総説
1.結合・解離を 操る 合成生物学―CID
とLID
合成生物学(synthetic biology)は,生命現象を「つ くる」ことで理解することを目指す学問分野であ る1),2).合成生物学は,大まかに「生命そのもの」を つくる研究と,生体内やそれを模した環境下で「生物 学的現象」をつくる研究とに分けられるが,筆者の研 究は後者に属し,特に細胞内の生物物理学的現象を時 間的・空間的に自在に操作する技術の開発に努めてき た. これら技術開発の基盤としたのが,細胞外からの 刺激でタンパク質間の結合・解離を操作する技術, CID(chemically-inducible dimerization)および LID(light- inducible dimerization)である(図 1)3),4).それぞれ小分 子化合物および光刺激によって,細胞外から非侵襲的 かつ迅速に細胞内の特定のタンパク質同士の結合を誘 導する.この特性を活かし,細胞機能を外部から操作 する「スイッチ」としての応用が進められてきた5). CID・LID には,それぞれに複数のバリエーション が存在する.本稿では,小分子化合物ラパマイシン刺 激で FKBP-FRB 間の結合を誘導する CID(図 1 左パネ ル)と,青色光刺激により SspB-iLID 間の結合を誘導 する LID(図 1 右パネル)を採用した.両者の最大の 相違は,ラパマイシン刺激による CID は,細胞外か らラパマイシンを除いてもすぐには細胞内の FKBP-FRB が解離せず,実質的に非可逆とみなせるのに対 し,SspB-iLID 間の LID は暗所で速やかに両タンパク 質が解離する可逆的な系である,という点にある. 光操作技術全盛の現在,この可逆性に由来する高い 時間・空間分解能を理由に,LID は CID よりも優れ ている,という議論が目立つ.しかし一方で,持続的 な刺激が容易であることや,医薬品として広く認可さ れ,生物個体応用への障壁が低いというラパマイシン 特有の利点から,CID の技術的価値もまだ捨てたも のではない,というのが筆者の意見である.実験のし やすさや応用事例の要請に応じて,適切な方を選択す る,という適材適所の運用が妥当であろう. 本稿では CID および LID を基盤に,生きた細胞内 でペプチド鎖のゾル―ゲル相転移を操作する合成生物生きた細胞内に人工のハイドロゲル凝縮体をつくる
合成生物学技術
iPOLYMER
中村秀樹
1,2 1京都大学大学院工学研究科合成・生物化学専攻生物有機化学研究室 2ERATO浜地ニューロ分子技術Synthetic Formation of Hydrogel Condensates in Living Cells
Hideki NAKAMURA1,2
1Department of Synthetic Chemistry and Biological Chemistry, Graduate School of Engineering, Kyoto University 2JST ERATO Hamachi Innovative Molecular Technology for Neuroscience
In a previous report, the author developed a synthetic biology molecular tool, iPOLYMER, which produces hydrogel condensates con-sisting of two polypeptide chains in living cells, in a stimulus-dependent manner. Functionalizing the iPOLYMER polypeptides with an RNA-binding domain successfully reconstituted a typical example of biomolecular condensates, stress granules, in living cells under a stress-free condition. In the current review, achievements made possible by iPOLYMER are summarized, with reference to a brief histo-ry of biomolecular condensate research. Future prospects, as well as technical challenges, of synthetic biological approaches to the novel class of intracellular structures are also described.
synthetic biology / biomolecular condensates / phase transition/separation / hydrogels / stress granules / iPOLYMER/iPOLYMER-LI
図 1
タンパク質の結合を操作する技術 CID と LID.生きた細胞内の特 定のタンパク質間の結合を,1 分以内で操作することができる.
なった研究が存在する.まず 2009 年,Anthony Hyman の研究室から,線虫 C. elegans の生殖細胞分化に関わ る RNA 顆粒 germ line P granule が,相転移によって凝
縮する液滴のように振る舞うと報告された9).さらに 同グループから核小体も同様の振る舞いを示すことが 報告され10),これらの構造の物理的性質や,相転移・ 相分離現象との関連にスポットライトが当てられた. 相転移・相分離メカニズム①:天然変性タンパク質 しかしこの段階では,これらの細胞内構造の物理的 性質が,どのような分子機構を基に実現されているか は示されていない.2012 年にふたつのグループから 相次いで報告された研究は,この点を明らかにするも の で あ っ た.Steven L McKnight の グ ル ー プ は,bio-tinylated isoxazole という小分子で細胞・組織抽出液を 処理すると,多数の RNA 結合タンパク質と mRNA を 含む沈殿を生じることを発見した11),12).この沈殿の 構成分子が RNA 顆粒とよく一致することから,彼ら はこの沈殿をいわば人工の in vitro RNA 顆粒として詳 細な検討を行った.特に重要な結果として,この in
vitro RNA 顆粒に含まれる RNA 結合タンパク質の多く
が low complexity domain という,比較的限られたアミ ノ酸から構成される天然変性ドメインを持つこと,さ らにこの天然変性ドメインを高濃度で精製すると,ハ イドロゲル凝縮体を形成することを示した. 相転移・相分離メカニズム②:多価相互作用ドメイン ほぼ時を同じくして,Michael K Rosen のグループ は,細胞骨格の制御に関わるタンパク質に見られるふ たつのドメイン,proline-rich motif(PRM)と SH3 を それぞれタンデムに連結したふたつの精製ポリペプチ ド鎖が試験管内で液滴を形成すること,この液滴内部 の電子顕微鏡画像がハイドロゲルで見られるような高 い電子密度を示すことを報告した13).彼らはさらに, 生きた細胞内でもこれらのペプチド鎖が凝縮体を形成 することを確認した. これらの先行研究から浮かび上がってくるのは,生 体分子間に働く多価の相互作用が,液滴の相分離やゾ ル―ゲル相転移のような物理化学的現象を介して生体 分子凝縮体のダイナミックな集合・離散を実現すると いう,細胞内秩序形成の新たな機構である.筆者ら は,タンパク質の相転移・相分離を時空間的に自在 に操作・擾乱する技術の開発を通じて,この新たな 視点の確立に貢献することを目標に,合成生物学技術 iPOLYMER の開発に着手した. 学技術 iPOLYMER6)について,その背景と現在までに 得られた成果,今後の展望をご紹介したい. 2. 生体分子凝縮体の「再発見」と相転移/相分離 私たちの身体をかたち作る細胞は,多種多様な分子 から成り立っている.これらの分子は,細胞内に一様 に存在するのではなく,各々の生理機能を発現する場 所に局在する.互いに関連した機能を有する多数の生 体分子が局所的に集合することで,細胞内に様々な機 能的サブコンパートメントを形成し,生化学反応の効 率化を実現している. 細胞内サブコンパートメントとして最もよく知られ ているのが,古典的な細胞内小器官(オルガネラ)で ある.脂質二重膜の境界で囲まれ,構成分子の精緻な 輸送システムによって維持されるこれらの小器官は, それぞれに固有の生理学的過程を担う.ゲノム DNA からの転写に関わる分子群は核に,好気的エネルギー 代謝に関わる分子群はミトコンドリアに,という具合 に,特定の過程に関わる分子群は,その過程を担う小 器官に局在する. 生体分子凝縮体=膜を持たないダイナミックな構造 一方,最近の細胞生物学研究で注目を集めている 「新たな」細胞内サブコンパートメントが,「生体分子 凝縮体(biomolecular condensates)」あるいは「非膜型 オルガネラ(membrane-less organelles)」と呼ばれるク ラスである7),8).古典的オルガネラと異なり,これら は脂質二重膜からなる物理的境界を持たず,周囲の環 境との間に活発な物質交換が存在する.このため,凝 縮体の多くはその必要性に応じてダイナミックに集 合・離散することが可能である. 代表的な例として,核小体や,後述するストレス顆 粒などの様々な RNA 顆粒,さらには神経変性疾患で 見られる封入体などが挙げられる.先に「新たな」と 括弧つきで書いたのは,これら個々の生体分子凝縮体 そのものは,多くの場合古くから研究された細胞内構 造である,という事情による.「生体分子凝縮体」と いう細胞内コンパートメントは,新規に「発見」され たというより,ある共通項の発見を通して「再発見」 された分類であり,その共通項こそが,最近方々で 耳にする「液―液相分離(liquid-liquid phase separation, LLPS)」などの相転移・相分離現象なのである.
相転移・相分離した構造としての「再発見」
凝縮体は融合を繰り返しながら徐々に成長し,24時 間後には 10 μm 程度に達するものもあった.これは RNA 顆粒など生理的な生体分子凝縮体と同様の振る 舞いとサイズであることから,iPOLYMER が細胞内に 「人工の生体分子凝縮体」を形成する技術となりうる可 能性を示している.そこで筆者らは,iPOLYMER を用 いて細胞内で「人工のストレス顆粒(stress granules)」 の形成実験を試みることにした. ストレス顆粒は,ストレス条件下にある細胞の細胞 質に形成され,多数の RNA 結合タンパク質と mRNA からなる RNA 顆粒の一種である14).異なる複数のス トレスによってその形成が同様に誘導され,集積した mRNA の翻訳が抑制されることから,細胞のストレ ス耐性に関わるのではないかと提唱されてきたが,ス トレス耐性における生理的意義を端的に示す結果は未 だ得られていない.一方で,最近の研究ではストレス 顆粒の異常形成が神経変性疾患の病態につながるとい う報告があり,病理的観点からも注目されている15). 実験的な扱いやすさから,ストレス顆粒は最も研究さ れた生体分子凝縮体のひとつでもある. 筆者らは,ストレス顆粒の構成タンパク質のひとつ である TIA-1 に着目した.TIA-1 は,RNA 顆粒構成タ ンパク質の例にもれず,RNA 結合ドメインである 3. 細胞内にハイドロゲル凝縮体をつくる;
iPOLYMER
そこで筆者らは,前述のRosenらの論文13)を参考に, CID や LID で用いるタンパク質ドメインをそれぞれ タンデムに結合したペプチド鎖を用いることで,相転 移を介したタンパク質の集合が刺激依存的に再現でき ると予想し,iPOLYMER(induced production of ligand-yielded multivalent enhancers)をデザインした(図 2).タンデム FKBP 鎖およびタンデム FRB 鎖を精製し, 混合してラパマイシンを投与すると,速やかにタンパ ク質凝集が生じた.この凝集を遠心・回収して様々な 物理的検証を行った結果,得られた凝集は 75% の含 水率を有し,6 nm 以下のメッシュ(ポア)サイズを 持つハイドロゲルであることを明確に示すことができ た.そこで同じ iPOLYMER ペプチド鎖を生きた細胞 内で発現し,蛍光イメージングによる検討を行った. 細胞外にラパマイシンを投与すると,10 分間で細 胞 内 に 凝 縮 体 の 形 成 が 確 認 さ れ た(図 3a). こ の iPOLYMER 凝 縮 体 は,in vitro で の 検 証 と は 異 な り GFP などのタンパク質が内部に浸透するのを排除し ないが,輸送小胞などより大きな細胞内構造の運動を 阻害した.また,光刺激依存型の iPOLYMER-LI(light-inducible)を細胞内に発現して検証を行ったところ, 局所的かつ可逆的に凝縮体を形成する実験に成功した (図 3b).形成された凝縮体は,一般に液滴のように 滑らかな境界線を持たず,むしろ in vitro での検討が 示唆するように,ハイドロゲル様の物質と考えられ た. 図 2 合成生物学技術 iPOLYMER/iPOLYMER-LI のデザイン.CID および LID を基に,使用されるタンパク質それぞれをタンデムに結合し たふたつのポリペプチド鎖からなる.刺激依存的にポリペプチド 鎖が“架橋”され,ハイドロゲル凝縮体を形成する.タンデムに 結合するタンパク質ドメインの数を調整することで,相互作用の 価数を直観的に調節可能である. 図 3 iPOLYMER/iPOLYMER-LI による細胞内凝縮体形成.a)COS-7 細胞 に iPOLYMER ペプチド鎖を発現し,ラパマイシンを細胞外に投与 した.刺激前(上パネル)には一様に分布していた CFP-FRBx5(緑) と YFP-FKBPx5(赤)が,刺激後 15 分(下パネル)までに明確な 凝縮体を生じていることが分かる.b)光刺激依存型 iPOLYMER-LI による可逆的かつ局所的な凝縮体形成.左列パネル青色四角形で 示した領域を順次光刺激した.YFP-SspBx6(緑)と mCherry-iLIDx6 (赤)が,同一の細胞内で可逆的かつ局所的に凝縮体を形成してい る.スケールバー;10 μm.文献 6 より改変.
of Medicine 井上尊生(Takanari Inoue)プロフェッサー の研究室で開発した合成生物学技術 iPOLYMER につ いてご紹介した.iPOLYMER は上記のように,生き た細胞内にいわば「人工のストレス顆粒」をストレス なしで「つくる」ことに成功した.これは筆者の知る 限り,生理的な生体分子凝縮体を細胞内で人工的につ くり出した世界初の事例である.なお,iPOLYMER に よる人工ストレス顆粒形成実験手順の詳細について は,近くプロトコルを出版予定である17). 一方で iPOLYMER には,凝縮体形成にポリペプチ ド鎖の高レベル発現が必要であり,形成の時間スケー ルやサイズについても細胞間のばらつきが大きい,な ど技術的課題も存在する.いくつかの観点から検証を 行い,iPOLYMER を,よりロバストに動作する汎用 性の高い技術へと進化させるべく改良を試みている. また,iPOLYMER と同時期に,同様に生体分子凝 縮体の集合過程を生きた細胞内で人工的に再現すると いう合成生物学的アプローチの報告が他にも相次い だ.iPOLYMER をはじめ,前述の Clifford P Brangwynne
らの研究18)を含むこれらの報告について,特に液滴・ ハイドロゲル・非可逆的凝集など凝縮体の物理的性質 と,その生理学的役割との相関を軸に総説を出版した ので,興味を持たれた読者はご一読いただきたい19). これら人工の生体分子凝縮体を「つくる」技術に加 えて,内在性の凝縮体を「こわす」技術の開発は,生 体分子凝縮体の生理的機能を探索する上で必須であ る.筆者の最近の研究では,細胞内のアクチン重合を 自在に操作する技術をストレス顆粒に応用し,ストレ ス条件下の細胞中でストレス顆粒を離散させることに 成功した(現在論文投稿中)20).離散の詳細な機構や ツールとしての汎用性などまだまだ課題は山積だが, 今後の発展に期待したい. 相転移・相分離と生体分子凝縮体は,猛烈な勢いで 発展を続けている研究分野であり,本稿ではスペース の関係上ご紹介できなかった重要な研究や知見が数多 くある.この場を借りてお詫び申し上げる. 最後になったが,iPOLYMER の開発を共同で担当 した Albert A. Lee,遅々として進まない筆者の研究を 忍耐強くサポートしてくださった Johns Hopkins Uni-versity 井上尊生プロフェッサー,畑違いの研究者であ る筆者を受け入れてくださった現所属の浜地格教授, そして両研究室のメンバーに最大限の謝意を表して, 本稿の締めくくりとしたい.
文 献
1) Bashor, C. J., Collins, J. J. (2018) Annu. Rev. Biophys. 47, 399-423.
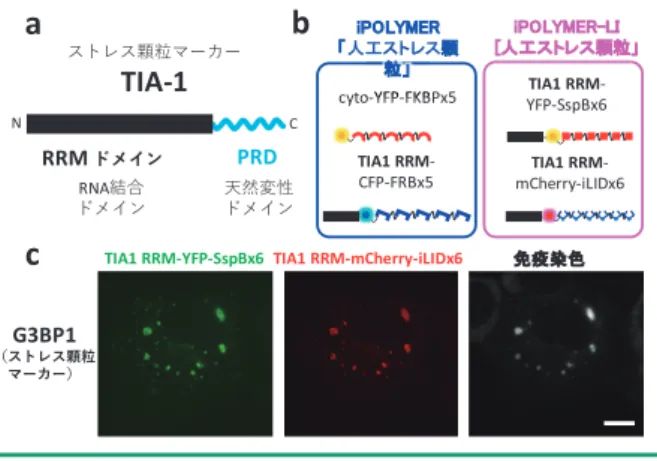
RRM ドメインと prion-related domain(PRD)と呼ばれ る天然変性ドメインからなり(図 4a),この PRD は ハイドロゲルを形成することが示されている11).さら に,TIA-1 は PRD を酵母のプリオンタンパク質由来 の別の天然変性ドメインに置き換えてもストレス顆粒 形成の足場として機能することが報告されていた16). そ こ で iPOLYMER の ペ プ チ ド 鎖 に TIA-1 の RRM ドメインを融合させ,細胞内で RNA 結合ドメイン を含む iPOLYMER 凝縮体を刺激依存的に形成した (図 4b).ストレス顆粒マーカー 4 種(PABP1, G3BP1,
eIF3b, eIF4G),ストレス顆粒非マーカー 1 種(ribosomal P-antigen)の計 5 種類の RNA 結合タンパク質につい て免疫染色を行ったところ,TIA-1 由来の RRM ドメ インを含む iPOLYMER 凝縮体には全てのストレス顆 粒マーカーが集積した(図 4c)一方,ストレス顆粒に 集積しない ribosomal P-antigen の集積を認めなかった. この結果はラパマイシン依存型の iPOLYMER,および 光刺激依存型の iPOLYMER-LI の双方で同様に認めら れ,細胞内の RNA 結合ドメインを含む iPOLYMER 凝 縮体は,ストレス条件下で形成される生理的ストレス 顆粒の特異的分子構成を再構成する「人工ストレス顆 粒」であることが示された6). 4.生体分子凝縮体の合成生物学;まとめとこれから
本稿では,筆者が米国Johns Hopkins University School
図 4 iPOLYMER による「人工ストレス顆粒」の再構成.a)ストレス顆 粒マーカータンパク質 TIA-1 のドメイン構造.N 末端側に位置す る RNA 結合ドメイン,RRM ドメインを用いて実験を行った.b) iPOLYMER/iPOLYMER-LI を用いた「人工ストレス顆粒」再構成のデ ザイン.TIA-1 由来の RRM ドメインを iPOLYMER ポリペプチド鎖 の N 末端に融合し,小分子あるいは光刺激で細胞内に凝縮体を形 成する.c)ストレス顆粒マーカー G3BP1 の iPOLYMER-LI「人工ス トレス顆粒」への特異的集積.複数の RNA 結合タンパク質につい て免疫染色を行い,光刺激によって形成された凝縮体への集積を 検証した.スケールバー;10 μm.文献 6 より改変.
総説 中村秀樹(なかむら ひでき) 京都大学大学院工学研究科・JST-ERATO 浜地ニュー ロ分子技術特定助教 2006 年東京大学大学院理学系研究科博士課程修 了(物理学専攻),理化学研究所研究員,早稲田 大学助教,Johns Hopkins University 研究員を経て 2019 年より現職. 研究内容:合成生物学・細胞生物学・生物物理学 連絡先:〒 615-8530 京都市西京区京都大学桂 工 学研究科附属桂インテックセンター 308 号室 E-mail: [email protected] URL: http://www.sbchem.kyoto-u.ac.jp/hamachi-lab 中村秀樹
15) Zhang, K. et al. (2018) Cell 173, 958-971.e17. DOI: 10.1016/ j.cell.2018.03.025.
16) Gilks, N. et al. (2004) Mol. Biol. Cell 15, 5383-5398. DOI: 10.1091/mbc.e04-08-0715.
17) Nakamura, H., Methods Mol. Biol., in press
18) Bracha, D. et al. (2019) Nat. Biotechnol. 37, 1435-1445. DOI: 10.1038/s41587-019-0341-6.
19) Nakamura, H. et al. (2019) J. Biochem. 166, 13-27. DOI: 10.1093/ jb/mvz028.
20) Nakamura, H. et al. (2020) bioRxiv. DOI: 10.1101/2020.03.30. 016360.
DOI: 10.1146/annurev-biophys-070816-033903.
2) Ausländer, S. et al. (2017) Angew. Chem. Int. Ed. Engl 56, 6396-6419. DOI: 10.1002/anie.201609229.
3) DeRose, R. et al. (2013) Pflugers Arch. 465, 409-417. DOI: 10.1007/s00424-012-1208-6.
4) Niu, J. et al. (2016) Biophys. J. 111, 1132-1140. DOI: 10.1016/ j.bpj.2016.07.040.
5) Kim, A. K. et al. (2016) Sci. Signal. 9, re1. DOI: 10.1126/scisignal. aac4779.
6) Nakamura, H. et al. (2018) Nat. Mater. 17, 79-89. DOI: 10.1038/ nmat5006.
7) Banani, S. et al. (2017) Nat. Rev. Mol. Cell Biol. 18, 285-298. DOI: 10.1038/nrm/2017.7.
8) Wheeler, R. J., Hyman, A. A. (2018) Philos. Trans. R. Soc. Lond. B. Biol. Sci. 373, 20170193. DOI: 10.1098/rstb.2017.0193.
9) Brangwynne, C. P. et al. (2009) Science 324, 1729-1732. DOI: 10.1126/science.1172046.
10) Brangwynne, C. P. et al. (2011) Proc. Natl. Acad. Sci. USA 108, 4334-4339. DOI: 10.1073/pnas.1017150108.
11) Kato, M. et al. (2012) Cell 149, 753-776. DOI: 10.1016/j.cell. 2012.04.017.
12) Han, T. W. et al. (2012) Cell 149, 768-779. DOI: 10.1016/j.cell. 2012.04.016.
13) Li, P. et al. (2012) Nature 483, 336-340. DOI: 10.1038/nature 10879.
14) Kedersha, N. et al. (2013) Trends Biochem. Sci. 38, 494-506. DOI: 10.1016/j.tibs.2013.07.004.