九州大学学術情報リポジトリ
Kyushu University Institutional Repository
シェーグレン症候群とミクリッツ病の病態形成にお けるThサブセットの関与に関する研究
前原, 隆
Graduate School of Dental Science, Kyushu University
https://doi.org/10.15017/21990
出版情報:Kyushu University, 2011, 博士(歯学), 課程博士 バージョン:
権利関係:
シェーグレン症候群とミクリッツ病の病態形成
における Th サブセットの関与に関する研究
A study on an involvement of Th subsets-related molecules in the pathogenesis of Sjögren’s syndrome and Mikulicz’s disease.
2012年
九州大学大学院歯学府歯学専攻
口腔顎顔面病態学講座 顎顔面腫瘍制御学分野 前原 隆
指導教員
九州大学大学院歯学研究院
口腔顎顔面病態学講座 顎顔面腫瘍制御学分野 中村 誠司 教授
本研究の一部は下記の学術雑誌に投稿中である。
Selective Localization of Th Subsets and B cells in Labial Salivary Glands from Primary Sjögren’s Syndrome Patients
Takashi Maehara, Masafumi Moriyama, Jun-Nosuke Hayashida, Akihiko Tanaka, Shouichi Shinozaki, Kaori Matsumura, Yoshiaki Kubo, Shinsuke Ieda, and Seiji Nakamura
Submitted to Rheumatology.
2010 年 第 64 回 口腔科学会総会にて
「シェーグレン症候群の病変局所における Th サブセットの局在」
として本研究の一部を発表し、学会賞優秀発表賞を受賞した。
略語表
AIP: autoimmune pancreatitis (自己免疫性膵炎) Bcl-6: B cell lymphoma-6
CCR: CC chemokine receptor (CCケモカインレセプター) CXCR: CXC chemokine receptor (CXC ケモカインレセプター) DEPC: dietyl pyrocarbonate
Foxp3: forkhead box p3
GAPDH: glycelaldehyde-3-phosphate dehydrogenase GATA3: GATA binding protein 3
IFN-γ: interferon-γ (インターフェロン-γ) Ig: immunoglobulin (免疫グロブリン) IL: interleukin (インターロイキン) LSG: labial salivary gland (口唇腺) MD: Mikulicz’s disease (ミクリッツ病) mRNA: messenger RNA
PBS: phosphate-buffered saline
PBMC: peripheral blood mononuclear cells (末梢血単核球) PHA: phytohemagglutinin
PCR: polymerase chain reaction
PSC: primary sclerosing cholangitis (原発性硬化性胆管炎) RORC2: retinoic acid-related orphan receptor C2
SS: Sjögren’s syndrome (シェーグレン症候群) T-bet: T box transcription factor
TGF-β: transforming growth factor-β (トランスフォーミング増殖因子-β) Tfh: Follicular helper T (瀘胞性ヘルパー T)
Th: T helper (ヘルパー T)
Th0: T helper 0 (ヘルパー T type 0) Th1: T helper 0 (ヘルパー T type 1) Th2: T helper 0 (ヘルパー T type 2) Th17: T helper 0 (ヘルパー T type 17)
TNF-α: tumor necrosis factor-α (腫瘍壊死因子-α) Treg: T regulatory (制御性 T)
LCM: Laser capture microdissection
目 次
要旨
緒言
材料と方法
結果
<研究 1>
SS 患者の LSG における Th サブセットの発現と局在の検討
1-1. LSG 全 体 に お け る Th サ ブ セ ッ ト の 発 現 に つ い て
1-2. LSG の 病 変 局 所 に お け る Th サ ブ セ ッ ト の 発 現 に つ い て
<研究 2>
MD 患 者 の LSG に お け る GC 形 成 お よ び IgG4 産 生 に 関 わ る分子の検討
2-1. GC 形 成 と Th サ ブ セ ッ ト と の 関 連 つ い て
2-2. IgG4 産 生 に 関 わ る 分 子 に つ い て
考察
謝辞
参考文献
5 9
26 23
56 47 14
35
55 43
要 旨
シェーグレン症候群 (Sjögren’s syndrome: SS) は、唾液腺や涙腺などの外分 泌腺が特異的に障害を受ける臓器特異的自己免疫疾患である。導管周囲への リンパ球浸潤を特徴とし、さらに病態進展とともに胚中心 (germinal center:
GC) を形成することもあり、様々な浸潤様式を呈する。SSの発症や病態進展 の機序についての詳細は不明な点が多いが、ヘルパー T (Th) 細胞の産生する サイトカインの関与が報告されている。そこで、本研究では第一に、自己免 疫疾患に重要な役割を果たすといわれている Th サブセットが産生する特異 的なサイトカインおよび転写因子に着目し、SS 患者の口唇腺 (labial salivary
gland: LSG) におけるこれらの分子の発現について検討した。さらに、病態進
展のメカニズムを解析するために、Laser Capture Microdissection (LCM) を用 いて同一患者の浸潤様式の異なる部位を分離採取し、Th サブセットの発現と 局在について検討を行った。
一方、ミクリッツ病 (Mikulicz’ disease: MD) は、涙腺や唾液腺の腫脹を特 徴とし、病理組織学的類似性から SS の一亜型として認識されてきた。しか し近年、MD に高免疫グロブリン (Immunoglobulin: Ig) G4 血症や腺組織への
IgG4 陽性形質細胞の浸潤が認められることから、MD は SS と全く異なっ
た機序で生じる疾患であることが示唆されている。また、MD のもう一つの 特徴として、腫脹した腺組織におけるGC の形成が SS より高頻度に認めら れることが挙げられる。最近の報告では、一般にGC 形成には Tfh が産生す る IL-21 が必須とされており、さらに IgG4 へのクラススイッチにも関与す るといわれている。そこで、本研究では第二に、MD 患者の LSG における
IL-21 の発現を検索し、GC 形成や IgG4 産生との関連について検討を行った。
以下に本研究で得られた結果をまとめた。
1. SS 患者の LSG おける Thサブセットの発現と局在の検討
SS 患者 54 例と健常者 16 例を対象とした。real-time polymerase chain reaction (PCR) 法を用いて Th1、Th2、Th17、regulatory T: Treg、および follicular helper T: Tfh 特異的なサイトカインおよび転写因子について messenger RNA
(mRNA) 発現を検討したところ、SS 患者の LSG では健常者と比較して、全
ての Th サブセット関連分子の発現が亢進していた。また、SS 患者をリンパ 球の浸潤程度で軽度と重度の 2 群に分け比較したところ、重度では Th2 関 連分子 (IL-4、GATA binding protein 3: GATA3) および Tfh 関連分子 (B cell
lymphoma-6: Bcl-6) の発現が亢進していた。次に病態進展のメカニズムを解析
するために、同一患者での導管周囲の病変局所と GC 形成部における Th サ ブセットの発現を比較検討した。SS 患者 54 例のうち、LSG の新鮮結切片 において GC の形成が認められた 8 例を対象とした。新鮮凍結切片におい て導管周囲に浸潤しているリンパ球を GC (-) とし、GC および GC 周囲の リンパ球を GC (+) として、それぞれ LCM を用いて分離採取した。同一患 者での GC (-) と GC (+) の 2 群において real-time PCR 法を用いて、Th サ ブセット関連分子の mRNA 発現を検討した。その結果、Th1 関連分子 (interferon-γ: IFN-γ、IL-12、T box transcription factor: T-bet) および Th17 関連 分子 (IL-17、retinoic acid-related orphan receptor C2: RORC2) の発現は、GC (-) で亢進していた。一方、Th2 関連分子 (IL-4, GATA3) および Tfh 関連分子
(IL-21、Bcl-6) の発現は、GC (+) で亢進していた。Treg 関連分子については、
両群間で有意な差は認めなかった。免疫組織化学染色でも同様に、IFN-γ お よび IL-17 は GC (-) に、IL-4 および Bcl-6 はGC (+) に強く発現していた。
また、forkhead box p3 (Foxp3) の発現は、両群間で明らかな差は認められなか
った。
2. MD 患 者 の LSG に お け る GC 形 成 と IgG4 産 生 に 関 わ る 分 子 の 検討
MD 患者12 症例、SS 患者 15 症例、健常者 15 症例を対象とした。検討 した MD は、全症例 LSG のリンパ球浸潤程度が重度であるため、比較対象 とした SS は、全て重度の症例を選択した。まず real-time PCR 法を用いて、
Tfh 関連分子 (IL-21、Bcl-6、CXC chemokine receptor: CXCR5) について検討 を行った。その結果、すべての Tfh 関連分子の発現は、SS および MD とも 健常者と比べて有意に亢進していた。また、免疫組織化学染色にてそれらの 局在をみたところ、SS 患者ではすべてのTfh 関連分子は、GC のみに強く発 現していた。一方 MD 患者では、Bcl-6 および CXCR5 は、SS 患者と同様 に GC に強く発現していたが、IL-21 は LSG 全体に強い発現を認めた。つ まり、MD における IL-21 産生細胞は Tfh の他にもある可能性が示唆され
た。IL-21は主にTfhが産生するといわれているが、その他にもIL-21 産生能
を有するとの報告がある Th2 および Th17 についても、同様の検討を行った。
その結果、MD では、免疫組織化学染色にて Th2 関連分子 (IL-4、CC chemokine receptor 4: CCR4、c-Maf) は、IL-21 と同様にGC だけでなく LSG 全体に強い発現を認めた。一方、Th17関連分子 (IL-17) は、ほとんど発現を
認めなかった。また、MD の IgG4 産生とIL-21 との相関をみたところ、IL-21
の mRNA 発現量は、IgG4 の mRNA 発現量および IgG4 陽性細胞率と正の
相関を認めた。
研究 1. より、SS 患者の唾液腺では Th サブセットの局在が認められ、Th1 および Th17 が導管周囲に集積することで SS の発症に関与し、Th2 および Tfh が集積することで病態の進展、特に GC の形成に関与していることが示 された。また研究 2. より、MD 患者の唾液腺では Th2 を主体とする IL-21 の産生により高頻度に GC が形成され、IgG4 へのクラススイッチが誘導さ れることが示唆された。
緒 言
シェーグレン症候群 (Sjögren’s syndrome: SS) は、唾液腺や涙腺などの外分 泌腺にリンパ球が浸潤し、それに伴い腺組織が特異的に障害を受ける臓器特 異的自己免疫疾患である。主症状として、ドライマウス (口腔乾燥) やドライ アイ (乾燥性角結膜炎) がある。病理組織学的には、導管上皮へのリンパ球浸 潤を特徴とし、病態進展とともに異所性に胚中心 (germinal center: GC) を形 成することもあるなど、様々な浸潤様式を呈することがわかっている (1, 2)。 さらに、高 γ グロブリン血症や悪性リンパ腫などの腺外症状が出現すること があるため、リンパ増殖性病変とも称されている (3)。免疫組織学的には、唾 液腺に CD4 陽性ヘルパー T (helper T: Th) 細胞を主体としたリンパ球浸潤 が認められ、これらの細胞が SS の発症や病態進展に重要な役割を担ってい ると考えられている (4, 5)。しかしながら、その病因や病態進展についてはい まだ不明な点が多く、SS の発症や病態進展に CD4 陽性 Th 細胞がどのよう に関与しているかを明確にすることは、SS の病態解明に重要であると考えら れる。
一般に、免疫系のバランスとその恒常性は CD4 陽性 Th 細胞により保 たれているが、その Th 細胞群のバランスの破綻が、様々な自己免疫性疾患 の原因となっていると考えられている。Th 細胞は、分泌するサイトカインや 発現している転写因子の違いから機能的に異なるいくつかのサブセットに分 類されており (6-11)、少なくとも 6 つのサブセットとして Th type 0 (Th 0)、Th type 1 (Th1)、Th type 2 (Th2)、Th type 17 (Th17)、濾胞性 T (follicular helper T:
Tfh)、および制御性 T (regulatory T: Treg) 細胞が報告されている (12)。Th1 は
インターロイキン (interleukin: IL)-12 によって誘導され、特異的な転写因子 T box transcription factor (T-bet) を発現して (13)、IL-2、インターフェロン-γ (interferon-γ: IFN-γ)、および腫瘍壊死因子-α (tumore necrosis factor-α: TNF-α) を産生し (14)、主に細胞性免疫を担っている。Th2 は IL-4 によって誘導され、
特異的な転写因子 GATA binding protein 3 (GATA3) を発現して (15)、IL-4、IL-5、
IL-13 を産生し、主に体液性免疫を担っている。また、Th0 は他の Th サブ
セットへと分化しうる前駆細胞と考えられている。近年報告されている Th17 は IL-1β、IL-6、および IL-23 によって誘導され、特異的な転写因子 retinoic acid-related orphan receptor C2 (RORC2) を発現して IL-17 を産生し、種々の 自己免疫疾患に関連があるとされている (16-18)。Tfh は特異的な転写因子 B cell lymphoma-6 (Bcl-6) を発現して IL-21 を産生し、GC の形成や自己免疫疾 患の病態に関与しているとされている (19)。また、Treg は transforming growth factor-β (TGF-β) によって誘導され、特異的な転写因子forkhead box p3 (Foxp3) を発現して IL-10、TGF-β を産生し (20, 21)、他のエフェクター Th 細胞の免疫 応答を調節するとされている (22-24)。これらの Th サブセットが産生するサイ トカインによって、他のサブセットの細胞群の増殖や機能をお互いに抑制す ることで調整し合い、免疫系のバランスを保っている (11, 25-28)。このように、
自己免疫疾患における Th サブセットの発現パターンを解析することは、そ の病因や発症のメカニズム解明に寄与すると考えられる。われわれの過去の 研究では、SS 患者の口唇腺 (labial salivary gland: LSG) における Th1/Th2 バ ランスについて検討しており、その発症には Th1 が、病態進展には Th2 が 重要な役割を果たしていることを報告してきた (29)。
また、SS 患者の唾液腺導管周囲には、リンパ球が浸潤するだけでなく、
病態進展とともに GC を形成することがあり (30-33)、LSG において GC を形 成している SS 患者では、血清中の抗 Ro/SS-A 抗体、抗 La/SS-B 抗体、お よび免疫グロブリン (Immunoglobulin: Ig) G が高値であるとの報告がある (34)。 一般に GC は、Th 細胞の刺激により抗原に高い親和性をもつ B 細胞が活性 化され、増殖することにより形成される。GC ではIgV 領域の高頻度体細胞 突然変異と高親和性 Ig の産生 (クラススイッチ) が生じており、その周囲に は、B および T 細胞が集積している。最近 Theander らは、LSG に GC の 形成がみられる SS 患者では、悪性リンパ腫の発症頻度が高くなることを報 告している (2)。したがって、 SS のGC 形成に至るメカニズムを明らかにす ることは、SS の発症から悪性リンパ腫までの病態進展の機序を解明するのに 重要であると考えられる。
そこで研究 1. では、SS の病態と Th サブセットの関わりを明確にするた めに、まずは従来報告のある Th1、Th2 に加えて、Th17、Tfh、および Treg に も着目し、LSG 全体における Th サブセット関連分子 (特異的なサイトカイ ンおよび転写因子) の発現を検討した。しかし LSG 全体での解析は、LSG 内 の様々なリンパ球浸潤様式をまとめて解析するということになる。よって、
正確にSSの病態進展のメカニズムを解明するためには、同一患者の経時的・
複数回の LSG の採取(生検)が必要であるが、倫理的観点から困難をとも なう。そこで、本研究では同一患者の浸潤様式が異なる部位を採取し、それ ぞれの病変局所のリンパ球を解析することで、複数回の採取をせずに SS の 病 態 進 展 モ デ ル を 確 立 す る こ と 試 み た 。 具 体 的 に は laser capture
microdissection (LCM) (35, 36) を用い、同一患者の生検組織で導管周囲に浸潤し たリンパ球および GC 形成部のリンパ球をそれぞれ分離採取し、Th サブセ ットの発現と局在について検討した。
次に、われわれは SS の一亜型として認識されてきた (37) ミクリッツ病
(Mikulicz’s disease; MD) にも注目した。MD は、病理組織学的に涙腺・唾液
腺の導管・腺房周囲に著明な炎症性細胞浸潤を認めることから、 SS の典型 像と類似点が多い。しかし MD では、抗 Ro/SS-A 抗体や抗 La/SS-B 抗体な どの疾患特異的自己抗体がほとんどの症例で陰性であり、ステロイドが著効 することなど臨床的に SS と異なる所見が多い (38)。さらには近年、高 IgG4 血症や腺組織への IgG4 陽性形質細胞の浸潤が認められることが報告されて おり (39)、SS と異なった疾患と認識されるようになってきた。また、MD と 高頻度に合併する自己免疫性膵炎 (autoimmunne pancreatitis: AIP)、硬化性胆管 炎 (primary sclerosing cholangitis: PSC)、および間質性腎炎などについても、そ の病理組織学的特徴が MD に非常に類似しており、ステロイドへの反応性も 同じであることから、共通する病態として『IgG4 関連疾患』と総称されるよ うになった (39)。この IgG4 関連疾患は日本から提唱されたものであり、内科、
眼科、病理、そして口腔外科を主体とした病態解明のための研究班を厚生労 働科学研究『難治性疾患克服研究事業』として推進され、われわれも研究協 力者として『唾液腺病変の病態解明』のプロジェクトの一旦を担っている。
われわれの過去の研究結果では、MD の病態形成には SS と異なり、Th2 が 産生する IL-4 と Treg が産生する IL-10 がその病態形成や IgG4 産生に重 要であることを明らかにしてきた (40)。また MDの病理組織学的所見として、
唾液腺では SS より高頻度に GC を形成するという特徴がある。前述のとお り、GC 形成には Tfh が産生する IL-21 が 重要であることが報告されてい ることから (19, 41)、IL-21 が MD の病態形成に何らかの関与があるのではない かと推察された。最近の研究では、ヒトやマウスの B 細胞を IL-4 と IL-21 で共刺激すると、IgG1 産生からMD の特徴であるIgG4 産生へとクラススイ ッチが引き起こされることが報告されている (42, 43)。さらに末梢血単核球 (peripheral blood mononuclear cells: PBMC) を IL-4 と IL-21 で刺激すると、
IL-10 の産生が増加するといわれている (44)。これまでの報告をあわせて考え
ると、MD の IgG4 産生には、IL-21 も関与している可能性が考えられる。
しかしながら、MD 患者の唾液腺において、病態の根幹である GC 形成およ び IgG4 産生における IL-21 の関与についての報告はいまだない。
そこで研究 2. では、MD の LSG における IL-21 の発現を検索し、高頻 度の GC 形成および IgG4 の産生との関連について検討を行った。
材料と方法
1. 対象患者および組織採取
対象は、平成 19 年から平成 23 年に九州大学病院顎口腔外科を受診し、
一次性 SS と診断された 54 症例 (男性 3 症例、女性 51 症例、平均年齢:
61.2歳)、MD と診断された 12 症例 (男性 3 症例、女性 9 症例、平均年齢:
61.8 歳)、唾液分泌機能障害が認められない健常者 16 症例(男性 4 症例、
女性 12 症例、平均年齢: 58.4 歳)とした。SS および MD 患者の LSG は、
生検時に採取し (45)、健常者の LSG は、粘液貯留嚢胞摘出時に採取した病変 周囲の健常 LSG を用いた。LSG は、免疫組織学的解析用にはパラフィンワ ックスに包埋し、mRNAの解析用には直ちに液体窒素を用いて凍結し、-80℃
で保存した後に実験に用いた。
2. SS および MD の診断
SS の診断はすべて厚生省シェーグレン診断基準 (1999年改定基準) に準 じて行ったが (46)、すべての患者はヨーロッパとアメリカでも用いられる診断 基準によっても SS と診断された (47)。一方、MD の診断は日本シェーグレン 学会にて承認された診断基準 (48, 49) に準じて行った。SS と MD の LSG に おけるリンパ球浸潤程度には、石川・小守の分類により ±、+、++、+++ と 評価したが (50)、MD は全症例で浸潤度 +++ であったため、 MD との比較 に用いた SS は浸潤度 +++ の 15 症例 (男性 3 症例、女性 12 症例、平均 年齢: 61.4 歳) を対象とした。
3. LSG の RNAの抽出および complementary DNA (cDNA) の合成
RNA の抽出には acidified guanidinium-;henol-chloroform (40, 51, 52) 法を用いた。
まず、 LSG に Torizol® Reagent (Invitrogen, California, USA) を 1 ml 加え、ホ モジナイザーを用いて粉砕した。0.2 ml のクロロホルム(和光純薬、大阪、
日本)を加えて撹拌後、15 分間静置した。4 ℃、15,000 rpm で 15 分間遠心 した後に RNA を含む水層を採取し、これに 1 ml のイソプロパノール(和 光純薬)を加えて撹拌後、4 ℃、15,000 rpm で 15 分間遠心し、上清の除去 後に得られた RNA ペレットを 80% EtOH(和光純薬)で洗浄後乾燥させ、
50 μlの 0.1% dietyl pyrocarbonate (DEPC) 処理水に溶解した。その後、吸光度 計にて RNA の濃度を測定した。
cDNA の合成には、DEPC 処理水に約 3.0 μg の total RNA、40 U/μl の Recombinant RNasin® Ribonuclease Ingibitor (Promega, Madison, WI, USA) を 0.5 μl 、0.5 μg/μl の pd (T)1218 を 1 ml、10 mM PCR Nucleotide Mix (以上、
Amersham Pharmacia Biotech, Piscataway, NJ, USA) を 1 ml、250 mM トリス塩酸 塩 (pH 8.3)、375 mM KCl および 15 mM MgCl を含む反応緩衝液を 4 μl、100 mM dithiothretiol を 2 ml、200 U/ml の SUPERSCRIPT™ II RNasse H Reverse Transcriptase (以上、Life Technologies, Rockville, MD, USA) を 0.5 μl 加えて合 計 20 ml とし、42 °C で 1 時間インキュベートした。その後、95˚C で 5 分間 加温して酵素を失活させ、直ちに氷冷した。これを DEPC 処理水で 2 倍に 希釈し、サイトカイン、ケモカインレセプター、転写因子の mRNA の解析 に用いた。
4. Laser Capture Microdissection (LCM) による病変組織の分離採取 54 症例の SS 患者のうち、口唇腺新鮮凍結標本にて GC の形成が認めら れた 8 症例を対象とした。図 2a に示すように、GC の形成を伴わずに導管
周囲に浸潤しているリンパ球を GC (-) とし、GC および GC 周囲のリンパ 球を GC (+) と定義し、それぞれ LCM を用いて分離採取した。口唇腺新鮮 凍結切片 (13 μm) をポリエルリジン (SIGMA、P8920) にてコーティングした フォイル付きスライドガラス (90 FOIL-SL25) に貼り付け作製した。切片を EtOH/CH3COOH (19 : 1) 溶液にて固定後、75 % EtOH に 30 秒、滅菌水にて 20 秒洗浄後、HistoGene Staining Solution (LCM Frozen Section Staining Kit
(ARCTURUS) で 20 秒間染色した後、滅菌水にて 30 秒間洗浄を行った。さ
らに、75 % EtOH、95 % EtOH、100 % EtOH に各 30 秒間浸漬させ脱水処理
を行い、Xylene に 5分間浸漬させた後にドライヤー冷風にて 30 秒乾燥させ
た。その後直ちに Leica Microsystems Japan (AS-LMD; Leica Microsystems Japan, Tokyo, Japan) を用いて、LCM を行い (35, 36)、病変組織の分離採取を行った (図
2b)。組織採取では、導管上皮を含まないようにした。さらに GC (+) におけ
る採取範囲は、GC とその周囲に浸潤しているリンパ球も含めて採取行うた めに、GC の半径を r とし、1.5 × r までの範囲で採取を行うこととした。UV レーザー照射により分離された組織は、QIAzol Lysis Reagent (Invitrogen) を 50 μl 入れたチューブ (CapSure HS LCM Cap (TaKaRa Code AL204) のキャ ップに回収し、直ちに RNA の抽出を行った。
5. LCM による病変組織の RNAの抽出および cDNA の合成
LCM により回収した組織の RNA の抽出は、RNeasy Lipid Tissue Mini Kit
(QIAGEN) を用いて行った。まず LCM にて回収した組織の入ったチューブ
に QIAzol Lysis Reagent (Invitrogen) を 950 μl 加え、室温にて 5 分間放置し た。200 μl のクロロホルム(和光純薬、大阪、日本)を加えて 15 秒間撹拌
後、室温にて 3 分間静置した。4 ℃、12,000 rpm で 15 分間遠心した後に RNA を含む水層を採取し、600 μl の 70 % エタノール(和光純薬)を加えて 撹拌後、2 ml のカラムチューブへ入れ、20 ℃、10,000 rpm で 20 秒間遠心 した。次に 700 μl のBuffer RW1 (QIAGEN) をカラムチューブへ入れ、 20 ℃、
10,000 rpm で 15 秒間遠心した。さらに 500 μl の Buffer RPE (QIAGEN) を カラムチューブへ入れ、20 ℃、10,000 rpm で 15 秒間遠心した後、500μl の Buffer RPE をカラムチューブへ入れ、20 ℃、10,000 rpm で 2 分間遠心した。
新しい RNA 保管用チューブをセットし、30 μl の0.1% DEPC処理水をカラ ムチューブへ入れ、 20 ℃、10,000 rpm で 1 分間遠心した。回収した RNA が 溶解した DEPC 処理水をカラムチューブへ再度入れ、20 ℃、10,000 rpm で 1 分間遠心した。その後、吸光度計にて RNA の濃度を測定した。cDNA の合 成は前述と同様の方法で行った。
6. Reverse transcriptase (RT) – polymerase chain reaction (PCR) 法による LCM を行った組織のmRNA発現の解析
LCM を行った組織は、ごく微量であるためサイトカインおよび転写因子の 発現を確認するためにRT-PCR を行った。RT-PCR は滅菌水に template DNA を 30 ~ 50 ng、10×Taq DNA Polymerase Buffer を 1.25 μl、25 mM 塩化マグネ シウム溶液を1.0 μl、5 units/μl Taq DNA polymerase を 0.1 μl (以上、Bio Basic)
2.0 mM dNTPmix (Toyobo) を 0.5 μl、センスおよびアンチセンスプライマーを
それぞれ 0.5 μl 加えて全反応量を 13.5 μl とした。反応条件は、熱変性は 94℃ で 1 サイクル目が 3 分、2 サイクル目以降は 30 病間で行い、伸長反 応は 72 ℃、30 秒間とした。得られた PCR 産物を 1.8 % アガロースゲル
(Invitrogen life Technologies) 上で電気泳動を行い、エチジウムブロマイド染色 後に、紫外線により可視化したものを図 2c に示す。
7. real-time polymerase chain reaction (PCR) 法によるサイトカイン、ケモ カインレセプター、転写因子の mRNA発現の解析
real-time PCR はBrilliant® Ⅱ SYBR® Green QPCR Master Mix (Stratagene, USA) を用いて行った。滅菌水に Master Mix を 10 μl、template DNA を 10 ng、
20 pMセンスおよびアンチセンスプライマーをそれぞれ 0.5 μl 加え、全反応
量を 20.0 μl とした。反応条件は、熱変性は 95 ℃ で 1 サイクル目が 5 分、
2サイクル目以降は 10 ~ 30 秒間で行い、伸張反応は 72 ℃、10 ~ 30 秒間と し、全 45 サイクルの増幅を行った。mRNAの発現を解析する分子は、Th1 に 特異的なサイトカインである IFN-γ と IL-12 および転写因子である T-bet、
Th2 に特異的なサイトカインであるIL-4 とIL-5、ケモカインレセプターであ る CC chemokine receptor (CCR4)、および転写因子である GATA3、 Th17 に 特異的なサイトカインである IL-17 および転写因子である RORC2、Treg に 特異的なサイトカインである IL-10 と TGF-β および転写因子である Foxp3、
そして Tfh に特異的なサイトカインである IL-21、ケモカインレセプターで あるCXC chemokine receptor 5 (CXCR5) および転写因子である Bcl-6、そして IgG と IgG4 とした。反応条件は、熱変性は95℃で1サイクル目が10分間、
2サイクル以降は10秒間で行った。伸張反応は72℃で30秒間とし、全て45 サイクルの増幅を行った。各プライマー配列を表 1 に示す。陽性対照として は、健常者の新鮮PBMCを、10mg/ml Phytohemagglutinin (PHA) を加えたRPMI 1640 (Life Technologies) 培養液中で72 時間刺激し、そこから抽出したmRNA
から合成したcDNA を用いた。また、各症例間でサイトカインおよび特異的 転写因子のmRNA発現量を定量化するために、それぞれの mRNAの発現量 はハウスキーピング遺伝子である β-actin mRNAの発現量を用いて補正し、相 対的発現量を算出した。
8. 免疫組織学的解析
パラフィン切片 (5 μm) を作製し、通常のストレプトアビジン-ビオチン法 による免疫組織化学染色をCSA II (Biotin-Free Catalyzed Amplification) System (Dako, Cambridge, UK) を用いて行った (40, 53)。サイトカイン、ケモカインレセ プターおよび特異的転写因子の解析に使用した抗体は、抗 IL-4 ポリクロー ナル抗体 (clone: ab9622, Abcam, Cambridge, UK)、抗 IL-10 ポリクローナル抗 体 (clone: ab34843, Abcam)、抗 IFN-γ ポリクローナル抗体 (clone: ab9657, Abcam) 、 抗 IL-17 ポ リ ク ロ ー ナ ル 抗 体 (sc-7927, SANTA CRUZ BIOTECHNOLOGY, INC, CA, USA)、抗 IL-21 ポリクローナル抗体 (clone:
LS-C401; LifeSpan BioScience)、抗 c-Maf 抗体 (clone: ab77071, Abcam)、抗 Foxp3 モノクローナル抗体 (clone: mAbcam 22510; Abcam)、抗 Bcl-6 モノク ローナル抗体 (clone: ab9479; Abcam)、抗 CXCR5 モノクローナル抗体 (clone:
ab89259; Abcam)、抗 CCR4 モノクローナル抗体 (MAB1567, R&D systems) を用いた。IgG4/IgG 陽性細胞の解析には抗 IgG4 モノクローナル抗体 (The Binding Site, Birmingham, UK)、抗 IgG ポリクローナル抗体 (A0423, Dako) を 用いた。なお、顕微鏡用デジタルカメラ DP25 (オリンパス株式会社、東京、
日本) を用いて、各抗体の陽性細胞を撮影し、記録した。
9. IgG4 陽性細胞率の算出
IgG4 陽性細胞率は、強拡大 5 視野で IgG4 陽性細胞と IgG 陽性細胞をそ
れぞれ計測し、各視野の IgG4 陽性細胞数を IgG 陽性細胞数で除したものを 平均した (47, 48)。
10. 統計
統計処理にはMann-Whitney の U 検定および Spearman 順位相関係数を用 いた。なお、統計解析ソフトとして JMP software version 8 (SAS Institute, USA) を使用し、P < 0.05の場合を統計学的有意差ありとした。
結 果
研究 1.
SS 患者の LSG おける Th サブセットの発現と局在の検討
1-1. LSG 全体における Th サブセットの発現について
各症例間で LSG における Th サブセット関連分子 (IFN-γ、T-bet、IL-4、
IL-5、GATA3、IL-17、RORC2、IL-10、Foxp3、IL-21、Bcl-6) の mRNA 量を 比較した。
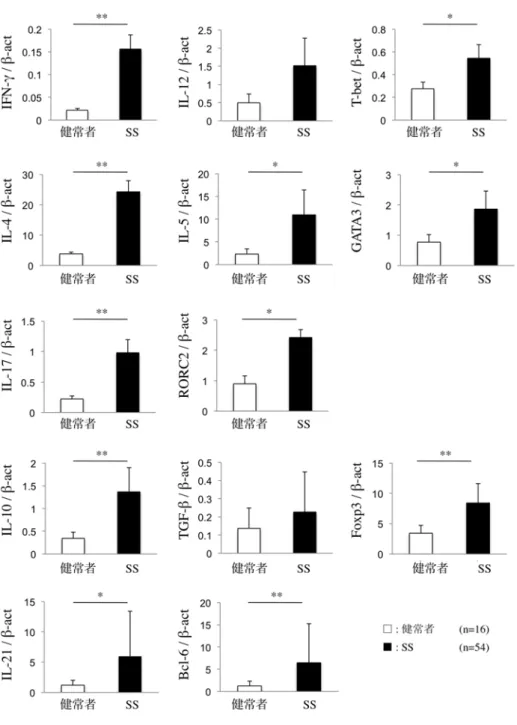
SS 患者の LSG では健常者の LSG と比較して、すべての Th サブセット 関連分子 (IFN-γ、T-bet、IL-4、IL-5、GATA3、IL-17、RORC2、IL-10、Foxp3、
IL-21、Bcl-6) のmRNA発現量が有意に亢進していた (図1a、P < 0.05, P < 0.01;
Mann-WhitneyのU 検定)。
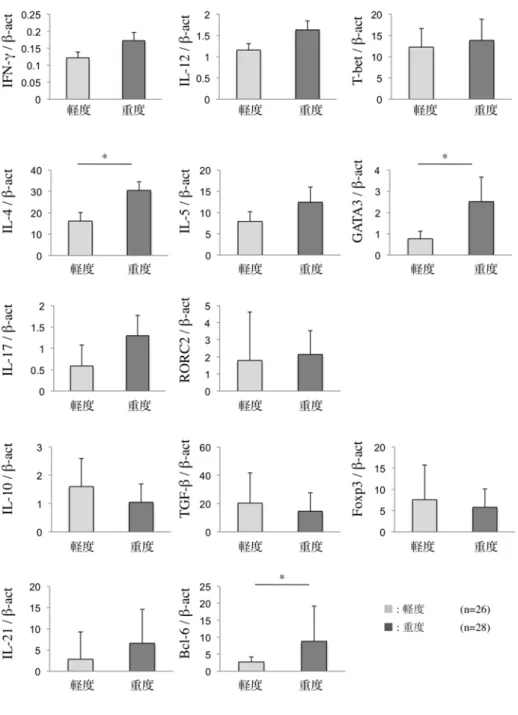
次に、病態の進展と関連分子の関連をみるために SS のリンパ球浸潤程度 との関連を検討したところ、Th1、Th17、および Treg 関連分子の発現は、リ ンパ球浸潤程度との間に有意な相関はみられなかった。しかしながら、Th2 関 連分子 (IL-4、GATA3) と Tfh 関連分子 (Bcl-6) の発現は、リンパ球浸潤が 重度の症例で有意に亢進していた (図1b、P < 0.05, P < 0.01; Mann-Whitneyの U 検定)。
図 1a. SS 患者と健常者の LSG における Th サブセット関連分 子(サイトカイン、転写因子)の mRNA 発現量
定量的 real-time PCR を行い、β-actin に相対的な mRNA 発現量を算出した。
SS 患 者 と 健 常 者 間 で Mann-Whitney の U 検 定 を 行 っ た (*P<0.05、
**P<0.01)。実験方法は「材料と方法」に示す。
図 1b SS 患者の LSG における Th サブセット関連分子(サイ ト カ イ ン 、 転 写 因 子 ) の mRNA 発 現 量 と リ ン パ 球 浸 潤 程 度 と の 関連
定量的 real-time PCR を行い、β-actin に相対的な mRNA 発現量を算出した。
リンパ球の浸潤程度は石川・小守の分類に基づき、軽度 ( 、+) と重度 (++、
+++) に分類して、2 群間で Mann-Whitney の U 検定を行った (*P<0.05、
**P<0.01)。実験方法は「材料と方法」に示す。
1-2. LSG の病変局所における Th サブセットの発現について
SS患者 54 症例のうちパラフィン切片にて GC の形成が認められたのは、
12 症例 (22 %) であり、この頻度は過去の報告と類似していた (31)。この 12 症例の LSG における組織学的所見の代表的症例を図2a に示すが、全ての症 例で導管周囲のリンパ球浸潤と、腺房の破壊およびアポトーシスがみられ、
さらに GC の形成を確認した。また、12 症例のうち新鮮凍結切片で GC の 形成が認められた8 症例 (表2) を対象に LCM を行った。
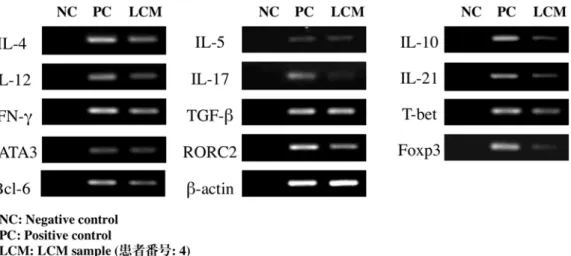
一般にGC は、B 細胞、濾胞樹状細胞、および T 細胞からなり、Ig 産生 形質細胞の産生の場となっており、その周囲には B 細胞や T 細胞が集積し ている。SS の LSG には、リンパ球の浸潤が重度になると GC が形成され ることから、導管周囲に浸潤した部分のリンパ球を GC (-) と定義し、GC を 形成している部分のリンパ球を GC (+) と定義して (図2a)、それぞれの病変 組織を LCM にて分離採取 (図 2b) した。LCM により採取された組織量は ごく微量であるため、Th サブセット関連分子(サイトカイン、転写因子)の 発現については real-time PCR にて確認した (図2c)。新鮮凍結切片において、
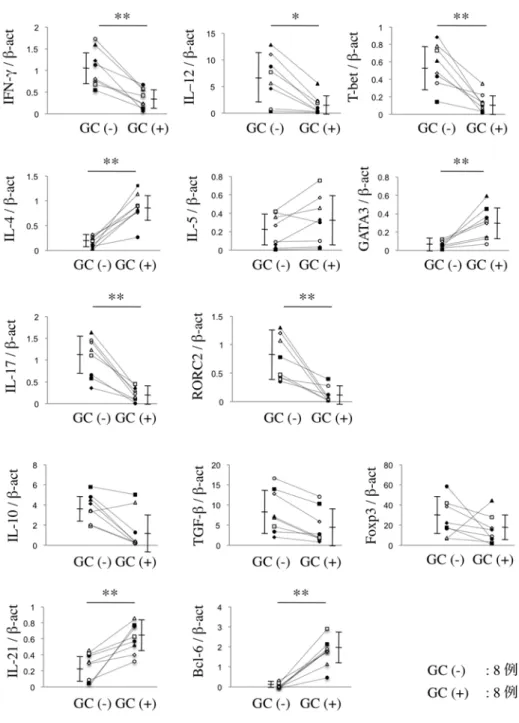
同一患者間の GC (-) と GC (+) の 2 群で、Th サブセット関連分子の mRNA 発現を、real-time PCR にて比較検討した (図3)。相対的 mRNA 発現 量でみると、Th1 関連分子 (IFN-γ、IL-12、T-bet) と Th17 関連分子 (IL-17、
RORC2) の発現は、GC (-) で有意に亢進していた。また、Th2 関連分子 (IL-4、
GATA3) と Tfh 関連分子 (IL-21、Bcl-6) の発現は、GC (+) で有意に亢進し ていた。Treg 関連分子 (IL-10、TGF-β、Foxp3) の発現は、両群間で有意な差 は認められなかった (P < 0.05, P < 0.01; Mann-WhitneyのU 検定)。
次に、mRNA 発現が有意に亢進していた Th サブセット関連分子の局在を 明らかにするために、免疫組織化学染色を行い GC (-) と GC (+) で比較検討 した (図4a)。その結果、IFN-γ および IL-17 は、ともに GC (-) に強い発現 を認めた (A-D)。一方、IL-4 および Bcl-6 は、ともに GC (+) に強い発現を 認めた (E、F、I、J)。また Foxp3 の発現は散在しており、局在は認められな かった (G、H)。Bcl-6 は GC (+) に強い発現を認めたが、この Tfh の転写因 子である Bcl-6 (54) は、GC 内の B 細胞に発現しその維持に必須であるとの 報告もあり(55)、Bcl-6 陽性細胞が Tfh 細胞であるとは断定できない。そこで
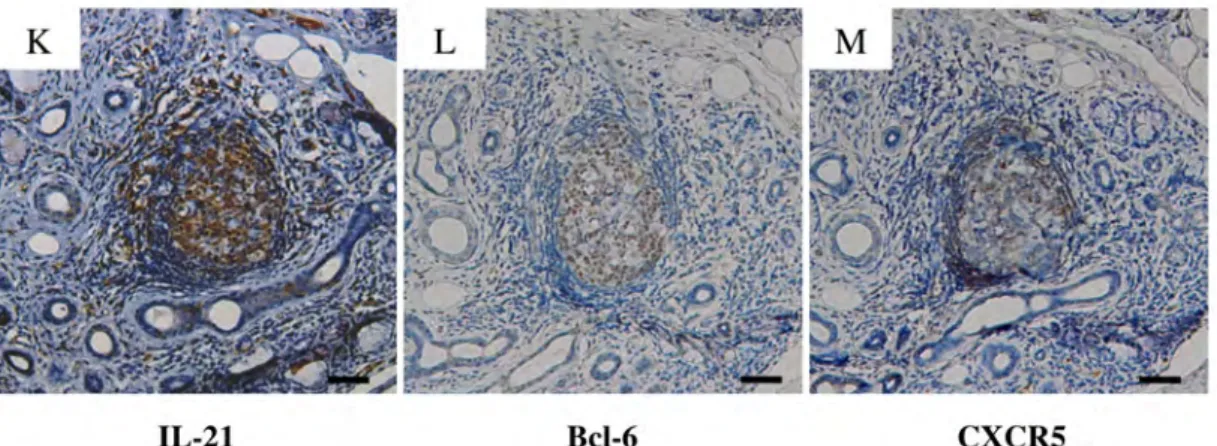
GC (+) における Tfh に関連するその他の特異的な分子 (IL-21、CXCR5) の
発現もみることとした (図4b)。その結果 IL-21 および CXCR5 も、GC (+) に 強い発現がみられた (K、M)。
これらの結果をまとめると、SS 患者の LSG では、全ての Th サブセット 関連分子の発現が健常者と比較して有意に亢進していた。また、SS の重度症 例では軽度症例と比較して、Th2 および Tfh 関連分子の発現が亢進していた。
さらに、LSG の導管周囲では Th1 および Th17 の発現が亢進しており、GC 形成部では Th2 および Tfh の発現が亢進していた。
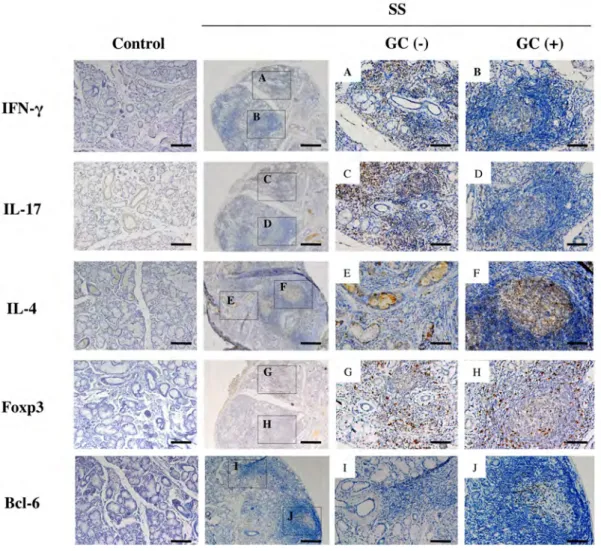
図 2a SS 患者 (石川・小守の分類: +++) の LSG における病理組 織学的所見
(パラフィン包埋標本を用いた HE 染色: scale bar 100μm)
すべての症例で導管周囲へのリンパ球浸潤と、腺房細胞の破壊が認められ、
GC の形成 (緑点枠) がみられた。赤点枠で囲む部分を、導管 (黄矢印) 周囲 に浸潤した部分のリンパ球として GC (-) とし、GC (緑点枠) とその周囲のリ ンパ球を含んだ部分のリンパ球 (青点枠) を GC (+) と定義した。
図 2b SS 患者の LSG における LCM を用いた組織採取
(代表症例の新鮮凍結標本を用いた HistoGene Staining Solution 染色: scale bars 200μm)
同一症例の LSG における GC (-) と GC (+) をそれぞれ、 LCM を用いて分 離採取した。その際、導管 (黄矢印) を含まないように注意した。また、GC (+) における採取範囲 (青枠) は、GC(緑枠)の半径を r としたら、その 1.5 × r までとした。実験方法は「材料と方法」に示す。
図 2c SS 患者において LCM を行った組織におけるサイトカイ ン、転写因子の mRNA の増幅産物
RT-PCR を用いて PCR 産物を 1.8% アガロースゲル上で電気泳動を行い、
エチジウムブロマイド染色をした後、紫外線により可視化した。実験方法は
「材料と方法」に示す。
図 3 SS 患 者 の LSG か ら LCM を 用 い て 分 離 採 取 し た GC (-) と GC (+) の 2 群間における Th サブセット関連分子 (サイトカ イン、転写因子) の mRNA 発現量
定量的 real-time PCR を行い、β-actin に相対的な mRNA 発現量を算出した。
同一患者の LSG の導管周囲病変局所より GC (-) と GC (+) を、それぞれ分 離 採 取 し て 、2 群 間 で Mann-Whitney の U 検 定 を 行 っ た (*P<0.05、
**P<0.01)。実験方法は「材料と方法」に示す。
図 4a SS 患者および健常者の LSG における Th サブセット関 連分子 (サイトカイン、転写因子) の発現
(代表症例のパラフィン包埋標本を用いた免疫組織化学染色: scale bar 100μm (A-J)) IFN-γ および IL-17 は、 SS の GC (+) よりも GC (-) に強い発現がみられ た (A-D)。一方 IL-4 および Bcl-6 は、GC (-) よりも GC (+) に強い発現が みられた (E、F、I、J)。Foxp3 は、両群間で明らか発現の差はみられなかっ た (G、H)。実験方法は「材料と方法」に示す。
図 4b SS 患者の LSG の GC 形成部における IL-21、Bcl-6、CXCR5 の発現
(代表症例のパラフィン包埋標本を用いた免疫組織化学染色: scale bar 100μm)
IL-21 および CXCR5 も、GC (+) に強い発現がみられた (K、M)。実験方法
は「材料と方法」に示す。
研究 2.
MD 患 者 の LSG に お け る GC 形 成 お よ び IgG4 産 生 に 関 わる分子の検討
2-1. GC 形成と Th サブセットとの関連について
MD 症例の臨床所見および血液学的所見を表 3 に示す。男女比は男性 3 例、女性 9 例 (男女比=1:3) と女性に多い傾向であった。主訴は涙腺およ び唾液腺の腫脹が多く、その腺腫脹は全症例弾性硬で圧痛がなく、3 ヶ月以 上持続していた。MD には AIP (56) や PSC (57) など他の IgG4 関連疾患を合 併している症例が多くみられた。血清学的検査では、MD のほとんどの症例 で高 γ グロブリン血症を認め、血清 IgG4 値は測定した症例では全症例にお いて高値を示した。MD における抗 SS-A/SS-B 抗体は全症例陰性であった。
今回結果は示していないが、SS では高 γ グロブリン血症を呈したが、抗核 抗体および抗 SS-A/SS-B 抗体は全例陽性であった。なお、検討した MD 患 者は全症例リンパ球浸潤程度が重度であるため、比較対象とした SS も全症 例重度の症例 (石川・小守の分類 +++) を選択した。
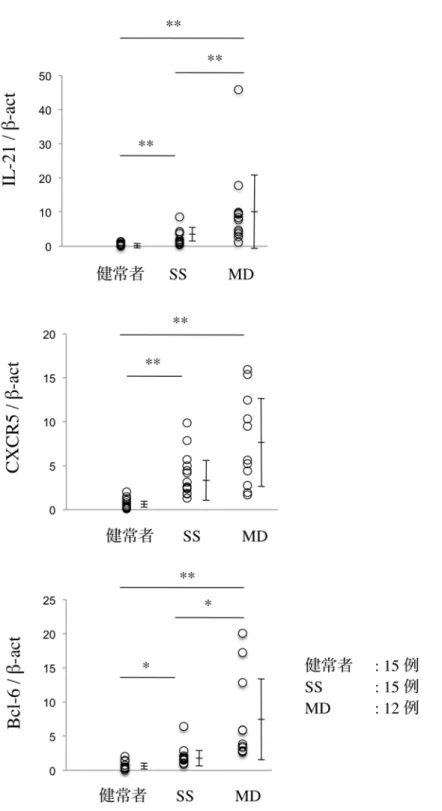
まずは、SS と MD の LSG において、real-time PCR 法および免疫組織化 学染色法にて、IL-21 とその主な産生細胞であり GC 形成に深く関与してい るTfh (19) の発現を、比較検討した。SS および MD 患者では、IL-21、CXCR5、
および Bcl-6 の発現は、健常者と比較して有意に亢進していた。さらに、MD 患者の IL-21 および Bcl-6 の発現は、SS 患者と比較して有意に亢進してい た (図5a、P<0.05、P<0.01; Mann-Whitney の U 検定) 。
まず、組織像を示すが (図 5b)、SS では、導管周囲のリンパ球浸潤と腺房 および導管の破壊と消失がみられ、なかには GC を形成しているものもみら れた (B)。一方 MD では、SS と同様に強いリンパ球の浸潤を認めるものの、
GC の形成が高頻度で認められ、SS で典型的な導管の破壊はほとんど認めら れなかった (C)。次に、real-time PCR 法にて発現が有意に亢進していた分子 の LSG 内での局在を明らかにするために、免疫組織化学染色を行った (図 5b)。SS では、GC にIL-21、CXCR5、および Bcl-6 の発現を認めた (E、H、
K)。一方 MD では、SS と同様に GC に IL-21、CXCR5、および Bcl-6 の 発現を認めたが (F、I、L)、IL-21 は、GC の形成に関わらず LSG 全体に、
強い発現を認めた (F)。つまり、MD における IL-21 産生細胞は Tfh の他に もある可能性が示唆された。IL-21 は主に Tfh が産生するといわれているが、
その他にもIL-21 産生能を有するとの報告がある Th2 (58) および Th17 (59) に ついても、同様の検討を行った。
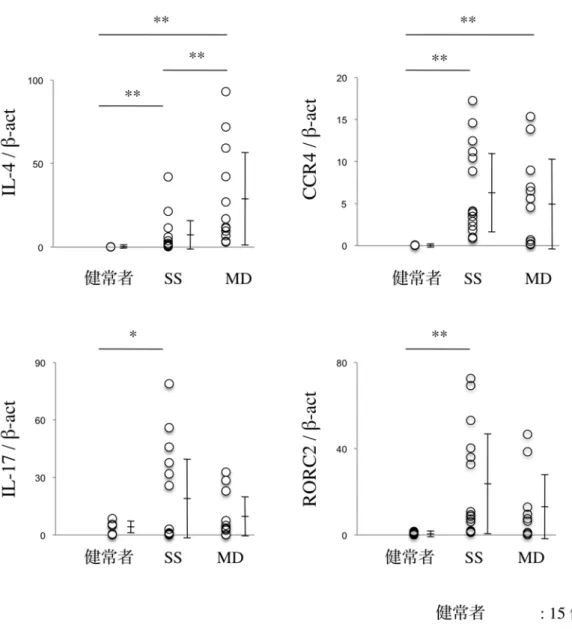
まずは、SS と MD 患者の LSG において、real-time PCR 法にて、Th2 関 連分子 (IL-4、CCR4) と Th17 関連分子 (IL-17、RORC2) の mRNA 発現を 比較検討した。 その結果 SS では、Th2 関連分子 (IL-4、CCR4) と Th17 関 連分子 (IL-17、RORC2) が、健常者と比較して有意に亢進していた。 一方、
MD では、Th2 関連分子 (IL-4、CCR4) が、健常者と比較して有意に亢進し ていた。さらに MD では、IL-4 が SS と比較して、有意に亢進していた (図 6a、P<0.05、P<0.01; Mann-Whitney の U 検定)。
次に、Th2 および Th17 関連分子の局在を明らかにするために、免疫組織 化学染色を行った (図 6b)。その結果 SS では、IL-4 および CCR4 の発現が
LSG 全体に認められ、特に GC で強い発現がみられた (B、E)。さらに Th2 の転写因子 (60) であり、IL-21 産生に直接関与しているといわれている c-Maf
(61) も、GC で発現がみられた (H)。IL-17 は GC で弱い発現がみられた (K)。
一方 MD では、Th2 関連分子 (IL-4、CCR4、c-Maf) は、IL-21 と同様に (図 5b F)、GC および LSG 全体に強い発現がみられた (C、F、I)。一方 IL-17 の 発現は、ほとんどみられなかった (L)。
図 5a SS、MD、および健常者の LSG における IL-21、CXCR5、
Bcl-6 の mRNA 発現量
定量的 real-time PCR を行い、β-actin に相対的な mRNA 発現量を算出し、
SS、MD、および健常者のそれぞれ 2 群間で Mann-Whitney の U 検定を行
った (*P<0.05、**P<0.01)。実験方法は「材料と方法」に示す。
図 5b SS、MD、および健常者の LSG における IL-21、CXCR5、 Bcl-6 の発現
(代表症例のパラフィン包埋標本を用いた免疫組織化学染色: scale bar 100μm) SS および MD の GC では IL-21、CXCR5、および Bcl-6 の強い発現を認 めた (E、F、H、I、K、L)。さらに MD では、GCの形成にかかわらず、LSG 全体に IL-21 の強い発現を認め (F)、同部位には CXCR5 および Bcl-6 の発 現は、認められなかった (I、L)。実験方法は「材料と方法」に示す。
図 6a SS、MD、および健常者の LSG における IL-4、CCR4、IL-17、 RORC2 の mRNA 発現量
定量的 real-time PCR を行い、β-actin に相対的な mRNA 発現量を算出し、
SS、MD、および健常者のそれぞれ 2 群間で Mann-Whitney の U 検定を行
った (*P<0.05、**P<0.01)。実験方法は「材料と方法」に示す。
図 6b SS、MD、および健常者の LSG における IL-4、CCR4、 c-Maf、IL-17 の発現
(代表症例のパラフィン包埋標本を用いた免疫組織化学染色: scale bar 100μm) SS では、IL-4 および CCR4 は、LSG 全体に発現を認めたが、特に GC で 強かった (B、E)。c-Maf は、GC で特に強い発現が認められた (H)。IL-17 は GC で弱い発現がみられた (K)。一方 MD では、GC と LSG 全体に IL-4、
CCR4、および c-Maf の強い発現を認め (C、F、I)、IL-17 はほとんど発現が
みられなかった (L)。実験方法は「材料と方法」に示す。
2-2. IgG4 産生に関わる分子について
前述のとおり、IL-21 が、培養 B 細胞からの IgG4 産生のクラススイッチ に関与しているといわれている (42, 43, 44)。そこで IL-21 と病変局所の IgG4 産 生との相関について検討した。
まず、SS および MD の LSG を用いて、IgG4 の mRNA 発現量と IL-21 の mRNA 発現量との関連について検討したところ、SS では両者の間に、相 関は認められなかったが、MD では正の相関を認めた (図7a)。
次に、SS および MD の LSG における IgG4 陽性形質細胞の局在を明ら かにするために、免疫組織化学染色を行った (図7b)。その結果 SS では、IgG4 陽性形質細胞の浸潤はごくわずかであった。これに対し MD では、びまん性 に浸潤が強く認められた (図7b)。
最後に、MD の IgG4 陽性細胞率と IL-21 の mRNA 発現量の関連につい て検討を行ったところ、両者の間に正の相関関係を認めた (図7c)。
以上の結果より、MD における唾液腺病変局所では、IL-21 が IgG4 産生 に関与している可能性が示唆された。
図 7a SS 患 者 と MD 患 者 の LSG に お け る IgG4 と IL-21
mRNA 発現量の相関
定量的 real-time PCR を行い、相対的な mRNA 発現量を算出した。IgG4 と IL-21 と の 相 関 関 係 を 示 し て い る 。Spearman 順 位 相 関 係 数 (*P<0.05、
**P<0.01). NS: not significant実験方法は「材料と方法」に示す。
図 7b SS 患者および MD 患者の LSG における IgG、IgG4 陽性 細胞の発現
(代表症例のパラフィン包埋標本を用いた免疫組織化学染色: scale bar 50μm) SS と MD ともに強いリンパ球の浸潤を認める。MD では、IgG4 陽性細胞 は LSG 全体にびまん性に存在していた。IgG4 陽性細胞率を算出した。実験 方法は「材料と方法」に示す。
図 7c MD 患 者 の LSG に お け る IgG4 陽 性 細 胞 率 と IL-21 の
mRNA 発現量の相関
定量的 real-time PCR を行い、相対的な mRNA 発現量を算出した。IgG4 陽 性細胞率と IL-21 との相関関係を示している。Spearman 順位相関係数 (*P<0.05、**P<0.01). 実験方法は「材料と方法」に示す。
考 察
Mosmann と Sad がサイトカイン産生様式により Th1 細胞と Th2 細胞に
大別できる事を報告して以来 (62)、多くの免疫反応による病態が Th1 病ある いは Th2 病という概念で理解されるようになり、Th1/Th2 バランスが疾患の 発症、進行、予後に重大な影響を及ぼすことが明らかにされてきた。しかし、
Th1・Th2 のいずれでもない IL-17 を産生する Th17 と呼ばれる新たな Th
サブセットの発見により、これまで Th1 より産生される IFN-γ や TNF によ って制御されていると考えられてきた SS、慢性関節リウマチ、多発性硬化症、
乾癬などの多くの自己免疫疾患が、この Th17 によって制御されていること が明らかにされている (63)。さらに、免疫系には免疫抑制機能を有した Treg が存在し、自己免疫、炎症、アレルギーといった病的な免疫応答を抑制的に 制御することで免疫恒常性の維持に重要な機能を担っていることが明らかに されてきた (64)。そして最近では、二次リンパ組織濾胞胚中心に存在し、Ig の 産生を調節する Tfh が報告された (19)。この Tfh は IL-21 を産生する特異な Th サブセットであり、その寛容の破綻が自己反応性 B 細胞を活性化し、自 己免疫疾患の発症を誘導することが示されている (65, 66)。従来 SS の発症には、
Th 細胞が重要な役割を担っていることが指摘されており (4, 5)、この Th 細胞 が、唾液腺上皮細胞に発現した HLA class Ⅱ 分子を認識することで炎症が惹 起されると考えられている (67)。つまり SS の病態形成のメカニズムとこれら の Th 細胞 (Th1、Th2、Th17、Treg、Tfh) との関わりを明確にすることは、
SS の発症や病態進展のメカニズムを解明することに役立つと考えられる。そ こで本研究では、研究 1. として SS 患者の LSG における Th サブセット
の発現と局在を明確にすることとした。以下にSSとそれぞれの Th サブセッ トとの関連について考察を述べる。
・Th1、Th17細胞について
SS の病態形成には、主として Th1 が強くかかわっていると理解されてき た。これは、SS のモデルマウスにおいて Th1 を欠損させると SS の臨床症 状や病理学的所見において病態が改善する報告や (68)、SS 患者の LSG のサ イトカイン発現を調べた結果、Th1 関連のサイトカイン (IFN-γ、IL-2、IL-12、
IL-18) が高発現するなどの報告から Th1 関連疾患であるとのコンセンサス
を得ている (69)。また、われわれは過去の研究で (1, 29, 52, 70, 71)、Th1 細胞と標的 組織である導管上皮が相互に刺激し合って、種々のサイトカインを産生する ことが、SS の発症と維持に重要であり、その結果、導管上皮の破壊が生じる ことを報告している。さらに近年では、Th17 細胞も SS の LSG に多数認め られ、Th17 の分化・増殖に必須とされているサイトカイン (IL-1β、IL-6、
TGF-β、IL-23) の発現が病変局所で亢進しているなど、Th17 を分化・増殖す
る環境にあるといわれている (69, 72, 73)。今回の研究結果では、SS 患者のLSG で Th1 および Th17 関連分子の発現が亢進していたが、リンパ球浸潤程度と の関連は認めなかった。また、LCMを用いた病変局所の解析では、導管上皮 周囲に浸潤したリンパ球において、 Th1および Th17関連分子が亢進してい た。ヒトの自己免疫疾患においては、初期段階の病変局所を探すことは困難 であるために、その病変局所における Th 細胞の働きは不明な点が多いが (69,
74, 75)、本研究によりSS の導管上皮周囲に浸潤した Th1 と Th17 両者が、病
態進展ではなくその発症に重要な役割を担っていることが強く示唆された。
・Th2、Tfh細胞について
われわれの過去の研究では、病態進展には Th2 が産生するサイトカインが 重要な役割を果たしていることを示してきた (29)。つまり Th2 が加わってく ると、自己抗原反応性の B 細胞が増殖し、SS のリンパ増殖性病変としての 病態の進展に結びつくと考えている。さらに、EBウイルス (Epstein-Barr virus)
(76)、サイトメガロウイルス (Cytomegalovirus) (77)、レトロウイルス (Retrovirus)
(78) などの組織指向性ウイルスに関連する環境要因が、偽リンパ腫や高 γ グ ロブリン血症などの病態形成のリスクを増加させ、最終的には SS を B 細 胞リンパ腫へと進展させる。また最近では、Tfh に関する見解も集積され、
マウスにおいて IL-4 ならびに IL-21 が共同して体液性免疫を担っていると いわれている (79)。また SS の血清中のIL-21 は、リンパ球の浸潤が軽度の症 例と比較して、重度において亢進しているとの報告もある (80)。今回の研究で も、リンパ球の浸潤が重度の症例において、Th2 や Tfh 関連分子の発現が亢 進しており、さらにLCMを用いた病変局所の解析では、GC を形成している リンパ球において、Th2 および Tfh 関連分子の発現が全症例で亢進していた。
これらの結果より、導管上皮に Th1 および Th17 が集積し、それに加えて Th2 および Tfh が集積してくることで、病態が進展し、特にGC の形成に重 要な役割を果たしていることが強く示唆された。
・Treg 細胞について
一方で、Treg 細胞と SS との関わりについては過去の報告でも見解が一致
していない。SS 患者の LSG ならびに末梢血における Treg の減少が唾液腺
病変の病因に関与している (81) という報告がある。一方で、Treg は、SS の 末梢血において健常者と比べ上昇している (82) という報告もある。さらに Th1 と Th2 は、Treg による抑制を受けやすいが、Th17 は、Treg の免疫抑 制活性に耐性であるといわれている (83)。最近の報告では、SS をリンパ球浸 潤程度で 3 群 (軽度、中等度、重度) で分けて比較したところ (84, 85)、リンパ 球浸潤が軽度から中等度に進展すると、Treg の発現は亢進していたが、逆に 重度になると低下するというものであった。今回の結果では、Treg はリンパ 球浸潤程度間では有意な差は認められなかったが、健常者と比較するとSS 患 者で発現が亢進しており、これは病態形成を抑制するように Treg が働いて おりネガティブフィードバック機構が存在する可能性が考えられる。これら を考えあわせると、SS における Treg の免疫学的役割は、発症や病態の進展、
特に GC の形成に直接関与してはいないと考えられるが、病態進展のステー ジによっては働きが異なる可能性があることが示唆された。
以上の結果により、SS の唾液腺局所において Th サブセットの明確な局在 を認めることが明らかになった。これらのことから、SS の病態モデルについ て考えてみると、まず Th1 細胞と Th17 細胞が標的組織である導管上皮と相 互に刺激し合い、種々のサイトカインを産生することによって、SSの発症と 維持に重要な役割を果たし、その結果、標的組織である唾液腺の破壊を生じ る。その後、Tfh 細胞や Th2 細胞が集積することにより、GC の形成が促進 され、さらには B 細胞の分化、増殖、および抗体産生が刺激され、SS のリ ンパ増殖性病変としての進展に結びつくということが示唆された。病態進展 の機序を解明するには、更なる解析が必要であるが、さらに研究が進めば、
病態進展の抑制に有効な治療戦略に繋がるのではないかと考えている。また SSの複雑な因果関係がさらに解明されれば、病態進展に関わる特異的なサイ トカインネットワークを遮断する薬物を用いた、治療法が確立するかもしれ ない。
一方、MD は SS と同様に、唾液腺・涙腺にリンパ球の著明な浸潤を伴う 腫脹を呈するが、前述のように、病理組織学的特徴として SS より高頻度に GC 形成が認められる。この GC 形成には IL-21 が必須であり、IL-21 によ って B および T 細胞に Bcl-6 を発現させ、さらに B 細胞には、形質細胞 へ転換するのに必要な B lymphocyte-induced maturation protein-1 (Blimp-1) も 発現させるといわれている (41)。つまり、IL-21 は、Ig の産生および GC の 形成に深く関与していることになり、MD の病態に深く関与している可能性 があると推察される。
IL-21 の主な産生細胞は、活性化 CD4+ T 細胞であるといわれている (86)。
過去に、Wurster らは、Th 細胞のなかで、Th1 細胞と Th2 細胞のいずれの 細胞分画が IL-21 産生細胞なのかについて検討し、Th2 が Th1 に比べて IL-21 mRNA を強く発現しており (58)、IL-21 の主要な産生細胞は Th2 であ ると考えられた。しかし Th17 細胞が同定されると、IL-21 は Th17 細胞分 化誘導条件下で主に産生され、その自己増殖因子として機能することが報告 され (59)、Th17 が主な産生細胞であると考えられるようになった。しかしな がら、興味深いことに Th17 細胞における IL-21 の発現は Th17 細胞の転写 因子である RORγt 非依存性であり IL-17 とはことなる制御を受けていると いわれていた (87)。最近、須藤らはIL-21 産生細胞の多くが Th17 に特異的な
IL-17 の発現を欠いていたことから、IL-21 を産生している細胞は Th17 と異 なる細胞集団であり、その細胞集団が Tfh 細胞の特徴と似ていることを報告 した (88)。最近ではいくつかの報告でTfh 細胞が、IL-21 を特異的に産生する といわれている (65, 89, 90)。また IL-21 レセプターは、B 細胞と NK 細胞、さ らに一部の T 細胞株に発現している (86)。今回の研究結果からも、SS では
IL-21 と Tfh 関連分子ともに GC に限局していることから、Tfh が主体とな
りIL-21 を産生し、 GC の形成に関与していることが考えられ、過去の報告
(65, 89, 90) とも矛盾しないものであった。しかしながら、MD では興味深いこと
に、Tfh関連分子はGCに限局していたものの、IL-21はGC の形成に関わら ず、LSG 全体に強い発現を認めた。そこで、IL-21 の産生細胞(Th2、Th17)
について検討すると、Th2関連分子のみにIL-21 の発現と同様にLSG 全体に 発現しており、MD のGC 形成や病態形成には、Th2 を主体とするIL-21 の 過剰な産生が関与していることが示唆された。
Ig の中で最も多い IgG は 1 から 4 のサブクラスを持つが、健常人の血 中における IgG4 は、総 IgG のおよそ 4% 前後と最も量が少ない (91)。IgG4 は、抗原に対する親和性が低く、さらに補体の C1q 部に結合できないといっ た特徴を持っている (92)。IgG4 の産生は、Th2 が産生する IL-4が B 細胞に 作用して誘導されることが知られているが、同時にその IL-4 は IgE の産生 も誘導している。しかし最近の研究では、アレルギー患者に対する特異的免 疫療法を行うと、その抗原特異的な IgG4 が上昇することが明らかになって きた。この作用機序として、Th2 や Treg が産生する IL-10 が、IL-4 による IgE 産生を抑制し、かわりに IgG4 産生を促進するというクラススイッチを
引き起こすことが報告されている (93, 94)。また、IL-10 そのものが B 細胞の
IgG1 産生を抑制することで、かわりに IgG4 の産生を促進させることも併せ
て報告されている (95)。われわれの最近の研究でも、Th2 が産生する IL-4 と
Treg が産生する IL-10 が、 MD の特徴である IgG4 へのクラススイッチに
関与していることを報告しているが (40)、 前述のとおり、IL-4 や IL-10 に加 えて IL-21 も IgG4 へのクラススイッチに関与していることが明らかにな
ってきた(42-44)。さらには、IL-4 と IL-21 の共刺激により、IL-10 産生が上昇
するという報告もあり (44)、本研究でもMD における高 IgG4 血症および腺組 織中への著明な IgG4 陽性細胞浸潤とIL-21 産生との関係について注目した。
その結果、MD における IgG4 の発現と IL-21 の発現には正の相関を認めた ことから、IgG4 の産生にはIL-21が深く関与している可能性が示唆された。
以上のように、MD と SS は免疫学的に明らかに異なる病態を呈しており、
MD は SS と比較して Th2 優位な疾患であることが推察される。これまで のわれわれの報告 (40) もふまえて、 MD の病態形成のメカニズムを考えると、
Th2 が産生する IL-21 が過剰に発現することで、B 細胞および T 細胞が活 性化され、Bcl-6 の発現が誘導される。さらには Tfh が産生する IL-21 も加 わり、唾液腺に GC が多数形成される。その GC 内では、Th2 が産生する IL-4 と IL-21 に加え、 Treg が産生する IL-10、そして Tfh が産生する
IL-21 が、B 細胞を IgG4 産生形質細胞へ分化させ、高 IgG4 血症へと至る
という病態モデルを考えることができる。
本研究のように、腺組織における Th サブセットの発現や局在を明確にす ることは、MD の病態解析に有用であることが示唆されたが、発症や病態進
展のメカニズムを解明するには、MD 患者の腺組織を用いたin vivoでの検討 や MD モデルマウスの開発など、さらなる検討が必要である。
謝 辞
稿を終えるにあたり、御懇篤なる御指導を頂きました九州大学大学院歯学 研究院口腔顎顔面病態学講座顎顔面腫瘍制御学分野の中村誠司教授に深甚な る謝意を表します。
また直接御指導頂きました林田淳之將先生、森山雅文先生に深謝致します。
さらに常に励ましの言葉を頂きました同分野の皆様に深く感謝致します。
最後に、蔭ながら研究を支えてくれました家族と妻に感謝致します。