早稲田大学審査学位論文 博士(スポーツ科学)
Effects of endurance exercise
on serum fibroblast growth factor 21 level
持久性運動が
血清 Fibroblast growth factor 21 濃度に及ぼす影響
2016年1月
早稲田大学大学院 スポーツ科学研究科
谷口 祐一
TANIGUCHI, Hirokazu
研究指導教員: 樋口 満 教授
Contents 1. Background
2. Cardiorespiratory fitness and visceral fat are key determinants of serum fibroblast growth factor 21 concentration in Japanese men
[1] Introduction
[2] Materials and Methods [3] Results
[4] Discussion and Conclusion
3. Endurance exercise reduces hepatic fat content and serum fibroblast growth factor 21 levels in elderly men
[1] Introduction
[2] Materials and Methods [3] Results
[4] Discussion and Conclusion
4. Conclusion and suggestions for future research 5. Acknowledgments
6. References
…
…
…
…
…
…
…
…
…
…
…
…
…
… 1
2 2 3 7 14
19 19 20 24 30 32 33 33
1 1. Background
Elevated cardiometabolic morbidity, such as diabetes and cardiovascular disease, is a worldwide problem (1), and there has been considerable interest in methods that prevent the development of these cardiometabolic diseases. The onset of these disorders is strongly associated with obesity (1); however, higher cardiorespiratory fitness (CRF) level reduces cardiometabolic risk regardless of obesity status (2,3). Although endurance exercises that increase CRF are also important for preventive strategies to reduce cardiometabolic morbidity, the underlying mechanism as to how CRF and endurance exercise prevent cardiometabolic diseases has not been fully explained. The elucidated mechanisms of the beneficial effects of exercise may provide a novel approach to treating cardiometabolic disease. Thus, there is great concern about acute and chronic exercise-induced adaptations in both, animal and human studies.
Because the beneficial effects of exercise are systemic, it is likely that unidentified or unexplained peptide hormones are concerned with CRF and/or endurance exercise. In accordance with the idea of exercise-induced mediators, recent studies have suggested that endurance exercise induced circulating cytokines that have important roles in homeostasis, specifically in adipose tissue thermogenesis (4,5). The significant exercise-induced changes in circulating cytokine concentrations (4,5) and the therapeutic effects of markedly elevated cytokines level has previously been documented in experimental animals (4,6). However, the question remains as to whether these cytokines play key roles in human metabolism in the same manner as in experimental animals. It is therefore necessary to obtain evidence that connects experimental results to human health in cytokine research.
Recently, there has been increased interest in fibroblast growth factor (FGF) 21 metabolism (7,8). Research has shown that circulating FGF21 level was increased by acute endurance exercise in both, rodents and humans (5). Previous animal studies have reported that temporary increases in serum FGF21 improve glycolipid metabolism and obesity (6,9); thus, the exercise-induced changes in serum FGF21 level may partially explain how endurance capacity
2
and exercise prevent cardiometabolic morbidity. On the other hand, it was also observed that chronically high levels of serum FGF21 produce an FGF21-resistant state (10,11), resulting in earlier onset of cardiometabolic disease (12-14). This result suggests that unlike the effects of acute exercise, regular exercise may play an opposing role in circulating FGF21 concentrations.
However, these studies did not examine the association of serum FGF21 levels with CRF and regular endurance exercise. This study therefore aimed to evaluate the association of serum FGF21 levels with CRF and regular endurance exercise.
2. Cardiorespiratory fitness and visceral fat are key determinants of serum fibroblast growth factor 21 concentration in Japanese men
[1] Introduction
Fibroblast growth factor-21 (FGF21) is an important metabolic regulator with multiple beneficial effects on glucose homeostasis and lipid metabolism in animal models of diabetes and obesity (6,9). The finding that injection of LY2405319, an FGF21 variant, for 28 days significantly improved glucose homeostasis and alleviated dyslipidemia in patients with obesity and type 2 diabetes (15) has led to considerable recent interest in the physiological effects of FGF21 in humans.
Although the beneficial effects of circulating FGF21 are known, several cross-sectional studies have demonstrated a positive correlation between the circulating FGF21 level and body mass index (BMI) in humans (16,17). Animal models of obesity also exhibit elevated endogenous levels of FGF21 (10,11), suggesting that a high serum level of FGF21 may not be beneficial in the obese state. To address this discrepancy, previous animal studies have investigated the existence of FGF21 resistance in dietary-induced obese mice (10,11). In addition to elevated serum FGF21 levels, the dietary-induced obese mice display a lower level of FGF21 receptors and a severely impaired signaling response to FGF21 in the liver and
3
adipose tissue (10,11). Therefore, a chronically elevated serum level of FGF21 likely represents resistance to and reduction of the therapeutic effect of FGF21, which may lead to abnormal glucose and lipid metabolism. In fact, prospective studies have revealed that an elevated level of serum FGF21 precedes the development of type 2 diabetes in both European and Asian populations (12,13).
On the other hand, voluntary wheel running exercise for 36 weeks was recently reported to reduce the blood FGF21 level in a rat model of obesity (18). In that study, the level of FGF21 receptor expression in the liver was significantly higher in exercised Otsuka Long-Evans Tokushima Fatty rats than in sedentary Otsuka Long-Evans Tokushima Fatty rats, suggesting that regular exercise prevents FGF21 resistance in target organs (18). Although the effect of regular exercise on FGF21 resistance has not been evaluated in humans, the ability of regular exercise to increase CRF and reduce the risk for type 2 diabetes (2,19) and metabolic syndrome (20) is well documented. This suggests that a high CRF level might prevent FGF21 resistance and thus the onset of many chronic diseases. However, no association between the CRF level and the circulating FGF21 concentration has yet been documented. Furthermore, although visceral fat accumulation has been convincingly shown to be an independent predictor of metabolic risk factors (21), the contribution of visceral fat area (VFA) to the serum FGF21 level in humans is poorly understood.
Therefore, this study aimed to determine the relationship between the serum FGF21 concentration and the CRF level. Because obesity has been suggested to induce FGF21 resistance, we also examined whether the degree of obesity affects the relationship between the serum FGF21 concentration and the CRF level.
[2] Materials and Methods Subjects
The first group of study participants consisted of 166 Japanese men aged 30–79 years (middle-
4
aged and elderly). Subjects were originally recruited for a separate study examining the effects of aging and exercise on the relationship between genetic factors and metabolic syndrome risk (22); samples obtained during that study were reanalyzed in the present study. The participants had no history of diabetes or cardiovascular disease and were free of other chronic diseases such as cancer, chronic kidney failure, nonalcoholic steatohepatitis, and autoimmune disorders.
Eleven subjects (6.9%) were using lipid-lowering medication. Fourteen subjects (8.8%) with prediabetes [defined as fasting plasma glucose (FPG) levels of 110–125 mg/dL or glycated hemoglobin (HbA1c) levels of 5.7%–6.4%] were included. The current and/or former smoking status was recorded using a questionnaire. Daily alcohol intake was assessed using a brief, selfadministered diet history questionnaire.
To determine whether long-term endurance training influences the circulating FGF21 level, we also recruited a group of men aged 19–29 years consisting of 21 endurance-trained athletes (ET: rowers, n=10; triathletes, n=10; and cycle racers, n= 1) and 25 healthy untrained men (UT). None of these younger participants had any history of chronic disease or medication use. The ET participants regularly performed 1022 ± 399 minutes of endurance training per week.
All participants provided written informed consent to participate before enrolling in the study. This research project was approved by the Ethics Committee of Waseda University.
Anthropometric characteristics
Body weight and body fat percentage (as assessed by bioelectrical impedance analysis) were measured using an electronic scale (Inner Scan BC-600; Tanita, Inc), and height was measured with a stadiometer (YL-65; YAGAMI, Inc). BMI and fat free mass (FFM) were calculated from the measurements of body weight, body fat percentage, and height. The VFA and sc fat area (SFA) were measured by magnetic resonance imaging (MRI) (Signa 1.5 T; General Electric, Inc) as described previously (22). The imaging conditions included a T1-weighted spin-echo
5
and axialplane sequence with a slice thickness of 10 mm, a repetition time of 140 milliseconds, and an echo time of 12.3 milliseconds. The cross-sectional area of the VFA and SFA at the umbilical level was determined using image-analysis software (Slice-o-matic 4.3 for Windows;
Tomovision). The coefficient of variation for the cross-sectional area at the umbilical level was 0.4%.
Cardiorespiratory fitness
CRF was assessed via a maximal graded exercise test using a cycle ergometer (Ergomedic 828E; Monark; or Aerobike 75XLII; Combi) and quantified as the peak oxygen uptake (VO2peak). The graded cycle exercise began at a workload of 45–90 W, which was then increased by 15 W/min until the participant could no longer maintain a pedaling frequency of 60 rpm. The heart rate and rating of perceived exertion were monitored each minute during exercise. During the incremental portion of the exercise test, the participant’s expired gas was collected and the O2 and CO2 concentrations measured and averaged over 30-second intervals using an automated gas analyzing system (Aeromonitor AE-300; Minato Medical Science). The highest value of VO2 recorded during the exercise test was considered the VO2peak (mL·kg-1·min-1). Middle-aged and elderly subjects were subsequently divided into low- and high-CRF groups according to the median VO2peak value of each age group, as follows (in mL·kg-1·min-1): 30–39 years, 41.8; 40–49 years, 36.3; 50–59 years, 38.7; 60–64 years, 32.7;
65–69 years, 30.4; and 70–79 years, 28.4.
Collection and analysis of blood samples
The participants were instructed not to engage in any intensive exercise on the day previous to blood sampling. Blood samples were collected between 8:30 and 11:00 AM after a 12-hour overnight fast and then centrifuged at 3000 × g at 4°C for 15 minutes. The serum and plasma were collected and stored at -80°C until analysis. The serum enzymatic activities of aspartate
6
aminotransferase (AST), alanine aminotransferase (ALT), and γ-glutamyl transferase (γ-GTP), and the concentrations of total cholesterol (total-C), low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), triglycerides (TG), free fatty acid (FFA), apolipoprotein C-III (ApoCIII), fasting plasma glucose (FPG), insulin, and HbA1c were determined by BML, Inc. The homeostasis model assessment of insulin resistance (HOMA-IR) value was calculated from the fasting concentrations of plasma glucose and serum insulin as follows: HOMA-IR = [fasting glucose (mg/dL)] - [fasting insulin (μU/mL)]/405. The serum FGF21 concentration was determined using a commercially available ELISA kit (DF2100;
R&D Systems, Inc) according to the manufacturer’s instructions. The detectable range of the assay was 17.0–2410.9 pg/mL. The intra- and inter-assay coefficients of variation reported by the manufacturer were 2.9%–3.9% and 5.2%–10.9%, respectively.
Statistical analysis
All statistical analyses were performed using SPSS version 21.0 (SPSS, Inc). The Kolmogorov-Smirnov test was performed to assess the normality of data distribution, and several variables were log or square root transformed prior to analysis to obtain a normal distribution of values. A Student’s t test (for normally distributed data), the Mann-Whitney U test (for nonnormally distributed data) and the χ2 test (for categorical data) were used to evaluate the differences between the low- and high-CRF groups of middle-aged and elderly subjects and between the ET and UT groups of young subjects. Associations among the variables were detected using Pearson’s correlation coefficients. Partial correlation analysis adjusted for age and BMI was also performed.
We performed stepwise multiple linear regression analyses to identify the factors influencing FGF21 in the middle-aged and elderly subjects. Age, alcohol consumption, current or former smoking status, use of lipid-lowering medication, and the variables that showed significant partial correlation with FGF21 after adjustment for age and BMI were included in
7
the linear regression model. Because a considerable number of the young subjects had a serum FGF21 level below the limit of detection of the assay, a multiple logistic regression analysis was performed to assess the odds ratio (OR) and 95% confidence interval (CI) for undetectable serum FGF21 with respect to the VO2peak and VFA values. All measurements and calculated values are presented as the mean ± SD (for normally distributed variables) or median (interquartile ratio) (for nonnormally distributed variables), and the level of statistical significance was set at P < 0.05.
[3] Results
Comparison of the characteristics of the middle-aged and elderly participants between the low- and high-CRF groups
After exclusion of subjects with a serum FGF21 level below the limit of detection of the ELISA, as well as two subjects with an extremely high FGF21 level (1913.8 and 2410.9 pg/mL), a total of 160 participants were analyzed. The serum FGF21 level among these 160 subjects ranged from 17.0 to 628.9 pg/mL. Because the Kolmogorov-Smirnov test showed that the serum FGF21 level was normally distributed (P = 0.075) whereas the log-transformed FGF21 level was not normally distributed (P < 0.001), we used the raw FGF21 values in subsequent analyses.
VO2peak and the HDL-C level were higher in the high-CRF than in the low-CRF group, whereas BMI, body fat percentage, VFA, SFA, the TG, HbA1c, and insulin levels and HOMA-IR were lower in the high-CRF group than in the low-CRF group (Table 1, P < 0.05).
The serum FGF21 level was also significantly lower in the high-CRF group than in the low-CRF group (Table 1, P = 0.007).
8
Table 1. Characteristics of the middle-aged and elderly subjects
Variable Low CRF High CRF p*
N 80 80
Age (years) 62.0 ± 11.5 61.4 ± 11.9 0.746
Height (cm) 169.8 ± 6.7 170.7 ± 6.4 0.351
Body weight (kg) 69.7 ± 9.6 67.9 ± 8.6 0.233
BMI (kg/m2) 24.1 ± 2.5 23.2 ± 2.3 0.022
Body fat (%) 21.7 ± 4.6 19.3 ± 4.1 0.001
VFA (cm2) 121.9 ± 40.6 97.0 ± 42.7 <0.001
SFA (cm2) 125.5 ± 45.5 104.4 ± 45.3 0.004
FFM (kg) 54.3 ± 6.0 54.6 ± 5.6 0.755
VO2peak (mL·kg-1·min-1) 28.3 ± 5.2 37.4 ± 5.8 <0.001
AST (IU/L) 24.0 (20.0–27.0) 24.0 (21.0–28.8) 0.348
ALT (IU/L) 20.0 (16.0–27.0) 19.0 (15.0–24.0) 0.467
γ-GTP (IU/L) 29.0 (23.0–41.0) 26.0 (21.0–45.0) 0.354
Total-C (mg/dL) 214.3 ± 31.5 211.8 ± 34.8 0.633
HDL-C (mg/dL) 55.0 (49.0–63.8) 65.0 (55.0–71.8) <0.001
LDL-C (mg/dL) 127.0 ± 28.3 120.4 ± 28.2 0.143
TG (mg/dL) 101.5 (71.0–138.0) 78.0 (62.0–101.0) <0.001
FFA (mEq/L) 0.62 (0.42–0.77) 0.51 (0.40–0.72) 0.094
ApoC-III (mg/dL) 10.6 ± 3.8 10.0 ± 2.8 0.230
FPG (mg/dL) 96.8 ± 10.0 96.0 ± 8.1 0.559
HbA1c (%) 5.05 ± 0.30 4.91 ± 0.22 0.001
Insulin (µU/mL) 6.1 (4.2–8.2) 4.2 (3.2–6.3) <0.001
HOMA-IR 1.41 (0.99–1.96) 1.04 (0.73–1.48) 0.001
FGF21 (pg/mL) 277.5 ± 118.2 228.6 ± 107.0 0.007
Alcohol consumption (g/day) 14.1 (2.6–39.1) 23.3 (9.8–43.4) 0.884
Current or former smoking status (%) 58.8 38.8 0.017
Lipid-lowering medication use (%) 5.0 8.8 0.534
Prediabetes (%) 13.8 3.8 0.025
Data are the mean ± SD or median (IQR) values. Prediabetes was defined as FPG 110–125 mg/dL or HbA1c 5.7–6.4 %. *Low CRF vs. high CRF. Boldface indicates significance.
9
Associations of the serum FGF21 level with CRF, body composition, and metabolic parameters in middle-aged and elderly men
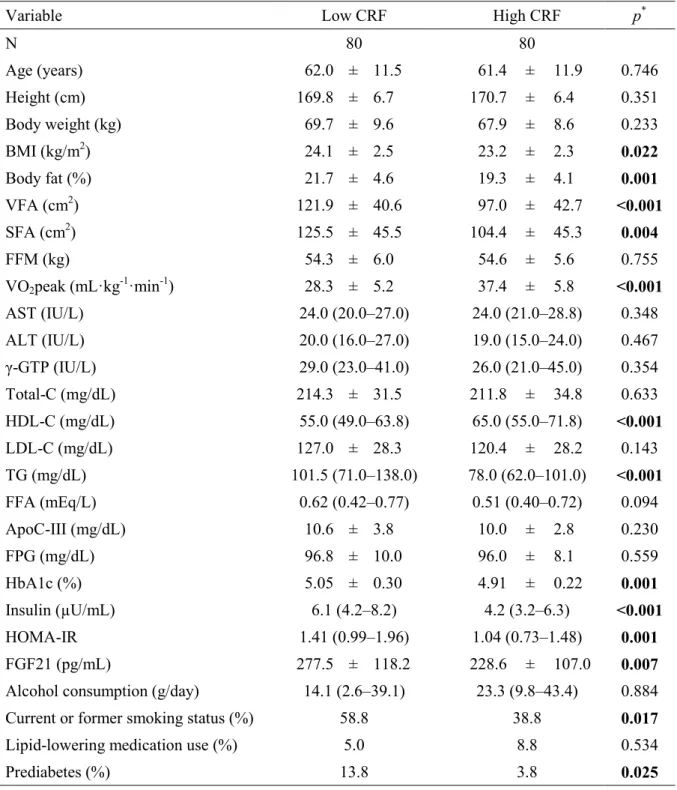
Table 2 presents the correlations of the serum FGF21 level with other variables. Our cross-sectional study showed that the serum FGF21 level correlated negatively with VO2peak (r
= 0.355, P < 0.001) (Figure 1A). This correlation remained significant after adjustment for age and BMI (r = 0.314, P < 0.001).
Furthermore, the serum FGF21 level correlated positively with VFA (r = 0.487, P <
0.001) (Figure 1B). This correlation remained significant after adjustment for age and BMI (r = 0.494, P < 0.001). Although SFA correlated with serum FGF21 level (r = 0.187, P = 0.018), this correlation was lost when adjusted for age and BMI. The serum FGF21 level also correlated positively with the ALT (r = 0.160, P = 0.044), γ-GTP (r = 0.257, P < 0.001), TG (r = 0.352, P <
0.001), and ApoC-III levels (r = 0.289, P < 0.001), even after adjustment for age and BMI.
Stepwise multiple regression analysis was performed to elucidate the independent predictors of the serum FGF21 level (Table 3). We included VFA, VO2peak, the ALT, γ-GTP, TG, and ApoC-III levels, age, alcohol consumption, current or former smoking status, and use of lipid-lowering medication as independent variables. In the best-fit model, VFA was most strongly associated with the serum FGF21 level (β = 0.360, P < 0.001), whereas VO2peak was an independent negative predictor of the serum FGF21 level (β = 0.174, P = 0.019). The serum TG level was also positively associated with the serum FGF21 level (β = 0.206, P = 0.005).
10
Table 2 Correlations of the serum FGF21 level with the other variables in middle-aged and elderly subjects
FGF21 FGF21
(age- and BMI-adjusted)
r p r p
Age (years) 0.144 0.069
BMI (kg/m2) 0.133 0.093
Body fat (%) 0.146 0.065 0.026 0.747
VFA (cm2) 0.487 <0.001 0.494 <0.001
SFA (cm2) 0.187 0.018 0.150 0.059
FFM (kg) 0.019 0.812 -0.028 0.727
VO2peak (mL·kg-1·min-1) -0.355 <0.001 -0.314 <0.001
AST (IU/L) 0.101 0.204 0.112 0.162
ALT (IU/L) 0.120 0.131 0.160 0.044
γ-GTP (IU/L) 0.245 0.002 0.257 0.001
Total-C (mg/dL) 0.061 0.441 0.045 0.572
HDL-C (mg/dL) -0.143 0.072 -0.137 0.085
LDL-C (mg/dL) 0.085 0.285 0.067 0.402
TG (mg/dL) 0.364 <0.001 0.352 <0.001
FFA (mEq/L) 0.090 0.257 0.076 0.342
ApoC-III (mg/dL) 0.303 <0.001 0.289 <0.001
FPG (mg/dL) 0.131 0.102 0.092 0.257
HbA1c (%) 0.134 0.093 0.083 0.306
Insulin (µU/mL) 0.038 0.635 -0.026 0.743
HOMA-IR 0.048 0.549 -0.025 0.761
Data are the Pearson’s correlation coefficients. The AST, ALT, γ-GTP, HDL-C, TG, FFA, Insulin, and HOMA-IR data were log-transformed prior to analysis. Boldface indicates significance.
11
Figure 1. Correlations of the serum FGF21 level with the VO2peak (A) and VFA (B) in middle-aged and elderly subjects.
Table 3. Stepwise multiple linear regression analysis of FGF21
Dependent variable Independent variable β p
FGF21 VFA (cm2) 0.360 <0.001
TG (mg/dL) 0.206 0.005
VO2peak (mL·kg-1·min-1) -0.174 0.019 β: Standardized coefficient. The model was adjusted for age, alcohol consumption, current or former smoking status, and use of lipid-lowering medication. TG was log-transformed prior to analysis. Alcohol consumption was square root-transformed prior to analysis. Model r2 = 0.327, P
< 0.001. Boldface indicates significance.
12
Effect of endurance training on the serum FGF21 level in young men
The VO2peak was higher in ET men than in healthy UT men (P < 0.001), whereas age, VFA, and SFA were lower in the ET than in the UT group (Table 4, P < 0.001). Surprisingly, 21 (45.7%) subjects in the young group had a serum FGF21 level below the limit of detection of the assay. Therefore, we compared subjects from the ET and UT groups for whom FGF21 was not detected. Although measurable serum FGF21 levels did not differ significantly between the groups (P = 0.106), more subjects in the ET than in the UT group had levels of serum FGF21 that were below the limits of detection (Table 4, P = 0.001).
Because VO2peak and VFA were independent predictors of the serum FGF21 level in the middle-aged and elderly men, we performed multiple logistic regression analysis to examine whether the extremely low levels of serum FGF21 among the ET participants were due to a high VO2peak or low VFA. After adjustment for age, VFA, and TG, the OR for a serum FGF21 level below the limit of detection increased significantly by 1.114 (95% CI 1.001–1.239, P = 0.047) per increment of VO2peak value (mL·kg-1·min-1) (Table 5). After adjustment for age, VO2peak, and TG, VFA was also independently associated with the occurrence of a serum FGF21 level below the limit of detection (OR 0.938, 95% CI 0.882–0.998, P = 0.044).
13 Table 4. Characteristics of young subjects
Variables Healthy untrained Endurance trained p*
N 25 21
Age (year) 23.8 ± 2.3 20.9 ± 1.0 < 0.001
Height (cm) 172.9 ± 5.6 173.1 ± 5.3 0.874
Body Weight (kg) 70.2 ± 8.4 68.1 ± 6.4 0.354
BMI (kg/m2) 23.5 ± 2.9 22.7 ± 1.6 0.246
VFA (cm2) 43.0 ± 19.7 21.5 ± 12.0 < 0.001
SFA (cm2) 112.3 ± 67.4 48.9 ± 20.1 < 0.001
VO2peak (mL·kg-1·min-1) 43.6 ± 5.3 59.6 ± 4.2 < 0.001
AST (IU/L) 21.0 (18.0–29.5) 27.0 (21.0–36.0) 0.024
ALT (IU/L) 18.0 (13.0–28.0) 23.0 (17.5–35.5) 0.120
γ-GTP (IU/L) 22.0 (17.5–31.0) 20.0 (15.5–23.5) 0.164
Total-C (mg/dL) 179.5 ± 30.0 169.8 ± 24.3 0.239
HDL-C (mg/dL) 57.0 (50.5–66.6) 63.0 (52.0–72.5) 0.216
TG (mg/dL) 67.0 (58.0–90.0) 53.0 (40.5–69.5) 0.076
FFA (mEq/L) 0.51 (0.39–0.81) 0.44 (0.37–0.57) 0.298
FPG (mg/dL) 84.3 ± 5.6 84.3 ± 4.8 0.973
HbA1c (%) 4.89 ± 0.28 4.90 ± 0.26 0.881
Insulin (µU/mL) 4.3 (3.4–5.7) 4.0 (3.4–5.3) 0.544
HOMA-IR 0.92 (0.71–1.28) 0.84 (0.70–1.15) 0.627
FGF21 (pg/mL) 219.9 (73.4–337.9) 74.2 (22.4–191.3) 0.106
FGF21 levels below the detection
limit of the assay (%) 24.0 71.4 0.001
Data are mean ± SD or median (IQR) values. *Healthy untrained vs. Endurance trained. Boldface indicates significance.
Table 5. Odds ratio for a serum FGF21 level below the limit of detection of the assay with respect to the VFA and VO2peak
OR 95% CI p
VFA (cm2) 0.938 0.882–0.998 0.044
VO2peak (mL·kg-1·min-1) 1.114 1.001–1.239 0.047 Data are the OR (95%CI). The model was adjusted for age and TG. TG was log-transformed prior to analysis. Boldface indicates significance.
14 [4] Discussion and Conclusion
The present study revealed that CRF is negatively associated with the serum FGF21 level. In contrast, a higher circulating FGF21 level was associated with greater visceral at content. This was a first investigation of the relationship of CRF on the serum FGF21 concentration in humans.
FGF21 is reported to be expressed primarily in the liver (23). Because hepatic expression of FGF21 is directly regulated by peroxisome proliferator-activated receptor (PPAR) -α, which is up-regulated by fatty acids (24), serum FGF21 level in humans is increased by both overfeeding (25) and fasting (26). FGF21 signaling has been investigated at the molecular level in vitro and in vivo and appears to act via cell surface classic FGF receptors complexed with β-klotho (27-29). β-Klotho is expressed mainly in the liver and adipose tissue (27), consistent with the pattern of FGF21 activity. FGF21 has been shown to stimulate hepatic fatty acid oxidation (30,31), adipose tissue glucose uptake (6), adipose thermogenic gene expression, and browning of white adipose tissue (32). Therefore, the therapeutic efficacy of FGF21 is likely affected by the expression of tissue-specific FGF receptors.
Recent animal studies have suggested the existence of FGF21 resistance, and obese rats with FGF21 resistance exhibit elevated serum FGF21 accompanied by reduction of FGF receptor expression in the liver and adipose tissue (10,11). A high level of circulating FGF21 has also been documented in obese people (16,17,33), and elevated serum FGF21 has been reported to be an independent predictor of type 2 diabetes (12,13) and metabolic syndrome (13). It is therefore plausible that chronic disease is induced by FGF21 resistance-mediated metabolic decline, whereas a lower basal FGF21 concentration suggests sensitivity to FGF21 and thus resistance to chronic diseases. Because regular exercise was reported to prevent FGF receptor down-regulation and serum FGF21 level elevation in obese model rats (18), the present study examined the relationship between the fitness level and serum FGF21 concentration in humans.
In agreement with the animal study, the serum FGF21 level was significantly lower in the high
15
than in the low fitness group (Table 1). Furthermore, the circulating FGF21 concentration correlated negatively with the CRF level (Figure 1A). These results suggest that regular exercise that increases aerobic capacity also improves FGF21 sensitivity in humans as seen in rodents.
The previous animal study did not eliminate the effect of obesity on FGF21 resistance because the body weight and epididymal fat mass were significantly lower in the trained than in the sedentary rats (18). However, our stepwise multiple linear regression analysis showed CRF to be a significant predictor of the serum FGF21 level independent of the VFA in middle-aged and elderly subjects (Table 3), suggesting that higher aerobic capacity itself is associated with greater FGF21 sensitivity. Therefore, the benefits of regular exercise may be due in part to prevention of FGF21 resistance.
Although obesity is known to be associated with elevated circulating FGF21 level in humans (16,17,33), the association between body fat distribution and the FGF21 level has not been fully elucidated. Our data clearly showed that the serum FGF21 level was the most robustly associated with VFA among the measures of adiposity such as BMI, body fat percentage, and SFA (Table 2). The multiple linear regression analyses in both older and young subjects suggested that VFA is the predominant determinant of the serum FGF21 level, although CRF was also independently associated with serum FGF21 (Table 3). Several studies have reported that waist circumference and waist to hip ratio, both indices of abdominal obesity, were no longer associated with the circulating FGF21 level after adjustment for BMI (16,34).
However, these simple anthropometric measures do not always reflect the VFA (35). In the present study, adjustment for BMI did not affect the association between VFA measured by MRI and the serum FGF21 level (Table 2). Therefore, a chronically high circulating FGF21 level is probably induced by visceral rather than overall obesity. Although the release of inflammatory adipokines and FFA from excess VFA is recognized as the primary mechanism linking VFA and metabolic abnormalities (21,36), FGF21 resistance is another possible explanation for the important role of VFA in the development of various chronic diseases.
16
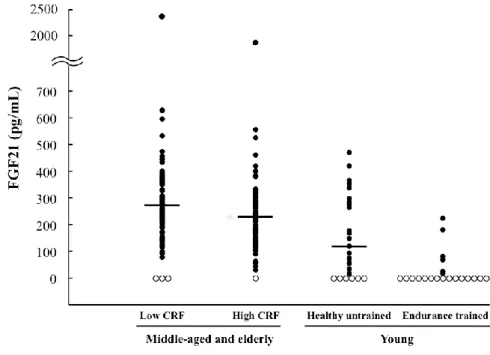
To understand further the effect of CRF and VFA on the serum FGF21 level, we recruited healthy untrained and endurance-trained individuals. As shown in Table 4 and Figure 2, the proportion of participants with a serum FGF21 level below the limit of detection was significantly higher in the ET than in the UT participants (71.4% vs 24.0%, P < 0.001).
Furthermore, the median serum FGF21 concentration was higher in the middle-aged and elderly subjects with high CRF than in the young untrained subjects (231.8 pg/mL vs 118.8 pg/mL, P = 0.019). This can mainly be explained by an age-related increase in VFA (Table 6). Consistent with the result in middle-aged and elderly participants, our multiple logistic regression analysis in the young subjects revealed that the CRF and VFA were independently associated with an undetectable serum FGF21 level (Table 5, P < 0.05), supporting our hypothesis that the CRF level is independently associated with FGF21 sensitivity. A previous German study also reported that there were many undetectable levels of serum FGF21 and found a more favorable metabolic profile in the subjects with FGF21 concentrations below the limit of detection than in those with measurable circulating FGF21 (37). Therefore, an undetectable level of FGF21 may be common in both European and Asian populations and may indicate greater sensitivity to FGF21.
Acute exercise training or a short-term supervised physical activity program has been shown to increase the serum FGF21 level (5,38). These previous studies consistently observed that exercise increased the circulating FFA level and concluded that the exercise-induced increase in FFA might explain the induction of FGF21. However, long-term aerobic exercise increases CRF and tends to reduce the release of FFA from adipose tissue during submaximal exercise (39). In the present study, CRF was not actually associated with the serum FFA level after adjustment for age and BMI (Table 2, r = 0.073, P = 0.369). Therefore, short-term and long-term exercise seem to have completely different effects on the serum FGF21 concentration response.
17
Figure 2. Distribution of the serum FGF21 concentration in young, middle-aged, and elderly subjects. Closed circles (●) indicate detectable serum FGF21 levels, and open circles (○) indicate levels below the limit of detection. Horizontal lines represent the median values (including undetectable serum FGF21 levels, which were considered zero). Levels for the middle-aged and elderly men: low CRF, n = 84, 266.7 pg/mL; high CRF, n = 82, 231.8 pg/mL; young men: healthy untrained, n = 25, 118.8 pg/mL; endurance trained, n = 21 (median value was below the limit of detection). The six middle-aged and elderly subjects excluded from the statistical analyses due to an undetectable serum FGF21 level or an extremely high FGF21 level are included here.
Table 6. Multiple linear regression analysis of FGF21 in middle-aged and elderly subjects with high CRF and untrained young subjects with detectable FGF21 (n = 99)
Dependent variable Independent variable β p
FGF21 VFA (cm2) 0.554 <0.001
VO2 peak (mL·kg−1·min−1) −0.243 0.029 β: Standardized coefficient. The model was adjusted for age. Model r2 = 0.313, P <
0.001. Boldface indicates significance.
18
In accordance with previous studies (16,34,40), we showed that the serum FGF21 level correlates with the serum levels of TG and hepatic enzymes, including ALT and γ-GTP (Table 2), that are well-accepted biochemical markers of liver injury. Obese mice exhibit decreased hepatic FGF21 sensitivity (10), and human patients with non-alcoholic fatty liver disease have high levels of circulating FGF21 as well as elevated ALT or γ-GTP levels (41,42).
The elevation of serum levels of TG and hepatic enzymes in the FGF21-resistant state is consistent with such findings. We also revealed that the level of ApoC-III, a key regulator of TG metabolism, correlated positively with the serum FGF21 level (Table 2). ApoC-III decreases the clearance of circulating TG by inhibiting lipoprotein lipase activity (43). Interestingly, the activation of PPAR-α, a primary regulator of FGF21, decreases ApoC-III expression (44).
Administration of FGF21 or a variant decreased the circulating levels of both ApoC-III and TG in monkeys and humans (9,15), suggesting that a marked reduction in ApoC-III might contribute to the decrease in TG. Therefore, the association between the serum TG and FGF21 levels may be mediated in part by up-regulation of ApoC-III induced by FGF21 resistance.
The present study has several limitations. First, our sample size was relatively small, which might have led to type 2 error. Second, we did not evaluate hepatic fat content despite a previous study that showed it to be independently associated with the circulating FGF21 level (45). The association between the CRF and serum FGF21 levels should be re-examined with hepatic fat content as a covariate. Finally, most the participants in this study were free from chronic diseases. Further investigation is needed to clarify whether a high CRF level is also associated with a low serum FGF21 level, even in patients with chronic diseases such as type 2 diabetes and cardiovascular disease.
In conclusion, the present study revealed that CRF is negatively associated with the serum FGF21 level in middle-aged and elderly Japanese men. We also demonstrated that young ET men have extremely low levels of serum FGF21. These findings provide novel evidence that regular aerobic exercise and a high CRF level improve health by combatting an underlying
19 resistance to FGF21.
3. Endurance exercise reduces hepatic fat content and serum fibroblast growth factor 21 levels in elderly men
[1] Introduction
Ectopic fat accumulation in the liver increases the risk of developing type 2 diabetes mellitus and cardiovascular disease (46-48) and is observed in elderly subjects (49). The underlying mechanisms between the age-related increase in hepatic fat and the onset of cardiometabolic diseases have not been fully understood. However, recent studies have suggested that hepatic fat accumulation impairs the effects of FGF21 and thereby increases the risk for these chronic diseases in elderly people.
The hepatokine FGF21 plays a therapeutic role in obesity and metabolic homeostasis (50), whereas a high level of FGF21 at rest has been considered to represent a FGF21-resistant state that indicates impaired FGF21 response to target tissues (10,11). It was reported that higher hepatic fat content was strongly associated with higher serum FGF21 levels(42,45,51), and the FGF21 resistance was exacerbated in older individuals (52,53). Moreover, the elevated circulating FGF21 level was an independent predictor of type 2 diabetes (12,13) and
cardiovascular disease (14). These results therefore suggest that FGF21 resistance caused by hepatic fat accumulation is an important risk factor for cardiometabolic diseases, and it is likely that the risk of FGF21 resistance increases with age.
Previous studies revealed that a higher level of CRF acquired by regular endurance exercise was associated with both lower hepatic fat content (54,55) and serum FGF21 level (53).
Although these results suggest that endurance exercise modulates hepatic fat content and FGF21 resistance, whether the exercise-induced change in hepatic fat substantially affects serum FGF21 concentration has not been evaluated. Moreover, there is no clear evidence that
20
endurance exercise modulates age-related hepatic fat accumulation and FGF21 resistance in elderly subjects. Because Asian populations have a high risk of hepatic fat accumulation and FGF21 resistance despite lower BMI (56-58), it is also important to examine whether endurance exercise reduces hepatic fat content and FGF21 resistance regardless of obesity status.
Therefore, the purpose of this study was to evaluate whether endurance exercise modulates hepatic fat content, thereby resulting in improvement of the FGF21-resistant state in elderly Japanese men. This study provides key insights into how endurance exercise prevents the onset of cardiometabolic diseases that are associated with FGF21 resistance.
[2] Materials and Methods Subjects
Thirty-three elderly Japanese men (aged 62–76 years) were recruited from temporary
employment agencies. The subjects had no diagnosis of diabetes or cardiovascular disease and were free of hepatic diseases such as hepatic cancer, hepatitis, and non-alcoholic steatohepatitis;
but two subjects had high levels of fasting glucose (≥126 mg/dL) at baseline. Four subjects (12.1%) were using lipid-lowering medication, and their medication status was not changed throughout the study. All participants provided written informed consent to participate before enrolling in the study, which was approved by the Ethics Committee of Waseda University. The study was conducted in accordance with the Declaration of Helsinki.
Study Design
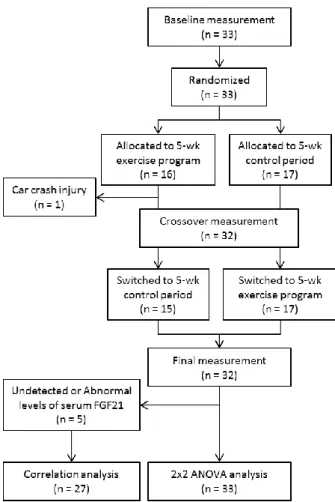
As shown in Figure 1, the study was a randomized crossover trial. The subjects were randomized by an independent observer to either a 5-week endurance exercise program or a control period during which they were instructed not to undertake any formal exercise or change their level of general physical activity. After 5 weeks, the subjects switched to the other regimen for 5 weeks. All subjects were free-living and were instructed to consume their habitual diet
21
throughout the study. Measurements were performed to determine cardiorespiratory fitness, hepatic fat content, VFA, SFA, other anthropometric characteristics and blood biochemical parameters at baseline (0 weeks), crossover (5 weeks) and at the end of the study (10 weeks).
The measurements were performed at least three days after the previous exercise session in order to exclude the acute effects of exercise. Subjects refrained from medication on the morning of each measurement. Diet and nutritional intake were also assessed using a brief, self-administered diet history questionnaire. Daily physical activity, excluding the exercise program, was assessed using the International Physical Activity Questionnaire and expressed as metabolic equivalent minutes per week.
Figure 3. Flow diagram of the participants in the randomized crossover trial.
22 Cardiorespiratory Fitness
CRF was assessed via a maximal graded exercise test using a cycle ergometer (Aerobike 75XLII; Combi Wellness, Inc) and quantified as the maximal oxygen uptake (VO2max). The graded cycle exercise began at a workload of 40–60 W, which was then increased by 30 W every 3 min until exhaustion. Heart rate and rating of perceived exertion were monitored each minute of measurement. During the exercise test, the participant’s expired gas was collected and the O2 and CO2 concentrations were measured and averaged over 30-second intervals using an automated gas analysing system (Aeromonitor AE-310S; Minato Medical Science). The highest value of VO2 recorded during the exercise test was considered the VO 2max (mL·kg-1·min-1).
Endurance Exercise program
Participants undertook a supervised, progressive, 5-week aerobic exercise program, which met current guidelines for adult prescribed exercise (59). Exercise comprised of three cycle
ergometer sessions per week. Exercise intensity gradually increased such that subjects exercised at a power output designed to elicit 60% of pre-exercise VO2max for week 1, 70% for weeks 2 and 3, and 75% for weeks 4 and 5. The exercise time was 30 minutes for week 1 and 2, and 45 minutes for week 3 to 5. At the first exercise session and when exercise intensity was increased, the exercise intensity was confirmed by measuring O2 consumption for 5 min, beginning 5 min after the start of exercise.
Anthropometric Characteristics
Body weight and body fat percentage (as assessed by bioelectrical impedance analysis) were measured using an electronic scale (Inner Scan BC-600; Tanita, Inc), and height was measured with a stadiometer (YL-65; YAGAMI, Inc). BMI was calculated from measurements of height and body weight.
23 Magnetic resonance imaging and spectroscopy
Hepatic 1H magnetic resonance spectroscopy (MRS) was performed using 1.5 T whole-body scanner (Signa 1.5 T; General Electric, Inc) with 8-channel body array coil. A single-voxel (30×30×20 mm3) for spectroscopy was located in the right lobe of liver, avoiding major blood vessels. Voxel shimming was performed to optimize the homogeneity of the magnetic field. The proton spectra of liver were acquired using the point-resolved spectroscopy technique (TR = 3000 milliseconds, TE = 35 milliseconds, 64 measurements, 1024 sample points) Acquisition was synchronized to the respiratory cycle and triggered at the end of expiration. MRS data were quantified using LCModel version 6.3. Fat content was defined as the signal intensities at 0.9, 1.3 and 1.6 ppm relative to the signal intensity of water at 4.7 ppm. The coefficient of variation between measurements was 2.0%.
The VFA and SFA were measured by MRI as described previously (53). The imaging conditions included a T1-weighted spin-echo and axial plane sequence with a slice thickness of 10 mm, a repetition time of 140 milliseconds and an echo time of 12.3 milliseconds. The cross-sectional area of the VFA, SFA at the umbilical level, was determined using
image-analysis software (Slice-o-matic 4.3 for Windows; Tomovision). The coefficient of variation for the cross-sectional area at the umbilical level was 0.4%.
Blood Sampling and Analysis
Blood samples were collected after an overnight fast and then centrifuged at 3000 × g at 4°C for 15 min. Serum and plasma were collected and stored at ˗80°C until analysis. The serum
enzymatic activities of AST, ALT and γ-GTP and the concentrations of TG, FFA and FPG were determined by BML, Inc. The serum FGF21 concentration was determined using a
commercially available ELISA kit (DF2100; R&D Systems, Inc) according to the
manufacturer’s instructions. The intra- and inter-assay coefficients of variation reported by the manufacturer were 2.9%–3.9% and 5.2%–10.9%, respectively.
24 Statistics
All statistical analyses were performed using SPSS version 22.0 (SPSS, Inc.). The Kolmogorov-Smirnov test was performed to assess the normality of data distribution, and non-normally distributed data were log-transformed prior to analysis. Changes in VO2max, hepatic fat content, VFA, SFA, and other anthropometric and blood biochemical variables during the exercise program and control period were compared using 2×2 crossover ANOVA.
The model consisted of the within-subject factor “treatment” (exercise, control) and the
between-subject factor “sequence” (exercise–control, control–exercise). Differences in physical activity level and dietary intakes at baseline, 5 weeks and 10 weeks were assessed by repeated one-way ANOVA (for normally distributed data) or Friedman’s test (for non-normally
distributed data). Relationships between changes in all variables during exercise and control periods were determined by Pearson’s correlation coefficients. All measurements and calculated values are presented as the means ± standard deviation (for normally distributed data) or median (interquartile range) (for non-normally distributed data) values. The level of statistical
significance was set at P < 0.05.
[3] Results
Baseline characteristics of the subjects are shown in Table 7. One subject did not complete the exercise program because he was injured in a car crash that was not related to this study. The FGF21 data from five subjects were not used because they exhibited extreme levels of serum FGF21 (FGF21 level beyond the range of 31.3–2000 pg/mL) or changes in serum FGF21 level exceeding 2 standard deviations from the baseline FGF21 level during intervention. Therefore, full data were obtained for a total of 27 participants. The missing values were replaced by mean values, and 2×2 crossover ANOVA was performed using the data from 33 participants (Figure 3).
25
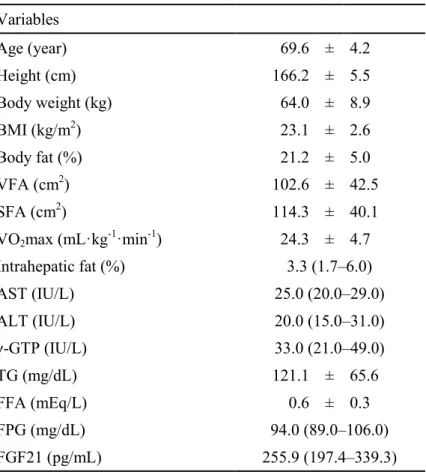
Table 7. Baseline characteristics of the subjects (n = 33)
Variables
Age (year) 69.6 ± 4.2
Height (cm) 166.2 ± 5.5
Body weight (kg) 64.0 ± 8.9
BMI (kg/m2) 23.1 ± 2.6
Body fat (%) 21.2 ± 5.0
VFA (cm2) 102.6 ± 42.5
SFA (cm2) 114.3 ± 40.1
VO2max (mL·kg-1·min-1) 24.3 ± 4.7
Intrahepatic fat (%) 3.3 (1.7–6.0)
AST (IU/L) 25.0 (20.0–29.0)
ALT (IU/L) 20.0 (15.0–31.0)
γ-GTP (IU/L) 33.0 (21.0–49.0)
TG (mg/dL) 121.1 ± 65.6
FFA (mEq/L) 0.6 ± 0.3
FPG (mg/dL) 94.0 (89.0–106.0)
FGF21 (pg/mL) 255.9 (197.4–339.3)
Data are mean ± standard deviation or median (interquartile range) values.
26
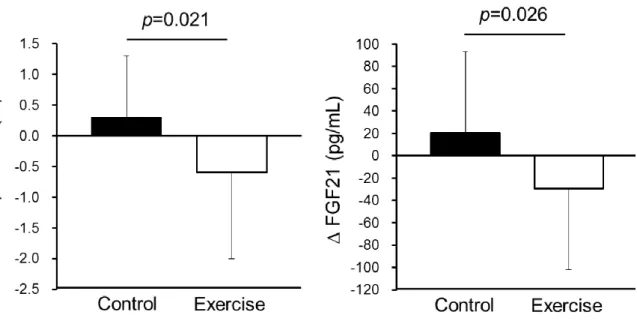
The changes in variables during the 5-week endurance exercise and control periods are summarized in Table 8. We compared the absolute values of changes in each variable between the exercise and control periods using 2×2 crossover ANOVA for the factors, treatment and sequence. The 5-week endurance exercise increased VO2max, and the absolute change in VO2max was higher in the exercise period than in the control period (P < 0.001). In contrast, the 5-week endurance exercise decreased hepatic fat content and serum FGF21 levels, and absolute changes in the exercise period were higher than in the control period (P = 0.026 and P = 0.021, respectively; Figure 4 and Table 8). We did not detected a significant interaction between treatment and sequence for VO2max, intrahepatic fat and serum FGF21 level,
indicating no carryover effects due to the sequence of experiment.
We also examined daily physical activity level and dietary intakes in study subjects at baseline, after 5 weeks and at the final measurements. Physical activity level, total energy intake, alcohol intake and macronutrient intakes were not statistically changed during the 10-week intervention in both the immediate- and delayed-exercise groups (Data not shown).
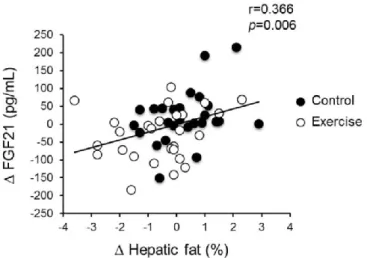
To identify the factors determining the change in serum FGF21 levels during intervention, we examined the correlations between absolute changes in each variable during exercise and control periods. As shown in Table 9 and Figure 5, only the change in hepatic fat content was significantly and positively correlated with changes in serum FGF21 levels (r = 0.366, P = 0.006). Changes in other anthropologic and blood biochemical parameters were not significantly correlated with changes in serum FGF21 levels.
27 Table 8. Changes in the variables during 5-week control and training periods
Control Exercise
Pre Post Δ Pre Post Δ p*
Body weight (kg) 64.1 ± 9.0 64.1 ± 8.6 0.0 ± 0.9 64.0 ± 8.8 64.0 ± 9.1 0.0 ± 0.8 0.881 BMI (kg/m2) 23.1 ± 2.5 23.1 ± 2.5 0.0 ± 0.4 23.1 ± 2.5 23.0 ± 2.6 –0.1 ± 0.4 0.750 Body fat (%) 20.9 ± 5.2 21.2 ± 5.3 0.3 ± 2.0 21.4 ± 5.3 20.8 ± 4.9 –0.6 ± 2.6 0.346 VO2max (mL·kg-1·min-1) 26.3 ± 4.4 25.0 ± 4.3 –1.0 (–1.8–0.0) 24.5 ± 4.1 27.1 ± 4.6 2.1 (0.8–3.7) < 0.001 VFA (cm2) 102.3 ± 44.0 103.5 ± 44.7 1.6 (–4.8–5.8) 103.3 ± 43.2 102.5 ± 44.5 0.2 (–6.6–6.0) 0.816 SFA (cm2) 115.3 ± 41.8 112.7 ± 39.0 –2.5 ± 14.7 114.4 ± 37.8 114.6 ± 42.7 0.2 ± 18.8 0.440 Intrahepatic fat (%) 2.7 (1.5–5.6) 2.9 (1.8–6.4) 0.3 ± 1.2 2.9 (1.7–6.5) 2.3 (1.3–5.4) –0.6 ± 1.4 0.021 AST (IU/L) 26.6 ± 8.6 28.2 ± 10.2 1.6 ± 4.6 28.5 ± 13.8 26.5 ± 8.7 –2.0 ± 7.5 0.067 ALT (IU/L) 20.0 (16.0–28.5) 19.0 (15.5–28.0) –1.0 (–3.5–2.0) 19.0 (16.0–31.5) 19.0 (15.5–25.5) –2.0 (–4.5–1.5) 0.416 γ-GTP (IU/L) 32.0 (21.5–46.0) 29.0 (20.0–47.0) –2.0 (–4.0–1.4) 33.0 (21.0–51.5) 31.0 (21.5–42.5) –2.0 (–5.5–1.0) 0.447 TG (mg/dL) 115.5 ± 63.8 115.9 ± 74.4 0.4 (–21.5–19.5) 122.2 ± 74.1 111.7 ± 66.2 –6.0 (–19.0–11.0) 0.843 FFA (mEq/L) 0.6 ± 0.2 0.5 ± 0.2 –0.1 ± 0.2 0.6 ± 0.2 0.6 ± 0.2 0.0 ± 0.2 0.721 FPG (mg/dL) 94.0 (88.0–106.5) 96.0 (91.0–107.5) 1.3 ± 6.0 97.0 (91.5–108.5) 99.0 (93.0–106.5) –0.7 ± 6.4 0.267 FGF21 (pg/mL) 231.6 ± 81.6 252.0 ± 98.5 20.4 ± 65.5 248.1 ± 88.5 218.5 ± 94.2 –29.6 ± 65.1 0.026 Data are mean ± standard deviation (for normally distributed data) or median (interquartile range) (for non-normally distributed data) values. Non-normally distributed data were log-transformed prior to analysis. Boldface indicates significance (P < 0.05). *P-values were obtained by 2×2 crossover ANOVA for the factor treatment.
28
Figure 4. Changes in hepatic fat content and serum FGF21 level during the control and exercise periods. Data are reported as mean ± standard deviation.
29
Table 9. Correlation of changes in serum FGF21 level and other variables.
Variables ∆ FGF21
r p
∆ Body weight (kg) 0.065 0.641
∆ BMI (kg/m2) 0.036 0.795
∆ Body fat (%) 0.223 0.105
∆ VO2max (mL·kg-1·min-1) -0.179 0.196
∆ VFA (cm2) -0.184 0.183
∆ SFA (cm2) -0.030 0.828
∆ Intrahepatic fat (%) 0.366 0.006
∆ AST (IU/L) 0.166 0.229
∆ ALT (IU/L) 0.111 0.423
∆ γ-GTP (IU/L) -0.147 0.288
∆ TG (mg/dL) -0.250 0.068
∆ FFA (mEq/L) -0.036 0.795
∆ FPG (mg/dL) 0.073 0.601
Data are Pearson’s correlation coefficients. VO2max, VFA, ALT, γ-GTP and TG were converted to log scale. Boldface indicates significance.
Figure 5. Correlation of changes in hepatic fat content and serum FGF21 level during the control and exercise periods.
30 [4] Discussion and Conclusion
The main findings of the present study were 5-week endurance exercise intervention decreased hepatic fat content without weight loss in elderly Japanese men, and change in serum FGF21 level was positively correlated only with change in hepatic fat content. These findings suggest that chronic endurance exercise improves FGF21 resistance by decreasing hepatic fat content regardless of obesity status and age. Age-related accumulation of hepatic fat content is strongly associated with metabolic disorder even in non-obese subjects (49,56,57), and elevated serum FGF21 level was strong predictor of cardiometabolic diseases (12-14). Therefore, our finding may contribute to prevention of age-related cardiometabolic diseases in non-obese elderly people.
Previous cross-sectional studies suggested that serum FGF21 level was independently associated with CRF, BMI, VFA and hepatic fat content (41,42,51,53). Only a study conducted by Yang et al. reported that a 3-month combined exercise program (endurance exercise + resistance exercise) significantly decreased serum FGF21 level in overweight middle-aged women (60). Although their study subjects had a significant loss in BMI during intervention, Yang et al. did not evaluate CRF, VFA and hepatic fat content, which may be key determinants of circulating FGF21 level. Therefore, evidence was lacking about the factors regulating the exercise-induced change in serum FGF21 level. In the present study, we showed that, although 5-week endurance exercise increased VO2max and decreased hepatic fat content, the reduction in serum FGF21 levels was significantly associated with a decrease in the hepatic fat content, but not changes in VO2max and other adiposity indices such as body fat percentage, VFA, and SFA. This result suggests that hepatic fat content is a dominant factor determining short-term exercise-induced reduction in serum FGF21 levels.
FGF21 is predominantly expressed in liver (23), and it stimulates hepatic fatty acid oxidation (30), adipose tissue glucose uptake (6), adipose thermogenic gene expression and browning of white adipose tissue (32) via classic FGF receptors complexed with β-klotho in an
31
endocrine or autocrine/paracrine manner (27-29,61). Because it has been documented that impairments of FGF21 receptors in the liver of obese mice have important roles in FGF21 resistance development (10,11), hepatic fat accumulation might also impair beneficial effects of FGF21 in the liver and white adipose tissue, leading to an increased cardiometabolic risk.
Interestingly, an animal study reported that drug-induced hepatic fat reduction enhanced
phosphorylation of hepatic AMP-activated protein kinase (AMPK), which was highly correlated with reduction of circulating FGF21 level (62). Another study demonstrated that vegetable extract reduced hepatic triglyceride as well as increased AMPK/sirtuin (SIRT) 1 signaling and FGF receptors 1, 3, and 4 (63). These results were in agreement with exercise-induced
beneficial effects in the liver. Indeed, chronic endurance exercise was reported to induce AMPK activation and SIRT1 expression in the liver of obese mice (64). Therefore, it is likely that endurance exercise modulates hepatic fat content and AMPK/SIRT1 signaling, thereby improving FGF21 resistance, which may be partly due to upregulation of FGF receptors.
Although we previously reported that high CRF was significantly associated with lower serum FGF21 levels independently of indices of adiposity such as VFA in middle-aged and elderly Japanese men (53), the present study demonstrated that endurance exercise-induced decrease in serum FGF21 level was determined by the reduced hepatic fat content rather than the elevated CRF in elderly men. Several studies reported that high CRF was independently associated with low hepatic fat content (54,55), and we also confirmed that endurance exercise modulated both CRF and hepatic fat content. Therefore, the association between CRF and serum FGF21 level in our previous cross-sectional study might be in part mediated by hepatic fat content. Although elevation of CRF caused by short-term intervention was not associated with decrease in serum FGF21 level in the present study, epidemiologic studies indicated that long-term maintenance of high CRF level is strongly associated with reduced risk of
cardiometabolic diseases (65,66), which may be in part explained by the prevention of FGF21 resistance. Therefore, we cannot rule out the possibility that long-term maintenance of high
32
CRF level by endurance exercise improves FGF21 resistance and prevents onset of
cardiometabolic diseases. Long-term exercise intervention or epidemiologic studies focusing on longitudinal change in CRF and circulating FGF21 level are needed to address this issue.
The present study has several limitations. First, our sample size was relatively small, which might have led to type 2 error. Second, our study included only male subjects.
Hamaguchi et al. reported that incidence of nonalcoholic hepatic steatosis was sharply increased with aging particularly in premenopausal women (67). Therefore, further investigation is needed to confirm our findings in older women. Third, we used a short-term endurance exercise
program to examine the changes in hepatic fat and serum FGF21 level without weight loss. It is unclear whether long-term prospective cohort studies support our finding that endurance exercise and/or physical activity can prevent a FGF21-resistant state.
In conclusion, the present study revealed that a 5-week endurance exercise program decreased hepatic fat content and serum FGF21 level without weight loss in elderly Japanese men and that an endurance exercise-induced reduction of serum FGF21 level was mediated by reduction of hepatic fat content. These findings suggest that endurance exercise improves FGF21 resistance without weight loss and provide new evidence that chronic exercise itself but not weight loss has beneficial effects on the prevention of age-related cardiometabolic diseases.
4. Conclusion and suggestions for future research
The results of the present cross-sectional study revealed that elevated serum FGF21 levels were positively associated with central adiposity, whereas high CRF levels were related to lower serum FGF21 concentrations. Furthermore, the five-week endurance exercise program significantly reduced hepatic fat content in elderly men, which was associated with decreased serum FGF21 levels (68). These results suggest that higher CRF and regular endurance exercise play key roles in preventing an FGF21-resistant state, which may partly explain the mechanism by which endurance exercise reduces the risk of cardiometabolic disease.
33
Because this cross-sectional study did not measure intra-hepatic content, further research is needed to examine whether CRF, VFA, and hepatic fat content are individually associated with FGF21 resistance. In addition, we determined that endurance exercise improved FGF21 resistance in elderly subjects; however, it is difficult for elderly and obese individuals to perform at high exercise intensity (75% of VO2max at end of the program). It is therefore necessary to examine the effects of low and moderate intensity exercise on hepatic fat content and FGF21 resistance.
Moreover, the present study revealed that changes in serum FGF21 levels were only associated with changes in hepatic fat content in the short-term exercise intervention, whereas VFA was a key factor associated with serum FGF21 levels in the cross-sectional study, which may indicate long-term effects of FGF21 resistance. Because FGF21 plays an important role in adipose tissue thermogenesis (32) and obesity (6, 9), it is likely that hepatic fat accumulation induces an increase in visceral fat mass accompanying an impaired FGF21 response in adipose tissue. Therefore, it is necessary to evaluate whether FGF21 resistance is associated with development of central adiposity.
5. Acknowledgements
The author thanks Ms. Xiaomin Sun, Dr. Zhen-Bo Cao, Dr. Satomi Oshima, Mr. Ryuken Ise, Mr.
Takafumi Kubo and Ms. Tomoko Ito for their assistance. Dr. Kumpei Tanisawa has been greatly tolerant and supportive during the research and subsequent writing. I am also grateful to all subjects for their participation in this study, and particularly grateful for the comments given by Prof. Shizuo Sakamoto and Prof. Isao Muraoka. In addition, assistance and advice given by Prof.
Mitsuru Higuchi has been a great help in preparing this manuscript.
6. References
(1) Maruthur NM, Gudzune K, Hutfless S, Fawole OA, Wilson RF, Lau BD, Anderson CA,
34
Bleich SN, Segal J. Avoiding weight gain in cardiometabolic disease: a systematic review.
Journal of obesity 2014; 2014:358919
(2) Sawada SS, Lee IM, Muto T, Matuszaki K, Blair SN. Cardiorespiratory fitness and the incidence of type 2 diabetes: prospective study of Japanese men. Diabetes care 2003;
26:2918-2922
(3) Church TS, LaMonte MJ, Barlow CE, Blair SN. Cardiorespiratory fitness and body mass index as predictors of cardiovascular disease mortality among men with diabetes. Archives of internal medicine 2005; 165:2114-2120
(4) Bostrom P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, Rasbach KA, Bostrom EA, Choi JH, Long JZ, Kajimura S, Zingaretti MC, Vind BF, Tu H, Cinti S, Hojlund K, Gygi SP, Spiegelman BM. A PGC1-alpha-dependent myokine that drives brown-fat-like
development of white fat and thermogenesis. Nature 2012; 481:463-468
(5) Kim KH, Kim SH, Min YK, Yang HM, Lee JB, Lee MS. Acute exercise induces FGF21 expression in mice and in healthy humans. PloS one 2013; 8:e63517
(6) Kharitonenkov A, Shiyanova TL, Koester A, Ford AM, Micanovic R, Galbreath EJ, Sandusky GE, Hammond LJ, Moyers JS, Owens RA, Gromada J, Brozinick JT, Hawkins ED, Wroblewski VJ, Li DS, Mehrbod F, Jaskunas SR, Shanafelt AB. FGF-21 as a novel metabolic regulator. The Journal of clinical investigation 2005; 115:1627-1635
(7) Kharitonenkov A, Adams AC. Inventing new medicines: The FGF21 story. Molecular metabolism 2014; 3:221-229
(8) Itoh N. FGF21 as a Hepatokine, Adipokine, and Myokine in Metabolism and Diseases.
(9) Kharitonenkov A, Wroblewski VJ, Koester A, Chen YF, Clutinger CK, Tigno XT, Hansen BC, Shanafelt AB, Etgen GJ. The metabolic state of diabetic monkeys is regulated by fibroblast growth factor-21. Endocrinology 2007; 148:774-781
(10) Fisher FM, Chui PC, Antonellis PJ, Bina HA, Kharitonenkov A, Flier JS, Maratos-Flier E.
Obesity is a fibroblast growth factor 21 (FGF21)-resistant state. Diabetes 2010;
35 59:2781-2789
(11) Hale C, Chen MM, Stanislaus S, Chinookoswong N, Hager T, Wang M, Veniant MM, Xu J.
Lack of overt FGF21 resistance in two mouse models of obesity and insulin resistance.
Endocrinology 2012; 153:69-80
(12) Chen C, Cheung BM, Tso AW, Wang Y, Law LS, Ong KL, Wat NM, Xu A, Lam KS. High plasma level of fibroblast growth factor 21 is an Independent predictor of type 2 diabetes: a 5.4-year population-based prospective study in Chinese subjects. Diabetes care 2011;
34:2113-2115
(13) Bobbert T, Schwarz F, Fischer-Rosinsky A, Pfeiffer AF, Mohlig M, Mai K, Spranger J.
Fibroblast growth factor 21 predicts the metabolic syndrome and type 2 diabetes in Caucasians. Diabetes care 2013; 36:145-149
(14) Ong KL, Januszewski AS, O'Connell R, Jenkins AJ, Xu A, Sullivan DR, Barter PJ, Hung WT, Scott RS, Taskinen MR, Keech AC, Rye KA. The relationship of fibroblast growth factor 21 with cardiovascular outcome events in the Fenofibrate Intervention and Event Lowering in Diabetes study. Diabetologia 2015; 58:464-473
(15) Gaich G, Chien JY, Fu H, Glass LC, Deeg MA, Holland WL, Kharitonenkov A, Bumol T, Schilske HK, Moller DE. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell metabolism 2013; 18:333-340
(16) Zhang X, Yeung DC, Karpisek M, Stejskal D, Zhou ZG, Liu F, Wong RL, Chow WS, Tso AW, Lam KS, Xu A. Serum FGF21 levels are increased in obesity and are independently associated with the metabolic syndrome in humans. Diabetes 2008; 57:1246-1253 (17) Dostalova I, Kavalkova P, Haluzikova D, Lacinova Z, Mraz M, Papezova H, Haluzik M.
Plasma concentrations of fibroblast growth factors 19 and 21 in patients with anorexia nervosa. The Journal of clinical endocrinology and metabolism 2008; 93:3627-3632 (18) Fletcher JA, Meers GM, Laughlin MH, Ibdah JA, Thyfault JP, Rector RS. Modulating
fibroblast growth factor 21 in hyperphagic OLETF rats with daily exercise and caloric
36
restriction. Applied physiology, nutrition, and metabolism = Physiologie appliquee, nutrition et metabolisme 2012; 37:1054-1062
(19) Wei M, Gibbons LW, Mitchell TL, Kampert JB, Lee CD, Blair SN. The association
between cardiorespiratory fitness and impaired fasting glucose and type 2 diabetes mellitus in men. Annals of internal medicine 1999; 130:89-96
(20) LaMonte MJ, Barlow CE, Jurca R, Kampert JB, Church TS, Blair SN. Cardiorespiratory fitness is inversely associated with the incidence of metabolic syndrome: a prospective study of men and women. Circulation 2005; 112:505-512
(21) Fox CS, Massaro JM, Hoffmann U, Pou KM, Maurovich-Horvat P, Liu CY, Vasan RS, Murabito JM, Meigs JB, Cupples LA, D'Agostino RB, Sr., O'Donnell CJ. Abdominal visceral and subcutaneous adipose tissue compartments: association with metabolic risk factors in the Framingham Heart Study. Circulation 2007; 116:39-48
(22) Tanisawa K, Ito T, Sun X, Ise R, Oshima S, Cao ZB, Sakamoto S, Tanaka M, Higuchi M.
High cardiorespiratory fitness can reduce glycated hemoglobin levels regardless of polygenic risk for Type 2 diabetes mellitus in nondiabetic Japanese men. Physiological genomics 2014; 46:497-504
(23) Nishimura T, Nakatake Y, Konishi M, Itoh N. Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochimica et biophysica acta 2000; 1492:203-206 (24) Lundasen T, Hunt MC, Nilsson LM, Sanyal S, Angelin B, Alexson SE, Rudling M.
PPARalpha is a key regulator of hepatic FGF21. Biochemical and biophysical research communications 2007; 360:437-440
(25) Heilbronn LK, Campbell LV, Xu A, Samocha-Bonet D. Metabolically protective cytokines adiponectin and fibroblast growth factor-21 are increased by acute overfeeding in healthy humans. PloS one 2013; 8:e78864
(26) Galman C, Lundasen T, Kharitonenkov A, Bina HA, Eriksson M, Hafstrom I, Dahlin M, Amark P, Angelin B, Rudling M. The circulating metabolic regulator FGF21 is induced by