© 2013 The Japan Society of Histochemistry and Cytochemistry AHC
Acta Histochemica et Cytochemica 0044-5991
1347-5800
Japan Society of Histochemistry and Cytochemistry Tokyo, Japan
AHC13002 10.1267/ahc.13002 Regular Article
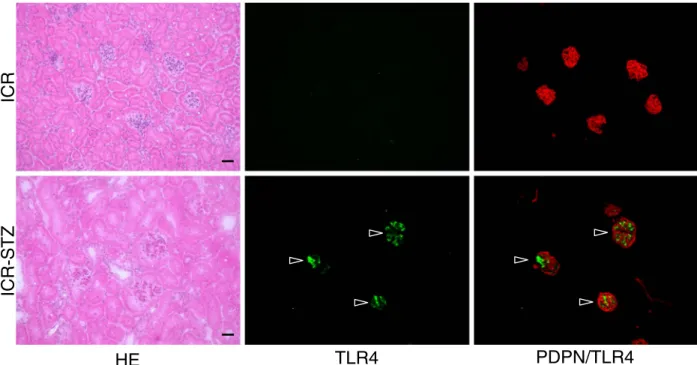
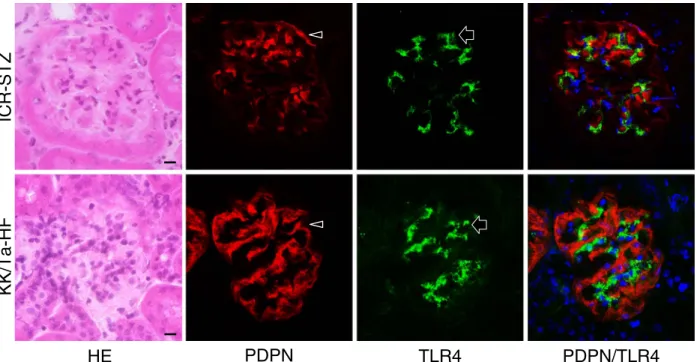
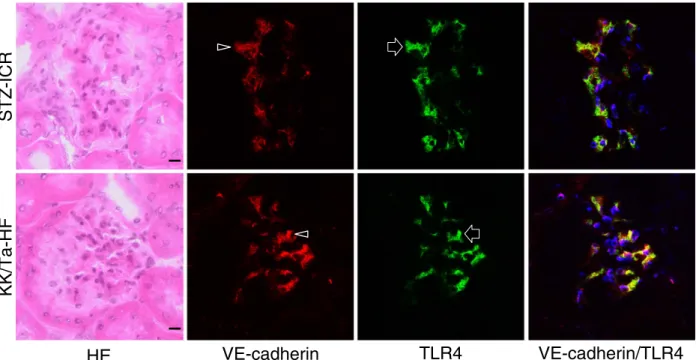
Expression of Toll-Like Receptor 4 in Glomerular Endothelial Cells under Diabetic Conditions
Shunsuke Takata 1 , Yoshihiko Sawa 2 , Takanobu Uchiyama 1 and Hiroyuki Ishikawa 1
1
Department of Oral Growth & Development, Fukuoka Dental College, 2–15–1 Tamura, Sawara-ku, Fukuoka 814–0193, Japan and
2