DOI: http://dx.doi.org/10.14947/psychono.35.15
ヒトはなぜ
3Dを見ることができるのか?
―ヒト脳内背側視覚経路に沿った階層的な3D情報処理過程―
番 浩 志
a, b a 国立研究開発法人情報通信研究機構脳情報通信融合研究センター b 大阪大学大学院生命機能研究科How does human perceive 3D visual world?:
Hierarchical processing of 3D depth cues along the dorsal visual pathway
Hiroshi Ban
a, ba Center for Information and Neural Networks, National Institute of Information and Communications Technology b Graduate School of Frontier Biosciences, Osaka University
A fundamental challenge in visual psychology and neuroscience is to understand how 2D retinal inputs are re-constructed into a coherent and stable 3D visual world. In this review paper, I will introduce our recent fMRI studies on 3D depth perception in which we investigated 1) how and where in the brain multiple depth cues are integrat-ed, 2) the development of visual 3D cue integration at the level of neural representation, and 3) how the outputs of neurons responding to local and simple elements are progressively transformed to encode the critical features of spa-tially-extensive objects. Based on those studies, I will discuss on the roles of dorsal visual areas to infer the computa-tional hierarchy that supports the 3D estimation, a property important for recognizing our visual world and plan-ning actions. Furthermore, I would like to discuss how 3D vision psychology/neuroscience studies can contribute to the engineering fields to develop future display devices and information and communication technologies.
Keywords: 3D, depth, stereo vision, cue integration, fMRI
1. は じ め に ヒトは顔の左右水平に約6–7 cm離れた2つの眼を有す る。よって外界を両眼で観察した際には,眼と注視点と を結ぶ輻輳角の違いにより,左右の網膜にそれぞれ少し ずつ異なる像が入力される。この「網膜像差」は,観察 者の注視点と視物体との位置・距離に対応して様々に変 化するため,そのずれをうまく検出し,ずれと奥行きと の対応を計算することができれば,左右の網膜に投影さ れた2つの2次元の像から3D (3 Dimensional; 3次元)構 造を再構築することができる。すなわち,我々ヒトが豊 かで滑らかな3次元視覚世界を知覚できるのは,脳が左 右の2次元網膜像の対応問題を解いているからである。 3D 視の神経科学・心理学研究においては,特にこの 「両眼視差」手掛かりが注目され,電気生理学的な手法 やfMRI (functional magnetic resonance imaging; 磁気共鳴 影像法)などの脳イメージング手法を駆使し,脳のどの 部位のどのような活動によって視対象の3D構造が復元 され,知覚へと至るのかが重点的に研究されてきた (DeAngelis, Ohzawa, & Freeman, 1991; Ohzawa, DeAngelis,
& Freeman, 1990; Orban 2011; Preston, Li, Kourtzi, & Welch-man, 2008)。一方で,ヒトは両眼視差のみならず,陰影 や運動,テクスチャのきめ,物体同士の大小関係・重な り,パースペクティブといった手掛かりからも豊かな 3D 構造を知覚できることがわかっているが,それら 「単眼性」あるいは「絵画的」な視覚情報からヒトがど のように立体を再構成しているのかは研究が十分に進ん でいない。また,両眼視差手がかりと運動など,複数の 奥行き手掛かりが組み合わされて呈示された際に,ヒト Copyright 2016. The Japanese Psychonomic Society. All rights reserved. Correspondence address: Room 3A4, Center for
Informa-tion and Neural Networks (CiNet), NaInforma-tional Institute of In-formation and Communications Technology, 1–4 Yamada-oka, Suita, Osaka, 565–0871, Japan. E-mail: ban.hiroshi@ nict.go.jp
脳がそれら複数の手がかりをどのように統合しているの かに関しても明確な仕組みは明らかになっていなかっ た。 絵画的な奥行き手掛かりは,両眼視差のように計算で 明示的に奥行き構造(注視点からの距離)が求められる 手掛かりではないため(不良設定問題となる場合が多 い),3D視にそれほど大きな影響を及ぼすことはないと 思われるかもしれない。しかし,ヒトの両眼間の距離 (約6–7 cm)は視対象の奥行き方向の拡がりに対して非 常に小さく,この構造上の制約により,数十∼数百メー トル離れた物体に対して両眼視差から正確な3D構造を 検出することは非常に困難である。そのような遠くの視 対象の奥行きを検出する際には,単眼性の絵画的手掛か りが両眼視差以上に重要な情報として働くことがわかっ ている。ヒトが広大な自然風景を見た際に,滑らかで連 続した3次元空間を知覚できるのは,両眼・単眼両者の 手掛かりを脳がうまく繋ぎ合わせているからである。ま た,絵画的な奥行き手掛かりは知識や文脈の影響に依存 する。例えば,陰影手掛かりは,光は上方から物体を照 らす,という知識・経験と組み合わさって物体表面に豊 かな凹凸感を生み出す。また,陰影と両眼視差とが組み 合わさって構成された視物体の凹凸感は,両眼視差のみ で定義された凹凸感よりも精確に知覚されることがわ かっている。よって,3D (立体)感を生み出すヒト脳内 の奥行き情報処理機構を真に理解するためには,両眼手 掛かりのみではなく,絵画的掛かりやそれに付随する文 脈が脳のどの部位のどのような働きで奥行きへと変換さ れるのかを解明し,手掛かりの違いによる脳活動の共通 性・差異を捉えるための新しい研究パラダイムの構築が 必要である。 本稿では,基礎心理学会フォーラム「脳イメージング で探る3次元の世界」での筆者の講演をベースに,両眼 視差手がかりと絵画的な 3D手がかり(陰影,テクス チャ,運動など)がヒト脳内で統合される過程,その 3D手がかり統合機構の脳内発達過程,そしてヒト脳内 の背側視覚経路の階層性に沿って局所的な両眼視差情報 が物体の大局的な傾き平面へと変換されていく過程,を 調べた我々の最近のヒトfMRI研究について紹介したい。 紙面の都合で各研究結果の導出に用いられた方法の詳細 には触れないが,それぞれの研究が何を目指したのか, その背景をできる限りわかりやすく紹介するように努め た。少しでも3D視研究の面白さを伝えることができれ ば幸甚である。 2. ヒト脳内における複数の3D手掛かりの統合 ヒトが絵画的な単眼性手掛かりから立体感を知覚する 例として,上では陰影を挙げたが,今度は飛行機が離陸 する時に窓から外の風景を眺める場合をご想像いただき たい。滑走路や空港に立ち並ぶ建物など,観察者である 自身の眼前に位置する視対象は,自分(飛行機)の運動 方向とは逆向きに高速に遠ざかっていく一方で,遠くに 見える木々や山は,あたかも自分の動きに追従するかの ようにゆっくりと動きながら,次第に自分から遠ざかっ ていくかのように見えるだろう。このように,自己の運 動と注視点,そして視対象との距離によって,視対象の 運動方向・速度は様々に変化して知覚され,その違いか らもヒトは外界の豊かな3D構造を知覚できる(「運動視 差」手掛かりと呼ぶ)。 では,ヒト脳は単眼性の手掛かりをどのように3D構 造へと変換しているのだろうか。また,両眼視差と単眼 性の3D手掛かりとが同時に呈示された際,ヒト脳はそ れらの複数の手掛かりをどのように統合しているのだろ うか。この謎が解明されれば,例えば両眼視差手掛かり を用いた3D視が困難な人間に対し,絵画的手掛かりを 強調することで立体視をサポートするシステムを作るこ とができるかもしれない。同様にして,現行の3Dテレ ビは両眼視差手掛かりのみを用いているが,映像の中の 絵画的な3D手掛かり(陰影など)を強調するなどして, より豊かで臨場感 れる次世代の3Dテレビの開発も可 能となるかもしれない。逆に,詳細は省くが,両眼視差 手掛かりに変調を加えるだけの操作で,物体表面の光沢 感知覚が変化すること(Muryy, Welchman, Blake, & Flem-ing, 2013)や,そうした両眼視差手掛かりの変調から得 られる光沢感を処理する脳部位も報告されている(Sun et al., 2016)。ゆえに,両眼視差と2次元の奥行き手掛か りがどのように脳内で統合されているのかを調べること は,神経科学・心理学のみならず工学的な応用分野に とっても非常に重要な研究テーマである。 そこで我々のグループは,両眼視差手掛かりと運動視 差手掛かりのそれぞれがヒト脳内のどこで,どのように 統合されているのかを調べるために,fMRIを用いたヒ ト脳機能計測を行った(Ban, Preston, Meeson, & Welch-man, 2012)。3D手掛かりに限らず,ヒトは2つの視覚手 掛かりが同時に与えられた場合に,それらをうまく統合 して効果的(ここで「効果的」とは,手掛かりが複数同 時に与えられた場合に,単一の手掛かりを使う場合と比 べてより詳細な判別が可能になったり,判別速度が促進 されたりすることを指す)な処理ができることがわかっ
ている。この統合の際には,おおまかに2つの方略が考 えられる。これらの方略を奥行きの差異の判別(2つの 物体のうち,どちらの物体がより近くに置かれているの か,といった判別)に合わせて図示したものがFigure 1 である。この図において,横軸と縦軸は90度に直交(2 つの手掛かりが独立していることを示す)し,横軸は両 眼視差による奥行き(遠近)の判別センサの応答,縦軸 はもう1つの手掛かり,例えば運動視差による奥行きの 判別センサの応答を示す。ここで,各軸において遠近の 両センサの分散が小さい,あるいは距離が遠いほど奥行 き判別の精度は高くなり,その距離はd′で表すことが できる。このとき,2つの手掛かりが与えられ,それら が線形に統合される場合,その判別感度は,ピタゴラス の3角形の定理に基づき, ′ ′ ′ d d d 2 2 両眼視差+運動視差= 両眼視差+ 運動視差 (1) となる。心理学的な研究により,手掛かりの統合のいく つかはこの非常に単純なモデル(独立統合; independent integration)でうまく説明できることがわかっている。 一方で,もし2つの手掛かりが何らかの形で「融合」さ れた場合はどうなるだろうか。その場合,手掛かりが融 合された後の「融合判別センサ」の分散は,単一の手掛 かりを処理するセンサの分散よりも小さくなる。ここで はこれを「融合統合 (fused integration)」と呼ぶ。このと き,より精度の高い 3D構造の知覚が可能となるため, 融合統合は生物の生存上有利なメカニズムであるといえ る。では,ヒトの脳は複数の奥行き手掛かりが与えられ た際に,どこかでこの「融合統合」を行っているのだろ うか。それとも,ヒト脳は異なる手掛かりを独立に処理 し,「独立(線形)統合」をしているのだろうか?これ を調べる目的で,我々は被験者が1. 両眼視差のみで定 義される遠近2つの奥行き平面,2. 運動視差のみで定義 される遠近2つの奥行き平面,3. 両眼視差と運動視差の 2つの手掛かりで定義される遠近 2つの奥行き平面,の 3条件の視覚刺激を観察している際の脳活動を計測し, 得られた脳活動に対して SVM (Support Vector Machine) による機械学習の解析手法を適用することで,それぞれ
の呈示条件における 3D平面の遠近判別感度を求めた。
その結果を図示したものがFigure 2である。この図にお いて,横軸は各視覚野を示し,図中の横線は,両眼視 Figure 1. Linear integration and fusion model. Schematic illustration of linear and fusion cue integration models. The
差,運動視差それぞれが単一に呈示された場合に得られ た奥行き判別精度を基に算出した理論的な「独立統合」 モデルの精度である。この結果より,V1, V2, V3といっ た初期の処理段階では,2つの手掛かりが同時に呈示さ れた場合の奥行き判別感度は「独立統合」モデルでうま く予測できることがわかる。一方,視覚野V3B/KOにお いては,2つの手掛かりが同時に呈示された場合の奥行 き判別感度が,「独立統合」のモデルを上回っているこ とが明らかになった。 では,運動視差以外の2次元の絵画的な奥行き手掛か りはどこで融合されているのだろうか(あるいは,融合 されていないのだろうか)。これを調べる目的で,上述 の運動視差の融合研究と同じパラダイムを用いて,両眼 視差と陰影情報(Dovencioglu, Ban, Schofield, & Welchman, 2013),あるいはテクスチャのきめの勾配情報(Murphy, Ban, & Welchman, 2013)が同時に呈示された場合のfMRI 脳活動計測を行った。結果,興味深いことに,両眼視差 と対になる手掛かりが何であれ,それらはV3B/KOで融 合されていることが明らかになった。さらに重要なこと に,2つの手掛かりが同時に呈示された際のfMRIデータ からV3B/KOの活動を抽出し,その判別精度の上昇度合 いを見ることで,ヒトが実際に奥行きを判別する際のパ フォーマンスをうまく予測することが可能であることも 示された(Dovencioglu et al., 2013)。脳には視覚に関連 する多くの部位があるにもかかわらず,たった1つの小 さな領野の活動を調べるだけで,その個人の奥行き判別 パフォーマンスが予測できたことは,驚くべき結果であ る。この V3B/KOの活動をより正確に定式化できれば, 3D映像呈示装置の臨場感など,工学的なデバイスの評 価指標として利用できる可能性がある。 3. ヒト脳内における複数の 3D手掛かり機構の発達 ここで,次の疑問が生じる。V3B/KOにおける複数の 3D手掛かりを「融合」する働きは,ヒト脳が生得的に 獲得している機能なのであろうか。心理学的な発達研究 によると,乳児はすでに視覚手掛かりに隠された規則性 を発見する能力が備わっていることが知られている (Van de Walle & Spelke, 1996など)。一方で,手掛かりを 大人と同じように統合するには,経験に基づいた比較的 長い発達過程が必要であることが示されており,様々な 感覚統合タスクにおいて,その種類によらず子どもが最 適な統合を実現できるのは10∼12歳頃だということが 明らかになりつつある(Nardini, Bedford, & Mareschal, 2010; Nardini, Bequs, & Mareschal, 2013)。この手掛かり統合の 発達にかかる時間(言語的な概念を覚え始めるよりも時 Figure 2. Results for the quadratic summation test. The upper graphs show prediction performance for near versus far
dis-crimination. The lower graphs are the same data as an integration index. Here, zero indicates the minimum bound for fu-sion (that is, the prediction based on quadratic summation). The center of the bowtie represents the median, the shaded area depicts 68% confidence values, and the upper and lower error bars 95% confidence intervals. The graphs were modi-fied from Ban et al., 2012.
間がかかっている)は,子どもが日常生活を送るなかで 常に様々な視覚刺激を見続けていることを考えると,信 じられないほど長いものである。さらに,自然界を観察 する際に常に複数の視覚手掛かりが重なり合って呈示さ れる(例えば,両眼視差と線分の勾配など)ため,統合 を学ぶための十分な経験が得られているはずである。で は,なぜ視覚的な手掛かりの統合にこれほどの時間がか かるのだろうか。 この謎を解明するため,我々は心理行動実験とfMRI 脳機能計測を組み合わせた研究を行った(Dekker et al., 2015)。最初に,3D手掛かり統合にも同じような遅い発 達過程が見られるのかを調べるため,6∼12歳の144名 の子どもに協力してもらい,上で述べた両眼視差と運動 視差の統合実験と同じパラダイムを用いて,どの年齢層 の子どもから2つの手掛かりを「融合」する(注: ここ でいう「融合」とは,「線形(独立)統合」のモデルを 越える手掛かりの統合を意味する。10.5歳未満の子ども も線形統合は行っている)ことができるのかを調べた。 その結果,他のモダリティあるいは手掛かりの統合を調 べた先行研究と同じく,両眼視差と運動視差が融合でき るのも10.5歳以降の子どもからであることが示された。 ここで,10.5歳以降しか3D手掛かりが「融合」でき な い理 由 と し て,2 つの仮説が考えられる。1 つは, 我々が前節で紹介した V3B/KOにおける3D手掛かり融 合機能が未発達なため,融合自体ができていない可能性 である。もう 1つの仮説は,V3B/KOにおける手掛かり の融合は実現されているものの,それをうまく読み出す 能力が発達していないとする仮説である。これらの2つ
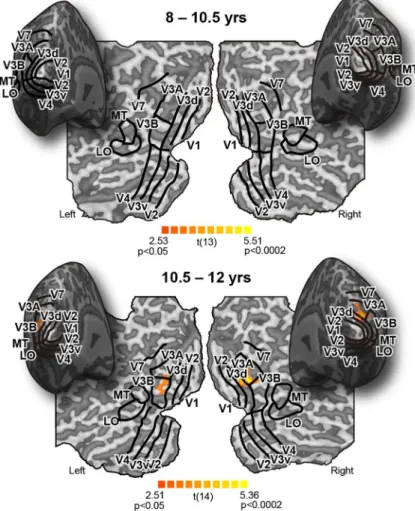
Figure 3. Searchlight results (accuracy for dual cue condition is higher than that for quadratic summation model). ROIs from two representative subjects are super-imposed on the group result inflated and flattened cortical sheets. Significant re-gions may therefore be slightly misaligned with respect to their labels. Areas where both integration indices were signifi-cantly above zero are colored. Modified from Dekker et al., 2015.
のうち,どちらの仮説が正しいのかを検証するため,行 動実験に参加した協力者から29名(8∼12歳)に再度協 力を依頼し,両眼視差と運動視差からなる奥行き刺激を 観察中に fMRIによる脳機能計測を行った。その結果, 非常に興味深いことに,V3Bにおける「融合統合」は 10.5 歳以下の子どもには全く観察されなかった。一方 で,10.5歳以降の子どもでは,大人の場合と同じく, V3B/KOにおいてのみ,2 つの手掛かりを「融合」する かのような活動が観察された(Figure 3)。これはすなわ ち,10.5歳未満の子どもが2つの3D手掛かりを「融合」 できないのは,脳の中で手掛かりが融合されていてもそ れを読み出すことができないからではなく,そもそも V3B/KOが大人のようには発達しておらず,手掛かりを 融合できていないことに起因することを意味する。 この結果は,子どもの学習方法の改善などの点からも 示唆に富むものである。繰り返しになるが,大人の場 合,複数の3D手掛かりが呈示された際,それらを融合 することにより,それぞれの手掛かりが単独で呈示さ れた場合の線形統合を越える 3D弁別能力を得られる。 すなわち,複数の手掛かりを同時に利用することで外界 の3D構造をより精確に知覚している。一方で,10.5歳 未満の子どもは手掛かりの融合ができないため,同じ視 対象を観察しても,大人ほどには精確に3D構造を知覚 することができない。よって,例えば野球などのスポー ツのトレーニングにおいて,大人と同じように「いろい ろな手掛かりを総合的に考慮して判断する」努力をする よう指示したとしても,複数の手掛かりをそもそも最適 に融合できないため,パフォーマンスは劣ってしまう。 これは,身体能力の差とはまた異なる子どもの能力の限 界を示すものである。特に10.5歳未満の子どもには年齢 にあった適切なトレーニング法を与えなければならない かもしれない。 4. ヒト脳内の背側視覚経路に沿った 階層的な3D情報処理過程 以上,ヒト脳内における3D手掛かりの統合過程を調 べた研究を紹介した。では,V3B/KOでの3D手掛かり の統合に至るまでの過程で,視野内に点在する両眼視差 情報はどのように物体の3D表面へと変換されているの だろうか。最後に,初期視覚野V1から中・高次の視覚 野 V3Aへの背側処理経路に沿って,局所的な両眼視差 情報が大局的な物体表面の傾きへと階層的に変換されて いく流れを可視化したヒトfMRI研究を紹介する(Ban & Welchman, 2015)。従来の3D視に関する脳イメージング
研究の多くは,ランダムドットステレオグラム(Ran-dom Dot Stereogram; RDS)法を用いて両眼視差手掛かり を付与することで,注視面から見て近く,あるいは遠く に「平面」を呈示し,その遠近に対する脳活動の違いを 調べてきた。しかしながら,単純な 3D「平面」刺激の 場合,RDS刺激に含まれるローカルなドット1つの奥行 きと刺激全体のグローバルな平面の奥行きが一致してい るため,観察者は数個のドットのみからターゲット刺激 が呈示された奥行き面を推測できてしまう。このため, 3D平面刺激はローカルな両眼視差情報の統合を調べる 目的には適切ではない。よって,本研究では RDSから なる様々な角度を持った傾き面をターゲット刺激として 用いた。傾き面からなる RDS刺激の場合,たとえ観察 者が一部のドットから奥行きを知覚できたとしても,刺 激全体に散らばるドット群の奥行き情報を統合できなけ れば傾き面の正しい角度を知覚することはできない。 実際の実験では,main条件として,Figure 4に示され
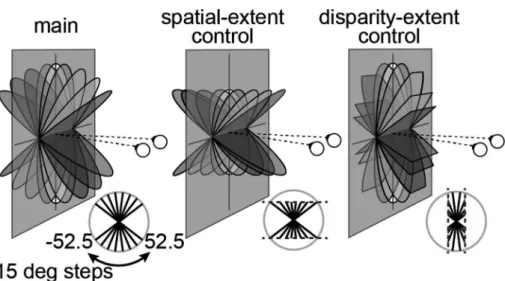
た8種類の傾き面刺激(−45 degから135 degまで15 deg
刻みで水平軸に沿って円盤を回転させたもの)を呈示し た。ここで,これらの8つの刺激は,傾きの大小によっ て両眼視差を有するドット群の網膜上での拡がりが異な る(ただし,背景にも RDS刺激を呈示しているので, 単眼では見分けがつかない)。この網膜像の違いが脳活 動に及ぼす影響を検証するため,spatial-extent control条 件として,ターゲット刺激の網膜像を統一した8種類の 刺激も用意した。さらに,main条件の刺激は,傾きの 大小によって刺激に含まれる両眼視差の最大値も大きく 異なる(ターゲット刺激が傾くほどに,刺激はより大き な両眼視差を有する)。傾きの違いではなく,刺激が有 する両眼視差の最大値の違いが及ぼす影響も検証するた め,disparity-extent control条件として,ターゲット刺激 の奥行き空間での拡がりを制限した刺激も8種類用意し た。この操作の後,spatial-extent, disparity-extentの両条 件にはそれぞれ 2つずつmain条件と同じ刺激が含まれ るため,条件数を減らし,合計で8種類の傾き×3条件 −4=20種類の刺激を被験者に呈示した際のfMRI脳活動 計測を行った。 解析には最近のfMRIデータ解析において主流になり つ つ あ る多 変 量 解 析 法 の 1 つ,MVP-similarity analysis (Multi Voxel Pattern similarity analysis; マルチボクセルパ
ターン類似度解析)を用いた(Kreigeskorte, et al., 2008)。 この解析手法は,V1, V2,といった各視覚野をROI (Re-gions Of Interest; 関心領域)として同定した後,そのROI 内のボクセル応答のパターンの相関を全条件間で計算 し,その相関値を類似度行列表として表現するものであ る。この類似度行列表を見れば,その ROI内の処理内
容,あるいはそのROI内でどのような視覚特徴が保持さ れているのかを捉えることが可能となる。例えば,今回 の研究の場合,ある視覚野(=ROI)でターゲット刺激 の傾き(角度)が表現・処理されているならば,その ROIの相関パターンは,spatial-extent条件やdisparity-ex-tent条件でコントロールした刺激の網膜像の拡がりや両 眼視差の大小に影響を受けず,傾き角度が同じ刺激間で は高い相関が観察されるはずである。一方,もしある視 覚野がローカルな網膜像の拡がりの違いのみを反映した 活動を示すならば,そのROIの相関パターンは,網膜像 の拡がりを統一したspatial-extent条件の刺激間では刺激 の傾き角度の違いによらず非常に高くなるはずである。 この解析を実際のデータに適用した結果がFigure 5で ある。実際には 10以上のROIを解析しているが,統計 的な手法を適用した結果,図に示された V1, V2, V3d, V3Aのみが刺激に対して頑健なパターンを示した領野と して信頼できる応答パターンを示した。興味深いこと に,これら初期から中・高次の視覚野は,脳内の背側視 覚経路の階層性に沿った領野であり,今回の実験に限れ ば,腹側経路の関与する証拠は得られなかった。これら V1からV3Aに至る類似度パターンを観察すると,V1で はそれほど特徴的なパターンが観察できないが,V2で は行列表の四隅に特徴的なパターンが観察され,V3Aで は3つの刺激条件によらず傾き面の角度が似ていれば高
Figure 4. Stimuli illustrations. Diagrams of the parametric slant variations in the three experimental conditions: Main: simu-lated rotation of a physical disc; Spatial control: projection height in the image plane was constant as slant was manipusimu-lated; Disparity control: the mean (unsigned) disparity was constant as slant was varied. Modified from Ban et al., 2015.
Figure 5. Results of multi-voxel pattern similarity analysis. Empirical similarity pattern matrices from areas V1, V2, V3d, and V3A. Slant angle varies in an ordered fashion for the three experimental conditions (main, spatial control, disparity control). Modified from Ban et al., 2015.
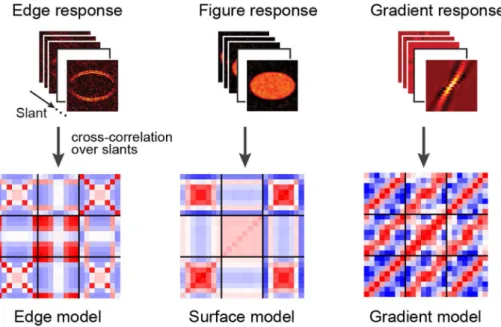
い相関パターンが観察される(角度が異なれば負の相関 パターンが観察される)ことが見て取れる。 これらの類似度行列パターンをより定量的に評価する ため,神経活動のシミュレーションを行い,シミュレー ション結果から類似度パターンの説明を試みた。具体的 には,(1)ターゲット刺激のエッジ(3D空間でのギャッ プ),(2)刺激の表面(網膜上でのターゲット刺激の拡 がり),(3)刺激の傾き(角度),のそれぞれの特徴のう ち,どれか1つのみを表象するようなROIがあるとすれ ば,そのROIの類似度パターンはどのようなものになる のかをシミュレーションとモデリングによって求めた。 続いて,それら3つの類似度行列モデルを重み付け線形 加算することにより,実際のfMRI脳機能計測で得られ たV1からV3Aまでの類似度行列パターンを説明できる か,重回帰法による分析を行った。その結果がFigure 6 である。V1ではローカルな情報(3D物体のエッジ)が 処理され,V2では 3D物体の表面(網膜での拡がり), V3ではそれらの手掛かりが徐々に必要なくなり,最後 にV3Aでは物体表面の傾きが表現されていることが定 量的に明らかになった(Figure 6)。 これまでのモデリング研究により,2次元の画像に含 まれる物体を同定する際には,画像からエッジを抽出 し,エッジの内部を埋め(図地の分離や表面の同定), 物体が何であるかを同定する,といった階層的な処理が 提案され,神経科学的な研究により,実際に脳内の腹側 経路に沿って階層的な処理機構が存在することが明らか になっていたが,我々の研究は3D情報を階層的に処理 する経路が背側視覚経路に沿って存在することを明らか にした。この研究結果は解剖学的にも非常に興味深い。 前節では複数の 3D手掛かりがV3B/KOで統合されてい ることを示した研究を紹介した。V3AはこのV3B/KOに 隣接する領野であり,この領野で処理されたグローバル な両眼視差情報がV3B/KOへと入力されていると考えら れる。また,V3B/KOの前部には,運動知覚を司るとさ れるV5/MT+野も位置しているため,V3B/KOにおける
Figure 6. Illustration of the modeling approach. The outputs for the three different models are calculated for all of the stimuli presented to the observers. These model outputs are used to create simulated similarity matrices in the same manner as the empirical fMRI data. Modified from Ban et al., 2015.
Figure 7. Regression analysis of the empirical similarity matrices. The weights of each model are plotted for ar-eas V1, V2, V3d, and V3A. Modified from Ban et al., 2015.
異なる3D手掛かりの統合は,これら隣接する領野から の入力を受けて成立していると考えられる。 5. 今後の展望 本稿では,ヒト脳が3D視覚手掛かりをどのように処 理・統合しているのかに関する我々の最近の研究を紹介 した。我々ヒトが日常生活を送る上で視知覚が当然のよ うに3D空間で成立していることを考えれば,ヒトの3D 視の理論自体はごく単純で,すでに研究され尽くされた かのように思われていたかもしれない。しかしながら, 実際にはヒトがなぜ3D世界を知覚できるのか,その詳 細な脳内の情報処理機構はまだ解明されていないのが実 情である。特に絵画的あるいは単眼性の手掛かりから 3D知覚を実現する脳内メカニズムは,知識や経験など の曖昧模糊とした要因に依存するため,「こうした刺激 に対してはこうした知覚が生じる」という知覚現象の単 純な羅列を越えた記述を可能とする方法論すらまだ確立 されていない。今後,ニューラルネットワークや人工知 能研究などの最新の知見と心理学・神経科学的な手法を 組み合わせることで,この分野に新たなブレイクスルー がもたらされるかもしれない。 謝 辞 本稿で紹介した研究のうち,両眼視差と運動視差手掛か りの統合研究(Ban et al., 2012)は,Wellcome Trust (095183/ Z/10/Z), BBSRC (C520620), JSPS (H22,290)の補助を受け た。奥行き手掛かり統合の発達に関する研究(Dekker et al., 2015) は,UK ESRC (RES-061-25-0523), Wellcome Trust (095183/Z/10/Z), JSPS (KAKENHI 26870911), NIH
(R01-MH-081990), Royal Society Wolfson Research Merit Award, The Swire Trust, NIHR Biomedical Research Centre at Moor-fields Eye Hospital, UCL Institute of Ophthalmology の補助 を受けた。両眼視差の階層的な処理経路に関する研究 (Ban et al, 2015)は,Wellcome Trust (095183/Z/10/Z), JSPS (H22.290, KAKENHI 26870911)の補助を受けた。
引用文献
Ban, H., & Welchman, A. E. (2015). fMRI analysis-by-synthe-sis reveals a dorsal hierarchy that extracts surface slant. Journal of Neuroscience, 35, 9823–9835.
Ban, H., Preston, T. J., Meeson, A., & Welchman, A. E. (2012). The integration of motion and disparity cues to depth in dorsal visual cortex. Nature Neuroscience, 15, 636–643. DeAngelis, G. C., Ohzawa, I., & Freeman, R. D. (1991). Depth
is encoded in the visual cortex by a specialized receptive field structure. Nature, 352, 156–159.
Dekker, T. M., Ban, H., Van der Valde, B., Sereno, M. I., Welchman, A. E., & Nardini, M. (2015). Late development of cue integration is linked to sensory fusion in cortex. Cur-rent Biology, 25, 2856–2861.
Dovencioglu, D., Ban, H., Schofield, A. J., & Welchman, A. E. (2013). Perceptual integration for qualitatively different 3-D cues in the human brain. Journal of Cognitive Neuroscience, 25, 1527–1541.
Kreigeskorte, N., Mur, M., & Bandettini, P. (2008). Representa-tional similarity analysis―connecting the branches of sys-tems neuroscience. Frontiers in System Neuroscience, 2, 4. doi: 10.3389/neuro.06.004.2008
Murphy, A. P., Ban, H., & Welchman, A. E. (2013). Integration of texture and disparity cues to surface slant in dorsal visual cortex. Journal of Neurophysiology, 110, 190–203.
Muryy, A. A., Welchman, A. E., Blake, A., & Fleming, R. W. (2013). Specular reflections and the estimation of shape from binocular disparity. Proceedings of the National Acade-my of Sciences of the USA, 110, 2413–2418.
Nardini, M. Bedford, R., & Mareschal, D. (2010). Fusion of vi-sual cues is not mandatory in children. Proceedings of the National Academy of Sciences of the USA, 107, 17041–17046. Nardini, M., Begus, K., & Mareschal, D. (2013). Multisensory
uncertainty reduction for hand localization in children and adults. Journal of Experimental Psychology; Human Percep-tion and Performance, 39, 773–787.
Ohzawa, I., DeAngelis, G. C., & Freeman, R. D. (1990). Stereo-scopic depth discrimination in the visual cortex: Neurons ideally suited as disparity detectors. Science, 249, 1037– 1041.
Orban, G.A. The extraction of 3D shape in the visual system of human and nonhuman primates. Annual Review of Neu-roscience, 34, 361–388 (2011).
Preston, T. J., Li, S., Kourtzi, Z, & Welchman A. E. (2008). Multivoxel pattern selectivity for perceptually relevant bin-ocular disparities in the human brain. Journal of Neurosci-ence, 28, 11315–11327.
Sun, H-C., Di Luca, M., Ban, H., Muryy, A., Fleming, R. W., & Welchman, A. E. (2016). Differential processing of binocu-lar and monocubinocu-lar gloss cues in human visual cortex. Jour-nal of Neurophysiology, 115, 2779–2790.
Van de Walle G. A., & Spelke, E. S. (1996). Spatiotemporal in-tegration and object perception in infancy: Perceiving unity versus form. Child Development, 67, 2621–2640.