1 INTRODUCTION
Alveolar reconstruction of bony defects, including use of autogeneous bicortical mandibular symphyseal bone grafts in patients with cleft lip and palate is a widely accepted treatment regimen
1. Autogenous bone grafts are the gold standard for such reconstruction because of their osteoconductive, osteoinductive, and non-immunogenic properties
2. Although autologous bone grafting is widely performed for bone reconstruction in the maxillofacial field, it is invasive and there is a limited amount
of collectable bone. Therefore, bone formation using bone marrow-derived mesenchymal stem cells (MSCs) has attracted attention as an alternative procedure.
MSCs are fibroblast-like cells isolated from the bone marrow, which attach to tissue culture surfaces
3. Human MSCs are multipotent and have been differentiated into several kinds of mesodermal tissues, including the bone, fat, and cartilage in vitro
4. Induced pluripotent stem cells (iPSCs) and embryonic stem cells (ESCs) have great
Induction of Mesenchymal Stem Cells-like Cells Derived from Human Gingival iPS
Cells into Osteoblast-like Cells
Yuji NAKAYAMA
1, Yoshiya HASHIMOTO
2, Yoshitomo HONDA
3, and Naoyuki MATSUMOTO
11
Department of Orthodontics,
2Department of Biomaterials,
3
Institute of Dental Research, Osaka Dental University, Osaka, Japan
SYNOPSIS
Mesenchymal stem cells (MSCs) have limited capacity to proliferate in vitro, making it very difficult to acquire sufficient cell numbers for implantation. The objective of this study was to induce MSC-like cells (MSLCs) derived from human gingival induced pluripotent stem (HG-iPSCs) into osteoblast-like cells. We derived MSLCs from HG-iPSCs under feeder-free conditions. MSLCs expressing MSC-specific markers treated with osteoblast differentiation factor expressed osteoblast-specific mRNA and protein level when cultured with or without bone morphogenetic protein (BMP). BMP-2 had a positive effect on the expression of early indicators of osteogenic differentiation. However, the medium without BMP-2 promoted calcium deposition, which occurs at the terminal stage of osteogenic differentiation. MSLCs are likely to have higher proliferative capacity, and hence, in vitro-differentiated osteoblast-like cells and appropriate scaffolds can be used for bone tissue engineering.
Key words: mesenchymal stem cells-like cells, mesenchymal stem cells, human
gingival, iPS cells, osteoblast-like cells
potential in tissue engineering because they proliferate indefinitely while retaining their pluripotency, and can differentiate into all cell types found in the body
5. However, there are many problems to be resolved regarding these pluripotent cell types, including ethical issues and tumorigenic transformation.
Recently, some groups have successfully established MSC-like cells (MSLCs) from ESCs/iPSCs
6-9. MSLCs derived from iPSCs are expected to be a promising new stem cell population incorporating the advantages of both iPSCs and MSCs. Hynes et al.
6demonstrated that MSLCs were successfully generated from human iPSCs, and that MSLCs exhibited the capacity to regenerate periodontal tissues in a rat periodontal defect model.
In our previous study
10, human gingival iPSCs (HG-IPSCs) were generated using non-integrating episomal plasmid vectors, and were successfully differentiated into MSLCs. The MSLCs expressed key MSC-associated markers, and had the capacity to undergo tri-lineage differentiation.
However, the osteoblast differentiation potential of MSLCs was not elucidated.
The aim of this study was to induce the differentiation of MSLCs into osteoblast-like cells with or without the
presence of bone morphogenetic protein, and to elucidate their osteoblast differentiation potential using gene and protein assays.
MATERIALS AND METHODS
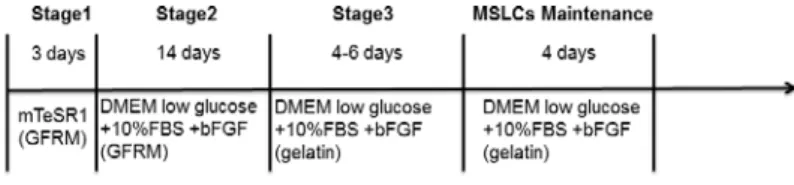
Derivation of HG-iPSCs into MSLCs Derivation of HG-iPSCs into MSLCs was carried out using the method of Umezaki et al.
10( Fig. 1 ) . Briefly, HG-iPSCs were cultured in mTeSR1 (StemCell Technologies, Vancouver, BC, Canada) on growth factor-reduced Matrigel (GFRM; BD Biosciences, San Jose, CA, USA) without feeders for 3 days. The medium was replaced with Dulbecco's modifed Eagle's medium (DMEM)-low glucose (Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS; Gibco) and 10 ng/mL Basic fibroblast growth factor (b-FGF) (PeproTech, Rocky Hill, NJ, USA), and changed every 2 days. Cells were trypsinized (0.025% trypsin Gibco, Grand Island, NY, USA) and passaged after 2 weeks and transferred to the same medium on a gelatin-coated plate.
After passaging three times, the cell morphology changed to a fibroblastic shape (Fig. 2). This study was approved by the ethics committee of Osaka Dental University (approval number: 110763).
Fig. 1 The method for differentiation of HG-iPSCs into MSLCs.
Fig. 2
Morphology of MSLCs derived from HG-iPSCs.
Scale bar: 200 μm.
3 Flow cytometry for MSLCs
We investigated whether HGF-iPSCs can be derived into MSLCs. Cells (2.5 × 10
5cells/mL) were obtained after trypsinizing (0.025% trypsin Gibco, Grand Island, NY, USA). Cell surface antigen staining was performed in phosphate buffered saline (PBS) with 2% human serum albumin. The cell suspension was incubated with specific antibodies for 30 min at 4 °C. Murine anti-human antibodies were used at the recommended concentrations. We investigated the following human mesenchymal markers of MSLCs: CD44 (400112; BioLegend, San Diego, CA, USA), CD73 (344006; BioLegend), CD90 (12-0909-42; Thermo Fisher Scientific Inc., Waltham, MA, USA), CD105 (323206; BioLegend), CD34 (343510; BioLegend), and CD45 (304008; BioLegend). The stained cells were analyzed with FACSVerse (BD Biosciences, San Jose, CA, USA), and data were analyzed using the FlowJo software (BD Biosciences).
LIN28 gene expression
LIN28 expression was evaluated to detect of residual undifferentiated hiPSCs using the QuantStudio 3D digital PCR system (ddPCR; Thermo Fisher Scientific Inc.) with the QuantStudio 3D Master Mix (Thermo Fisher Scientific Inc.) and gene expression assay (Thermo Fisher Scientific Inc.). The cycling conditions were as follows: 10 min at 96 C, 39 cycles of 2 min at 60 C, 30 s at 98 C, and 2 min at 60 C. The assay ID of TaqMan Gene Expression Assay and the assay ID used for the qPCR and digital PCR systems is Hs00702808_s1 (Applied Biosystems, Foster City, CA, USA).
Osteogenic differentiation of MSLCs To evaluate the osteogenic potential of MSLCs at passage 3, they were replated in standard medium in a
24-well plate at a density of 5 × 10
4cells/well and incubated at 37 °C with 5% CO
2. At confluency, cells were cultured in either control medium;
DMEM-high glucose supplemented with 10% FBS (Gibco) and antibiotic/
antimycotic mixed stock solution (control group), osteogenic medium; control medium supplemented with 50 μM L-ascorbic acid 2-phosphate (Sigma- Aldrich, St. Louis, MO, USA), 10 mM
-glycerophosphate (Nacalai Tesqe, Kyoto, Japan), and 100 nM dexamethasone (non BMP group) (Wako Pure Chemical Industries, Osaka, Japan); or osteogenic medium with BMP-2; osteogenic medium supplemented with 100 ng/mL BMP-2 (BMP group)( PeproTech, Rocky Hill, NJ, USA) for days 7, 14, and 21.
Quantitative real-time polymerase chain reaction (PCR)
Total RNA was isolated using a QIAcube (QIAGEN, Limburg, The Netherlands) according to the manufacturer’s protocol, and single-stranded cDNA was synthesized using a Transcripor Universal cDNA Master (Roche). mRNA levels were analyzed by quantitative real-time PCR using a TaqMan
Gene Expression Assay (COL1A1, Hs00164004_m1; ALP, Hs00240993_
m1; RUNX2, Hs01047973_m1; OCN, Hs00609452_g1; Applied Biosystems) on a StepOnePlus PCR system (Applied Biosystems). The glyceraldehyde-3- phosphate dehydrogenase (GAPDH) gene was co-amplified as an internal standard (human GAPDH endogenous control; Applied Biosystems). Gene expression was measured using the
Ct method.
DNA content analysis
DNA content was measured on days 7,
14, and 21. The medium was removed
and the cells were washed twice with
PBS. Then, 300 μL of 0.2% Triton X-100 was added to each well, and the cells were removed for DNA extraction. DNA content was measured by Quant-iT™
PicoGreen dsDNA Reagent and Kit (Invitrogen, Life Technologies, Carlsbad, CA, USA). Fifty microliters of a sample was mixed with a DNA-binding fluorescent dye solution (0.5 μL Picogreen reagent in 100 μL 1× Tris- EDTA buffer), and the fluorescence intensity was measured by a microplate reader (Excitation 450 nm/Emission 510 nm; SpectraMax M5; Molecular Devices, Sunnyvale, CA, USA).
Measurement of alkaline phos- phatase (ALP) activity
ALP activity was measured by LabAssay™ ALP kit (Wako Pure Chemical Industries). One-hundred microliters of working assay solution (6.7 mmol/L p-nitrophenylphosphate disodium) was added to 20 μL of the same sample used for DNA measurement, and then mixed thoroughly and incubated at 37 °C with 5% CO
2for 15 min. After 80 μL of stop solution (0.2 mol/L sodium hydroxide) was added to the mixture, the absorbance was measured at 405 nm with the SpectraMax
M5. The levels of ALP were normalized to the amount of total DNA in the cell lysates.
Measurement of osteocalcin (OCN) MSLCs were cultured under the same conditions as described in DNA content analysis, and then the expression of OCN in both cell types was determined by a Human Gla-Osteocalcin High Sensitive EIA Kit (Takara Bio Inc., Shiga, Japan). On days 7, 14, and 21 of culture, the medium was removed, cells were washed with PBS, 300 μL of 10% formic acid was added to each well, and cells were removed. Samples (100 μL) were added to each well of an anti-OCN
antibody-coated microtiter plate and then incubated at 37C for 2 h. After three washes with PBS, 100 μL of a peroxidase-conjugated anti-OCN antibody solution was added to each well, followed by incubation at room temperature for 1 h. After four washes with PBS, 100 μL of substrate solution (tetramethylbenzidine) was added to each well, followed by incubation at room temperature for 15 min. After 100 μL of stop solution was added to each well, the absorbance was measured at 450 nm with the SpectraMax
M5.
.
Alizarin red staining
On day 21 of the culture period, the medium was removed and cells were fixed with 95% ethanol for 10 min at room temperature. After the fixing solution was removed, the cells were rinsed with water and stained with alizarin red S (Sigma-Aldrich) for 30 min at room temperature. After staining, the cells were rinsed with water six times.
After 400 μL of 10% formic acid was added to each well for 10 min, the absorbance was measured at 415 nm with the SpectraMax
M5.
Statistical analysis
All experiments were conducted in quadruple and repeated at least twice.
All data were expressed as the mean and standard deviation. Differences were evaluated by analysis of variance with Tukey’s test. Differences were considered significant at p < 0.05.
RESULTS
Differentiation of HG-iPSCs into MSLCs
We investigated whether the
fibroblast-like cells differentiated from
iPSCs were MSLCs. MSLCs and bone
marrow-derived MSCs (BMMSCs) were
analyzed by flow cytometry for the
expression of human mesenchymal
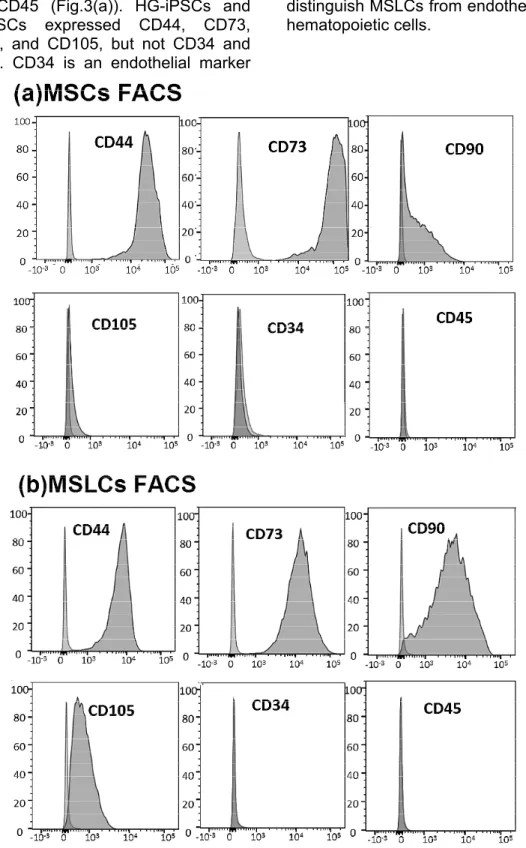
5 markers, such as CD44, CD73, CD90, and CD105, and endothelial and hematopoietic markers such as CD34 and CD45 (Fig.3(a)). HG-iPSCs and BMMSCs expressed CD44, CD73, CD90, and CD105, but not CD34 and CD45. CD34 is an endothelial marker
and CD45 a hematopoietic marker, both of which are not expressed in MSCs (Fig.3(b)). Therefore, we used them to distinguish MSLCs from endothelial and hematopoietic cells.
Fig. 3 Flow cytometry analysis of MSC-related surface markers (CD44, CD73, CD90, and CD105) and hematopoietic markers (CD34 and CD45). (a) MSCs (b) MSLCs.
Gene expression analysis LIN28 expression
A ddPCR that directly quantifies DNA/
mRNA copies has been developed. The ddPCR takes advantage of water- in-oil droplet templates, and specific cleavage of TaqMan probes generates a strong fluorescence signal to divide a 20-μL mixture of sample and reagents into
~20,000 droplets. The copy numbers of LIN28-positive expression were only 11.5 per 20,000 copies, which was not expressed.
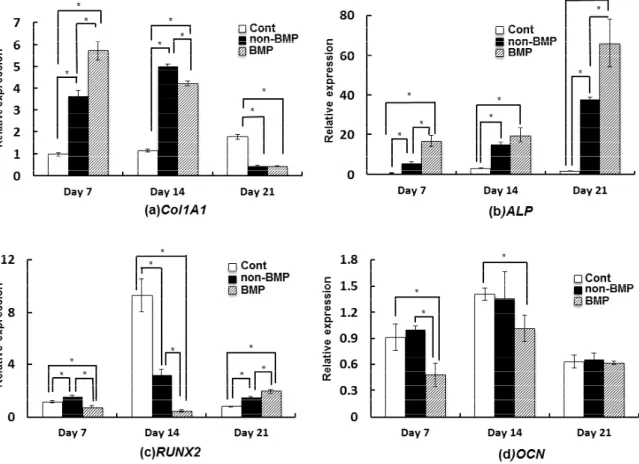
Col1A1 expression
Col1A1 expression of MSLCs cultured in BMP and non-BMP groups was the highest on days 7 and 14, respectively (p < 0.05). However, Col1A1 expression of MSLCs cultured in control group was the highest on day 21 (p < 0.05) (Fig.
4(a)).
ALP expression
ALP expression of MSLCs cultured in BMP group was the highest on all days cultured, as compared to other groups (p < 0.05) (Fig. 4(b)).
RUNX2 expression
RUNX2 expression of MSLCs cultured in non-BMP, control, and BMP groups (control medium) was the highest on day 7, 14, and 21 (p < 0.05), respectively (Fig. 4(c)).
OCN expression
OCN expression of MSLCs cultured in BMP group was the lowest on day 7. No significant difference was observed in the OCN expression between different groups on days 21 (Fig. 4(d)).
Fig. 4 mRNA expression profiles: (a) Col1A1 expression (b) ALP expression (c) RUNX2 expression and (d) OCN expression of MSLCs cultured in control, non-BMP, and BMP. Data are expressed as the mean and standard deviation. *p < 0.05.
7 ALP activity analysis using Enzyme-Linked ImmunoSorbent Assay
ALP activity of MSLCs cultured in BMP group was the highest on day 14. ALP activities of MSLCs cultured in BMP and non-BMP groups were higher than those of control group on all days (p < 0.05) (Fig. 5(a)).
OCN content analysis using Enzyme-Linked ImmunoSorbent Assay
The OCN content of MSLCs cultured in BMP group was the lowest and the highest on days 7 and 21, respectively (p < 0.05) (Fig. 5(b)).
Alizarin red staining
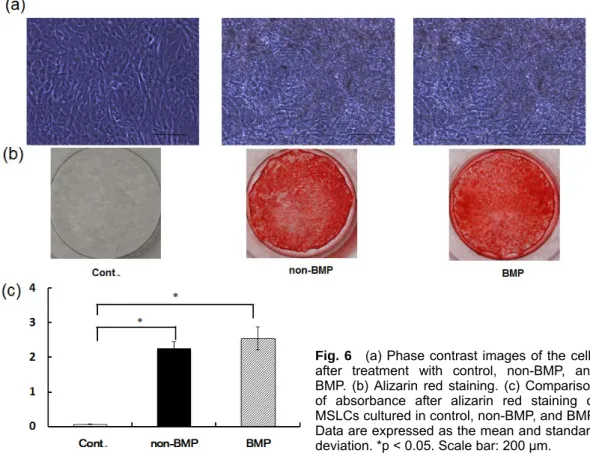
Under a phase contrast microscope,
Fig. 5 (a) ALP activity and (b) OCN contents of MSLCs cultured in control, non-BMP, and BMP. Data are expressed as the mean and standard deviation. *p < 0.05.
Fig. 6 (a) Phase contrast images of the cells after treatment with control, non-BMP, and BMP. (b) Alizarin red staining. (c) Comparison of absorbance after alizarin red staining of MSLCs cultured in control, non-BMP, and BMP.
Data are expressed as the mean and standard deviation. *p < 0.05. Scale bar: 200 μm.
some mineralized nodule formation was observed in MSLCs cultured in non-BMP and BMP groups on day 21 (Fig. 6(a)). Red areas stained by alizarin red are an indication of calcium deposi- tion. The plates containing MSLCs cul- tured in non-BMP and BMP groups for 21 days were stained strongly using alizarin red (Fig. 6(b)). No significant difference in absorbance was observed between non-BMP and BMP groups, however the absorbances in non-BMP and BMP groups were higher than that in control group (Fig. 6(c)).
DISCUSSION
Gingival tissues are routinely resected during general dental treatments, such as tooth extraction, periodontal surgery, and dental implantation, and are gener- ally treated as biomedical waste
11. Egusa et al.
12successfully derived iPSCs from human gingival using retro- viral transduction of transcription factors.
However, retroviral integration increases the risk of tumor formation, while in- tegration-free methods decrease the potential risk
13. The development of novel approaches for generating inte- gration-free iPSCs has eliminated the concern of integrating virus- associated genotoxicity in clinical applications
14. In our previous study, HG-IPSCs were generated with non-integrating episomal plasmid vectors and were differentiated into MSLCs. In the present study, the effective culture conditions for the induction of MSLCs derived from HG-IPSCs into osteoblast-like cells were investigated.
We differentiated HG-iPSCs into MSLCs using the culture method of Umezaki et al.
10. MSLCs expressed the human mesenchymal markers, but did not express endothelial and hematopoi- etic markers, such as CD34 and CD45, as confirmed by flow cytometry analysis.
Lin28 is highly expressed in human ESCs and, along with three other stem
cell marker proteins (OCT4, NANOG, and SOX2), can induce pluripotency in human somatic cells
15. LIN28 is a highly sensitive marker of residual undifferen- tiated human iPSCs (hiPSCs) in hiPSC-derived cell therapy products
16. The ddPCR assay using LIN28 as a target is able to detect extremely slight numbers of undifferentiated hiPSCs
16. In this study, MSLCs derived from HG-IPSCs did not express LIN28 might be a safe cell therapy.
Bone-like tissue is synthesized by BMMSCs cultured in medium containing ascorbic acid, -glycerophosphate, and the synthetic glucocorticoid dexa- methasone
17. Real-time PCR analysis showed that MSLCs cultured in non-BMP group containing osteoblast induction factor expressed the osteo- genic marker Runx2, suggesting that MSLCs retain the properties of osteoblast lineage-committed progenitor cells. The mature human BMP-2 is an acidic glycoprotein, and is well known to increase the expression of osteoblast indicators in pluripotent stem cell cul- tures
18. In the present study, the osteoblast differentiation potential of MSLCs cultured with or without BMP-2 was investigated. RUNX2 is a major target of the BMP pathway. In osteoblastic cells, transcription of the bone-specific OCN gene is principally regulated by the RUNX2 transcription factor
19. RUNX2 expression of MSLCs cultured with BMP-2 was higher than that of MSLCs cultured without BMP-2 on day 21. As a result, MSLCs cultured with BMP-2 showed the highest OCN content on day 21. Among the many proteins expressed early in osteogene- sis, Col1A1 and ALP are important early indicators of a cell’s osteogenic capacity.
Song et al.
20demonstrated that BMP-2 treatment had a strong positive effect on the secretion of early indicators, such as ALP, thereby supporting our results.
This suggests that BMP-2 treatment
9 may affect early osteogenic differentia- tion of MSLCs in vitro. OCN has been shown to be a late stage marker of osteoblastic differentiation
21, whereas calcium deposition is a terminal stage marker
22. -glycerophosphate is known to promote calcification of the extracel- lular matrix produced by osteoblasts
23, 24. The present study shows that
-glycerophosphate has strong positive effects on calcium deposition in MSLCs cultured without BMP-2.
In conclusion, BMP-2 had a positive effect on the expression of Col1A1 and ALP and ALP activity, which are early indicators of osteogenic differentiation, in MSLCs. However, culture of cells in medium without BMP-2 promoted calcium deposition, which occurs at the terminal stage of osteogenic differentia- tion. MSCs derived from mouse or human iPSCs have been reported by several groups
25, 26. Moreover, MSCs derived from iPSCs have greater cell proliferation capacity than BMMSCs (over 10-fold higher in telomerase activ- ity) and are able to proliferate for 120 population doublings without losing their self-renewal capacity and MSC charac- teristics
7. Alveolar reconstruction of large bony defects, such as a cleft lip and palate might require scaffold mate- rials. The combination of scaffolds and osteoblast-like cells differentiated from HG-iPSC-derived MSCs holds great potential for patient-specific bone tissue engineering therapy.
ACKNOWLEDGMENTS
This study was supported by MEXT/JSPS KAKENHI Grant Number 17K11770 and the Promotion and Mutual aid Corporation for Private Schools of Japan.
REFERENCES
1)
Weijs W, Siebers T, Kuijpers-Jagtman A, Bergé S, Meijer G, Borstlap W. Early secondary closure of alveolar clefts with mandibular symphyseal bone grafts and
β-tri calcium phosphate (β-TCP). Int J
Oral Maxillofac Surg 2010;39:424-429.
2)
Gimbel M, Ashley RK, Sisodia M, Gabbay JS, Wasson KL, Heller J, Wil- son L, Kawamoto HK, Bradley JP. Re- pair of alveolar cleft defects: Reduced morbidity with bone marrow stem cells in a resorbable matrix. J Craniofac Surg 2007;18:895-901.
3)
Colter DC, Class R, DiGirolamo CM, Prockop DJ. Rapid expansion of recy- cling stem cells in cultures of plas- tic-adherent cells from human bone marrow. Proc Natl Acad Sci U S A 2000;
97:3213-3218.
4)
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S, Marshak DR. Multilineage potential of adult human mesenchymal stem cells.
Science 1999;284:143-147.
5)
Shimada H, Hashimoto Y, Nakada A, Shigeno K, Nakamura T. Accelerated generation of human induced pluripo- tent stem cells with retroviral transduc- tion and chemical inhibitors under physiological hypoxia. Biochem Biophys Res Commun 2012;417:659-664.
6)
Hynes K, Menicanin D, Han J, Marino V, Mrozik K, Gronthos S, Bartold PM.
Mesenchymal stem cells from iPS cells facilitate periodontal regeneration. J Dent Res 2013;92:833-839.
7)
Lian Q, Zhang Y, Zhang J, Zhang HK, Wu X, Zhang Y, Lam FFY, Kang S, Xia JC, Lai WH, Au KW, Chow YY, Siu CW, Lee CN, Tse HF. Functional mesen- chymal stem cells derived from human induced pluripotent stem cells attenuate limb ischemia in mice. Circulation 2010;121:1113-1123.
8)
Raynaud CM, Halabi N, Elliott DA, Pasquier J, Elefanty AG, Stanley EG, Rafii A. Human embryonic stem cell derived mesenchymal progenitors ex- press cardiac markers but do not form contractile cardiomyocytes. PLoS One 2013;8:e54524.
9)
Zou L, Luo Y, Chen M, Wang G, Ding M, Petersen CC, Kang R, Dagnaes- Han- sen F, Zeng Y, Lv N, Ma Q, Le DQ, Besenbacher F, Bolund L, Jensen TG, Kjems J, Pu WT, Bunger C. A simple method for deriving functional MSCs and applied for osteogenesis in 3D scaffolds. Sci Rep 2013;3:2243.
10) Umezaki Y, Hashimoto Y, Nishishita N,
Kawamata S, Baba S. Human Gingival Integration-Free iPSCs; a Source for MSC-Like Cells. Int J Mol Sci 2015;
16: 13633-13648.
11) Egusa H, Okita K, Kayashima H, Yu G,
Fukuyasu S, Saeki M, Matsumoto T, Yamanaka S, Yatani H. Gingival fibro- blasts as a promising source of induced pluripotent stem cells. PLoS One 2010;
5:e12743.
12) Okita K, Matsumura Y, Sato Y, Okada A,
Morizane A, Okamoto S, Hong H, Nakagawa M, Tanabe K, Tezuka K-i. A more efficient method to generate inte- gration-free human iPS cells. Nat Methods 2011;8:409-412.
13) Su R-J, Baylink DJ, Neises A, Kiroyan
JB, Meng X, Payne KJ, Tschudy-Seney B, Duan Y, Appleby N, Kearns-Jonker M.
Efficient generation of integration-free iPS cells from human adult peripheral blood using BCL-XL together with Yamanaka factors. PLoS One 2013;8:
e64496.
14) Peng S, Maihle NJ, Huang Y. Pluripo-
tency factors Lin28 and Oct4 identify a sub-population of stem cell-like cells in ovarian cancer. Oncogene 2010;29:
2153-2159.
15) Kuroda T, Yasuda S, Matsuyama S,
Tano K, Kusakawa S, Sawa Y, Kawamata S, Sato Y. Highly sensitive droplet digital PCR method for detection of residual undifferentiated cells in car- diomyocytes derived from human pluri- potent stem cells. Regenerative Therapy 2015;2:17-23.
16) Aston BA, Allen TD, Howlett CR,
Eagelson CC, Hattori A, Owen M. For- mation of Bone and Cartilage by Marrow Stromal Cells in Diffusion Chambers in Vivo. Clin Orthop Relat Res 1980;151:294-307.
17) Rosen V. BMP2 signaling in bone
development and repair. Cytokine Growth Factor Rev 2009;20:475-480.
18) Sierra J, Villagra A, Paredes R, Cruzat F,
Gutierrez S, Javed A, Arriagada G, Olate J, Imschenetzky M, Van Wijnen AJ. Regulation of the bone-specific osteocalcin gene by p300 requires Runx2/Cbfa1 and the vitamin D3 re- ceptor but not p300 intrinsic histone acetyltransferase activity. Mol Cell Biol 2003;23:3339-3351.
19) Song SJ, Jeon O, Yang HS, Han D-K,
Kim B. Effects of culture conditions on osteogenic differentiation in human mesenchymal stem cells. J Microbiol Biotechnol 2007;17:1113.
20) Zhao J, Shinkai M, Takezawa T, Ohba S,
Chung UI, Nagamune T. Bone regen- eration using collagen type I vitrigel with bone morphogenetic protein-2. J Biosci Bioeng 2009;107:318-323.
21) Lian JB, Stein GS. Concepts of
osteoblast growth and differentiation:
basis for modulation of bone cell development and tissue formation. Crit Rev Oral Biol Med 1992;3:269-305.
22) Hessle L, Johnson KA, Anderson HC,
Narisawa S, Sali A, Goding JW, Terkeltaub R, Millán JL. Tissue- non- specific alkaline phosphatase and plasma cell membrane glycoprotein-1 are central antagonistic regulators of bone mineralization. Proc Natl Acad Sci U S A 2002;99:9445-9449.
23) Poliard A, Lamblin D, Marie PJ,
Buc-Caron MH, Kellermann O. Com- mitment of the teratocarcinoma-derived mesodermal clone C1 towards terminal osteogenic differentiation. J Cell Sci 1993;106:503-511.
24) Teramura T, Onodera Y, Mihara T,
Hosoi Y, Hamanishi C, Fukuda K.
Induction of mesenchymal progenitor cells with chondrogenic property from mouse-induced pluripotent stem cells.
Cellular Reprogramming 2010;12:249- 261.
25) Villa-Diaz LG, Brown SE, Liu Y, Ross
AM, Lahann J, Parent JM, Krebsbach PH. Derivation of mesenchymal stem cells from human induced pluripotent stem cells cultured on synthetic sub- strates. Stem Cells 2012;30:1174-1181.
(Received, November 7, 2017/
Accepted, December 12, 2017)
Corresponding author: