イカ筋肉のアデノシン脱アミノ酵素
13
0
0
全文
(2) Vol. 19, No, 1 Journal of Hokkaido University of Education (Section II A) September, 1968. Purification and Properties of Adenosine Deaminase from the IVtuscle of a Squid (^Ommastrephes sloani pacificusy*. Midori SAKAITOHI, Kuniko YOSHIDA and Sakari NARA*'". Chemical Laboratory, Hokkaido University of Education, Hakodate S?. isW?-. zisX^ : -< A%l^®77:'^ ^yfi7 ^@N? Summary Adenosine deaminase (adenosine aminohydrolase, EC 3. 5,4. 4) has been purified about 200-fold from the squid muscle by the following techiques : aqueous extraction, heating to 50°, precipitation with ammonium sulfate, and column chromatography on DEAE-cellulose.. The purified enzyme was chromatographically homogeneous and a bulk of the protein migrated paper-ionophoretically was highly active. The crude enzyme resisted to heat, but. the activity of the purified enzyme was quitely lost in half an hour at 50 °C. The optimal pH was observed at 7. 5 with phosphate buffer and Mcllvaine buffer. The apparent activation energy was found to be approximately 12,OOOcaVmol. The Michaelis-Menten constant was calculated to be 1. 39 x 10-5M. Finally, the effect of various additions on the enzymatic activity was described. Especially Hg2+ was a potent inhibitor whose Ki (dissociation constant of enzyme-inhibitor complex) was calculated to be 6. 5 X 10-6M in its competitive inhibition.. It is well known that adenosine deaminase (adenosine aminohydrolase, EC 3. 5. 4. 4) is distributed in varying concentration not only in the organs of animals, but also in bacteria.. Since this enzyme was shown to be present in the muscle of a rabbit by SchmidtD in 1928, it has been extensively studied enzymo-chemically and physiologically by many authors. Conway et al.W found its presence with 5/-adenylic acid deaminase (AMP aminphydro-. lase, EC 3.5.4, 6) in the blood and tissues of rabbit and suggested that it might be important in detoxifying the vasodepressant adenosine into relatively inactive inosine for the reason of its high concentration in the intestine. Kornberg et fl/.3) reported on the extract of calf intestinal mucosa that this enzyme deaminated slowly synthetic 2,6-diaminopurine riboside (2-aminoadenosine).. In regard to the detailed purification procedures of this enzyme from the calf intestinal mucosa was given by Brady and O'Cnnnell.4) They obtained first the homogeneous preparation. judged by the method of ultracentrifugal analysis. * A part of this work was presented at the 40th Annual Meeting of the Japanese Biochemical Society,. Osaka (1967). ** To whom requests for reprints should be submitted.. ( 55).
(3) Adenosine Deaminase of a Squid Muscle. And moreover, one of them, BradyS) reported in detail with his co-worker that the deamination of adenosine and deoxyadenosine was different a little from the organs of six mammalian animals in the view of a comparative biochemistry. Fischer et alfo) suggested. that the activity of this enzyme in the chicken duodenum could be increased 40-fold by an injection of adenosine. This result was not consistent with the observation performed by Gordon et al. who has found the similar finding rather in its embryos.7) Recently, Hoagland and Fischei-8) have partially purified adenosine deaminase from the chicken duodenum and observed some properties with particular emphasis on its inhibition by mercurials. Aikawa9) has reviewed the physicochemical properties of this enzyme from the clam mid-gut gland. Concerning to the non-specific adenosine deaminase, Nakanishi et al.W-W have. examined its specificity of substrates and inhibitors. On the other hand, the activity of this enzyme in plants was found in the dried barley roots by Fiers et al.W and the discussions relating to its presence in plant seeds has been arise in the recent paper, is) However, little has known on this enzyme from an invertebrate muscle. The present paper described the purification procedures and some enzymatic properties on this enzyme from the squid muscle.. MATERIAL AND METHODS Materials — Fresh squids (Ommastrephes sloani pacificns') were caught on the southern coast of Hokkaido from August to November, 1967.. All chemicals were of analytical grade and obtained from Wako Pure Chemical Industries Co. DEAE-cellulose (BROWN) was purchased from Seikagaku Kogyo Co. Filter paper (No. 5, 2X 30cm) for an ionophoresis was obtained from Toyo Roshi Co. Methods — Adenosine deaminase activity was measured by reading the decrease in the optical density at 265m,u according to the method of Kalcker.16) The standard assay system consists of 0.05 ml of 1.87xlO-3M adenosine solution in 0.1M phosphate buffer, pH 7. 0, 0.1 ml of the enzyme solution and the similar buffer solution to make 3 ml of the resulting solution in a quartz cell having a lcm light path.. The reaction was initiated by adding the enzyme solution and read at 265m/< for each 30-second intervals using a Shimadzu Model DU spectrophotometer. One unit of the enzyme. is defined at the amount which causes an initial rate of change in optical density (^fE^) ot 0.01 per minute under the condition mentioned above. Also ammonia liberated after the deamination was determined colorimetrically by a Nessler reagent for some experiments. Protein was determined by a biuret reactionl?) with using casein as a standard and also by a micro-Kjeldahl method. Specific activity was expressed as units per mg of protein.. The copper was determined colorimetrically by the method of Peterson and BollierlS) based on producing a blue complex between copper (cupric) and cupferazone. The migration of protein on the paper with veronal buffer (/<=0.1, pH 8.6) was performed under the following conditions ; 420 volts, 0. 3 mA/cm, and 3 hours by using a Toyo Roshi Type C apparatus.. ( 56 ).
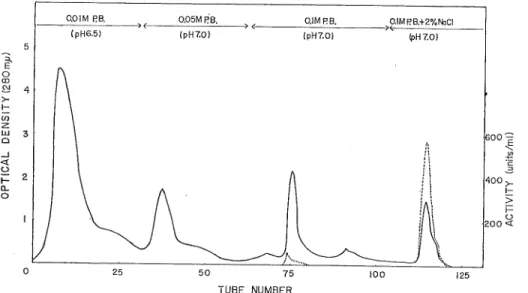
(4) M. SAKAUCHI, K. YOSHIDA and S. NARA. RESULTS Purification of Adenosine Deaminase Crude extraction from Muscle — Flesh muscles taken off from an alive squid were. minced and homogenized carefully in a homogenizer (Nihon Seiki Kogyo Co.) for 15 seconds with chilled water (1: 5 W/V). The resulting homogenates were centrifuged at 5,000 r.p.m. for 15 minutes. The precipitates were repeated by a similar procedure and the combined upper layer was called as Fraction 1. Heat Treatment — Each 300ml of Fraction 1, efficiently stirred, were heated to 50°±1° in a 500 ml stainless steel container by running warm water (55°±1°C). The suspension was kept at 50°±1°C for 3 minutes accurately and then chilled in a -10°C ice bach until the temperature dropped to 5°C. The denatured protein was centrifuged at 5,000 r.p.m. for 10 minutes and discarded. The slight opalescent supernatant was called as Fraction 2.. Ammonium Sulfate Precipitation and Dialysis-Solid ammonium sulfates were carefully added into the Fraction 2 to bring about 65% saturation. Much cloudy suspension was filtered through a Buchner funnel prepared by a large amounts of Hyflo Super-Cel, Deep. blue precipitates on the layer were collected immediately with a spatula and dissolved in a small amount of water. The resulting fairly bluish solution put into a cellophane tubing. was dialysed against chilled water (1: 20 V/V) for 20 hours with changing 3 times of the outer water. Many white precipitates appeared in the tubing during the dialysis.. Eliminating the precipitates by centrifuge, adenosine deaminase with a high activity was found in a pale blue supernatant liquid which was called as Fraction 3.. O.OIM P.B.. (pH6.5). 0.05M RB.. 0.1 MRB.. (pHT.O). IpHT.OI. O.IMf?B.+2%NaCI. (pH 7.0). i-. 0 £! 4 GO. < d. i400. Q.. 0. I-. >. 50. 75. 125. TUBE NUMBER Fig. 1. Column chromatography of the crude enzyme on DEAE-cellulose, DEAE-cellulose column, 2 x 50 cm, equilibratfid with 0.1M phosphate buffer (pH 6. 5) and then eluted with buffer as shown in this figure. Each tube, 6ml. Approx. 500 mg protein was loaded. : Protein measured by the change of optical density at 280m/<. : Enzymatic activity assayed by the change of optical density at 265m/<.. (57).
(5) Adenosine Deaminase of a Squid Muscle. DEAE-Cellulose Chromatography — Each 30 ml of the Fraction 3 was loaded onto a DEAE-cellulose column (2x50 cm) which was pre-equilibrated with 0.01M phosphate buffer solution, pH 7. 0. The elution of protein was subjected to a stepwise elution method and the buffer solution system at pH 7. 0 consisted of 0. OlM, 0. 05M, 0. 1M and 0.1M containing. sodium chloride (final concentration to 2%). A 6ml of the eluates was collected by a fraction collector at the rate of one ml per minute. A coloriess highly active enzyme fraction which was called as Fraction 4, was obtained from the last buffer solution as shown in Fig. 1. A blue fraction eluted with 0.1M buffer solution contained much protein but no enzymatic activity was observed. Rechromatography on DEAE-Cellulose — Fraction 4 was concentrated by saturated. ammonium sutfate solution (adjusted at pH 7. 0) and then dialysed against water till the desalting of dialysates entirely ceased. A 10 ml of this dialysates was rechromatographied on DEAE-cellulose (2x30 cm) which was prepared with 0.01M phoshate buffer solution,. 1500 luoo. 50. 100 150 200. 250. TUBE NUMBER Fig. 2. Rechromatography of Fraction 4 on DEAE-cellulose. The elution condition was almost the same as shown in Fig. 1 except the height of column (2 x 30 cm) and the ionic strength of the buffer. Each tube, 6 ml. Approx. 20 mg was loaded.- : Protein,. Table I. Ptirification of adenosme deamhiase from the squid musrle (200.S- muscle') Fraction. Volume. (ml). Activity (unit). Protein. (mg). Specific activity (units/rng). ^ds Purification. 1.. crude extract. 950. 57,000. 8,740. 6.5. 100. 2.. Heat-treatment. 830. 46,480. 1,992. 23.3. 81. 3.6. 3.. (NH4)siSO., 0.65saturation and dialysis. 60. 35,760. 510. 70.2. 62. 10.8. 4.. DEAE-cellulose chromatography. 16. 9,312. 19. 2 485.0. 16. 74.6. 5.. Rechromatography on DEAE-cellulose. 5. 7,270. 8 1,381.3. 12. 212,5. (58). 4.. 1.
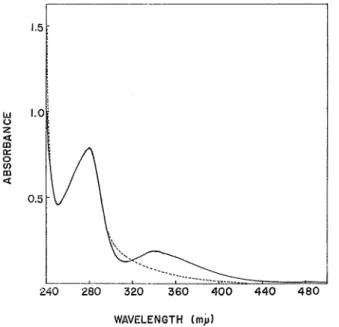
(6) M. SACAUCHI. K. YOSHIDA and S. NARA. pH 7.0. The buffer solution system used here was almost similer to the previous step except using 0. 3M buffer instead of 0.1M phosphafce buffer containing sodium chloride. The highest active enzyme preparation, named as Fraction 5, was observed in the eluates with 0.1M buffer solution as shown in Fig. 2. Overall results of the purification of the enzyme were summarized in Table I. Ultimately the enzyme preparation was purified more than 200 times as a specific activity.. Properties of Purified Adenosine Deaminase Purity — A homogeneity of the enzyme preparation has not been rigorously established yet, but it has been examined by paper-ionophoresis as given in Fig. 3. A bulk of protein migrated for anode was highly active. Also, a pattern observed by the column chromato-. graphy on DEAE-cellulose indicated a partial homogeneity.. t Fig. 3, Paper-ionophoregram of native enzyme. Filter-paper was prepared with veronal buffer (/<==0,1, pH 8.6). Condition of migration ; 420 volt, O.SmA/cm, 15 °, and 3 hours. Arrow indicates slot for addition of the sample. The distance of migration was about 8 cm for anode from arrow under the condition mentioned above,. Absorption Spectrum — A spectrum of the native enzyme preparation is shown in Fig. 4. Besides a major peak of an absorption at 280m/.<, a minor one around 340nv< was found in this figure. This minor component disappared immeadiately by adding a small amount of sodium dithionite, but recovered partially by an airation. Opiimal-PH'—Wect of pH on the enzymatic activity was carried out by using phosphate buffer (pH 4.0-9.0) and Mcllvaine buffer (pH 5.5-8.0). Each buffer solution was used in the reaction mixture instead of that in a standard assay system for a determination of the activity. A sharp peak appeared at pH 7.5. The results obtained are shown in Fig. 5. Stability — The enzyme preparation kept at 4°C was entirely inactive in one month, but its activity remained about S0% in deep-freezing. No loss showed in freezing even over one month, if ammonium sulfate was added to the preparation to make a final concentration of 10%, A pH-stability was shown in Fig. 6. The enzyme preparation. was diluted (1 : 10 V/V) with phosphate buffer solutions having various pHs (from 4 to 9) and was kept in a water bath at 35 °C. The enzyme showed the highest stability at pH 7.0 and retained its activity of 90% even after 6 hours.. (55).
(7) Adenosine Deaminase of a Squid Muscle. 1.5. u u z. < 00 a:. 0 0) m < 0.51-. 320. 360. 400 440 480. WAVELENGTH (mp) Fig. 4. Absorption spectra of native, and reduced enzyme. Specific activity, approx. 1,300. : native enzyme (0. 8 mg protein per ml) .- : enzyme reduced by Na-dithionite.. PH. Fig. 6. Effect of pH on enzymatic activity. The reaction mixture ( 3 ml) contained 0.1M phosphate buffer (-0—0-) or Mcllvaine buffer (-®—®-), 3.1xlO-3M adenosine, and about 6 units of the enzyme.. (60 ).
(8) M. SAKAUOHT. K. YOSHIDA and S. NARA. 100. TIME (hours) Fig. G. PH-stability of adenosine deaminase. The enzyme was 10-fold diluted with 0.1M phosphate buffer (pH 4.0-9.0) and kept in a water bath at 35 °C. The activity was measured by a standard method at various time intervals.. Also the stability was comparatively high at pH 6 or 8, but it dropped half in one hour at pH 5 or 9. At pH 4, no enzymatic activity showed after one hour. That is, a. stability of the enzyme was higher in a neutral state, but decreased remarkably in both acidic and alkaline sides. A thermostability was determined in the following procedure. Aliquots of the enzyme preparation were kept at pH 7.0 in a water bath adjusted with a. 100. ?^;=. •o •a-. -o——o 35°. -••-. 40°. 8C. .s. °s. t GO I—. u <. 45-. I 40 S£. I LL]. tK 20 s,50°. 10. 30. 60. 90. 1?0. TIME (minutes) Fig. 7. Thermostability of adensoine deaminase. The enzyme was kept at pH 7.0 in a water bath adjusted with a range of temperature from 35 °C to 50 °C. The activity was measured by a standard method at various time intervals as shown in this figure.. (62).
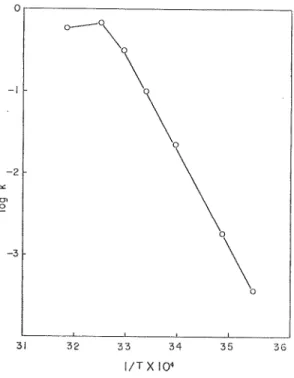
(9) Adenosine Deaminase of a Squid Muscle. temperature ranging from 35°C to 50°C. The remaining activity was determined by a usual method during constant time intervals.. The results obtained are given in Fig. 7. The highest thermostability was found at 35°C and 40°C, and the enzymatic activity of about 90% remained even after two hours.. The activity decreased by half in 2 hours at 45°C and perfectly in half an hours at 50°C. Effect of Temperature on the Reaction Velocity — An apparent activation energy of the enzyme reaction was approximately 12,000 caVmol as plotted after Arrhenius' equation. In this experiment, the reaction started by adding one ml of the enzyme preparation to. 1.25xlO-3M adenosine (final concentration) mixed with 2ml of 0.1M phosphate buffer solution, pH 7.0. A determination of the activity was carried out colorimetrically at 450m/-< by combing ammonia liberated with a Nessler's reagent at various temperatures for 10 minutes. The results obtained were shown in Fig. 8. It appears that a little denaturation of the enzyme occured at higher than 35 °C.. 34. 35. 36. 1/TXIO" Fig. 8. Arrhenius' plot of the temperature dependence of deaminase activity. The reaction started by adding 1 ml of the enzyme to 1.25xlO-3M adenosine mixed with 2ml. of 0.1M phosphate buffer (pH 7.0) and kept for 10 minutes at various temperatures. The activity was determined colorimetrically at 450m^.. Effect of Sttbsirate Concentration on the Reaction Velocity — Several concentrations of adenosine were prepared within the range from 8. 30 x 10-eM to 8. 30 x 10-5M. The enzymatic activity was measured by a standard assay system and the results obtained are shown in Fig. 9 according to a Lineweaver and Burk's plotl9)' and a Michaelis-Menten constant. ( 62 ).
(10) M. SAEAUOHI. K. YOSHIDA and S. NARA. was calculated to be 1.39xlO-5M phosphate buffer solution, pH 7.0. 2.5. 12. Fig. 9. Effect on substrate concentration on the initial reaction velocity. The reaction mixtures (3 ml) contained 0.1M phosphate buffer (pH 7. 0), 60 units of the enzyme and various concn. of adenosine. v. Units of the enzymatic activity, S, Mol. per liter.. 1.6. 1.6. 'IXIO-5M. 24 6 8 10 12. 1/(S)XIO'4 -6 -4 -2. 2468 Hg21-(XIOeM). Fig'. 10. Competitive inhibition by Hg?.+ and its Dixon's plot. The type of inhibition by Hg2+ was given by a Lineweaver and Burk's equation (left figure) and the value of Kt was calculated graphically (right figure). v, Units of the enzymatic activity. S, Mol per liter. In the right figure, A, B, C, D, and E represent the concentration of adenosine, that is, 0.83xlO-5M, 1.23X10-5M, 1.52xlO-5M, 2.08X10-5M, and 3.12 x 10-5M, respectively.. ( 63).
(11) Adenosine Deaminase of a Squid Muscle. Table II. Effect of various additions on the activity of adenosine deamhiase from the squid muscle The reaction mixture except adenosine was preincubated for 15 minutes at room temperature and then the reaction was started by mixing the substrate.. Additions. Final concentration. (M). m. Inhibition. Additions Final concentration. (M). Inhibition. (%). ( 5.0). NOs-. 1X10-3. ( 2.5). (10.2). so.^-. 1X10-3. ( 8.4) ( 8.0). Na+. 1X10-2. K+. 1X10-2. Ag+. 5xl0-6. 50,5. Mg2+. 1X10-2. ( 2.5). Ca2+. 1X10-2. ( 3.2). Co2+. 1X10-3. Ba2+. 1X10-3. Zn2+. 1X10-3. Hg2+,D. 3x10-8. 62.6. Pb2+. 3 x 10-''. Cu2+. 1X10-3. F-. 6x10-2. Pyrophosphate. 1X10-3. EDTA. 1 X 10-3. 12.6. ^-CMB". 1 X 10-5. 48.3. 5.0. Monoidoacetic acid. 1 X 10-3. 4.0. 4.3. Fericyanide. T x 10-a. 0. (15.2). Oxalic acid. 1X10-3. ( 7.6). Cysteine. 1 X 10-2. 45.7. Ascorbic acid. 1 x 10-*. ( 4.3) 0. 42.2. Hydroquinone. 1 X 10-3. ( 2.3). 57.5. H;A. 1 X 10-2. 0. 1) The preincubation time was used for 6 minutes. Figures in parlentheses represent acceleration, also in per cent,. Effect of Various Additions of ihe Reaction Velocity — Some additions including cathions, anions, oxidants, reducing agents, metal chelating agents and sulfhydryl binding agents were tested how they did affect on the reaction velocity. The concentration of additions was applied temporally in the range from 3xlO-6M to lx!0-ZM. The reaction mixture except adenosine was preincubated for 15 minutes at room temperature and then the reaction started by mixing the substrate. Hgz+ and p-CMB in mercurials were preincubated for 6 minutes, because a longer period in time intervals resulted in a decrease of its activity.. Results obtained are listed in Table II. Heavy metals were generally strongly inhibitory and especially, Hgz+, Ag+ and p-CMB had a potent inhibitory effect. The inhibition by Hg2+ is shown in Fig. 10, and also this figure20) indicates that Hg2+ was a competitive inhibitor for adenosine. It was much striking that the enzymatic activity in a low concentration of p-CMB was conversely accelerated as suggested by Fisher et al.s) That. is, the inhibition rate by p-CMB varied with time intervals of the preincubation. Concerning the inhibition by mercurials, the detailed experiments are in progress.. DISCUSSION As far as the purification of adenosine deaminase is concerned, an acetone fractionation adopted in the bovine intestinal mucosa'021), and an isoelectric point precipitation in the mid-gut gland of a clam9) were neither successful in this case because much denaturation occurred. While a calcium phosphate gel absorption in the beef livei-22) and Tenerds Cr2D seemed to be better, they were not applied here by reason of having a low yield. To obtain Fraction 3, the time interval of dialysis was taken for 20 hours under the condition. described previously and this fact appeared to be much important to elevate the yields in. (fi4).
(12) M, SAKAUOHI, K. YOSHIDA and S. NARA. the enzyme purification. Dialysing against water for more than 20 hours, many precipitates examplified as myosin or actomyosin in the squid muscle occurred gradually, but the time intervals resulted simultaneously in some inactivation of the enzyme. A column chromatography on DEAE-cellulose may be suitable for a separation of this enzyme from the other fractions, especially an unknown blue material containing approximately 6.5 w copper mg of protein at the highest. The purified enzyme gave one major band paper-ionophoretically and also indicated comparatively symmetrical curves in both the enzymatic activity and the protein as shown in Fig. 2, but no other check for the. homogeneity has been performed. Further detailed studies should be done and are still in progress. By a spectral analysis, the enzyme preparation has no absorption around 600m/-<. based on a blue color, but a different spectrum affected by sodium dithionite was observed at 340 rn/.i. It is not sure whether this spectrum is due to the enzyme itself or any contaminants.. The pH activity curves of this enzyme preparation are given in Fig. 5. It has an. optimum at pH 7, 5 which declined sharply on both acidic and alkaline sides. This result was something different from that of the rabbit (6.6), of the dog (6.8), of the clam and of the calf (7.0). In regard to thermostability, a remaining activity of the enzyme preparation showed about 90% even when maintained at 40 °C for 2 hours. This fact was in contrast to the behaviour of adenosine deaminase from a clam9) which was quite inactivated at 40 °C in 5 minutes.. It was evident that a Lineweaver and Burk's plot of the reciprocal of the initial velocity against the reciprocal of adenosine concentration gave a straight line as shown in Fig. 9. The value of Km for adenosine was calculated graphically as 1.39xlO-5M. Comparing this value with that of the other sources, it was almost the same order and especially it was close to that of the chicken duodenum deaminase (1.30xlO~5M). It is possible that this enzyme is classified as a sulfhydryl-enzyme, because it was. strongly inhibited by Hg2+, and p-CMB indicating the sulfhydryl nature of the enzyme and also activated by some reducing agents23\ Hg+z was competitively inhibitor/ at some range of its concentration for adenosine. However, it is noticeable that the enzymatic activity was observed to increase conversely, if less than 10-5M of p-CMB were incubated with the reaction mixtures. That the value of Ki for Hg2+ was smaller than Km for adenosine, might mean that the affinity of Hgz+ for the enzyme is larger than that of adenosine. According to Brady et al.,5) the activity of adenosine deaminase is generally low in a muscle of all species which are at least classified as vertebrates, but it appears to be high in the squid muscle. It is much interesting to ascertain whether or not some specificities may exist in the two classifications.. REFERENCES 1) G. Schmidt, Z. Physiol. Cliem., 179, 243 (1928) 2) D. J. Conway and R. Cooke, Biochein. /., 33, 479 (1939) 3) A. Kornberg and W. E. Pricer, Jr., /. Biol. Chem., 193, 481 (1951). ( 65 ).
(13) Adenosine Deaminase of a Squid Muscle 4) T. G. Brady and W. O'Connell, Biochim. Biophys. Acta, 62, 216 (1962) 5) T. G. Brady and C. I. O'Donovan, Comp. Biochem. Physiol., 14, 101 (1965) 6) J. R. Fisher, 0. P. Chilson and S. K. Chan, Biochim. Biophys. Ada, 58, 371 (1962) 7) M. W. Gordon and M. Roder, /. Biol. Chem., 200, 859 (1953) 8) V, D. Hoagland, Jr. and J. R. Fisher, /. Biol. Chem., 242, 4341 (1967). 9) T, Aikawa, Comp. Biochem. Physiol., 17, 271 (1966) 10) S. Minato, T. Tagawa and K. Nakanishi, /. Biochem., 58, 519 (1965) 11) S. Minato, T. Tagawa, M. Miyalu, B. Shimizu and K. Nakanishi, /. Biochem., 59, 265, (1966) 12) S. Minato, T. Tagawa and K. Nakanishi, /. Biochem., 60, 352 (1966). 13) S. Minato and K. Nakanishi, /. Biochem., 62, 21 (1967) 14) W. Piers and L. Vandendriessche, Arch. inter. Physiol. et Biochem., 68, 203 (1960). 15) T. G. Brady and V. J. Hegarty, Nature, 209, 1027 (1966) 16) H. M. Kalckar, /. Biol. Chein., 167, 461 (1947) 17) A. G. Gornall, C. J. Bardawill and M. M. David, /. Biol. Cliem., 177, 751 (1949) 18) P. E. Peterson and M. E. Bollier, And. Chem., 27, 1195 (1955) 19) H. Lineweaver and D. Burk, /. Am. Chem. Soc., 66, 658 (1934). 20) M. Dixon, Biochem. J., 55, 170 (1953) 21) W. Klein, Z. Physwl., Chein., 324, 163 (1961) 22) K. Hagiwara, K. Tsujimoto, M. Sakamoto and T. Hiroo, /. Female College of Shitennoji (in Japanese),. p. 102 (1966). 23) P. D. Boyer, "The Enzymes", ed. by P. D. Boyer, H. Lardy and K. Myrback, Academic Press Inc.,. New York, Vol. I. p. 511 (1959). ( 66 ).
(14)
図


+4


関連したドキュメント
An easy-to-use procedure is presented for improving the ε-constraint method for computing the efficient frontier of the portfolio selection problem endowed with additional cardinality
If condition (2) holds then no line intersects all the segments AB, BC, DE, EA (if such line exists then it also intersects the segment CD by condition (2) which is impossible due
The inclusion of the cell shedding mechanism leads to modification of the boundary conditions employed in the model of Ward and King (199910) and it will be
Many interesting graphs are obtained from combining pairs (or more) of graphs or operating on a single graph in some way. We now discuss a number of operations which are used
It is suggested by our method that most of the quadratic algebras for all St¨ ackel equivalence classes of 3D second order quantum superintegrable systems on conformally flat
This paper is devoted to the investigation of the global asymptotic stability properties of switched systems subject to internal constant point delays, while the matrices defining
Next, we prove bounds for the dimensions of p-adic MLV-spaces in Section 3, assuming results in Section 4, and make a conjecture about a special element in the motivic Galois group
The main problem upon which most of the geometric topology is based is that of classifying and comparing the various supplementary structures that can be imposed on a
