obtained from NEN Research Product (Boston, USA).Animals
For eggs and neonates, Gunpo x Shugyoku (Chapter I) was used. Newly deposited eggs were kept at 25°C to allow entering diapause. On day 30, the eggs were chilled to 5°C and, on day 120 of chilling, these were transferred to 25°C to permit post-diapause development. For larval fat bodies and muscles, eggs deposited from a hybrid (N137 x C137) were chilled and HCl-treated and the hatched larvae were reared on an artificial diet at 25°C as described in Chapter I.
Partial purification of aldolases
Sand
FFor aldolase S, eggs or larval fat bodies were homogenized in 10 volumes of 100 mM Tris-HCl buffer, pH 8.0, containing 10 mM EDT A with a glass-glass homogenizer at
4
°C. The homogenate was centrifuged at 12,000g for 20 min at 4°C. The supernatant wasfractionated by salting out with ammonium sulfate. The precipitate at 30 to 80% saturation was dissolved in a small amount of TE (50 mM Tris-HCl buffer, pH 8.0, containing 10 mM EDTA) and dialyzed for 8 hr at 4 oc against TE. To remove in-soluble materials the solution was centrifuged at 12,000g for 15 min. The supernatant was applied to a column of P-cellulose equilibrated with TE. The column was washed with TE until the absorbancy at 280 nm of the effluent dropped below 0.03. Aldolase activity was then eluted from this column with 100 mM NaCl dissolved in TE.
For aldolase F, neonates or muscles were homogenized in 10 volumes of PP (50 mM potassium phosphate buffer, pH 6.0) and centrifuged at 12,000g for 15 min at 4 °C. The resulting super
natant was applied (without salting out) to a P-cellulose column equilibrated with PP. The column was washed with PP and then eluted with 100 mM NaCl dissolved in PP.
The absorbancy was monitored at 280 and 260 nm during elution and the protein concentration of each fraction was estimated by the formula 1.45 x absorbancy at 280 nm- 0.74 x absorbancy at 260 run.
Gel filtration and SDS-PAGEs
Gel filtration was performed using Sephadex G-200 Superfine equilibrated with PP containing 0.5 M NaCl. SDS-polyacrylamide gel electrophoresis (SDS-PAGE) was done in 10% acrylamide according to Laemmli (1970) at 150 V for 1 hr. Proteins were stained with silver staining kit. For the estimation of molecular weight, the above mentioned size marker proteins (No. 17-0445-01) were co-migrated.
80
Determination of optimum pH, optimum temperature and thermo
stability
Optimum temperature was determined at pH
7 .5,and opti
mum pH at 30°C by the spectrophotometric method. For thermo
stability test, one volume of enzyme preparation was added to two volumes of 100 mM Tris-HCl buffer containing 10 mM EDTA and allowed to stand at 30°C for various period of time. For all experi
ments, the activity was assayed with FBP as a substrate.
2D gel electrophoresis
Eggs and organs were homogenized in the modified O'Farrell sample buffer containing 2% (V /V) Ampholine (pH 3 to 10), 2o/o (WN) Nonidet P-40, 9.5 M urea and 5% (V/V) 2-mercaptoethanol.
The extract was centrifuged at 12,000g for 10 min at 10°C and aliquots of the supernatant were subjected to O'Farrell type two
dimensional (2D) gel electrophoresis (O'Farrell, 1975). The first dimensional run was done with isoelectric focusing. The gels were poured to a height of 10 em in glass tubes (2 mm inside diameter) with a gel mixture composed of 9.2 M urea, 2% (W/V) Nonidet P- 40, 4% (W/V) acrylamide/N,N'-methylenebisacrylamide and 2%
(WN) Ampholines (1.5% pH 5 to
7and 0.5% pH 3.5 to 10). For the
running buffer, 0.02 M NaOH and 0.01 M H3P04 were used in the
anode and cathode, respectively. After a prerun at 200 V for 30 min
then at 400 V for 30 min, the samples were loaded onto the gels
and electrophoresis was performed at 800 V for 16 hr followed by
1,200 V for 15 min at 10°C. At the end of the run, the gels were
equilibrated for 2 hr in an
SDSsample buffer containing 62.5 mM
Tris-HCl buffer, pH 6.8, 5%
(VN)
2-mercaptoethanol, 2.3%(WN)
SDS and 10%(W N)
glycerol at room temperature and stored at -80°C. The gels were loaded on top of the slab gels for the second dimension. The slab was composed of a separation gel with 12%(WN)
acrylamide/N,N'-methylenebisacrylamide in 125 mM TrisHCl buffer, pH 6.8, containing 0.1% SDS. Electrophoresis was run at a constant voltage of 150
V
in 25 mM Tris, 192 mM glycine and 0.1% SDS, pH 8.3. Purified rabbit muscle aldolase A (40,000 in molecular weight, see ChapterI)
as a molecular marker was run in parallel.Western blotting of gels
Proteins were separated by the 2D-gel electrophoresis and transferred to PVDF paper, which was reacted with the anti-aldolase S serum as detailed in Chapter I. In the present case, the anti -serum was diluted at 1 : 500 with Milk-PBS and the washing of membrane was done twice with PBS containing 0.02% T-20.
Preparation of genomic
DNAfor hybridization
DNA was prepared according to the method described previ
ously (Kai
et a!.,
1992). Diapausing eggs (5 g) of the silkworm (Gunpo x Shugyoku) were frozen in liquid nitrogen and ground to a fine powder with a mortar and pestle. The frozen powder was transferred to 50 ml bottle and thawed in 4 ml extraction buffer (50 mM Tris-HCl buffer, pH 8.0, 500 mM EDTA, 0.5% sarcosine and 800 Jlg proteinase K). The mixture was stirred gently to disperse tissues at 37°C for 6 hr. Then, added 3.4 g of solid CsC12 and 250 Jll ethidiumbromide (100 mg/ml) were added. The mixture was82
stirred gently until dissolved. The solution was transferred to an ultracentrifuge tube and centrifuged for 48 hr at 38,000 rpm at 20°C in a Hitachi RPV65T rotator. After the centrifugation, the DNA band was recovered by suction into a syringe through a needle.
The solution was mixed with an equal volume of isoamyl alcohol.
The tube was shaken vigorously, the upper phase was removed and the aqueous phase extracted repeatedly until no red color remained.
The solution was dialyzed against TE (10 mM Tris-HCl buffer, pH 7.5, 1 mM EDTA) and concentrated with 2-butyl alcohol. DNA was precipitated with ethanol, air dried, dissolved in TE and stocked at -20°C until use.
Probe for hybridization
A eDNA clone for the human aldolase B gene was supplied by Dr. Y. Sugimoto of Seinan-Jogakuin Junior College. The clone including the full coding sequence had been inserted into EcoRI
Hindiii site of the plasmid pHABL 120-3. In this experiment, the plasmid was digested with Mboii and the resulting fragments were separated by agarose gel electrophoresis. A 672 bp fragment containing the active site sequence was used as a probe. It was labeled with random primed deoxyribonucleotide probes generated using the DNA labeling kit and biotilated dUTP.
Southern hybridization analysis
About 20 J.lg of DNA was digested extensively with EcoRI and Hindiii and subjected to electrophoresis on 1% agarose gel made up in 50 mM Tris, 1 mM EDT A and 48 mM boric acid, pH 8 .2. After electrophoresis, the digests on agarose gel were dena-
tured in 0.2 N NaOH plus 600 mM NaCl for 30 min, neutralized with 240 mM Tris-HCl buffer, pH
7.7,
containing 600 mM NaCl for 1 hr and blotted onto a GeneScreen membrane filter by the method described previously (Southernet a/., 1975).
For the hybridization, the blotted membrane filter was incubated at 42°C for 30 hr in 20 ml of 2 x Denhardt's solution (0.04% bovine serum albumin, 0.04%polyvinylpyrolidone and 0.04% Ficoll), 5 x SSC (
7
50 mM NaCl and75
mM sodium citrate), 1% SDS, 100 mg/ml heat denatured salmon sperm DNA, 0.1% sodium pyrophosphate and an aliquot of labeled probe DNA. After incubation the filter was washed twice with 0.5 xSSC and 0.1% SDS for 30 min at 50°C. The filter was dried in air and exposed to an X-Omat AR film using an detection kit according to the manufactures instructions.
Other methods
Determination of enzyme activity by zymogram or spectropotometry and other procedures including those for kinetic parameters were done as described in Chapter I.
RESULTS
Establishment of procedure for partial purification
Previous procedure for purification of aldolase proteins have generally utilized P-cellulose to which proteins were absorbed and then eluted with FBP at low temperature (Penhoet
et al.,
1969;Kochman and Kwiatkowska, 1972). This method, successfully applied to mammalian aldolases, was found to cause a confusion in
84
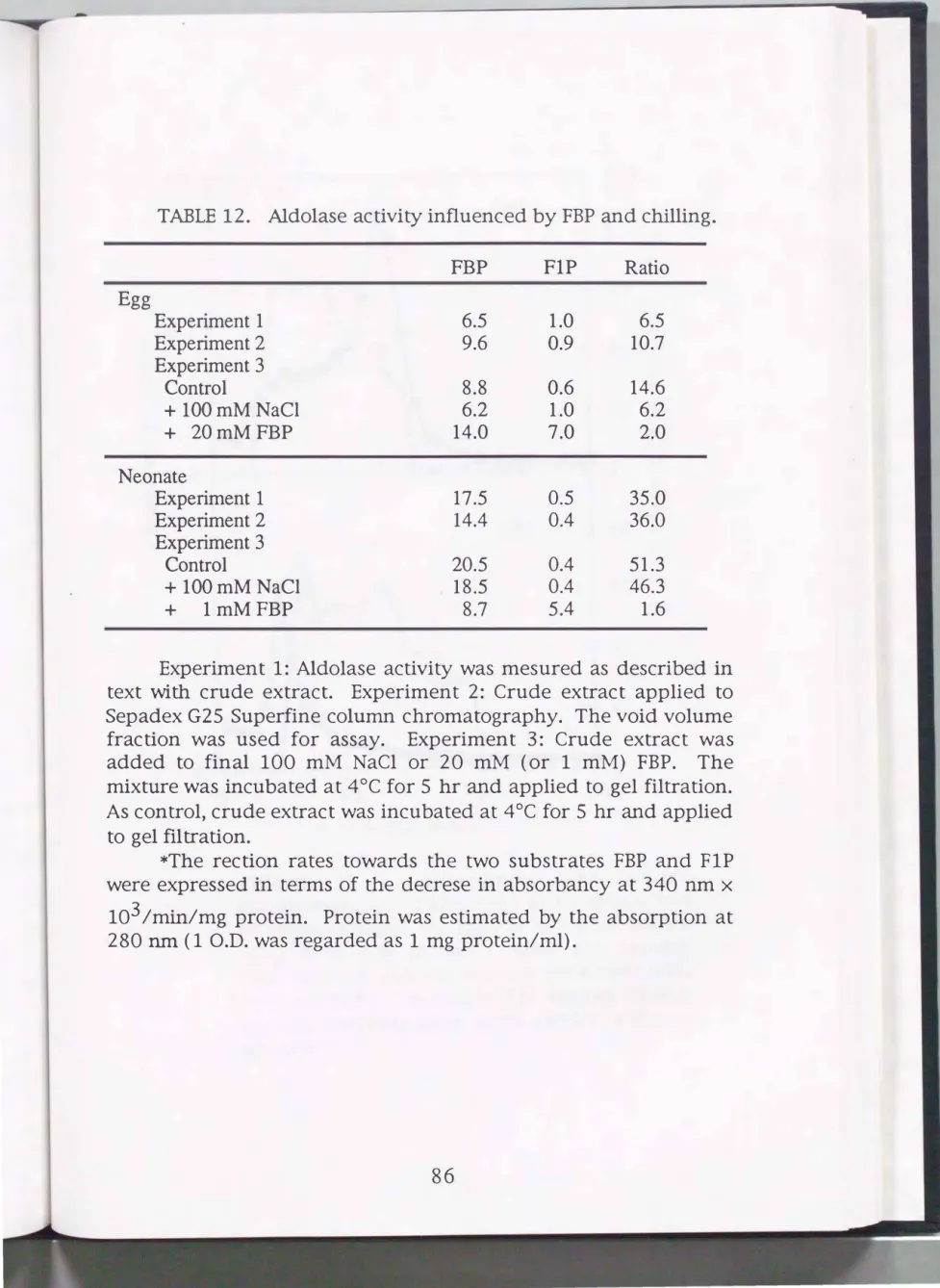
the case of the silkworm. The author inferred that the elution of aldolase with FBP from the P-cellulose column is inadequate. A test was made to see the influence of FBP at low temperature for the silkworm aldolase activity (Table 12). The crude extracts from eggs and neonates were incubated with FBP (final 20 mM for eggs and 1 mM for neonates) at 4°C for 5 hr. After incubation, the mixtures were passed through a column of Sephadex G-25 Superfine equilibrated with TE (this step aims at removing FBP). Because of unknown reason, the aldolase activities for PIP of the extracts after the FBP treatment were found to be activated. Moreover, the values of FBP/FlP activity ratio became low in particular for the neonates.
The values of 2 for S and 1.6 for F resembled the previously reported values of purified silkworm aldo�ase isozymes, 3 for S and 1 for F (Nishimura, 1992; Y ara, 1994 ). In contrast, the treatment with 100 mM NaCl did not affect the FBP/FlP activity ratio. Then the author avoided the use of FBP for elution at the chromatography step.
Partial purification of isozymes S and F
The type-S aldolase isozyme was partially purified both from eggs (day 1 after the start of embryogenesis) and larval fat bodies (day 3 of the 5th instar) by the procedures described in MATERIALS AND METHODS. The egg and larval enzymes were eluted at the same fractions in the P-cellulose chromatography (Fig.
15). The type-F enzyme was purified from neonates and larval muscles (day 3 of the 5th instar) by a procedure slightly different from that applied for S (see MATERIALS AND METHODS). Both enzymes again were eluted at the same fractions from the P-
TABLE 12. Aldolase activity influenced by FBP and chilling.
FBP FlP Rati o
Egg
Experiment 1 6.5 1.0 6.5
Experiment 2 9.6 0.9 10.7
Experiment3
Control 8.8 0.6 14.6
+ lOOmMNaCI 6.2 1.0 6.2
+ 20mMFBP 14.0 7.0 2.0
Neonate
Experiment! 17.5 0.5 35.0
Experiment2 14.4 0.4 36.0
Experiment3
Control 20.5 0.4 51.3
+ lOOmM NaCI 18.5 0.4 46.3
+ lmMFBP 8.7 5.4 1.6
Experiment 1: Aldolase activity was mesured as described in text with crude extract. Experiment 2: Crude extract applied to Sepadex G25 Superfine column chromatography. The void volume fraction was used for assay. Experiment 3: Crude extract was added to final 100 mM NaCl or 20 mM (or 1 mM) FBP. The mixture was incubated at 4°C for 5 hr and applied to gel filtration.
As control, crude extract was incubated at 4°C for 5 hr and applied to gel filtration.
*The rection rates towards the two substrates FBP and FlP were expressed in terms of the decrese in absorbancy at 340 nm x
103 /min/mg protein. Protein was estimated by the absorption at 280 nm ( 1 O.D. was regarded as 1 mg protein/ml).
86
0.2
E 0.1
0 c CX) N +-J ro
� u
c 0
..0 ro 0 (f) ..0 <{
0.1
0 0
100
(A)
8060
,..--...
... E
40 =>
""-/
E
20 � >.:>
� u
0 <(
(1) (J) ro
(B)
80 ""0 00 <(
yt-o
60\0
40booo
20
10 20 30
Fraction No.
FIGURE 15. Phosphocellulose column chromatography of aldolase S (after salting out of crude extracts). A, Eggs; B: larval fat bodies (day 3 of 5th ins tar). After the protein concentration fell, the column was eluted with 100 mM NaCl disolved_ in TE. Closed circles, protein concentration; open circles, aldolase activity.
cellulose column (Fig. 16). Each aldolase preparations after the P
cellulose column chromatography gave several bands on SDS
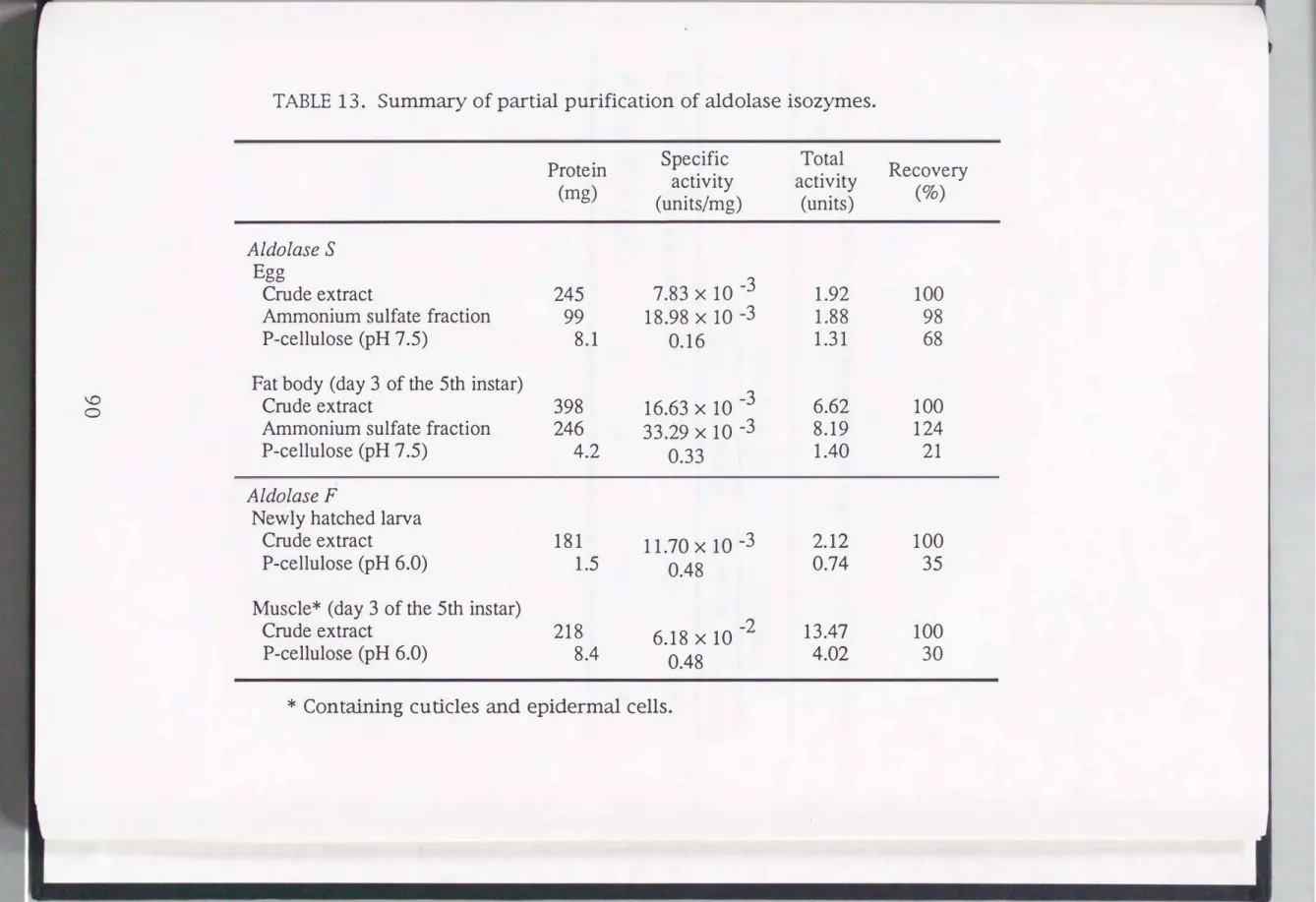
PAGE but a single activity band on zymogram analysis and a single protein peak on gel filtration. Because of this, taken together the fact that aldolase F is very unstable, further purification steps were not carried out. Table 12 summarizes the purification procedures.
The specific activities of the type S and F enzymes were over 8-fold that of the crude extract. The partially purified isozymes were analyzed for their enzymatic properties.
Molecular weight
For estimation of the molecular weight of S and F isozymes under non-denaturing conditions, the crude extract and partially purified aldolases were subjected to gel filtration. Aldolase activity was eluted as a single peak, whose molecular weight was calculated to be 160,000. SDS-PAGE analysis revealed that the partially puri
fied aldolases were at the molecular weight of 40,000.
Kinetic parameters
As listed in Table 14, aldolase S preparations from the two different sources gave comparable kinetic parameters to each other.
Similarly, different F preparations resembled each other. The Km values for FBP were 5 x 10-6 and 3 x 10-6 for type-S from eggs and fat bodies, respectively, and 7 X 10-6 and 5 X 10-6 for type-F from neonates and muscles, respectively. The apparently different Km values are actually within experimental errors. The FBP/F1P activity ratio was about 3 for S, and about 11-12 for F.
88
0.1
0.08
0.06
0.04 E
0 c 0.02
(X) N
� ro 0
� u c ro
..0 0.2
� 0
..0 (f)
<( 0.15
0.1
0.05
0 0
100
(A)
8060
40 ,..-..
E
...
20 :::> E
'-..-/
� �
0 .>
(B)
8060
40
20
0
10 20 30
Fraction No.
FIGURE 16. Phosphocellulose column chromatography of aldolase F (without salting out of crude extracts). A, neonates; B, larval muscles containing epidermal cells and cuticles (day 3 of 5 th ins tar). After t he protein concentration fell, t he column was eluted with 100 -mM NaCl disolved in
� u ro
<1>
(f) ro -o 0
<(
PP. Closed cir c les, pro tein concentration; open circles, aldolase activity.
\0 0
TABLE 13. Summary of partial purification of aldolase isozymes.
Protein Specific Total activity activity (mg)
(units/mg) (units)
Aldolase S Egg
7.83 X 10 -3
Crude extract 245 1.92
Ammonium sulfate fraction 99 18.98 X 10 -3 1.88
P-cellulose (pH 7 .5) 8.1 0.16 1.31
Fat body (day 3 of the 5th ins tar)
16.63 X 10 -3
Crude extract 398 6.62
Ammonium sulfate fraction 246 33.29 X 10 -3 8.19
P-cellulose (pH 7 .5) 4.2 0.33 1.40
Aldolase F
Newly hatched larva
Crude extract 181 11.70x1o-3 2.12
P-cellulose (pH 6.0) 1.5 0.48 0.74
Muscle* (day 3 of the 5th instar)
Crude extract 218 6.18 X 10 -2 13.47
P-cellulose (pH 6.0) 8.4 0.48 4.02
* Containing cuticles and epidermal cells.
Recovery
(%)
100 98 68
100 124 21
100 35
100 30
TABLE 14. Comparison of some properties of partially purified aldolase isozymes.
Aldolase S Aldolase F Egg Fat body Neonate Muscle FBP/F1P activity ratio 3.3 2.5 12.0 10.5
Km for FBP (�M) 5 3 7 5
pH and temperature optima
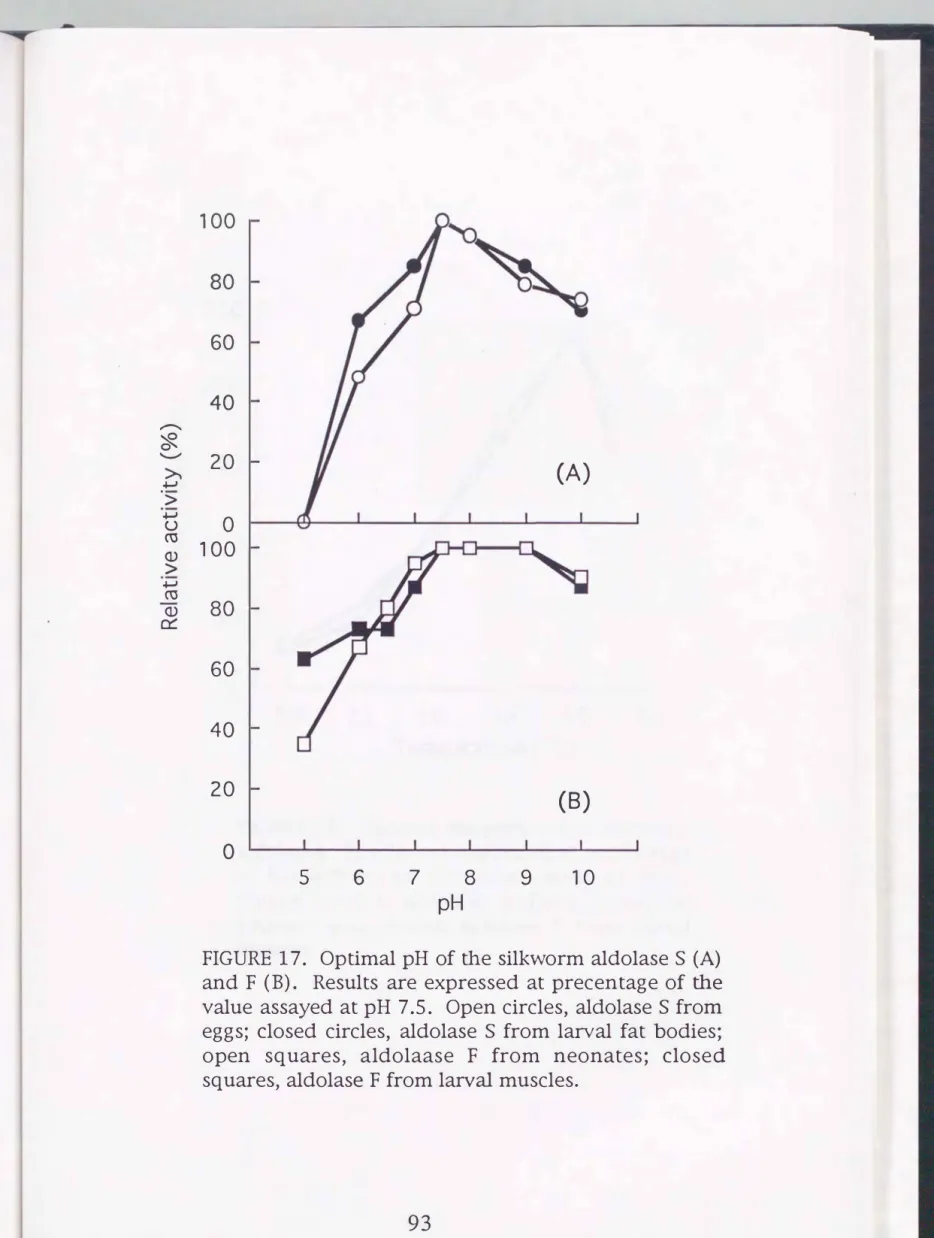
The pH for optimum activity for both enzymes is 7.5 (Fig. 17).
Aldolase F was characterized by the rather broader pH optimum than aldolase S. The partially purified S and F isozymes were determined for activity under the standard assay conditions but at varying temperatures. The both enzymes showed the highest catalytic activities at 50°C (Fig. 18).
Effect of temperature and other properties of partially purified silk
worm aldolases
Plots of S and F activities as a function of temperature gave different curves (Fig. 19). Aldolase F was more rapidly inactivated, than aldolase S, whose activity remained at 65% of the initial value after 4 hr at 30°C. One of the difference between class I and class II aldolases is that the latter are activated by K+ and NH4 + ions, and inhibited by EDTA (Rutter, 1964). The activity of silkworm aldolases was unaffected by 10 mM KCl and EDT A. These results repeatedly suggest silkworm aldolases are of the typical class I.
Immunological detection of
Sand
Fisozymes in crude extracts after
2Delectrophoresis
Preliminary experiments indicated that antiserum against purified type-S egg aldolase reacted with both of the two types of aldolase treated with the modified O'Farrell buffer (9.5 M urea, 2%
NP-40 and 2% 2-mercaptoethanol). Then eggs, neonates, fat bodies and muscles (plus cuticles and epidermal cells) were extracted for proteins with an O'Farrell buffer containing urea and the super
natants were subjected to O'Farrell type 2D gel electrophoresis.
92
� ..._.,
�
>.
�
'>
� u ro
> (1)
� ro 0::: (1)
100 80 60 40
20
(A)
0 100 80 60 40
20
(B)
0
5 6 7 8 9 10
pH
FIGURE 17. Optimal pH of the silkworm aldolase
S (A)
and F
(B).
Results are expressed at precentage of the value assayed at pH 7.5. Open circles, aldolaseS
fromeggs; closed circles, aldolase
S
from larval fat bodies;open squares, aldolaase F from neonates; closed squares, aldolase F from larval muscles.
....-...
'-" �
..., � '>
...,
u C'O
> (])
...,
C'O 0::: (])
250
200
150
100
50
0
10 20 30 40 50 60
Temparature
(°C)
FIGURE 18. Optimal temparature of silkworm aldolases. Results are expressed at percentage of the activity of the value assay at 30°C.
Closed circles, aldolase S from larval fat bodies; open circles, aldolase F from larval muscles.
100
�80
�
>.
..J.-J
:> 60
..J.-J
u ro
.� 0)
40
c ro E
� 20
0
0 20 40 60 240
Incubation time (min)
FIGURE 19. Thermal stability of silkworm aldolases. The partially purified preparations were each preincubated at 30°C for indicated periods of time and residual activity was assayed by spectrophotomeric method under the standard assay conditions. Closed circles, aldolase S from larval fat bodies; open circles, aldolase F from larval muscles.
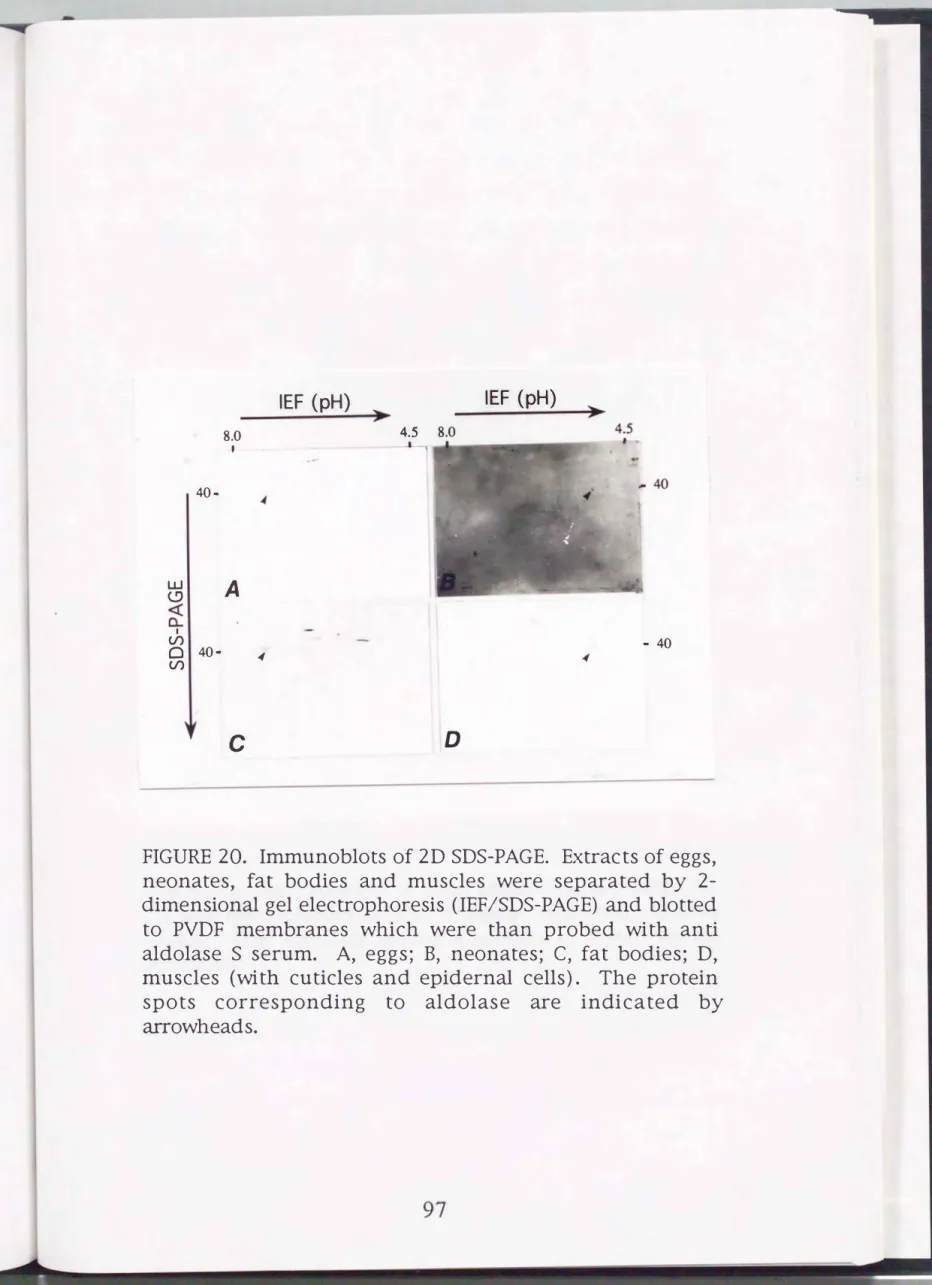
The separated proteins were analyzed by immunostaining using anti-aldolase S serum (Fig. 20). The serum reacted with several spots. The positive spots with a molecular weight of about 40,000 and pi of about 7.5 were detected in extracts from eggs (day 1 after the start of embryogenesis) and larval fat bodies (day 3 of the 5th instar). On the other hand, the spots with a molecular weight of about 40,000 and pi of about 5.2 were detected in extracts from neonates and larval muscles (day 3 of the 5th instar). These components are considered to be silkworm aldolase proteins. Thus it can be concluded that the pi values for S and F are 7.5 and 5.2, respectively.
An extract of the mid-gut, which gave the five bands on zymogram (Chapter III), was applied to 2D gel electrophoresis and reacted with anti-aldolase S serum. Only the spot with molecular weight of about 40,000 and pi of 7.5 was detected (Fig. 21 ). This organ must have both S and F and the reason why no F spot was observed remain unclear.
Properties of aldolases II, III and IV
Gel filtration of the partially purified aldolase S and F preparations indicated that their molecular weight was about 160,000. On SDS-PAGE the partially purified isozyme preparations gave a major band with a molecular weight of about 40,000. The author suggest that the silkworm aldolase isozymes are tetramers, and the five bands found in many organs are formed by the random tetrameric association of the two subunits, i.e. aldolase S = S 4, aldolase IV = S3F, aldolase III = S2F2, aldolase II = SF3 and aldolase F = F 4. Indeed, when the two partially purified isozyme
96
40- 8.0
'
w
A
(.9
<{
a.. I
V) 0 40- �
V)
c
IEF
(pH)
.,. IEF
(pH)
4.5 8.0
- 40
D
FIGURE 20. Immunoblots of 2D SDS-PAGE. Extracts of eggs, neonates, fat bodies and muscles were separated by 2- dimensional gel electrophoresis (IEF/SDS-PAGE) and blotted to PVDF membranes which were than probed with anti aldolase S serum. A, eggs; B, neonates; C, fat bodies; D, muscles (with cuticles and epidernal cells) . The protein spots corresponding to aldolase are indicated by arrowheads.
8.0
L1J
40
..._.(!) <(
a. I
(/) 0 (/)
I
IEF (pH)
4.5
t
FIGURE 21. Immunoblots of mid-gut proteins separated by 2D gel electrophoresis and stained with anti-aldolase serum. The plausible aldolase protein is shown by an arrowhead.
98
preparations from the larval fat bodies and muscles were mixed in
1
M NaCl and subjected to the three cycles of freezing (at -80°C for
1 hr)
and thawing, weak aldolase activity was detected between the S and F isozyme bands on zymogram analysis, although five bands were not resolved clearly (Fig.22).
Isozyme preparation S or F alone did not give any bands or smears besides the respective S or F band after the freezing and thawing.Detection of silkworm aldolase gene by Southern blot hybridization
The silkworm genomic
DNA
was subjected to electrophoresis and the gel was analyzed for the Southern blot hybridization with aeDNA
clone from human aldolaseB
as a probe (Fig.23).
TheeDNA
clone for human typeB
(Lane H) and D.melanogaster
aldolase (Lane
D)
exhibited the strong bands of hybridization.However, the silkworm genome gave only a trace band at about
1 0,000
bp (arrow of LaneB).
DISCUSSION
The type-S band was found to be the major aldolase compo
nent in eggs at early states of embryonic development and in the fat bodies of larvae, pupae and adults. On the other hand, the type-F band, which first appears after the head pigmentation stage of embryogenesis, was found to be distributed in the muscles (although the present muscle materials include cuticles and epidermal cells) of larvae, pupae and adults. Then, aldolase S was partially purified from eggs (on day
1
after the start of embryo-1 2 3
-
-S
F
+
FIGURE 22. Hybridization experiments of two types of silkworm isozymes. Aldolases S and F partially purified from the fat bodies and muscles, respectively, of larvae on day 3 of the 5th instar were used. Lanes 1, 2 and 3 contained the solutions of aldolase S, aldolase F and mixed aldolases S and F, respectively; each of the solutions was frozen and thawed in 1 M NaCl three times and subjected to zymogram analysis.
100
B D H M
Origin
23,130 (bp)
9,416 6,557 4,361 2,322 2,027
FIGURE 23. Southern blot analysis of B. mori
DNA.
GenomicDNA
ofB. mori (B), human type B aldolase
eDNA (H)
and D. melanogaster aldolaseeDNA (D)
were electrophoresed, blotted onto a filter and hybridized with Mboii fragment of human aldolase BeDNA
clone atlow stringency.
genesis) and larval fat bodies (on day
3of the 5th instar), whereas aldolase F from neonates and larval muscles (on day
3of the 5th instar). Previously aldolases S and F have also been purified (Nishimura, 1992). However, aldolase F was unstable with its recovery after purification being extremely low, and the comparison of enzymatic properties of the two aldolase isozymes was not fully conducted. Then, the methods of purification of F-type were modified (Yara, 1994), where the recovery from crude extracts of neonates was much improved. Using the purified preparations, Yara (1994) reported that the values of the FBP/F1P activity ratio for aldolases S and F were
3and 1, respectively. The present study with partially purified preparations gave the corresponding values of
3and 11-12, for S and F, respectively, indicating that previous value of 1 for F is inadequate. It is possible that molecular forms of silkworm aldolases (in particular F type) tend to be changed during purification, so that the apparent activities for substrate (in particular for F1P) are modified. The author found that avoiding long term incubation with FBP reduces a confusion. In addition, the fraction by salting out was omitted in the purification, since F-type isozyme is unstable.
It should be mentioned here that there were differences in value of the FBP/F1P activity ratio between the crude extracts and the partially purified preparations even within the same isozyme.
The crude S enzyme preparations gave the value of
6for eggs and 17 for fat bodies (see Tables
4and 9 of Chapters I and III, respectively) , but the partially purified S enzymes gave values of about 3 (Table 13 of the present chapter). Similarly, the crude F (neonates and muscles) gave the values of about 30 (Tables
4and
102
9),
whereas the partially purified F gave values of 1 1 to 12 (Table 13 ). Irrespective of the modification of the assay system, the values for the activity ratio obtained using crude extracts still seemed to have some inadequacy.The similarity in enzymatic properties, in behavior during P
cellulose chromatography, in electrophoreic mobility on zymogram and in 2D Western blotting analysis suggested that isozyme S (or F) from eggs (or neonates) are the same as (or very similar to) the corresponding ones from the post-embryonic organs; the author proposes that this is the case for all the S (or F) isozymes detected from different sources, although these points must be confirmed by structural analyses.
The Km values of silkworm aldolase isozymes for FBP (3 to 7
x 1 o-6) were similar to those of vertebrate and other insect aldolase isozymes but lower than that of He fix pomatia (snail) muscle aldolase (3 x 10-7; Buczylko et a/., 1980). Isoelectric focusing analyses showed several peak of partially purified silkworm aldolases (not shown). The pi values determined by 2D gel electrophoresis followed by Western blotting analysis were 7.5 and 5.2 for aldolase S and F, respectively. The pi values of aldolases from other insects were in the range of 4.9 to 6.0. Also rabbit aldolase exhibits pi of 6.1 (Bergmeyer et al., 1963). Table 14 sum
marized some properties of insect aldolases.
It is vertebrate class I aldolases that have the three isozymic forms. Three parental aldolases have been isolated from the rabbit:
isozyme A from the muscle (Taylor et al., 1948); isozyme B from the liver (Rajkumar et al., 1966); and isozyme C from the brain (Penhoet et al., 1967). These have a distinct substrate specificities,
which are well indicated by the FBP/F1P activity ratios. Aldolase S of B. mori can be considered to function similarly to the mam
malian liver-type isozyme B in view of the similarity of the physiological functions of insect fat body and mammalian liver, despite that the FBP/F1P activity ratio of aldolase S (about
3)
wasintermediate between those of aldolase B
( 1)
and the brain type aldolase C (12). The activity ratio of silkworm aldolase F resembled that of mammalian aldolase C but was different from that(50)
ofrabbit skeletal muscle aldolase A, although there is the possibility that enzyme F is a counterpart of the mammalian muscle isozyme.
The activity ratio to the two substrates of the silkworm F enzyme was reminiscent of the value
(10)
of D. melanogaster aldolase (Brenner-Holzach and Leuthardt,1967, 1969;
Malek et a!.,1985;
Shaw-Lee et a!.,
1992;
Kim et a!.,1992;
Kai et a!.,1992).
In addition, the activity ratio of aldolases isolated from larval inter
segmental muscle and adult flight muscles of the blowfly P. regina was
20
(Table15).
On the other hand, the ratio of the flight muscle aldolase from Ceratitis capitata, Schistocerca americana gregaria and Epiblema saidderiana were48, 37
and40,
respectively (Table14
), closer to the50
of the mammalian aldolase A. Aldolases having high values have not been isolated from the silkworm and the fruit fly.Through the characterization of chimerical enzymes as well as wild-type enzymes, a hypothetical evolutional process of vertebrate aldolases has been debated as follows (Kusakabe,
1994
). First, a prototype aldolase molecule was divide into two types, B prototype isozyme and A/C prototype isozyme by gene duplication. The prototype B isozyme was possibly adapted to the need of fructose104
�
0 Vl
TABLE 15. Summary of characteristics of insect aldolases.
Bombyx Ceratitis* 1 Drosophila *2 Epiblema*4 Phormia mori capitata melanogaster saidderiana regina
Schistocerca *7 americana gregaria
Source Egg Neonate Whole body Whole body
Fat body *5 Muscle *6
Muscle Muscle
Fat body Muscle (Pupa) (Larva)
Molecular weight
(Native) 160,000 160,000 158,000 (SDS-PAGE) 40,000 40,000 39,000
Km for FBP (�M) 5 5 4.3
FBP/F1P activity ratio 3 10 48
pi 7.5 5.2 6.6
Optimum pH 7.5 7.5- 9.0 7.4
* 1 Femandez-Sousa, J. M. et al. (1978)
*2 Brenner-Holzach and Leuthardt (1971)
160,000 160,000 160,000
40,000 37,800 40,000
27 1.13 3.0 5.7
20 oo*3) 40 5 20
5.0-5.1
6.8- 8.0 6.0 7.8 7.3
* 3 As estimated by the spectrophotometry method as describrd in Chapter I (Sugimoto, unpublished)
*4 Holden and Storey ( 1994)
* 5 Bauer et al. (1978), estimated using crude extracts.
*6 Levenbook et a/. (1973); Bauer et al. (1974)
*7 Storey et al. (1980)
155,000 40,000
4.0
37 4.9 7.3
metabolism by getting the high PIP-catalyzing activity with the FBP/FlP activity ratio of
1.Second, the A/C prototype isozyme was divided into A and
Cprototype isozymes via gene duplication. The proto-isozyme
Aacquired the high PEP-catalyzing activity with the FBP/FlP activity ratio of 50 and evolved to the modern type of isozyme A. Precise typing and molecular evolutionary aspects of aldolase isozymes in the insect world would give crucial points for the better understanding of the evolution of aldolase.
A difference in pH-activity profiles was observed between silkworm and other insect aldolases; the optimum pH for the silkworm enzymes were broader than those for other insects giving sharp patterns. The patterns for vertebrate aldolases are also broad.
However, this difference may be more apparent than real, probably due to the composition of the buffering reaction medium rather than to a unique property of the insect enzyme. The insect enzymes except for the silkworm enzyme were studied with a universal buffer containing acetate, phosphate and borate ions. Interestingly, the mammalian and silkworm aldolases were inhibited by phos
phate and borate; in this case glycylglycine buffer made the pH
activity profiles much more broader than other buffers (un
published results).
The molecular weight of the present aldolases was about
160,000
(and
40,000in SDS-PAGE), in agreement with the values for mammalian and
D. melanogasterclass I aldolases, which form tetramers (cf. Brenner-Holzach and Leuthardt, 1972; Malek
et al.,1985). Zymograms of the
B. morienzymes from different organs gave five distinct forms. In the vertebrate brain and other tissues aldolase A and C are co-expressed and produced the five members
106
of tetrameric forms (Lebherz and Rutter, 1969; Penhoet
eta/.,1966).
Similarly, the A-B hybrid forms were observed in the liver and kidney (Lebherz
andRutter, 1969; Penhoet
et al.,1966). Aldolase
Chas not been known to make hybrids with B-type
in vivo.However, when isozymes B and
Cwere co-expressed in
E. coli,the
five possible combinations of tetrameric forms were detected
(Kusakabe, 1994 ). An attempt to generate heteropolymers of
aldolases S and
Findicated that enzymatically active intermediate
forms were produced. The subunit hypothesis for isozyme systems
such as LDH (Markert, 1963) might be applicable to the present
aldolases, making all possible combinations: two homomeric
species (S4, F4) and three hybrids (S3F, S2F2, SF3). Some of the
larval, pupal and adult organs examined seemed to lack the
heterohybrids; the occurrence of the latter may depend upon the
conditions in the respective organs. LDH is coded by two different
genes; one for M subunit and the other for H subunit. The
functional LDH enzyme is a tetramer. If two subunits genes are
being expressed equally and hybridization is random, the expected
ratio of isozyme types are; 1(H4) : 4(H3M1) : 6(H2M2) : 4(HM3) :
1 (M4). However, muscle LDH is predominantly made of M
subunits, whereas heart LDH are composed of H subunits. The rate
of synthesis of LDH-M subunit does not vary greatly from tissue to
tissue. On the other hand, the degradation of LDH-M is about
twenty times greater in heart than in skeletal muscle (Fritz
et al.,1969). These findings suggested that the composition of LDH is not
controlled at the level of transcription or translation, but at the level
of enzyme degradation. In the case of the silkworm aldolase
system, the occurrence and distribution of aldolase isozymes seem
to have relevance to the extreme instability of isozyme F. What is the relation of the isozyme stability to the adaptation to the type of sugar metabolism is an open question to be investigated in near future.
The present findings clearly showed that the silkworm aldolase isozymes belong to class I and are similar to vertebrate and other insect aldolases in enzymatic properties. The amino acid sequences, which were deduced from the nucleotide sequence of eDNA or genomic clones, are highly conserved among the aldolase isozymes of different vertebrate species. In particular, the active site sequence are highly conserved through class I aldolases known to date (Kitajima et al.,
1990).
A eDNA clone of D. melanogaster aldolase was hybridized with fragments of mammalian aldolase B which contained a highly conserved region. D. melanogaster aldolase is60
to70%
homologous to mammalian aldolases (ShawLee et al.,
1992;
Kim et al.,1992,
Kai et al.,1992).
If B. morialdolase gene was analogous to D. melanogaster aldolase gene and more than
60%
homologous to mammalian aldolases, strong hybridization bands would be detected in Southern blot experiments. This prompted the author to detect the genome of B.mori containing an aldolase sequence by using an authentic class I aldolase probe. However, DNA from eggs of B. mori was faintly hybridized with this probe. Also RNAs from eggs during embryogenesis were weakly hybridized with the eDNA clones of human and D. melanogaster (Sugimoto et al.,
1992).
These results suggested that the silkworm aldolases have unique sequences compared with those of other species or that some inadequacy in108
the experimental procedures interfered the hybridization. The latter may be true as will be described in GENERAL DISCUSSION.
SUMMARY
Two types of aldolases, S and F, were partially purified; S from eggs and larval fat bodies, and F from neonates and larval muscles.
The Km values for FBP and their molecular weights of S and F were found to be similar to those of aldolases from vertebrates. The partially purified S and F preparations differed in FBP/FlP activity ratio, the values being about 3 for S and about 10 for F, suggesting that aldolase isozymes of B. mori play distinct roles on the metabolism of sugar-phosphates. Heterohybridization test of the S and F preparations in vitro produced intermediary migrating materials in zymogram. This observation indicated that the occur
rence of five components in the silkworm organs may be due to the formation of tetrameric hybrids with the S and F subunits. A trial to detect silkworm aldolase gene by Southern blotting gave only a sparse hybridization signal.
GENERAL DISCUSSION
In the present study, the author analyzed changes and distribution of fructose-1 ,6-bisphosphate aldolase of B. mori. The aldolase system has the advantage in that this enzyme plays a crucial role in glycolysis and its function is of interested. Moreover, aldolase has isozymes in a variety of molecular forms, which are potentially differentiated in physiological significance (Markert and Whitt, 1968; Markert et al., 1975).
As described in the first part of Chapter I, the spectrophoto
metric assay used previously was found to suffer from a non
specific change of absorbancy occurring before the addition of substrate to the reaction mixture. This inadequacy has not been pointed out in preceding papers, since most experiments for aldolase have so far been done with purified enzymes which scarcely produce non-specific changes. However, the estimation of aldolase activities using crude extracts is often convenient to understand facility the relative features among tissues. The author modified the assay method to be applicable to crude extracts. It works well also for purified preparations and found to give better results than the previous method. This modified method, which is not complicated, therefore must be enployed for all of the aldolase studies from now on.
The crude extracts from eggs, larvae, pupae and adults ex
hibited S and F bands of aldolase on zymogram. Thus, it was con
sidered that two fundamental types of aldolase isozymes are present in the silkworm. There was an interchange of these isozyme types from S to F at the head pigmentation stage of embryogenesis (see
110
the latter half of Chapter I). Aldolase S was observed in yolk of the eggs during embryogenic development until the stage of inter
change and aldolase F appeared in the embryo thereafter. In addi
tion, the transition from B to A type of hexokinase was also observed at the head pigmentation stage of embryogenesis (Chapter II). As to the human embryo, the interchanges of liver aldolase isozyme from A to B, the brain isozyme from A to C and the heart isozyme from C to A are known to occur at the period of birth.
These switchovers all occur gradually. In contrast, the switchover of aldolase and hexokinase during the silkworm embryogenesis is rather abrupt, and during interchange the aldolase and hexokinase activity levels exhibit a temporal decrease.
At the head pigmentation stage, or at a period close to this, changes in activity or content of various enzyme activities and other egg components are observable. These changes can be classified into four categories,
(1)
temporal decrease; not only the activities of aldolase and hexokinase but also those of chymotrypsin inhibitor (Aratake et al.,1990)
and alkaline phosphatase (Koga eta/.,1988)
take this pattern,
(2)
the onset of increase, shown in the activities of neutral and alkaline proteinases (Ikeda et al.,1990),
alkalinenuclease (Koga at al.,
1969),
mitochondrial glutamate-oxaloacetate transaminase and mitochondrial malate dehydrogenase (Yaginuma and Yamashita,1986)
the contents of eclosion hormone (Fugo et a!.,1985),
prothoracicotropic hormone B (Chen eta/.,1987)
and fibroin mRNA (Ohta et al.,1988), (3)
rapid decrease; overall protein and sugar components, e.g. vitellin, ESP, glycogen and trehalose (Zhu et al.,1986;
Okazaki and Yamashita,1981)
fall into this category,(4)
temporal increase; the ESP-specific proteinase activity rises and falls
sharply attaining a peak value at the period (Indrasith et al., 1988).
All these changes are thought to accompany disappearance of the yolk components and the activation of embryonic genes involved in differentiation of larval organs. The author proposes that these observable changes before hatching can be called the "prehatching transition."
The partially purified S and F preparations differed in FBP/F1P activity ratio, being 3 for S and 10 for F. On the other hand, previous measurements for the FBP/F1P activity ratio by using purified enzyme preparations have given perplexing results; the values were 3 for S and 1 for F (Nishimura, 1992; Y ara, 1994 ). The purification method applied in previous studies was suitable for the purification of aldolases from vertebrates and other insects.
However, the properties of B. mori aldolase isozymes seemed to be modified after the affinity chromatography with a P-cellulose column. The procedure includes the elution of aldolase with FBP, which is one of the substrates. After this step, the FBP/F1P activity ratio became low. There is the possibility that some specific changes in aldolase proteins are induced by FBP at low temperature.
Some properties of the two silkworm aldolases, except the FBP/F1P activity ratio and thermostability, were similar to those of mammalian aldolase isozymes. Aldolase F was much more un
stable than aldolase S and mammalian aldolases. Mammalian aldolase A may be the most stable one as far as the author's knowledge. Interestingly, a mutant human aldolase A, found in a patient with the hemal ytic anemia (erythrocyte deficiency), is unstable to heat than the wild-type molecule. The instability was
112
ascribed to an exchange of a single base (Kishi eta!., 1987). The difference between the silkworm aldolases in thermostability might be based on such a difference in the amino acid sequence.
Some organs of larvae, pupae and adults exhibited three additional zymogram bands between S and F (in total five bands).
In vitro mixing of the partially purified S and F preparations produced intermediary migrating materials, indicating that the occurrence of the five components may be due to the formation of heterotetramers of the S and F subunits (Fig.
24)
in the organs. Five types of A-B or A-C hybrid set have been detected in mammalian tissues (Rutter et a!., 1963; Penhoet et a!., 1966) and these can be produced by the random combination of the parental subunits in vivo (Penhoet et a!., 1967). In B. mori, some larval, pupal and adult organs, and eggs during embryogenic development, seemed to lack the hetrohybrids. The question to be solved is that whether the hybrids can be made under the genetic regulation of protein synthesis and/or the selective degradation of protein in B. mori organs.Isozyme expression corresponds to the physiological function in the tissues. The FBP/F1P activity ratio of aldolase is an important index depending upon the sugar-phosphate metabolism. This may be also true for the S and F silkworm isozymes, interchange during embryogenesis and e�pressed in an organ-specifically manner: S is the fat body type and F is the muscle type. Mammalian isozyme A with a high FBP/F1P activity ratio of 50 is distributed in the skeletal muscle where glycolysis is active, while mammalian isozyme B with a low ratio of 1 is localized in the adult liver where fructose utilization and gluconeogenesis are active (Rutter and Weber,
Subunits
s4V
(S)
IV Ill III (F)
FIGURE 24. Schematic representation of hybrid formation of silkworm aldolase. Arrow indicates the direction of electrophoresis; the perpendicular line, starting point.
114
1965).
Aldolase S was at a rather low ratio of 3, in favor of efficient utilization of F1P. This inference was supported by the observation that the activities of NADP-SDH and NAD-SDH, which are the enzyme for the polyol pathway generating F1P, were high in the young eggs and the fat bodies. On the other hand, aldolase F was as high as10
in FBP/F1P activity ratio, suggesting that it is less effective inF1P
utilization. This is in agreement with the fact that the NADP-SDH and NAD-SDH activities were low in neonates and in the muscles, where F1P utilization might be of less importance.As a whole, the changes and distribution in aldolase isozymes and other enzymes tested were considered to be well correlated with the carbohydrate utilization and glucose utilization in the eggs or the organs in which the respective enzymes are located.
Fig. 25 summarizes the polyol pathway and related routs with major enzymes responsible for the relevant reactions. Some additional features can be debated as follows. (A) Embryogenesis of diapausing eggs of the silkworm is arrested at the early embryonic stage (Yamashita and Yaginuma, 1991 ). At the initiation of diapause, glycogen present in the eggs is rapidly converted to sorbitol and glycerol and these polyols are maintained at high levels during diapause (Chino, 1958). Aldolas� S activity (adapted to F1P utilization) was detected in diapausing egg and this may be functioning in the formation of glycerol via F1P. When diapause is terminated and embryogenesis is restarted, sorbitol is used to resynthesized of glycogen and this conversion is promoted by NAD-SDH (Yaginuma and Yamashita, 1979). In addition, fructose was shown to be utilized during early post-diapause development (Okazaki and Yamashita, 1981 ). Again aldolase S may function
here. It is thus concluded that F 1P formed by the polyol pathway is catalyzed by aldolase S in diapusing and early embryogenic eggs.
A polyol phosphate, sorbitol 6-phosphate, is a probable precursor of sorbitol.
(B)
Sorbitol 6-phosphate is also detected in the larval hemolymph (Wyatt et al., 1963; Sakamoto and Rorie, 1979). The presence of polyol dehydrogenase and that of phosphatase activities also have been demonstrated in the hemolymph and fat body (Faukner, 1956). Changing activities of aldolase S in the fat body will be connected with the utilization of sorbitol 6-phosphate in the silkworm hemolymph.
(C) The reaction of NADP-SDH produces NADP and that of NAD-SDH produces NADH. The continuous fructose formation by the polyol pathway must be supported by the coupled consumption of NADP and NADH in the cytoplasm, since NADP-SDH and NAD
SDH are cytosolic enzymes. Generally, LDH is a good cytoplasmic acceptor of NADH. In the insect thoracic muscles, however, LDH is almost totally undetectable and a-glycerophosphate dehydrogenase is of vital importance in transferring cytosolic NADH into mitochondrial equivalents and thus ensuring the maintenance of the highest possible glycolytic flux (Kubita, 1958; Chefurka, 1958). A similar situation has been revealed in the silkworm eggs (Chino 1958, 1960; Kageyama, 1976), where LDH is low in activity and instead cytosolic a-glycerophosphate dehydrogenase and malate dehydrogenase, and in turn the a-glycerophosphate cycle and malate-aspartate cycle in mitochondria, are physiological acceptors of NADH (Chino 1960; Yaginuma and Yamashita, 1986). As to NADP, it can be consumed by G6PDH, extramitochondrial iCDH,
116