J Tokyo
(
63 (Extra)
Yilgsm-.,I)gsd,,8,o,il)
RELATIONSHIP BETWEEN HLA AND SEIZURE DISORDERS
AMONG CHINESE CHILDREN IN SOUTHERN TAIWAN
Chi-Hsiun CHIANG
Department of Pediatrics, Kaohsiung Medical College, Taiwan, Republic of China
(Received June 22, 1993)
The purpose of this study was to investigate the occurrence of human leukocyte antigens (HLA antigens) and haplotypes in children with different types of seizure disorders and their families. The sample consisted of 169 unrelated mentally normal
Chinese children in Southern Taiwan with seizure disorders, and 284 of their parents. The types of seizures were divided into idiopathic generalized seizures (74 cases), simple partial seizures (33 cases) and febrile convulsions (62 cases). HLA-Cw7 (p<O.OOI) was found to be significantly more common in both patients and thelr parents with simple
partial seizures and febrile convulsions. The prevalence of the HLA-Bw6 (p<O.OOI)
antigen was higher, to a statistically significant degree, in parents of children with
idiopathic generalized seizures as was HLA-Bw22 (p<O.OOI) in patients with simple
partial seizures. Only HLA-Cw3 (p<O.OOI) appeared to be present at a significantly lower frequency in parents of children with idiopathic generalized seizures than in controls. The prevalence of the All; Bw6 haplotype (p<O.OO05) was significantly higher in both patients and parents of children with idiopathic generalized seizures, and the A9; Bw6 haplotype (p<O.OOI) in parents of children with simple partial seizures. Among those
with febrile convulsions, there were no statistically significant increases in the
frequencies of these haplotypes, but the frequency of haplotype All; B40 was relatively low in parents. Since the HLA A2'; B40 and All; B40 were the most common haplotypes in normal controls, the relatively low frequency of the All; B40 haplotype in parents of
children,with febrile convulsions may support the hypothesis of "protecting" genes
which'somehow counter the effects of certain noxious mechanisms, and the lack of such a haplotype may point to an inherited susceptibility to seizure disorder. However, an investigation with HLA typing in larger populations. of epileptic individuals and their
families may give a more reliable and conclusive answer regarding the genetic
characteristics of seizure disorders. Furthermore; the results must be interpreted very
cautiously.
Introduction
Epilepsy is probably an inherited disorder. The fact that seizures sometimes occur with increased
frequency among families of patients with
epi-lepsy has long been recognized by clinicians and the public alike.
Several recent studies have provided evidence, both direct and indirect, that epilepsy is inherited.
For instance, clinical studies of epileptic Probands and their relativesi)2), electroencephalogrqphic (EEG) studies3>N6), twin studies7)"9), and pedigree studiesiO}Ni2) have provided direct evidence, and
studies on non-specific familial EEG
pat-terns4}i3}Ni5}, the inheritance of epilepsy in
animalsi6)i7), hereditary diseases associated with epilepsyi8)N2i), and chromosomal abnormalities as-sociated with epilepsy22}23) have contributed
-direct evidence. However, some of these data are still not entirely convincing, and reliable scientific data either confirming or refuting this hypothesis have not been easy to obtain.
In the recent years, the relationship of human leukocyte antigens (HLA antigens) to certain dis-eases has attracted interest since the discovery of
the close linkage between histocompatibility genes and those of the immune response in
ex-perimental animals24)25), and similarly in man, a
relationship has been found between diseases
with alteration of the immune system and
markers in the major histocompatibility complex
(MHC) chromosomal region, serum-defined
human leukocyte antigens (SDHL-A),lymphocyte-defined antigens, and mixed lymphocyte reaction antigens (LD MLR)26)N28). Therefore, it has been realized that MCH is not only intimately involved
in regulation of the immune system, but also
shows extreme polymorphism and is the most
reliable set of human genetic markers so far
found, and may thus be helpful for identifying
individuals at risk for epilepsy.
The purpose of this investigation was to study the distribution frequencies of HLA antigens and
haplotypes among Chinese children in Southern
Taiwan with seizure disorders and their families,
in hopes of identifying HLA correlations with
these disorders.
Subjects and Methods
The subjects comprised 169 unrelated, mentally
normal Chinese children in Southern Taiwan
with seizure disorders. A study of history, clinical
examination, routine laboratory investigations
and EEG were performed to exclude other causes.
No definite causes of seizures, inheritance of
known central nervous system (CNS) disorders, or
no chromosomal anomalies, were found and all parents were free of seizures. A total of 284
parents of the patients agreed to participate in the investigation, which was a prerequisite. All were enrolled consecutively at the Department of Pedi-atrics at Kaohsiung Medical College Hospital. Five hundred and eighty-four healthy, unrelated
indi-viduals were included as a control group for HLA
antigen analysis. They were selected randomly
from among persons who visited the hospital for minor surgical conditions (not of genetic origin)
and nonconsanguineous parents of children
ex-amined in the pediatric outpatient clinic for sim-ple nongenetic diseases. Among them, 584 volun-teers were tested for class I antigens and 134 of these 584 for class II antigens, in the latter period of this investigation, and haplotype frequencies were determined for 252 controls.
The types of seizures were classified in accord-ance with the International League Against Epi-lepsy (ILAE) recommendations29) into idiopathic generalized seizures (74 cases) and simple partial seizures (33 cases), and febrile convulsions (62
cases) were added to the study as a special
syndrome.
1) HLA-A, -B, -C, and -DR typing
HLA antigens were identified on lymphocytes separated from heparinized peripheral blood by means of the Ficoll-Hypaque density gradient
centrifugation technique30>, and the microcytoto-xicity test of Mittal et al.3i) and the Terasaki32)
panel plate method. DR typing was performed on
B cell-enriched peripheral mononuclear cells. The
B cell enrichment was accomplished by a method
based on the different mobilities of cells in a discontinuous density gradient of Percol133). The
microcytotoxicity test and the Terasaki-panel
plate method were also used.
The following HLA antigens were examined in
this investigation:
HLA-A
HLA-B
HLA-C
HLA-DR
The antisera
Corp, West Germany,
DR from C-six Diagnostics
1, 2, 3, 9, 10, 11, w19, w23, 24, 25, 26, 28, w32 w4, 5, w6, 7, 8, 12,-13, 14, 15, w16, 17, 18, w21, w22, 27, w35, 37, w38,
40, w44, w49, w51, w55, w56,
w60, w73
w2, w3, w4, w7 1, 2, 3, w4, 5, w6, 7, 2 + Qwl, MT2,MT3
were purchased from Boehring
and the antisera for HLA
, Inc., USA.
-E189-2) Statisticalmethods
HLA antigen frequencies (ab were determined
by direct counting. The association of a particular disease with a particular HLA antigen was quan-titated by calculating the "relative risk". The relative risk (RR) values were calculated accord-ing to the followaccord-ing equation34).
RR =Pd (1 ff Pc)/Pe (1 - Pd)
where Pd is the frequency in patients and P, that
in controls. RR indicates how many times more
frequently the disease occurs in a group of indi-viduals carrying the antigen relative to a group lacking it.
Gene frequency was estimated according to the following equation:
g=1-Vi:f
where g is the gene frequency and fthe antigen
phenotype frequency in the population.
For estimation of the haplotype frequencies
from the phenotype data, the equation of Ceppel-lini et al.35) was used:
X,j==P,Pj + D,j
were PiPj is the product of the gene frequencies of'
the antigens forming the haplotype and Dij the gametic association of these antigens. Since a
crossover within the HLA region is relatively rare, the demonstration that 2 siblings share the same 2 haplotypes, as judged on typing for HLA-A and B alone, usually implies that they are also identical
as to HLA-C, D, DR, and other genes within the
same region. This is fortunate, since most histo-compatibility testing laboratories can reliably type only the HLA-A and B antigens at the present
time36). Therefore, only HLA-A and B were
con-sidered for haplotype analysis in this study.
Linkage disequilibria, D (delta values), were
computed by subtracting from the haplotype fre-quency the product of the gene frequencies of the antigens describing the haplotype. The statistical significance of the linkage disequilibrium
coeffi-cient, "delta", was evaluated by means of an
approximate Chi-square calculated as follows:
4n (h-ggb)2
H-ggb
X2=
[1+
]2
gagb(2-g.) (2-gb) 2 (1-g,) (1-gb)where n is the number of subjects, h the estimated
haplotype frequency, and g, and gb the gene
・frequencies37}.
The Chi-square test with Yates' correction for
continuity was used throughout this
investiga-tion. Only values of p<O.OOI were considered
significant.
Results
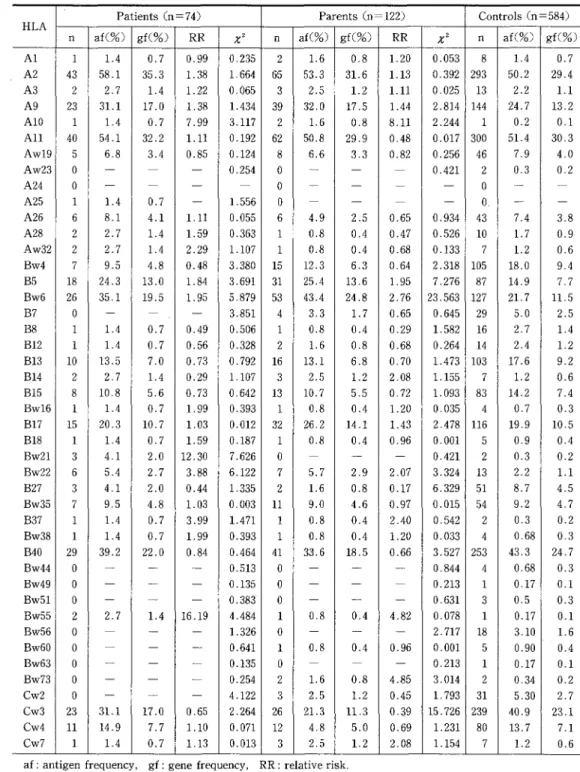
HLA antigens in patients
The HLA antigen distribution, gene frequencies and RR values for all 169 patients, with 3 different
types of seizures disorders, were studied, as
shown in Tables 1, 2, 3 and 4.
The HLA antigen did not exhibit a statistically significant distribution, except for a relatively high frequency and deviating RR value (positive
association) of antigen HLA-Bw21 (x2=7.626,
p<O.Ol, RR=・ 12.30) in patients with idiopathic
generalized seizures, as compared with normal
controls (Tables 1 and 5).
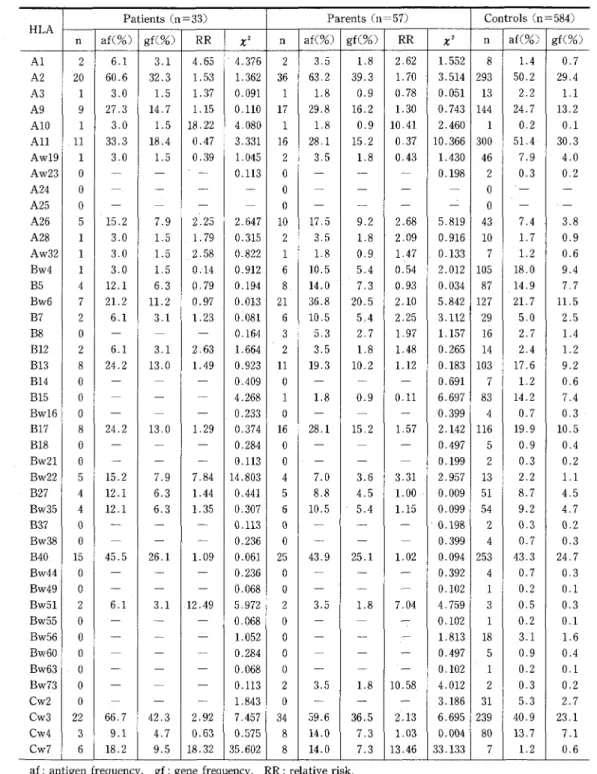
In patients with simple partial seizures, there were statistically significant high frequencies and deviating RR values (positive association) of
HLA-Bw22 (x2=14.803, p<O.OOI, RR=7.84) and -Cw7
(x2 :35.602, p<O.OO05, RR==18.32), and a relatively high frequenc'y and deviating RR value (positive association) of -Cw3 (x2=7.457, p<O.Ol, RR=2.92),
as compared with normal controls (Tables 2 and
5).
In patients with febrile convulsions, there was a statistically significant high frequency and devi-ating RR value (positive association) of HLA-Cw7 (x2=10.997, p<O.OOI, RR=7.23), and a relatively high frequency and deviating RR value (positive association) of -A2 (x2=10.402, p<O.O05, RR =2.70), , as compared with normal controls.
HLA antigens in parents
There was a statistically significant high fre-quency and deviating RR value (positive associa-tion) of HLA-Bw6 (x2==23.563, p<O.OO05, RR=2.76)
and a low frequency and deviating RR value
(negative association) of HLA-Cw3 (x2=15.726,
p<O.OOI, RR :O.39) in parents of idiopathic gen-eralized seizure cases, as compared with normal controls. HLA-B5 was found to show a relatively
-Table 1 Distribution of HLA antigens, gene frequencies, and RR values in idiopathic generalized selzures
Patients(n =74) Parents(n =122) Controls(n =584)
HLA
n af(%) gf(O/o)
RR
X2 n af(%) gf(O/o)RR
X2 n af(O/o) gf(%) Al I 1,4 O.7 0.99 O.235 2 1.6 O.8 1.20 O.053 8L4
O.7A2 43 58.1 35.3 1.38 1.664 65 53,3 31.6 1.13 O.392 293 50.2 29.4
A3 2 2.7
L4
1,22 O,065 3 2.5 1.2 1.11 O.025 13 2.2 1.1A9 23 31.1 17.0 1,38 1,434 39 32,O 17.5 1.44 2,814 144 24,7 13.2
AIO 1 1,4 O.7 7.99 3.117 2 1.6 O.8 8.11 2.244 1 O,2 O.1
All 40 54,1 32,2 1.11 O.192 62 50.8 29.9 O.48 O.O17 300 51.4 30,3
Aw19 5 6.8 3.4 O.85 O.124 8 6.6 3.3 O.82 O.256 46 7,9 4.0
Aw23 A24 A25 oo1 N-1.4 -LO.7 ... O.254 -1.556 ooo 7r- --- --- 0.421 --2oo. O.3 -" O.2
--A26 6 8.1 4,1 1.11 O.055 6 4.9 2.5 O.65 O.934 43 7.4 3.8
A28 2 2,7
L4
1.59 O.363 1 O.8 O.4 O.47 O.526 10L7
O.9Aw32 2 2.7 1.4 2.29 1.107 1 O.8 O.4 O.68 O.I33 7
L2
O.6Bw4 7 9.5 4.8 O.48 3.380 15 12.3 6.3 O.64 2.318 105 18.0 9,4
B5 18 24.3 13.0 1.84 3.691 31 25,4 13.6 1,95 7.276 87 14.9 7,7
Bw6 26 35,1 19.5 1.95 5.879 53 43.4 24.8 2.76 23.563 127 21.7 11,5
B7 o
m
-
.
3,851 4 3.3L7
O.65 O.645 29 5.0 2.5B8 1 1.4 O,7 O.49 O.506 1 O.8 O.4 O.29 1:582 16 2.7
L4
B12 1
L4
O.7 O.56 O.328 2 1.6 O.8 O.68 O.264 14 2.4 1.2B13 10 13.5 7.0 O.73 O.792 16 13.1 6.8 O,70 1.473 103 17.6 9.2 B14 2 2.7 1.4 O.29 1.107 3 2,5 1.2 2.08 1.155 7
L2
O.6Bl5 8 10.8 5.6 O.73 O,642 13 10.7 5.5 O.72 1.093 83 14.2 7.4
Bwl6 1 1,4 O.7 1.99 O.393 1 O.8 O.4 1.20 O.035 4 O.7 O.3
B17 15 20.3 10.7 1.03 O.O12 32 26.2 14.1 l.43 2.478 116 19.9 10.5
B18 1 1.4 O.7 1.59 O.187 1 O.8 O.4 O.96 O.OOI 5 O.9 O.4
Bw21 3 4.1 2.0 12.30 7.626 0
-m
-
O.421 2 O.3 O.2Bw22 6 5.4 2.7 3.88 6.122 7 5,7 2.9 2.07 3.324 13 2.2
Ll
B27 3 4,1 2.0 O.44 1.335 2
L6
O.8 O.17 6.329 51 8.7 4.5Bw35 7 9.5 4.8 1.03 O,O03 11 9,O 4.6 O.97 O.O15 54 9.2 4.7
B37 1 1,4 O.7 3.99 1.471 1 O.8 O.4 2.40 O,542 2 e.3 O.2
Bw38 1
L4
O.7 1.99 O.393 1 O.8 O.4 1.20 O.033 4 O.68 O.3B40 29 39.2 22.0 O.84 O.464 41 33.6 18.5 O,66 3.527 253 43.3 24.7 Bw44 o
T
-
m
O.513 o-
-
rm O.844 4 O.68 O.3Bw49 o
-
L
-
O.135 o-
--
O.213 1 O,17 O.1Bw51 o
-
T
-
O.383' o-
-
rm O.631 3 O.5 O.3Bw55 2 2.7 1.4 16.19 4.484 1 O.8 O.4 4.82 O.078 1 O.17 O.1
Bw56 o
-
m
-
1,326 o-
-
rm 2.717 18 3.10L6
Bw60 o
-
-
-
O.641 1 O.8 O.4 O.96 O.OOI 5 O.90 O.4Bw63 o rr
-
L
O.135 o-
-
-
O.213 1 e.17 O.1Bw73 o
m
-r
O.254 2L6
O.8 4.85 3.014 2 O.34 O.2Cw2 o
-
L
-
4,122 3 2.5 1.2 O.45 1.793 31 5.30 2.7Cw3 23 31.1 17.e O.65 2.264 26 21.3 11.3 e.39 15.726 239 40,9 23.1
Cw4 11 14.9 7.7 !.10 O.071 12 4.8 5.0 O.69 1.231 80 13.7 7.1
Cw7 1
L4
O.7 1.13 O.O13 3 2,5 1.2 2.08 1.154 7 1.2 O.6af: antigen frequency, gf: gene frequency, RR: relative risk,
high frequency (x2=7.276, p<O.Ol, RR=1.95) in the same seizure group (Tables 1 and 5).
There was a statistically significant high
fre-quency and deviating RR value (positive
associa-tion) of HLA-Cw7 (z2=33.133, p<O.OO05,
RR=13.46) in parents of simple partial seizure E191
-Table 2 Distribution of HLA antigens, gene frequencies, and RR values in simple partial seizures Patients (n=33) Parents (n=57) Controls(n =584)
HLA
n af(O/.) gf(%)
RR
X2 n af(%) gf(O/o)RR
X2 n af(O/o) gf(%) Al 2 6,1 3,1 4.65 4.376 2 3,5 l..8 2.62 1.552 8 1.4 O.7A2 20 60.6 32.3 1.53 1.362 36 63,2 39.3 1.70 3.514 293 50.2 29.4
A3 1 3.0
L5
1.37 O.091 1L8
O.9 e.78 O,051 13 2.2Ll
A9 9 27.3 14.7 l.15 O,110 17 29.8 16.2 1.30 O.743 144 24.7 13.2
AIO 1 3.0 1,5 18.22 4.080 1 1.8 O.9 10.41 2.460 1 O.2 O.1
All 11 33.3 18.4 O.47 3.331 16 28,1 15,2 O.37 10.366 300 5L4 30.3
Aw19 1 3.0
L5
O.39 1.045 2 3,5L8
O.43 1.43e 46 7.9 4.0Aw23 A24 oo T" -T L- O.113
m
oo -T -- -- 0.198"
2o O.3 -O,2 -A25 A26 o5 -15.2 -7.9 -2.25 -2.647 o10 -l7,5 m9.2 -2.68 =5.819 043 m7.4 -3.8A28 1 3.0
L5
1.79 O.315 2 3,5 1.8 2.09 O.916 10L7
O.9Aw32 1 3.0
L5
2.58 e.822 1L8
O.9 1,47 O.133 7L2
O.6Bw4 1 3.0 1,5 O.14 O.912 6 10.5 5.4 O.54 2.012 105 18.0 9.4
B5 4 12.1 6.3 O,79 O,194 8 14.0 7.3 O.93 O.034 87 14.9 7.7
Bw6 7 21.2 11.2 O.97 O.O13 21 36.8 20.5 2.10 5.842 127 21.7 ll,5 B7 2 6.1 3.1 1.23 e.osl 6 10.5 5,4 2・.25 3.1!2 29 5,O 2.5 B8 o
--
-
O.164 3 5,3 2.7 1,97 1.157 16 2.7L4
B12 2 6.1 3.1 2.63 1,664 2 3.5L8
l.48 O.265 14 2.4L2
B13 8 24.2 13.0 1.49 O.923 11 19.3 10.2 1.12 0.183 103 17.6 9.2 B14 o--
-
O,409 o -.L
-
O,691 7L2
O.6B15 o
-
-
`
4.268 1L8
O.9 O.11 6,697 83 14.2 7.4Bw16 o
-
-
-
O.233 o-
T
pt .O.399 4 0.7 O.3B17 8 24.2 13.0 l,29 O.374 16 28.1 15.2 1,57 2.142 116 19.9 10.5
B18 o
-
T
-
O,284 o-
r
-
O.497 5 O.9 O.4Bw21 o
m
-
m
0.113 o.
-
-
O.199 2 O.3 O.2Bw22 5 15.2 7.9 7.84 14,803 4 7.0 3.6 3.31 2,957 13 2.2
Ll
B27 4 12.1 6.3 1.44 O.441 5 8,8 4.5 1.00 O,O09 51 8.7 4,5
Bw35 4 12.1 6.3 1,35 O.307 6 10.5 5.4 1.15 O.099 54 9.2 4.7
B37 e
r
m
-
O.!13 od
m
-
G.I9g 2 O.3 O.2Bw38 0
-
-
L
O.236 oT
-
r
e.399 4 O.7 O.3B40 15 45.5 26.1 1.09 O,061 25 43.9 25.1 1.02 O.094 253 43.3 24.7
Bw44 o
-
m
-
O.236 oL
m
-
O.392 4 O.7 O,3Bw49 o rm
-
"
O.068 o-
-
r
O.102 1 O.2 O.1Bw51 2 6.1 3.l .12.49 5.972 2 3.5 1.8 7.04 4,759 3 ,O,5 O.3
Bw55 o
-
m
-
O,068 o-
-
-
O.102 1 O.2 O.1Bw56 o rm
L
m
1,052 o ne-
-
1.813 18 3.1 ,1.6Bw60 o
-
-
L
O.284 o-
-
-
O.497 5 O.9 O.4Bw63 o
-
-
-
O.068 o-
m
-
O.102 l 0.2 O.1 Bw73 o-
L
-
O.ll3 2 3.5 1.8 10.58 4.012 2 O.3 O.2Cw2 o
-
-
-
1.843 0L
`
-
3.186 31 5,3 2.7Cw3 22 66.7 42.3 2.92 7.457 34 59.6 36.5 2.13 6,695 239 40,9 23.1
Cw4 3 9.1 4,7 O.63 O,575 8 14.0 7.3 1,03 O.OQ4 80 13.7 7.1
Cw7 6 18.2 9.5 18.32 35.602 8 14.0 7.3 13.46 33.133 7 1.2 O.6 af: antigen frequency, gf: gene frequency, RR : relative risk,
cases, which was similar to that of patients with simple partial seizures, as compared with normal
controls, and a relatively high frequency and
deviating RR value (positive association) of HLA-Cw3 (x2=6.695, p<O.Ol, R =2.13) or relatively low
frequencies and deviating RR values (negative
-E192-Table 3 Distribution of HLA antigens, gene frequencies, and RR values in febrile convulsions Patients(ni=62) Parents(n= 105) Coritrols(n=:584)
HLA
n af(%) gf(%)
RR
X2 n af(%) gf(%)RR
X2 n af(O/o) gf(%) Al A2 o45 m72.6 N47.6 -2.70 O.863 10.402 o62 -59,O -36.0 -1.43 O.541 2.465 8293 1,4 50.2 O.7 29.4 A3 2 3.2 l.6 l.46 O.256 4 3.8 1.9 1.73 2.463 13 2.2 1.1A9 16 25.8 13.9 1.06 O.041 25 23,8 12.7 O.95 O.O04 144 24.7 13.2
AIO 1 1,6 O.8 9.56 O.561 3 2.9 1.4 17.15 6.950 1 O.2 O.1
All 23 37.1 20.7 O.56 O.214 36 34.3 18.9 O.49 O.633 300 51.4 30.3
Aw19 6 9.7 5.0 1,25 O.254 6 5.7 2.9 O.3! O.156 46 7,9 4.0
Aw23 A24 oo NN mm -m O.214
-oo 7m -- Lr O.132m
2o O.3 -O.2 -A25 A26 o11 -17.7 -9.3 N2.71 -6.594 o14 "13.3 rm 6.9 m1.94 m3.425 o43 rm 7.4 -3.8A28 2 3.2
L6
1.91 O.120 7 6.7 3.4 4.10 7.140 10L7
O.9Aw32 o
-
-
-
O.756 o-
-
-
O.475 7 1.2 O.6Bw4 15 24.2 12.9 1.46 1,047 22 21.0 11.1 1.21 O.346 105 18.0 9.4
B5 3 4.8 2.4 O,29 3.928 9 8,6 4.4 O.54 O.OOI 87 14.9 7.7
Bw6 17 27.4 14.8 1.30 1.042 25 23,8 12.7 1.12 O.117 !27 21.7 11.5
B7 1
L6
O.8 O.3! 1.422 7 6.7 3.4 l.37 2.681 29 5.0 2.5B8 o
'
'
m
1.741 2L9
1.0 O.69 O.030 16 2.7L4
B12 1
L6
O.8 O.67 O.155 2L9
LO
O.79 O.101 14 2.4 1.2B13 9 14.5 7.5 O.79 O,382 17 16.2 8.5 O.90 2.233 103 17.6 9.2・
B14 2 3.2
L6
O.75 1.687 2 1.9LO
1.60 1.Q65 7 1.2 O.6B15 3 4.8 2,4 O.31 3.489 5 4,8 2.4 O.30 1.136 83 14.2 7.4
Bw16 o
-
--
O.433 2L9
LO
2.82 2.483 4 O,7 O.3B17 13 21.0 11.1 1.07 O.041 19 17.1 9.0 O.89 2.481 116 19.9 10.5
B18 o
L
-
L
O.532 o-
-
-
O.344 5 O.9 O.4Bw21 o
"
'
'
O.214 o"
-
-
e.139 2 O.3 O.2 Bw22 4 6.5 3,3 3.03 2.437 8 7,6 2.9 3.62 7.031 13 2.2Ll
B27 6 9.7 5.0 1.12 O.060 13 12.4 6.4 1.48 O.853 51 8.7 4.5 Bw35 1 1.6 O.8 O,16 4.191 3 2.9 1.4 e.29 O.852 54
92
4.7B37 o
-
-
-
O.214 o rm-
m
O.132 2 O.3 O.2Bw38 o o o
r
O.431 1LO
O.5 1.39 O.403 4 O.7 O.3B40 34 54.8 32.8 1.59 3.012 47 44.8 25.7 1.06 O.077 253 43.3 24.7
Bw44 1 1.6 O.8 2.38 O.631 1 1.0 O.5 1.39 O.401 4 O,7 O.3
Bw49 o o o
-
O.113 eL
-
-
O.071 1 e.2 O.1Bw51 1
L6
O.8 3,l7 1.102 2L9
1.0 3.76 3.382 3 O.5 O.3Bw55 o
-
m
N
e.113 1LO
O.5 5.61 2.384 1 O.2 O.1 Bw56 o--
-
1.971 o rr'
fi 1.242 18 3.1L6
Bw60 o
-
-
-
O.531 1 LO O.5 l,11 O.223 5 O.9 O.4Bw63 o
-
-
-
O.113 o-
-
-
O.071 1 O.2 O.1Bw73 2 3,2 1.6 9.70 3.670 4 3.8
L9
11.52 8.765 2 O.3 O.2Cw2 1
L6
O.8 O.29 1.632 3 2.9 1.4 O,52 O.023 31 5.3 2.7Cw3 32 51.6 30.4 1.54 2.634 50 47.6 27.6 1.31 1.639 239 40.9 23.1
Cw4 6 9,7 5.0 O.68 O,794 7 6,7 3.4 O.45 O.182 80 13.7 7.1
Cw7 5 8.1 4.1 7.23 10.997 11 10.5 5.4 9.65 26,618 7
L2
O.6af:antigen frequency, gf:gene frequency, RR:relative risk.
'
association) of HLA-All (x2=10.366, p<O.O05, group(Table2).
RR=O.37) and HLA-B15 (x2 :6.697, p<O.Ol, RR= For febrile convulsions, HLA-Cw7 (x2==26.618,
O.11) were found in the simple partial seizure p<O.OO05,RR=9.65) was found to
-E193-cally significant high frequency among parents, as in patients, and the HLA-AIO (x2=:6.950, p<O,Ol,
RR==17.15), -A28 (x2=7.140, p<O.Ol, RR=4.10), -Bw22 (x2==7.031, p<O.Ol, RR=3.62), and -Bw73 (x2=8.765, p<O.Ol, RR=11.52) showed relatively
high frequencies among parents of febrile convul-sion cases, as compared with normal controls.
Haplotype frequency and linkage
disequi-librium
Haplotypes with significant linkage disequilib-rium in the different seizure disorders are listed in Tables 6, 7 and 8. A statisitcally significant high frequency and significant positive linkage disequi-librium of haplotype HLA AI1; Bw6 were noted in both patients (x2=18.906, p<O.OO05, D/1000==180) and parents ix2==21.737, p<O.OO05, d/1000==156),
Table 4 Distribution ・of HLA-DR antigens and gene frequencies in different types of seizure disorders Patients Controls
HLA
n af(O/o) f(%)RR
X2 n af(%) f(%) DRI 2 9.1 4.7 4.45 2.86 3 2,2Ll
DR2 4 18.2 '9.6 O.88 O.05 27 20.1 10,6 DR3 4 18.2 9.6 1.13 O.04 22 16,4 8.6DRw4
3 13.6 7,O 1.35 O.20 14 10.4 5,3 DR5 le 45.5 26,2 O.53 1.93 82 61.2 37,7DRw6
oT
"
m
O.50 3 2.2Ll
DR7 l 4.5 2.3 .O,29 1.57 19 14.2 7.4DR2+DQw1
o-
-
-
O.50 3 2.2Ll
MT2
2 9.1 4.7 6.6 1,89 2 1.5 O.8MT3
o・rr
-
-
O.17 1 O.8 O.4af1antigen frequency, gf:gene frequency,
RR : relative riskL
Table 5 Distribution and significance of HLA antigen frequencies and deviating RR values disorders in different seizure Significantly highfrequency (p<O.OOI)
RR
value Relatively highfrequency (p<O.Ol-O,O05)RR・
value Significantly lowfrequency (p<O.OO1)RR
value Relatively lowfrequency (p<O.Ol-O.O05)RR
value IdiopathicPatients Bw21 12.30 generalized (x2=7.626) seizuresParents Bw6 2.76 B5 1.95Cw3
O.39 (x2==23.563) (x2==7.276) (x2=15.726)SimplePatients.
Bw22 7,84 Cw3 2.92 partial.selzures (x2:=14.803) Cw7 18.32 (x2=7.457) (x2=35.602)Parents Cw7 13,46 Cw3 2.13 All O.37
(x2==33.133) (x2=6.695) (x2-10.366) B15 O.ll (x2=6.697)
FebrilePatients
Cw7 7,23 A2 2.70 convulsions (x2=10.997) (x2=10.402) Parents Cw7 9,65 AIO 17.15 (x2=26.618) (x2=6.95) A28 4.10 ' (x2=7.140) Bw22 3.62 (x2=7.03!) Bw73 11.52 (x2=8.765) RR : relative risk.and a relatively high frequency and significant
positive linkage disequilibrium of HLA A2;B5
(x2=7.736, p<O.Ol, D/1000=96) were found in
parents of idiopathic generalized seizure cases,
which were comparable to those of normal
con-trols (Tables 6 and 9).
' HLA A9; Bw6 w'as found to show a statistically significant high frequency with a significant
posi-tive linkage disequilibrium ix2=12.371, p<O.OOI, D/1000=74) in parents of simple partial seizure cases, and a relatively high frequency and signifi-cant positive linkage disequilibrium of HLA AI;
B17 (x2==9.991, p<O.O05, D/1000=57), A2; Bw22
(x2=6.808, p<O.Ol, D/1000==66) and A26; B35
(x2==7.348, p<O.Ol, D/1000=96) were found in
patients with simple partial seizures, as compared
-Table 6 Distribution of HLA-A B haplotype frequencies and linkage disequilibria in generalized seizures
idiopathic Patients(n=74) Parents (n=122) Controls(fi=314) Haplotypes
n hfllOOO DllOOO X2 n hf/1000 D/1000 X2 n hf/1000
AllBw4
oT
+
O.067 o-
L
O.391 1 3AliBw6
1 14 13 O.743 1 8 6 2.580 o-AliB13
oL
-
O.067 e-
-
O.390 1 3Al;B17
0'
+
-
oL
-
-
o-AilB35
o-
-
O,067 1 8 8 O.481 1 3Al;B40
o-
-
O.067 o-
-
O.390 1 3A2iBw4
5 68 51 1.487 8 66 46 O.428 21 67A2;B5
8 108 62 2.800 17 139 96 7.736 17 54A2IBw6
5 68 -1 1.043 18 148 70 4.206 24 76A2:B7
o-
-
O.067 1 8 3 O.481 1 3A2;B8
o-
-
O.067 oL
-
O.390 1 3A2iB12
1 14 12 O.991 1 8 6 O.041 2 6A2lB13
2 27 2 O.035 3 25 4 2.578 22 7A2iB14
1 14 9 1.914 2 16 12 2,242 1 3A2IB15
6 81 61 O.744 6 49 32 O.038 15 48A2;B17
7 95 57 O.142 13 107 62 1.293 23 73A2;B21
or
-
o.e67 oL
T
O.390 1 3A2lBw22
on
-
O.179 o"
rm 1.172 3 10A2;B27
or
-
e.523 oL
"
3.571 9 29A2iB35
2 27 10 O.252 4 33 19 O.O14 11 35A2]B37
o-
-
O,067 om
rm O.390 ! 3A2:B40
11 l49 71 O.197 14 115 57 2.642 56 178A2iBw51
o-
-
-
o-
-
m
om
A2;Bw56
or
v
O.112 o-
-
O.786 2 6A2iBw63
or
-
O.067 oL
-
O.390 1 3A2lBw73
on
-
L
o+
T
-
o-A3iB5
on
-
O.067 o"
-
O.390 1 3A3;Bw6
o-
-
O,067 2 16 13 2.242 1 3A3IBw22
2 27 27 4.096 2 16T
2.202 o-A3;B40
or
-
L
o-
16-
oT
A9lBw4
1 14 6 O.175 2 16 5 O.152 5 16A9;B5
3 41 19 2.042 5 41 17 O,738 8 25A9lBw6
1 14 -19 O.313 3 25 -18 O.784 4 13A9iB7
o'
-
0.236 o-
-
1,575 4 13A91B12・
o em-
O.236 1 8 7 O.163 4 13A9lB13
4 54 42 O.810 4 33 21 O.182 8 25A91B15
1 14 5 O.041 2 l6 6 O.153 7 22A9lB17
1 i4 -4 O.092 5 41 16 O.033 14 45A9lBw22
1 14 9 l.914 oL
L
o.3ge 1 3A9iB27
o-
.
O.284 o-
m
1.972 5 l6A9:B35
2 27 19 O.819 3 25 17 2,574 2 6A9iB40
6 81 44 1.672 10 82 50 O.125 26 83A9;Bw44
o-
-
O.112 eN
-
O,041 2 86A9iBw56
o-
m
O.236 o-
-
1.575 4 13A9;Bw60
o-
-
O.067 l 8 7 O.481 1 3A9;Bw73
o-
-
L
2 16 15 2.202 o-AIOlBw6
o"
-
O,067 om
'
O.390 1 3 AIOiB13 o-
L
-
2 16 15 2.202 o-AIO:B17 1 14 13 O.743 1 8 7 O.291 o ..."
AlliBw4
1 14 2 O.062 2 16-3
1.653 13 41-AlliB5
5 68 26 3.231 10 82- 41 2.366 14 45AlllBw6
18 243 180 18.906 28 230 l56 21.737 21 67AllIB7
om
-
O.401 1 8 3 O.972 7 22AllIB8
1 14 12 O.313 1 8 7 O.I63 4 13AlliB12 o
-T
O.112 o-
-
O.786 2 6A!1iB13 3 41 19 0.115 8 66 46 O.428 21 67
AlllB14 o
-
-
O.067 1 8 4 O.481 1 3AlllB15 1 14 -4 O.635 4 33 17 3.433 26 83
All;B17 3 41 7 O.343 7 57 15 O.621 17 54
All;B18 1 14 12 O.914 1 8 7 O.481 1 3
AlliBw21
o-
-
O.067 om
-
O.390 1 3All;Bw22
2 27 18 1,888 2 16 7 2.242 1 3All1B27 2 27 21 O.O12 2 16 14 2.172 17 54
AlllB35 2 27 12 O.122 3 25. 11 O.130 6 19
AlliB40 10 135 64 O.551 16 131 76 3.586 68 217
AlliBw44
o-
-
O.067 oT
-
O.390 1 3AlliBw51
o-
rm.t.-
o-
-
-
o-AlliBw56, o
-
-
O.067 om
+
O.390 1 3A19;Bw4
o+
-
O.112 o"
m
O.786 2 6A19;B5
o-
m
O.236 1 8 4 O.163 4 13A19;Bw6
1 14 7 O.551 1 8 -O.2 O.024 3 10A19;B7
o-
-
O.179 o-
-
1.172 3 10A19;B13 1 14 12 O.551 3 25 23 1.462 3 10
A19lB17 1 14 10 O.551 1 8 3 O.024 3 10
A19iBw22
o-
rmT
o-
-
-
or
A19iB27 o
-
-
O.067 or
-
O.390 1 3A19iB40 3 41 34 1.734 3 25 19 1.467 3 10
A19lBw55
1 14 14 O.743 1 8 8 2.580 o rmA23;B17 o
-
-
O.067 e -- in O.390 1 3A24;B17 o
-
-
T
o-
-
-
o rrA25iB5
1 14 13 1.914 oT
nt O.390 4 13A261Bw4
1 14 12 1.914 1 8 6 O,481 1 3A26IB5
o-
-
e.o67 o-
-
O.390 1 3A26iBw6
o-
-
O.112 o-
-
O.786 2 6A26iB7
o-
-
O.236 o-
-
1.575 4 13 A26iB13 o-
-
-
o-
-
-
o wu A26iB17 2 27 23 1L992 3 25 22 O.783 1 3A26lBw22
e"
N
O.067 om
-
O.39e 1 3A26iB27 o
-
-
O.067 o-
-
O.390 1 3A26lB35 1 14 12 O.551 2 16 15 O.362 3 10
A26lB40 o
-
-
O.179 1 8 3 O.024 3 10A26;Bw55
o-
-. O.067 o rm-
O.391 1 3A26;Bw56
o im-
O.067 o-
-
O.391 1 3'
A26;Bw73
o-
-
O.I12 o-
m
O.786 2 6A28;Bw6
om
m
-
o-
-
-
o,-A28iB7
o-
-
-
o-
-
-
o-A281B13 o
-
-
O.067 o-
-
O.390 I 3 A281B17 om
h
O,067 o-
-
O.390 1 3 A28lB40 o-
um e.112 oN
-
O.786 2 6・A281Bw49
o-
-
O.067 o-
m
O.390 1 3A32lB40 1 14 IO.9 e.743 o
-
thm
o-hf/1000 : haplotype frequencies per thousand, D/1000:delta values per thousand.
-E196-Table 7 Distribution of HLA-A-B haplotype frequencies and linkage disequilibria in simple partial selzures
Patients(n=33) Parents (n=57) Controls(n:=314)
Haplotypes n hf/1000 D!1000 X2 n hf/loee D/1000 X2 n hf!1000
Al1Bw4
o.
-
e.113 om
L
O.182 ! 3Al:Bw6
o-
-
-
o-
rm rm om
AllB13
o-
L
O.113 o-
-
O.182 1 3AliB17
2 61 57 9.991 2 36 33 5.435 o-Al:B35
o.
-
O.113 om
-
O.182 1 3Al;B40
o--
O.113 em
-
O.182 1 3A2iBw4
1 30 25 O.675 2 36 15 O.844 21 67A2lB5
3 91 71 O.743 4 71 42 O.236 17 54A21Bw6
1 30 -6 O.952 4 71 -10 O.031 24 76A2iB7
1 30 20 O.562 2 36 15 2.780 1 3A2lB8
o-
-
O.113 o-
m
O.182 1 3A21B12
1 30 o O.169 1 18 11 O.752 2 6A2lB13
2 61 19 O.044 3 54 l4 O.233 22 70A2;B14
o-
rm O.562 or
-
O.182 1 3A2lB15
o-
m
1.653 om
n
2.840 15 48A2iB17
2 61 19 O.075 2 36 -24 1.125 23 73A2IB21
o un-
O.113 o-
T
O.182 1 3A2lBw22
3 91 66 6.808 4 71 57 6.334 3 10A2lB27
1 30 10 O.023 2 36 18 O.074 9 29A2iB35
om
7
1.194 2 36 15 O.053 11 35A2;B37
o-
-
O.I13 o-
-
O.182 1 3A2;B40
5 152 68 O.020 5 89 -10 2.884 56 178A2lBw51
o-
-
rm o-
rmL
o-A2IBw56
o-
-
O.215 o-
-
O.372 2 6A2lBw63
o-
-
O.113 o-
-
O.182 1 3A2iBw73
or
L
-
1 18 11 e.963 o-A3;B5
om
-
O.113 oL
-
e.182 1 3A3lBw6
o"
ne O.113 om
-
O.182 1 3A3lBw22
o--
rm o-
-
-
o-A3iB40
1 30 13 2.058 1 18 i6 O.963 oT
A9:Bw4
0T
T
O.OOI o-
-
O.923 5 16A9lB5
o-
"
O.531 1 18 6 O.134 8 25A9iBw6
o-
-
O.028 6 107 74 12.371 4 13A9iB7
o-
-
O.028 o-
-
O.735 4 13A91B12
o-
-
O.028 1 18 15 O.083 4 13A9lB13
oL
-
o.Ieo o-
-
1.480 8 25A9IB15
oL
-
O.752 o--
1.301 7 22A9:B17
2 61 42 e.173 1 18 -7 O.912 14 45A9lBw22
1 30 18 O.562 1 18 12 1.861 1 3A9;B27
2 61 52 1.161 2 36 29 O.962 5 16A9lB35
1 30 21 1.802 1 18 9 O.752 2 6A9IB40
5 155 117 l.733 5 89 48 O.025 26 83A9lBw44
o-
-
O.215 o-
m
O.372 2 6A9lBw56
o rm rr O.028 o-
-
O.735 4 13A9IBw60
o-
-
O.113 eT
-
O.!82 1 3A9iBw73
o-
-
m
1 18 15 O.963 o"
AIO;Bw6
1 30 28 e.562 1 18 16 1.861 1 3 AIO:B13 o-
-
-
o-
m
-
o-AIO1B17 o
m
r
rm o-
-
-
o-All;Bw4
o-
-
1.422 1 18 10 O.766 13 41-E197-AlliB5
o 30 18 O.153 3 54 43 O.071 14 45AlllBw6
1 121 100 1.322 4 71 40 O.O14 21 67'All;B7 o pt
m
.O.752 1' 18 10 O.Q57 7 22AllIB8
o-
-
O.028 o-
-
e.735 4 13 AlllB12 or
7
O.215 o rr-
O.372 2 6AlliB13 1 61 37 O.029 2 36 20 O.844 21 67
AlliB!4 o
r
+
O.113 on
-
O.182 1 3AlliB15 o
-
-
2.950 1 18 17 1.405 26. 83.All1B17 o 30 6 O.344 1 18 -5 O.186 17 54
AllIB18 o
"
-
O.113 om
-
O.182 1 3All;Bw21
o-
-
O.113・ 0L
-
O.182 1 3AllIBw22
o-
"
O,113 o.-
m
O.182 1 3AlliB27 o
-
r
1.887 1 l8 11 1.404 17 54AlliB35 o pt un O.644 o
-
-
1.115 6 19All・]B40 4 121 73 1.634 4 71 33 5.704 68 217
AlllBw44
o .tnff-
O.113 o-
-
O.182 1 3AlllBw51
1 30 24 1.913 1 18 15 O.963 o-AlllBw56
o-
-
O.113 o"
-
O.182 1 3A19lBw4
o-
u
O.215 o-
-
O.372 2 6A19iB5
o'
rm O.028 oL
-
O.735 4 13A19lBw6
o-
-
O.322 o-
-
O.553 3 ioA19lB7
o-
-
O.322 1 18 17 O,297 3 10A19lB13 1 30 28 1,135 1 18- 16 O,297 3 10 A19lB17 o
-
'
O.322 o-
ua O.553 3 10A19iBw22
o-
-
L
o-
ne=
o -A19lB27 o"
-
O,113 om
-
O.182 1 3A19lB40 o
'
-
O.322 1 18 14 O,297 3 10A19;Bw55
o-
-
L
o-
-
`
e-A23lB17 o
-
-
O,113 o-
-
O.182 1 3A24iB17 o
-
m
-
o-
-
-
o-A25IB5
o"
-
O.113 om
-
O.182 1 3A26lBw4
o-
-
O.113 1 18 13 1,865 1 3A26IB5
o-
T
0.113 o-
rm O.182 1 3A26lBw6
o-
rm O.215 1 18 -1 O.752 2 6A26iB7
1 30 28 O.651 2 36 31 O.963 4 13A26IB13 1 30 20 1.913 1 18 9 O.963 o
-A26lB17 o-
L
O.028 1 18 4 O,083 4 13A26iBw22
o-
rm O,113 oT
-
O.182 1 3 A26IB27 ov
-
O.113 o-
-
O.182 1 3 A261B35 3 91 96 7.348 3 54 49 3.257 3 10 A261B40 o-
-
O.322 o-
T
O.553 3 10A26iBw55
o-
-
O;113 oT
-
O.182 1 3A26iBw56
o-
-
O.113 e-
m
O.182 1 3A26iBw73
o"
-
O.215 om
-
O.372 2 6A28:Bw6
o-
-
-
o-
-
r
oT
A281B7
o-
-
m
o-
r
-
o -A28;B13 1 30 28 0.562 1 18 16 1.865 1 3 A28lB17 o-
mt O.113 1 18 14 1.865 1 3 A28lB40 o-
=
O.215 o-
m
O.372 2 6A28iBw49
o an-
O.113 o rm-
O.182 1 3A32lB40 o
'
-
-
o rm-
m
om
hfllOOO:haplotype frequencies per thousand, D/1000:delta values per thousand.
-El98-Table 8 Distribution of HLA-A-B haplotype frequencies and linkage convulsions
disequilibria in febrile
Patients(n=62) Parents (n=105) Controls (n=314) Haplotypes
n hf/IOOO D/1000 X2 n hf/1000 D/1000 X2 n hf/1000
Al;Bw4
o-
-
O.862 oL
"
O.341 1 3AliBw6
o-
r
-
oN
-
-
o-Al;B13
0-
m
O.862 o-
-
O,341 1 3AllB17
o"
-
-
o-
-
N
oT
AllB35
om
L
O.862 o-
-
O.341 1 3AliB40
o-
L
O.862 o-
r
O.341 1 3A21Bw4
11 177 116 O.131 13 124 8 3.423 21 67A2iB5
o-
L
2.369 3 29 13 1.135 17 54A2iBw6
6 97 27 O.292 10 95 49 O.372 24 76A2iB7
o-
L
O.862 1 10 -2 O.677 1 3A2lB8
o-
-
O.862 om
-
O.341 1 3A2lB12
o-
T
O,106 1 10 6 O.115 2 6A2lB13
4 65 29 O.022 5 48 16 O.663 22 7A2;B14
1 16 8 1.644 1 10 6 O,677 1 3A2;B15
1 16 5 1.273 2 19 IO 1.673 15 48A2:BI7
4 65 12 O,431 5 48 16 O,832 23 73A2;B21
o-T
O.862 om
-
O.341 1 3A2lBw22
1 16 O.3 O.215 o-
-
1.015 3 10A2iB27
2 32 8 O.022 10 95 72 6.597 9 29A2iB35
1 16 12 O.602 1 10 5 1.846 11 35A2iB37
o"
-
O.862 o-
-
O,341 1 3A2iB4e
14 226 70 O.773 19 181 89 O.025 56 178A2iBw51
1 16-
O,862 2 19 15 2.673 o-A2lBw56
o-
`
O.106 om
m
O.671 2 6A2iBw63
o-T
O.862 o rm-
O.341 1 3A2lBw73
1 16 12 O.862 1 10 3 3.004 o uzA31B5
o"
-
O.862 o-
-
O.341 1 3A3lBw6
o-
-
O.862 o-
-
O,341 1 3A31Bw22
o-
L
-
o-
T
m
o rmA3lB40
1 16 11 O.862 1 10 5 3.004 o-A9iBw4
1 16 -2 O.068 4 38' 24 1.844 5 16A9iB5
o--
1.611 oT
n
O.733 8 25A9iBw6
1 16 -5 O.053 2 19 3 O.224 4 13A9:B7
o--
O.823 1 10 6 O.076 4 13A9;B12
o-"
O.823 o-
-
1.351 4 13A9lB13
2 32 22 O.092 2 19 8 O.144 8 25A9iB15
2 32 29 O.225 2 19 16 O.043 7 22A9iB17
4 65 50 O.451 4 38 27 O,082 14 45A9iBw22
o-
-
O.862 o-
-
O.341 1 3A9]B27
1 16 9 O,068 1 10 2 O,232 5 16A91B35
o-
L
O.453 o-
-
O.671 2 6A91B40
6 97 91 O.134 11 le5 72 O.475 26 83A9:Bw44
o-
-
O.453 o-
-
O.671 2 86A9iBw56
o"
-
O.823 er
-
1.35I 4 13A9IBw60
o-
7
O,862 o7
m
O.341 1 3A9iBw73
o-
-
-
2 19 17 2.673 o-AIOlBw6
i 16 15 1.644 3 29 27 3.028 1 3 AIOiB13 o-
-
-
om
-
-
o-AIOlB17 1 16 16 Or862 2 19 18 2.673 o
-AlliBw4
3 48 21 O,066 3 29 8 O.352 l3 41-E199-AlllB5
o nt -・ 2.873 5 48 40 O.022 14 45All;Bw6
7 113 84 1.592 11 105 81 1.654 21 67AlliB7
1 1・6 14 O.092 2 19 13 O.045 7 22All;B8
o-
m
O.823 o-
-
1.351 4 13 All;B12 o-
-
O.453 1 10 8 O.115 2 6 All1B13 2 32 17 1.085 5 48 32 O.502 21 17 AllIB14 1 16 13 1.644 1 10 8 O.677 1 3 AlllB15 2 32 27 1.922 2 19 15 5.133 26 83 All;B17 ・1 16 -7 O.915 2 19 2 2.242 l7 54 All;B18 o-
1 O.862 o-
L
O.341 1 3AlliBw21
o-
-
O.862 o.m
-
O.341 1 3AlliBw22
1 16 9 1.644 1 10 5 O.677 l 3AlllB27 2 32 22 O.526 3 29 27 1.133 17 54
All;B35 1 16 14 O.031 1 10 7 O.447 6 19
All;B40 7 113 45 2.868 10 95 46 6.870 68 217
AlliBw44
'1 16 14 1.644 1 10 9 O.670 1 3AlliBw51
o-
-
-
o rm"
-
o-AlllBw56
o-
-
O.862 o-
-
O.341 1 3A19lBw4
2 32 26 3.303 2 19 16 1.348 2 6A19iB5
1 16 15 O,052 1 10 9 O.077 4 13A19lBw6
o-
-
O.603 o-
-
1.015 3 10A191B7
o-
-
O.603 1 10 9 O.333 3 IOA19iB13 2 32 29 2.o3e 5 48 46 4.247 3 le
A19lB17 1 16 10 O.215 1 10 7 O.333 3 le
A19;Bw22
1 16 15, O.862 1 10 9 3.004 or
A19IB27 o rm
-
O.862 o-
-
O.341 1 3A19lB40 o
-
-
O,603 o-
-
1.015 3 10A19IBw55
o-
m
-
o-
-
-
oT
A23lBl7 o
-
-
O.862 om
-
O.341 l 3・A24lB17 o
-
m
m
1-
-
3.004 o-A25iB5
o-
-
O.862 om
-
O.341 1 3A26IBw4
1 16 4 1.644 2 19 11 1.007 1 3A26iB5
1 l6 14 1,644 1 10 7 O.671 1 3A26;Bw6
om
-
Q.453 1 10 9 O.115 2 6A26iB7
oN
-
O.823 oT
-
1.350 4 13 A26lB13 o-
-
-
o-
-
-
0-A26;B17 o
-
-
O.823 1 10 8 O.076 4 l3A26lBw22
om
-
O.862 o-
m
O.341 1 3 A26iB27 3 48 43 6.209 3 29 25 3,028 1 3 A26iB35 o-
-
O.603 oN
-
1.015 3 10 A26lB40 3 48 18 2,803 3 29 11 2.025 3 10A26iBw55
o-
-
O.862 o-
-
O.341 1 3A26lBw56
o rm-
e.862 o-
rm O.341 1 3A26iBw73
1 16 15 O.622 1 10 9 O.115 2 6A28iBw6
oN
-
-
o rr-
m
o-A28lB7
o-
-
-
o-
-
-
o rrA28;B13 o un nt O,862 o
-
r
O.341 1 3A28iB17 o
-
-
O.862 o-
-
O.341 1 3A28lB40 2 32 27 3.303 1 19 10 e.I15 2 6
A28;Bw49
o-
L
O.862 o-
.-
O.341 1 3 A32iB40 o-
-
-
o-
-
-
o-hfllOOO : haplotype frequencies per thousand, D/1000:delta values per thousand.
-E200-Table 9 Distribution and significance of haplotype disorders
frequencies and linkage disequilibria in different seizure
Significantly highfrequency (p<O.OOI) DllOOO Relatively highfrequency (p<O.O1-O.O05) DllOOO Significantly lowfrequency (p<O.OOI) DllOOO Relatively lowfrequency (p<O.O1-O.O05) D/1000 IdiopathicPatients
AlllBw6
180 generalized (x2=18.906)seizuresParents
AlliBw6
156A2lB5
96(x2=21.737) (x2=7,736)
SimplePatients
AliB17
57 partial (x2=9.991) .selzuresA2lBw22
66 (x2=6.808) A26lB35 96 (x2=7.348) ParentsA9iBw6
74 (x2=12.371) FebrilePatients convulsionsParents AlliB40 46 (x2=6.870)DllOOO:delta values per thousand.
Table 10 HLA-A ; B haplotype analysis in several families with different types of
disorders, in both patients and their siblings
seizure Patients Siblings Fathers Mothers
FamilyA Idiopathic g,ei",e.r,a.l,iZedFamiiyB
A9IB5iCw3
A2iB17
A2iBw4lCw3
A9iB40iCw7
A9lB5iCw3
A2lB17
A2iBw4iCw3
A9lB40iCw7
A2lB17
A2lB5iCw3
A9lB40lCw7
A9iBw73;Cw3
A9lB5lCw3
A26IB40 A2;BwhT4lCw3A2;BI7lCw3
SimpleFamilyC
partial.selzures
A2lB401Cw7
A26lB13lCw3
A2iB40lCw7
A26lB13iCw3
A21Bw6lCw7
A26lB13lCw3
A2lB40iCw7
A26lB7lCw3
FamilyD Febrile convulsionsFamilyE
A9lB40iCw3
AlllB35 A26lB27A30+311Bw22
A9lB40lCw3
AllIB35 A26lB27A30+31;Bw22
A91B40iCw3
A9lBw6
A30+3IlBw22
A9lB4e
AlliB35A3lB40
A26lB27 Bw22with normal controls (Tables 7 and 9).
Only a relatively low frequency and significant linkage disequilibrium of HLA AII; B40 (x2=6.870,
p<O.Ol, D/1000=46) were found in parents of
febrile convulsion cases (Tables 8 and 9).
Discussion
Although the influence of inheritance in the
etiology of seizure disorders has long been
recQg-nized and has been an issue of debate for many
years, our understanding of the genetic basis has been slow in developing and its relative roles in predisposition to or causation of various types of seizures have not been clearly established. A part of the prQblem probably arises from differences in
populations, seizure definitions, measurement
data, and analytical methods, but the complexity of the nervous system, especially in its
devel-opmental aspect, has also presented formidable
obstacles. However, from the many genetic
studies of epilepsy reviewed by Newmark and
Penry38), several important trends are apparent.
First, generalized seizure disorders (absence,
generalized tonic-clonic, and photic-induced
sei-zures) most often have been associated with a
family history of epilepsy. Benign Rolandic epi-lepsy, which is usually partial, and febrile sei-zures, which may be partial or generalized, are the only potentially inheritable partial seizure
dis-orders. Second, although many metabolic and
inherited disorders are associated with epilepsy,
no single metabolic abnormality predominates; E201
-epilepsy may arise via many mechanisms. Third, the most closely related family members are at
highest risk of inheriting epilepsy. The highest
rates of epilepsy (60-80%) in relatives of an
epileptic proband are found in monozygotic
twins7)8). Nevertheless, it seems that some
evi-dence for a strong genetic contribution to the manifestation of epiiepsy has been provided by
twin and EEG studiesi).
The genes controlling HLA are located on the
short arm of the 6th chromosome. The HLA
system consists of A, B, C, D and DR loci, which are responsible for extreme polymorphism in in-heritance, and serve as good genetic markers for research. More than 300,OOO,OOO genetically dif-ferent individuals can be found on the basis of the known alleles of the above mentioned loci and, of theSe, more than 30,OOO,OOO have distinguishable combinations of antigens39}. The frequencies of the main alleles at each of the HLA-A, -B, -C, and
-D (and-DR) loci in representative populations vary among different races. Antigens which are
frequent in one population may be absent or rare in another40). Furthermore, antisera which have
been well characterized in one population may
show unexplained extra reactivity, or a lack of reactivity, in other populations4i}. As shown in Tables 1, 2 and 3, the frequencies of HLA-A2, -Al1
and -B40 were higher, and those of HLA -AIO, Aw23, -Bw29, -Bw49 and -Bw55 were very low in the Chinese population examined in this study.
These results were very similar to those reports of Chen et al.`2) in Taiwan, and Fujii et al`3) inJapan, but markedly different from the two earlier re-ports44}45) of normal Taiwan Chinese. A difference in the HLA profile for the same area may in part be due to differences in the antisera used. HLA typing is used primarily for determination of HLA compatibility prior to transplantation, for paternity testing, for anthrbpological studies, and for establi.shing HLA-disease associations. Dis-eases associated with HLA antigens have several characteristics. In general, these diseases (1) have
an unknown cause and unknown
pathophysiolog-ic mechanism, with a hereditary pattern of
dis-tribution but weak penetrance; (2) are associated
with immunologic abnormalities; and (3) have
little or no effect on reproduction`6). However, several hypotheses have been advanced to explain HLA-disease associations. Three of these appear to be most likely. The first hypothesis holds that
HLA antigens are merely markers for irnmurte
response genes or immune suppress'ive genes. The immunologic response (or lack thereof) to an agent may indicate a predisposition to a disease. The
second hypothesis suggests that HLA antigens
may act as receptors for etiologic agents. If par-ticular HLA antigens act as receptors for viruses,
toxins or other foreign substances, and these substances are the etiologic agents for given diseases, then HLA-disease associations would
result. The third hypothesis is that of molecular mimicry. It postulates that a disease-associated HLA antigen is structurally and immunologically similar to the etiologic agent for the disease and further postulates one of two alternatives. The first alternative is that because of the similarity between the etiologic agent and the HLA antigen, no immune response is elicited, and therefore the etiologic agent produces the disease without any interference. The second alternative suggests that a vigorous immune response is elicited against the etiologic agent. Because of the similarity of the
agent and the HLA antigen, the immune response
is turned against the.HLA antigen, a・nd this
"autoimmuner' response then produces the
dis-ease46). As HLA antigens trigger the immune
defense mechanism, several diseases such as an-kylosing spondylitis (with HLA-B27) and juvenile
diabetes mellitus (with HLA DR3 and DR4) have
been shown to have a very close association with
the HLA antigens. On the other hand, some
diseases are transmitted with a certain HLA
haplotype, such as idiopathic hemochromatosis
and 21-hydroxylase deficiency, which are two
examples that have been shown to be transmitted as recessive HLA-associated diseases47)48).
In the early 1970s the first HLA and disease
associations were described, among which was
multiple sclerosis49). Since then, numerous HLA
-and disease associations have been reported, as well as several studies of the genetic aspect of human epilepsy47)50)'"58), concentrating on a possi-ble association between HLA and epilepsy types. The first such attempts to be published were the studies of Smeraldi and co-workers (1975)50} on the association of the Lennox-Gastaut syndrome with
antigen HLA-B7, and of Smeraldi and co-workers
(1976)5i} on mixed lymphocyte culture (MLC) in 15 unrelated children with the same syndrome. Some of the several subsequent investigations concern-ed with different types of seizure disorders, such as primary generalized epilepsy, partial epilepsy, petit mal epilepsy, etc, showed a good correlation to certain HLA antigens. For instance,
signifi-cantly increased HLA-B7 antigen and HLA A3; B7 haplotype in the Lennox-Gastaut syndrome52);
increased frequency of Al; B8 haplotype, HLA-B5
and -B8 antigens, and decreased frequency of
HLA-A2 antigen in 3 C/S SW absence
epi-lepsy53)54); increased frequency of HLA-B5 antigen in focal seizures54); decreased frequency of HLA
AI; B8 haplotype in benign epilepsy of
child-hood47); increased frequency of HLA-A9 antigen in generalized tonic-clonic and absences55} and
in-creased frequency of HLA-DR5 antigen in grand mal primary generalized seizures have been
re-ported. In investigations concerning this topic by Eeg-Olofsson et a147) and Hafez et a155}, an antigen
HLA-A9 prevalence of 25--47% was found among
European patients with idiopathic epilepsy, which was comparable to that in the Chinese population (31.1%) in this study (Table 1).
However, Howitz and Platz57) and Monaghan et
al.58) found no differences in HLA antigens
be-tween controls and probands with infantile
spasms or cryptogenic myoclonic epilepsy. As shown in Table 5, a statistically significant
high frequency and deviating high RR value
(7.23"--18.32, positive association) of HLA-Cw7
antigen were noted in both patients and their
parents for simple partial seizures and febrile convulsions. For other antigens, HLA-AIO, -Bw21, and -Bw73, etc, showing relatively high
frequen-cies, there were relatively high RR values
(II.52N17.15). This indicates that the higher the relative risk (above 1), the more frequent is the antigen among the patient population, which can be regarded as the chance an individual with the disease-associated HLA antigen has of developing the disease compared to an individual who lacks that antigen46).
The statistically significant HLA-A; B haplo-types, A9; Bw6 and All; Bw6, with significantly high delta values (D/1000=74-180, Table 9) found in this study imply that these particular HLA-A; B haplotypes occur in the seizure populations with frequencies which are higher than expected from the gene frequencies of the alleles in the same populations.
In spite of the high HLA-A2, -AIO and -B40
antigen frequencies (Tables 1, 2 and 3), and high
A2; B40 and All; B40 haplotype frequencies
(Tables 6, 7 and 8), which ,were very compatible
with the most commonly appearing antigens
men-tioned above, found by normal population analysis
in this study (a high Aw19; B17 haplotype fre-quency in Chinese59), and high Aw24; Bw52 and
Aw33; Bw44 frequencies in Japanese`3} have pre-viously been reported), the statistically significant
HLA antigens, -Bw6, -Bw22, -Cw3 and -Cw7, and
haplotypes, Al1; Bw6 and A9; Bw6, were found in the different seizure disorders (Tables 5 and 9).
These specific HLA antigens, -Bw6, -Bw22, -Cw3
and -Cw7, hOwever, may reflect an association or
linkage between the HLA system and genetic
control, and seem to be characteristic of patients with seizure disorders in this area.
Similarly, the relatively lower frequency of
haplotype All; B40 in febrile convulsions, which
was the most common haplotype in normal
con-trols in this study (Tabies 8 and 9), must thus provide some kind of biological advantage. It is
possible that All; B40 carries protector genes
against certain noxious mechanisms, e.g., infec-tious agents. A lack of this haplotype thus could
explain a lowered immunologic defensive
mecha-nism, resulting in increased risk for the appear-ance of seizure disorders47).
The. reason for linkage disequilibrium is
-E203-clear. Several hypotheses have been proposed to
explain the phenomenon of linkage
disequili-brium, including (1) a selective advantage of a given haplotype, (2) migration and admiXture of two populations, (3) inbreeding, and (4) random
drift46).
Immunological abnormalities in human epilep-tics have been reported60). An abnormal T-cell
function in drug-treated patients6i} and low serum levels, both drug-induced and drug-independent, of IgA62) have been described. A predisposition, presumably genetically determined, seems to be a prerequisite for the development of drug-induced IgA deficiency. An increased risk for developing
IgA deficiency was reported among individuals
with antigen HLA-A263)64). In the present inves-tigation, however, there was no statistically signi-ficant difference between any of the seizure dis-orders and the controls, regarding the Occurrence of HLA-A2 antigen, except for a relatively high
frequency (x2=10.402, p<O.O05, RR=2.70, as
shown in Table 3) in patients with febrile con-vulsions.
Since there are two unique HLA-B specificities,
called Bw4 and Bw6, which do not correspond to
conventional alleles of the HLA-A or HLA-B anti-gens, and their specificity resides on unique epi-topes on the HLA-B molecule (and on a few HLA-A
molecules), either Bw4 or Bw6 is expressed
to-gether with the HLA-B private specificity65); there-fore, further evaluation of the results concerning the statistically significant high frequency of the
HLA-Bw6 antigen in seizure disorders among the
Chinese population is very necessary.
The HLA Cw3 antigen was rather elusive in
this study. Despite the relatively high frequency and positive disease association <high relative
risk), in both patients and parents for simple
partial seizures, this HLA antigen still showed a statistically significant low frequency and
nega-tive disease association in parents of primary
generalized seizure cases (Table 5).
In the few publications on HLA and epilepsy,
there are no data on investigations of antigens of
the HLA-DR locus. Since it is known that the
antigens of the HLA-DR locus are of greater
importance for the' immune response than are
those of the other loci of the HLA system, the
finding of a statistically significant increased
frequency of HLA-DR5 (x2=13.08, p<O.OOI) in
patients with grand mal primary generalized seizures by Minev et a156}. can be accepted as
offering hope. But, no significant HLA-DR antigen was found in this study (Table 4).
Family studies involving haplotype analysis
were per'formed on several cases with different seizure disorders, siblings having seizure histories similar to those of the probands. All siblings had
the same haplotype as the respective probands
(Table 10). This・ haplotype, correspondence may point an inhertied susceptibility to these seizure disorders.
Nevertheless, it may be concluded that the
difficulty in demonstrating simple Mendelian
inheritance of seizure disorders with modern
techniques still argues in favour of multifactorial
inheritance. Investigations involving HLA
typing in larger populations of epileptic individ-uals and their families may provide a more reliable
and conclusive answer regarding the genetic
characteristics of seizure disorders.
Acknowledgement
The author is grateful to Professor Y. Fukuyama, Head of the Department of Pediatrics, Tokyo Women's Medical College, Japan, for critically reviewing the manuscript. The author is also grateful to Professor Y.H. Ma, forrner Head of the Department of Pediatrics, Kaohysiung Medical College, for his advice and en-couragement, and to Professor WJ. Liu, Head of the Department of Pediatrics and Clinical Laboratory, and
Miss W. Chiang, instructor of the Departrrient of
Clini-cal Laboratory, for their immunologiClini-cal assistance, and to Dr. FJ. Ko, instructor of the Department of
Pedia-trics, for her help in the statistical data processing, in
Kaohsiung Medical College.
Referecnes,
1) Annegers JF, Hauser WA, Elvebaek LR et al:
Seizure disorders in offspring of parents with a history of seizures.-a maternal-paternal difference?, Epilepsia