ヌクレオソームの構造とその情報
慶應義塾大学環境情報学部 斎藤輪太郎
(監修)・野崎慎 (著)
平成
23
年2
月7
日Epigenetics
21
世紀の科学は生命科学がリードすることになるだろう.「20世紀の夜明けに,アルベルト・アイ ンシュタインとマックス・プランクが宇宙の基本公式を記述していた頃, 生物学者はイヌがどのよ うにヨダレをたらすかに関する研究でノーベル賞を受賞していた」(Check Hayden, 2010)
という言 葉には,ワトソンとクリックがDNA
の二重らせん構造を発見してから今日まで, どれほどすさま じいはやさで生命科学が進歩してきたかが如実に表れている.そして,2003年のヒトゲノム解読以 降,現代の生命科学はゲノムワイドな解析を取り扱うことが可能となり, さまざまな知見が蓄積さ れている. また, 生命科学の研究の進展とともに, シーケンスやアレイといった技術が急速的に 発展し, 巨大なデータが蓄積され続けている. これらの巨大なデータをどのように統合的に扱い,新しい発見をするかという問題に対し, バイオインフォマティクスと呼ばれる情報解析の技術を用 いて生命を解明しようという学問に注目が集まっている.そして,最近の研究では
DNA
の一次配 列の変化を伴わずに後天的な修飾によって行われる「エピジェネティクス」と呼ばれる一群の制御 機構の存在が明らかとなった.この機構は真核生物における特徴的な制御方法であり,その代表格 としてDNA
のメチル化やヒストンの修飾が挙げられる. これらの修飾の多くは可逆的であり,非 常に柔軟な制御である. また組織の特異性も高く,多細胞生物の形成に大きく関わっていることが 示唆されている. ヒトゲノムの解読が終了した現在において, ゲノム配列だけからは知り得るこ との出来ないこれらのエピジェネティック制御の理解が生命科学研究の使命の一つに挙げられてい る. エピジェネティクスの研究はまだ歴史が浅く, 不明瞭な部分が大半を占めているが, ヒスト ンはDNA
を梱包し,核内に折り畳むだけではなく,そのポジションや修飾によって様々な機能を 持つことが明らかにされている. 特にヒストンの修飾の組み合わせによって様々な特徴的な機能を 引き出すというヒストンコード仮説はStrahl
とAllis
によって2000
年に提唱され, それを支持す るデータが蓄積されている.本冊子では,現在知られているヒストンに関する情報を網羅的に収集し,その総説について記述 した.エピジェネティクス研究はすさまじい速度で進展しているため, 網羅的な総説が存在してい ないことが問題となっている. そこで, エピジェネティクスの初学者はこの冊子を読むことで,も しくは現在世界中で繰り広げられているエピジェネティクス研究を理解できるように構成している.
特に後半においては,これらからの
20
年間でエピジェネティクス研究の軸となる問題について記述 した.ヒストンを取り巻くエピジェネティクス研究はすさまじい早さで進展しつつあり,これからの生 命科学への貢献が期待されるだろう.
2011
年1
月19
日 野崎慎目 次
1
ヒストン4
1.1
ヒストンの構造. . . . 4
1.2
ヌクレオソームの構造. . . . 5
1.3
ヒストンの進化. . . . 6
2
ヒストンコード7 2.1
ヒストン修飾とその機能. . . . 7
2.1.1
ヒストンのメチル化. . . . 9
2.1.2
ヒストンのアセチル化. . . . 11
2.1.3
その他のヒストン修飾. . . . 11
2.2
ヒストン修飾認識ドメイン. . . . 12
2.3
ヒストンクロストーク. . . . 13
3 DNA
メチル化とヒストン修飾15
4 small RNA
とヒストン修飾18
H2B
H4 H3
H2A
図
1:
ヒストンタンパク質の概略図黄色い丸い樽状のタンパク質がヒストンタンパク質を表している. このように
4
つのコアプ ロテインが2
コピーずつ集まりヒストンが形成される.1
ヒストン1.1
ヒストンの構造真核生物のゲノムは核内に折り畳まれている.ヒトゲノムに関しては, 塩基対だけで
2 m
の長さ に達するとされている. このような巨大なゲノムを核内に折り畳むために, クロマチンという繰 り返し構造を形成することが必要となる.ヒストンタンパク質はそのようなクロマチン構造の形成 で,中心的な役割を果たしている.ヒストンはH2A,H2B,H3,H4
という4
つのヒストンコアタ ンパク質が2
コピーずつ集合することでヒストン8
量体として形成される(図 1). そのヒストン 8
量体の周りを約147 bp
のDNA
が, 左巻きのスーパーヘリックスで巻き付くによって, クロマチ ンの構成単位であるヌクレオソームが形成される(Luger et al., 1997; Kornberg and Lorch, 1999)
.そ れぞれのヒストンコアタンパク質は球状のC
末端側とヒストンテールと呼ばれる鎖状のN
末端側 の2
つの特徴的な構造を持つ. それぞれ4
つのヒストンコアはhistone fold domain (HFD)
を共通 の構造ドメインとして持つ. これらは3
つのα-helices (α1,α2,α3)と, それを分断する2
つの ループ構造(L1,L2)
からなる. ヒストン同士はH3
とH4,H2A
とH2B
という逆行性のペアが それぞれ組み合わさることで,複合体を形成する(Arents and Moudrianakis, 1995; Luger et al., 1997;
Zlatanova et al., 2009)
.ヒストンを構成するアミノ酸残基をみるとそれぞれヒトにおいて,H2Aは
130 AA,H2B
は126
AA,H3
は136 AA,H4
は103 AA
からなっている.真核生物でのヒストンコアタンパク質の保存性は高く, どの生物のヒストンもほぼ同じアミノ酸によって構成されており, 少しの変異でも致 死に至ることが示されている. それぞれのヒストンコアには,N末端側から
30
アミノ残基付近ま でに多くの翻訳後修飾可能な残基が存在している.ヒストンにおける翻訳後修飾は細胞内で重要な図
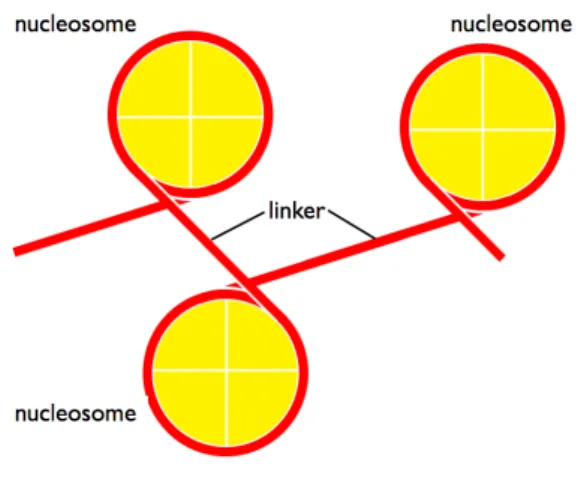
2:
ヌクレオソームとリンカーの概略図ヌクレオソームが周期的に形成され,それぞれのヌクレオソームはヒストンに巻き付かないリ ンカー領域によって分断されている.黄色い丸がヒストン,赤い線が
DNA
を表している.役割を果たすことが知られており, 集中的に研究が行われている1. ヒストンにおける修飾可能な 残基を紹介していく.まず,ヒトにおける
H3
内で現在報告されている修飾可能な残基はR2,K4,
K9, S10,K14,R17,K18,K23,R26,K27,S28,K36,P38,K79
である.左のアルファベッ トが残基の種類(リジン: K, アルギニン: R
,トレオニン: T ,セリン: S, グルタミン酸: E,プロリン: P),右の数字が開始メチオニンを除いた
N
末端側からのアミノ酸残基数を表している.H4
内ではS1,R3,K5,K8,K12,K16,K20
が報告されている.そしてH2A
内では,S1,R3,K5,T119
が,H2B内ではK5,K13,S15,T120
が報告されている(Bairoch et al., 2005)
.ここに 列挙した多くのアミノ酸残基をみても, ヒストンは多くの翻訳後修飾可能な残基を持っていること が伺える. これらの修飾可能なアミノ酸残基が,真核生物の細胞内において様々な機能と関連して おり,ヒストンが単なるDNA
を梱包するための部品ではなく, 多くの制御機構における要として 用いられている.1.2
ヌクレオソームの構造真核生物の染色体
DNA
は, ヌクレオソームとよばれるヒストンにDNA
が巻き付いた領域と,リンカーとよばれるヒストンに巻き付いていない
DNA
がむき出しの領域が繰り返すことでクロマ チン構造を形成している(Kornberg, 1974; Olins and Olins, 1974; Oudet et al., 1975)
.ヌクレオソーム では,ヒストンに約147 bp
のDNA
が1.67
周巻き付いて形成される(Luger et al., 1997).また,
それらヌクレオソームの間に存在するリンカー領域は約
50 bp
のDNA
からなっている(図 2) (Van
Holde, 1989; Yao et al., 1990)
.さらに,大域で染色体の構造を観ていくとクロマチン構造もその凝集度合いから, ユークロマチンとヘテロクロマチンと呼ばれる
2
つの構造タイプに分けることができ1修飾による制御については後述する.
る. ヘテロクロマチンはクロマチン同士が密に凝集している状態を指し, ユークロマチンはクロ マチン同士が凝集していない状態のことを指す. このようなクロマチン構造の変化は転写制御と関 連していることが知られている.
ゲノム上において,約
200 bp
の周期で現れるヌクレオソーム領域であるが,これらのポジション がどのように決まっているかについては様々な議論が行われている. 現在考えられているヌクレオ ソームポジションを決定する因子として,DNA配列選択性,DNAメチル化,ヒストン修飾,ヒス トンコアタンパク質のホモログであるヒストンバリアント,そしてクロマチンの構造に関係するク ロマチンリモデラーなどが挙げられている(Segal and Widom, 2009)
.この中でもDNA
配列選択性 については,盛んに研究されており,DNAとヒストンの親和性に関する研究で,DNA配列の塩基 組成の違いで少なくとも5000
倍の親和性の差があることが示されている.また,DNA配列はその 構造的な特徴から,塩基配列の組み合わせによって物理的に生じる曲がりやすさ(bendability)
や,DNA
のらせん構造の特徴性によって親和性が異なることも示唆されている(Drew and Travers, 1985;
Widom, 1992; Lowary and Widom, 1998; Widom, 2001; Thastrom et al., 2004; Gencheva et al., 2006)
.他にも
in vitro
の研究でin vivo
におけるヌクレオソームポジションを再現することができたり, 染色体
DNA
塩基配列だけから機械学習を用いた手法によりin vivo
におけるヌクレオソームをおおま かに予測できたりすることが可能となっている(Ioshikhes et al., 2006; Segal et al., 2006; Peckham et al., 2007; Yuan and Liu, 2008; Ogawa et al., 2010)
.しかし,DNAのどのような配列特徴がヌクレオ ソームの形成に重要であるかは明らかとされておらず, 今もなお研究が続けられている.1.3
ヒストンの進化真核生物において, クロマチン構造を形成するヒストンは重要なタンパク質であるが, ヒストン タンパク質がどこから派生し, 真核生物に現れたかについては不明瞭なままである. ヒストンタン パク質の保存性解析では,バクテリアでは未だヒストン様タンパク質が発見されていないものの
(最
近,ヒストン様タンパク質(HU)
が発見されたが詳細は不明である(Balandina et al., 2002)),古細
菌においてはヒストンのHFD
とホモログなドメインを持つタンパク質が発見されている(Sandman and Reeve, 2006; Garrett and Klenk, 2007).さらに, これらは古細菌の主な 3
つの門(ユリアー
キオータ門,クレンアーキオータ門,ナノアーキオータ門)で発見されており,真核生物と古細菌 の共通の祖先に存在していたという可能性が示唆されている(Sandman and Reeve, 2006).ヒスト
ン様のタンパク質の保存性に関して, さらに古細菌の中で詳しく調べていくと,HFDを含むタン パク質はユリアーキアでは確認することができるが,クレンアーキアの中ではケナルカエウムを除 くほとんどにおいては確認出来ないことが報告されている.これらの結果からHFD
を含むヒスト ンの祖先タンパク質は,1.
クレンアーキア以降失われた2.
ユリアーキアとクレンアーキアの分化後に生まれた3.
真核生物とユリアーキアの間で水平伝播が起きて, 真核生物から持ち込まれたの
3
つの仮説が考えられている. これらの検証がヒストン誕生に迫るためには必要であるが, 今 のところ古細菌において生じたタンパク質なのか, 真核生物において生じたタンパク質なのかはまだ分かっていない
(Sandman and Reeve, 2006)
. 真核生物におけるヒストンとの比較を行うと,R10,R19,K53
とT54
がDNA
への結合に重要であると予測されているが, その領域に関しては古細菌において高い保存性が示されている
(Decanniere et al., 2000)
.しかし,古細菌におけるヒストン は構造レベルでは真核生物におけるヒストンとよく似通っているが,配列レベルではそうとはいえ ない.さらに, 古細菌におけるヒストンは真核生物におけるヒストンのN
末端側鎖状ペプチドと ホモログな配列を持ち合わせていない.真核生物において, 鎖状のN
末端テールはヒストン修飾 と関連し, 細胞内における様々なエピジェネティクスな制御に必要とされている部位である. そ のような部位を古細菌では持ち合わせていないということは, ヒストンはどの時点においてN
末 端テールを獲得したのかという問題を提起する. また, 古細菌の一部で真核生物においてヒスト ンの脱アセチル化を行うhistone deacetylace (HDAC)
のホモログ遺伝子が発見されており, 古細 菌においても真核生物と同様にヒストン修飾を行う機構があるのではないかということが示唆されている
(Sandman and Reeve, 2006)
.これらの問題は,古細菌のゲノム解読やタンパク質の同定とともに明らかにされ,さらにそれらの機能同定によって, ヒストンとその修飾による制御機構のルー ツを明らかにするだろう.
2
ヒストンコード2.1
ヒストン修飾とその機能真核生物においてヒストンは
DNA
を梱包するためのタンパク質としてだけではなく,ヒストン のN
末端テールにおけるリジン, アルギニンなどにおける修飾によって,様々な細胞内のプロセ スと関連することが知られている. ヒストンで起きた翻訳後修飾から, それと相互作用するタン パク質によって様々な機能が引き出されるという「ヒストンコード仮説」がStrahl
とAllis
によっ て2000
年に提唱された(Strahl and Allis, 2000)
.ヒストンにおける翻訳後修飾の種類として, リン 酸化, メチル化,アセチル化, ユビキチン化,SUMO化,ADP
リボース化,脱アミノ化, ビオ チン化,そしてプロリンの異性化など, 多く修飾が知られている.このように, ヒストン上には 多くの修飾可能なアミノ酸が存在している. 修飾の種類や, 修飾サイトの組み合わせによって現 れる機能はこれまでに数多く同定されており, ヒストンコード仮説を支持している(図 3).
ヒストン修飾によって引き起こされる様々な制御はエピジェネティック制御の中核を担っている.
以前から転写開始点付近におけるヒストンのアセチレーションやメチレーションが転写制御に関わっ ていることは知られていたが, 詳しいことは不明瞭なままであった. しかし,近年におけるクロ マチン免疫沈降
(Chromatin immunoprecipitation: ChIP)
法, アレイ技術, シーケンシング技術 の発達に伴い, ゲノムワイドなヒストンの修飾状態の同定が可能となった. これにより, ヒスト ン修飾に関する様々な知見とデータが蓄えられてきている. ヒストンの修飾と関連する制御として は, 転写(転写開始, 伸張, 終結, スプライシング),DNA
修復,DNA複製, クロマチンの凝 縮などが挙げられる.転写の活性化は主にプロモーターへの転写因子の誘導,もしくは阻害が修飾 されたヒストンを認識するタンパク質が仲介することによって行われる.DNA修復では,DNA損 傷をうけた領域に特異な修飾を受けたヒストンを配置することで, それを認識するタンパク質を仲 介して行われる.DNA
複製とヒストン修飾の関係性については示唆されているものの,まだあま り研究が進んでいない状態である. クロマチンの凝縮に関しても特異的な修飾を受けたヒストンをA P
P
H3
H4
H2A
H2B
ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPH
SGRGKGGKGLGKGGAKRHRKVLRDNIQGIT
SGRGKQGGKARAKAKSRSSRAGLQFPVGRVHRLLRKGNY
PEPAKSAPAPKKGSKKAVTKAQKKDGKKRK
M M M
M
M M M M MM
M M
M M
A A A A
A A
P P
P
A A A
A A A A
A A A
A
図
3:
修飾することができるヒストンテール上のアミノ酸と修飾の種類それぞれのヒストンコアタンパク質において,修飾可能なヒストンテール残基とその修飾例 を示した.左側のアルファベットの並びは
N
末端アミン酸テールを示し,右側の大きな紺色の丸 はC
末端アミノ酸球形ドメインを示している.青い丸のA
はアセチル化,赤い丸のM
はメチル 化, 紫の丸のP
はリン酸化を表している.ここに示すように,それぞれ修飾の組み合わせは多 様である.認識するタンパク質によって行われる.このように多くのヒストン修飾が存在するが,以下修飾と それによって引き出される機能を一つ一つ紹介する.
2.1.1
ヒストンのメチル化ヒストンのメチル化はリジンとアルギニン上で起きる. 現在, ヒストンのメチル化は主に転写 と
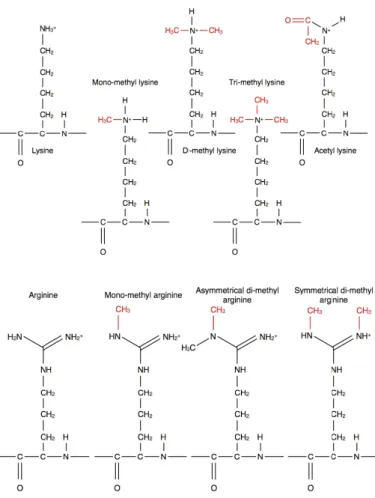
DNA
修復,クロマチンの凝集に関与していることが報告されている.それぞれのアミノ酸上に おいて,リジンは修飾基が1
つの場合のモノメチルリジン(Kme1),2
つの場合のジメチルリジン(Kme2),そして 3
つの場合のトリメチルリジン(Kme3)
と3
つのメチル化パターンが知られている.またアルギニンも修飾基が
1
つの場合のモノメチルアルギニン(Rme1),2
つのメチル基が非対 称な場合の非対称ジメチルアルギニン(Rme2a), そして 2
つのメチル基が対称な場合の対称ジメチ ルアルギニン(Rme2s)
と3
つのメチル化パターンが知られている(図 4). メチル化されるサイト
がヒストン上であっても,メチル基のつき方の違いで引き出される機能が異なる場合がある.リジ ンとアルギニンにおけるメチル化はプロモーターに転写因子を誘導することで転写の活性化に関与 し, またヘテロクロマチン化に関連するタンパク質を誘導することでヘテロクロマチン化を促し,転写の不活性化に関与する.
H3K4me
は転写開始との関係性についてもっとも研究が進められているヒストン修飾の1
つであり,主に転写開始点
(transcriptional start site: TSS)
付近において見られる.H3K4me3はその 中でも特にTSS
直近(-200 -50 bp)
に位置するヒストンにおいてよく観られ, ヌクレオソームの 除去と転写活性化に関与している.H3K4me1と2
も3
と同様に転写開始の活性に関連しているが,H3K4me3
がTSS
直近に頻度のピークを持つのに比べ,TSS から少し離れたところに頻度のピークを持つ.何故
TSS
周辺において,修飾数によってポジションが異なるかについてはまだ明らかに なっていない.H3K27
のメチル化も主にTSS
付近に位置するヒストンに観られる.H3K27me3は転写抑制に関わっていることが示唆されており,
Hox
遺伝子の発現のサイレンシングに関わっていることが知ら れている. 一方, モノメチル化されているH3K27me1
は抑制遺伝子よりも活性遺伝子付近に位置 していることが知られており,H3K27me3と違って転写活性との関係が示唆されている.このように
H3K27me1
と3
のようにヒストン上のポジションは同じであっても修飾するメチル基の個数が異なることでその関係する機能も異なっている例が示されており, これは, ヒストン修飾による 制御の複雑さを表しているといえる.
H3K9
のメチル化はヘテロクロマチンの形成と遺伝子の抑制に関与していることが知られている.H3K9me2
とme3
は抑制遺伝子付近(± 10 kb)
に主に位置する.H3K9me3 はHeterochromatin
protein 1 (HP1)
をプロモーター周辺に誘導し,ヘテロクロマチンを形成する.しかし,一方H3K9me1
は活性遺伝子周辺に位置することが確認されている. これは
H3K27
におけるメチル化と同様, ヒ ストンポジション上での修飾基数による制御の変化を表しているのかもしれない.H3K36me3
は活性遺伝子のTSS
下流の遺伝子内領域に位置することが知られており, 転写伸張に関係していることが示唆されている.そして活性遺伝子の
3’
末端でも見つかっていることから,リン酸化されたセリン残基
(S2ph)
を持つRNA
ポリメラーゼII (RNA polymerase II: Pol II)
との 関係性が示唆されている.またEAF3
タンパク質とRpd35
ジアセチラーゼ複合体を誘導し, 周囲図
4:
ヒストンにおけるリジンとアルギニンの修飾ヒストンのリジンとアルギニンにおける修飾について示した.上図はリジンのメチル化
(mono-
,di-
,tri-methyl)
とアセチル化(ac)
について,下図はアルギニンのメチル化(mono-
,2a-
,2s-
methyl)
について表している.のヒストンにおける脱アセチル化を行うことで, コード領域内部における不必要な転写開始を抑制 する働きがある
(Carrozza et al. 2005
;Cuthbert et al. 2004; Joshi and Struhl, 2005; Keogh et al., 2005)
.H3K79me3
は抑制プロモーターよりも活性プロモーター付近に位置することが示されている.H3R2のメチル化は核内受容体を仲介した転写を共活性化する
coactivator-associated arginine methyltrans-
ferase 1 (CARM1)
によって行われる. これにより,H3R2のメチル化が転写活性と関連することが示唆されているが,H3R2me1と
me2
共に活性遺伝子との関係性は見つかっていない. 以前の研究では
H4K20
のメチル化は抑制クロマチンの形成と関連していることが示唆されていた. しかし,最近の研究では
H4K20me1
とme3
はゲノム上での分布が異なり,H4K20me3がヘテロクロマチン に関連している一方で,me1
は活性遺伝子のプロモーターやコーディング領域に位置していること が多いことが報告されている. またDNA
修復にも関連していることが知られており,H4K20me
と 細胞周期チェックポイントタンパク質であるCrb2
が共役してDNA
修復が引き起こされる(Sanders et al., 2004)
. 共役するタンパク質であるCrb2
は,double tudor domainを通してH4K20me
を認 識してDNA
修復部位に誘導される(Botuyan et al., 2006)
. またCrb2
はDNA
修復部位周辺にお いて, リン酸化が誘導されたヒストンバリアントであるH2A.Xph
に対してBCRT
ドメインを通 して認識し,H4K20meとH2A.Xph
の2
つに結合することでその部位に安定的に留まる.ヒトにおいては
p53BP
1によって行われる.2.1.2
ヒストンのアセチル化ヒストンのアセチル化は転写,DNA修復,DNA複製といった様々な機能と関連している. メチ ル化と違い, アミノ酸に対して
1
つの修飾基を付けることしか出来ず, リジンのアセチル化(Kac)
しか存在しない. ヒストンのアセチル基はアセチル化を触媒するヒストンアセチルトランスフェラー ゼ(histone acetyltransferase: HAT)
と脱アセチル化を触媒するヒストンジアセチラーゼ(histone
deacetylase: HDAC)
といった2
つのタンパク質によって可逆的に行われている. 転写制御においてはメチル化による制御と違い, 今のところ転写抑制に関係する制御はなく,転写活性に関係する 修飾しか見つかっていない.H3K9ac,H4K12ac,
H2BK5ac,H4K16ac,H3K14ac
は転写活性と 関係することが知られている.ヒストンのアセチル化はDNA
修復との関係性についても研究が進 められている.G2 期において,DNA損傷がない場合はH3K56ac
が観られないが,DNA損傷が ある場合はH3,H4
のジアセチラーゼが抑制され,H3K56ac が多量存在することが報告されてい る(Celic et al., 2006;Maas et al., 2006)
.2.1.3
その他のヒストン修飾ヒストン修飾の種類として, メチル化とアセチル化の他にもリン酸化, ユビキチン化,
SUMO
化,ADPリボース化, 脱アミノ化,そしてプロリンの異性化が挙げられる. リン酸化はヒストン 中のセリンとトレオニンに起きる.H3S10におけるリン酸化はH3S10ph
を認識する他のヒストン 修飾タンパク質の足場となり, 転写,DNA修復, クロマチンの構造に影響を与える.哺乳類にお けるヒストンバリアントであるH2A.X
はダメージを受けたDNA
周辺(数千 kb)
に広く配置される ことでATP
依存性リモデリング活性を持つINO80
複合体を誘導し,DNA修復を行うことができ る. またH2AS129ph
とH4S1ph
はそれぞれMec1, Casein kinase II
というタンパク質を仲介し,非相同末端再結合を通して,二本鎖切断修復によって
DNA
修復を行う. ユビキチン化はH2A
とH2B
中のリジンに起こる大きな修飾である. リン酸化同様に他の修飾の足場になることが知られ ており, 転写,DNA修復に関与することが知られている.uH2AK119 (H2AK119がユビキチン化 を受けている状態を指す)は, ポリコーム群タンパク質(Polycomb group protein: PcG)
内に見つ かっているBmi/Ring1A
タンパク質を仲介して転写抑制に関与している(Wang et al.,2006). 逆に uH2BK120
はヒトにおいてはRNF20/RNF40
とUbcH6
を仲介し,出芽酵母ではRad6/Bre1
を仲 介して転写の活性化に関与している(Zhu et al.,2005).しかし,多くのメカニズムや機能性は不明 であり, クロマチンに追加因子を誘導することで, もしくはその大きさから物理的にクロマチン をオープンにしておくことによって制御と関係していると考えられている.他の2
つの修飾につい てはあまり研究が進んでいないが,SUMO化はヒストン中のリジンに付加可能であり,H4,H2A,H2B
において修飾を受ける残基が見つかっている.ユビキチン化と同様, 大きな修飾である. ア セチル化とユビキチン化と競合し, その結果転写抑制に関連しているとされている(Nathan et al.,
2006).ADP
リボース化はグルタミン酸に付加可能である. 今のところ,H2BE2ar1においてのみ見つかっている.
ADP
リボース化はmono-か poly-で負荷することが可能であり, それぞれ MARTs (MonoADP-ribosyltransferase)
,PARPs (Poly-ADP-ribosepolymerase)
という酵素によって行わ れている.転写制御と関係しているとされているが,そのメカニズムや詳しい機能性は分かってい ない(Hassa et al. 2006).脱アミノ化はPeptidyl arginine deiminase type IV (PADI4)
酵素によっ てH3
とH4
におけるアルギニンがシトルリン(Cit)
へと変化する修飾である.メチル化可能なアル ギニンをシトルリンに変化させることによってメチル基の付加を阻害することができ, アルギニン のメチル化による転写制御に影響を与えると考えられている. またin vitro
におけるPADI
4 酵 素の研究により,RmeとCit
は可逆的であることが示唆されている(Hidaka et al., 2005). プロリ
ンはcis
型とtrans
型の2
つの構造異性体を持ち, プロリンの異性化はP-cis
からP-trans
へと異 性化する修飾である. 構造が変化することでポリペプチドの構造が変化し, 他の部位への修飾を 阻害する.現在H3P38
におけるプロリンの異性化が見つかっており,これはH3K36
へのSet2
に よるメチル化を阻害し,転写制御に関係する(Chen et al. 2006).しかし,他の修飾同様, その詳
しいメカニズムや他の機能性については知られていない.これからの研究で, ここに列挙した以外 のヒストン修飾, もしくは列挙したけれども未だ知られていないヒストンの機能が明らかとなって いくだろう.2.2
ヒストン修飾認識ドメインヒストン修飾はそれだけでは機能を持たず, 修飾部位を認識して結合するドメインを持ち, 機能 を仲介するタンパク質が重要となる. 既に多くのドメインが発見されており, それらは真核生物で は普遍的に存在していることが知られている. これからドメインの種類と認識するヒストン修飾部 位について紹介していく.
bromodomain
は約110
アミノ酸残基からなるドメインである.多くの クロマチン関連タンパク質で発見されており, リン酸化されたリジンと相互作用することが知られ ている. クロマチンのリモデリングや転写の活性化に関連するヒストンのアセチル化を担うHAT
と共役するco-activator
のほぼ全て(例: p300
やCBP-associated factor)
においてbromodomain
が見つかっている.chromo (CHRomatin Organization Modifier) domainはハエで見つかった約60
アミノ酸残基からなるドメインである. このドメインを持つタンパク質の多くはクロマチンの構造 を凝集させることでヘテロクロマチン化に関連している
(Zeng and Zhou, 2002). chromodomain
を持つタンパク質は3
つのグループに分けることができる.1つ目はN
末端にchromodomain
を持 ち,さらにchromoshadow domain
と呼ばれる領域を持つタンパク質である. 例としてショウジョウ バエにおけるSu(var)205,HP1
などが挙げられる.2つ目は1
つのchromodomain
を持つもので例 としてDrosophila protein Polycomb (Pc)
が挙げられる.3つ目はタンデムなchromodomain
を持 つもので, 例として哺乳類におけるChromodomain-helicase-DNA-binding protein1-4 (CHD1-4)
が挙げられる.chromodomainを含むタンパク質はメチル化されたヒストン(H3K9me)
に結合する ことができる(Kornberg and Lorch, 1999; Wyrick et al., 1999; Bannister et al., 2001; Jacobs and Khorasanizadeh, 2002; Min et al., 2003; Tajul-Arifin et al., 2003; Joshi and Struhl, 2005).PHD (Plant Homeo Domain) finger
はシロイヌナズナのホメオドメインタンパク質であるHAT3
におい て発見されたCys4-His-Cys3
モチーフ配列からなるドメインである.PHD fingerは約50 ?80
アミ ノ酸残基からなり,ヒトにおいては100
以上のプロテインがこのドメインを持つことが分かってい る(例: p300,CBP,Polycomb-like protein).bromodomain
やchromodomain
などの他のドメイ ンと共役して働くことが多い.ING2,YNG1とNUR
などのいくつかのタンパク質におけるPHD finger
はH3K4me3
に結合することが報告されている(Pascual et al., 2000; Bienz, 2006; Shi et al., 2006; Taverna et al., 2006).また SMCX
というPHD finger
を持つタンパク質がH3K9me3
に結合 することが報告されていることから,PHD fingerはメチル化されたリジンに結合できるのではない かということが示唆されている.Tudor domainはメチル化されているヒストンに結合する(Huang et al., 2006). ヒストンジメチラーゼを含む JMJD2A
のTudor domain
がH3K4me
とH4K20me
に結合することが報告されている.Malignant brain tumor (MBT) repeatsはH3
とH4
における メチル化したリジンに結合するモジュールである.MBT repeatsは約100
アミノ酸残基からなり,MBT
を持つCGI-72
はH3K4me1
とH4K20me1
に結合することが報告されている(Eryilmaz et
al., 2009).Trithorax (TRX)
ヒストンメチルトランスフェラーゼ複合体のサブユニットで保存されている
WDR5
が持つWD-40 repeat
はH3K4me0-3
に結合することが知られている(Couture et
al., 2006).14-3-3
タンパク質はH3S10ph
に結合することが示されており, リン酸化トレオニンとリン酸化セリンに結合することが可能であることが示唆されている
(Macdonald et al., 2005; Winter et al., 2008; Karam et al., 2010). このように多くのヒストン認識結合ドメインとその結合サイト
が知られており, これらのドメインとヒストン修飾が相互作用することで初めてヒストンコードが 可読なものとなる. まだ知られていないヒストン認識結合ドメインの解明や, 現在知られていて もまだわかっていない修飾部位への結合の研究がヒストンコード仮説解明のためには重要である.2.3
ヒストンクロストークヒストンの修飾間での修飾クロストークは様々なヒストンコードを構成すると考えられている.
ここで述べるクロストークは, ヒストン上における修飾がヒストン上の他の部位の修飾に影響を与 える場合を指す. いくつかの場合で,ある
1
つの残基における修飾が他の残基に対する修飾能力を 変えることが知られている(Lee and Zhang, 2008; Lee et al., 2010). よく知られている例を見てい
くと,H3S10phがヒストンアセチルトランスフェラーゼであるGcn5
を促し,H3K14のアセチル化(H3K14ac)
が行われるというヒストンクロストークを挙げることができる(Cheung et al., 2000; Lo et al., 2000).Gcn5
にはヒストン修飾認識ドメインであるbromodomain
が存在しているので,こ れを仲介してH3S10ph
に結合し,H3K14
をアセチル化するものと考えられる.またH3K14
の修飾 状態を除去して, 周囲の修飾状態を調査する研究により,H3K14における修飾の除去はH3K4me3
が特異的に失われるも,H3K4me1
と2
は失われないことが示された(Nakanishi et al., 2008).こ
れらの結果より,H3K4のメチル化はその修飾数によって異なる経路を持っているということが示 唆されている. このように,付加するメチル基数によって経路が異なっているということは大変興 味深く,ヒストンコードをより一層複雑なものとしている. ヒストン修飾酵素はよくマルチサブユ ニット複合体で発見され, それらのサブユニットはヒストンを修飾する部位とヒストン修飾を認識 して結合する部位に分けられることが多い.NuAヒストンアセチル化酵素複合体のサブユニットのYng1
におけるPHD finger
は,メチル化されたH3K4
を認識してH3K14
のアセチル化のためにアセチル化酵素を誘導する
(Martin et al., 2006; Taverna et al., 2006). また,Yng1
と関連してい るING2
もメチル化されているH3K4
に結合することができる.ING2
はヒストン脱アセチル化酵 素内に存在することが知られており, メチル化されたH3K4
は様々な修飾, もしくは脱修飾のた めの足場になっていることが分かる(Shi et al., 2006). クロストークは修飾, もしくは脱修飾を
誘導するだけではなく, あるアミノ酸残基における修飾がその周辺のアミノ酸残基への酵素による 認識を阻害する例も知られている. 例えばH3R2
にメチル化が起こると酵母ではSet1/COMPASS
に,哺乳類ではCOMPASS-like
複合体によって行われるH3K4
のメチル化を阻害することが報告 されている(Guccione et al., 2007; Kirmizis et al., 2007). このように様々な方法によってヒスト
ン上における修飾同士が相互作用する. また, ヒストン修飾は酵素以外の因子の誘導も阻害する ことが知られている. 例えばHP1
はヘテロクロマチンに関連するタンパク質であり,メチル化さ れているH3K9
に結合することができるが,付近のH3S10
がリン酸化されていると結合できない ことが示されている(Fischle et al., 2005; Mateescu et al., 2008). そして, 同一なヒストンコア
タンパク質におけるアミノ酸残基の修飾間クロストークだけではなく, 別のヒストンコアタンパク 質上における修飾とクロストークを行うtrans-histone
クロストークが存在する例が示されている.FOSL1
の転写調節においてH3
のリン酸化がRNA Pol II positive transcription elongation factor
(P-TEFb)
に必要なH4
のアセチル化を誘導することが示されている(Zippo 2009).この制御に関
するキナーゼ活性を持つ
PIM1
はH3S10
をリン酸化することが知られているが,H3S10に関わる キナーゼとして他にもMSK1
とMKS2
が知られている.最近の研究で,PIM1によるリン酸化とMSK1/2
によるリン酸化では, タイミングとその領域が異なることが示されている.MSK1/2はFOSL1
のプロモーター付近において,H3S10のリン酸化を仲介するが,PIM1はH3S10
のエンハンサー付近において,MSK1/2によるプロモーター付近の
H3S10
のリン酸化後に行われることが 示された. またFOS1
のエンハンサーにおけるH4K16ac
はH3S10ph
を受けるヒストンと一致し ており,RNA interference (RNAi) によってPIM1
をノックダウンすることで,H4K16 のアセチ ル化も失われることから,H3S10とH4K16
がtrans-histone
クロストークを行っていることが示唆 されている.14-3-3 (γ,ε,ζ)タンパク質は上記したがH3S10ph
を認識して結合し,H4K16のア セチルトランスフェラーゼであるMOF
を誘導することが知られている.14-3-3
ε,ζ は,FOSL1 のエンハンサーとプロモーター両方において確認できる(Karam et al., 2010). しかし,14-3-3
γ だけは, エンハンサーにおいてのみMOF
を誘導し,プロモーターには誘導しないことが報告されている.MOFを誘導することによって起こる
H4K16ac
を持つヒストンは, リン酸化されたリジ ンに結合することのできるbromodomain
を持つbromodomain-containing protein 4 (Brd4)
によっ て結合することが可能となる.それに続き,Brd4と関連してP-TEFb
が誘導され,Pol IIを転写 伸張促進のためにPol II
のリン酸化を行う.これら一連の結果より,Zippoらはtrans-histone
クロ ストークが転写制御に関与していることを明らかにした(Zippo et al., 2009). 他にも,H3K4me,
H3K79me
とH2B
ユビキチン化の関係を挙げることができる.最近ではuH2B
が直接H3K79me
に関与していることが示されており
(McGinty et al., 2008),uH2B
がH3K4
メチル化複合体であるCOMPASS
を誘導し,H3K4のメチル化を起こすというクロストークも知られている(Shilatifard,
2006).これは酵母で見つかった現象であるが,最近の研究では哺乳類で広く使われているクロス
トークであることが示唆されている(Kim et al., 2009). またこれらの H3K4me,H3K79me
,そ してuH2B
は活性化されている遺伝子付近に集中していることも示されている(Martin and Zhang, 2005; Minsky et al., 2008). そして, 同ヒストンにおけるテール間の修飾のクロストークだけでは
なく, 周辺のヒストンとのクロストークも興味深い.uH2Bが周辺の別ヒストンの修飾に影響を与 えるかという実験では,uH2B
を持つヒストンでしかH3K79me
は確認されず,別ヒストンへの修 飾影響はないことが示唆された.このことからuH2B
に結合するタンパク質はそのヒストンにおい てのみH3K79
をメチル化することができると考えられる(van Leeuwen et al., 2002; Henry et al., 2003; Klose and Zhang, 2007). ヒストンクロストークの解析は複雑なヒストン修飾ネットワーク
を解き明かすために必要であり, ここで記述したように様々なヒストン修飾間でのクロストークが 既に示されている. しかしこれらは知られている修飾の一部の関係性でしかない.1つ目の問題と して, ヒストンは4
つのコアタンパク質が2
コピーずつ集合することで形成されているが,それぞ れ同一ヒストン内に存在する2
つのH3
やH4
における修飾がどのように相互作用するかというこ とはまだ示されていない.2つ目の問題としてuH2B
の研究で少し行われたが, 隣り合うヒストン 同士でどのようなクロストークを行うかということも示されていない.3つ目の問題として, まだ 時系列データが蓄積されておらず実際のインタラクションの同定が難しいことが挙げられる.これ らの1
つ1
つの解析が複雑なヒストン修飾ネットワークを解き明かす鍵となるであろう.3 DNA
メチル化とヒストン修飾DNA
メチル化とヒストン修飾は真核生物においてエピジェネティック制御を担う2
つの核であ る.DNAメチル化は原核生物, 真核生物を問わず見られる現象であり,進化的にも古くから存在 することが知られている.DNAメチル化は原核生物において制限酵素による切断から守るための 自己と非自己の認識に使われており, 転写制御との関係性は知られていない. 一方真核生物におい ては, ヘテロクロマチン化などに関連し, 転写やトランスポゾンの抑制などに関与していること が知られており, ヒストン修飾と同様に組織特異的なパターンを示し,分化や組織特的な制御に重 要であることが知られている. これら原核生物が持つDNA
メチル化酵素と真核生物が持つDNA
メチル化酵素はホモログであることが知られている. また, 真核生物では全ての種においてDNA
メチル化酵素が保存されている訳ではない. 酵母や線虫などのモデル生物はこれらのメチル化およ びメチル化酵素を持ち合わせていないことが知られており,進化の上で欠失したと考えられている.DNA
メチル化酵素が認識し, メチル化されるDNA
配列は様々であるが,動物では一般的にCpG
という
di-nucleotide
におけるシトシンが修飾されることが知られている.シトシンはメチル化されて
5-メチルシトシンになった後に脱アミノ化されるとチミンになってしまうため,DNA
の修復が困難になる.そのため
CpG
のdi-nucleotide
は真核生物における進化の上で減少していることが 知られており,CpGが多量存在する領域はCpG island
と呼ばれている.CpG island はプロモー ターの上流などに見られることが知られている. 最近ではCpG
のみならずCHG
やCHH
のシト シンがメチル化されていることが明らかとなった. また植物ではシトシン以外にアデニンのメチル 化が知られている. 一方ヒストンは上記したように, バクテリアには存在せず, 古細菌もしくは 真核生物以降で獲得されたと考えられている. このことからも古くから存在するDNA
メチル化と ヒストン修飾は違う経路を辿って進化してきたことが伺える. しかし,最近となってこの2
つの エピジェネティクスな修飾が高等生物においては,お互い影響を及ぼし合っている例が報告されて いる(Cedar and Bergman, 2009). ヒストン周辺の DNA
メチル化を誘導することが知られている 修飾としてH3K9me
とH3K27me
が挙げられる. また, 逆にDNA
メチル化を阻害することが知 られている修飾としてH3K4me
やアセチル化されたヒストン全般が挙げられる. 最近の研究ではDNA
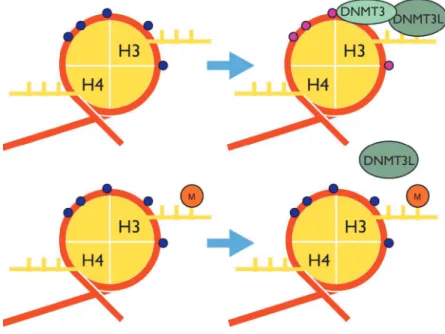
メチル化のプロファイルがヒストン修飾を仲介して決められているというモデルが強く支持さ れている(Cedar and Bergman, 2009).このモデルでは,DNA
メチル化のパターンはde novo
なDNA
メチル化が起こる前にヒストン修飾によって決定されていることを示唆している(Ooi et al., 2007)
.de novo
なDNA
メチル化はDNA methyltranseferase (DNMT) 3A
とDNMT3B
の複合 体とDNMT3L
によって行われることが知られている(Ooi et al., 2007).DNMT3L
はメチル基付 加の活性を失っているが,ヒストンH3K4
に結合して,DNAメチルトランスフェラーゼをDNA
に 誘導し,DNAメチル化の起点になることが知られている.しかし,H3K4にメチル化を受けたヒ ストンはDNMT3L
のH3
への結合を阻害することができる(Ooi et al., 2007). この DNMT3L
の 結合を阻害するH3K4
のメチルトランスフェラーゼは,DNAに直接結合したPol II
に誘導される(Ooi et al., 2007). これらより転写が活性化している状態では, プロモーター周辺において DNA
のメチル化を行うことができないことが示唆されている
(図 5). 他にもヒストン H3K9
のメチルト ランスフェラーゼであるG9a
とヒストン脱アセチル化活性を持つ酵素の複合体によるde novo
なDNA
メチル化の誘導が知られている.この複合体は局所的なヒストンの脱アセチル化を行い, 転 写を抑制することが知られている. 同じ残基上において, メチル化とアセチル化を同時に行うこと はできないが, ヒストンの脱アセチル化によりヒストン上の修飾をリセットすることで,G9a
によ るH3K9
のメチル化を可能にする.H3K9meを持つヒストンはchromodomain
によって結合する ことが可能であり, そのドメインを持ちヘテロクロマチン化形成を促進させることが知られているHP1
のヒストンへの結合が可能となる.最終的にG9a
を含んだ複合体がDNMT3A
とDNMT3B
を誘導し,de novoなDNA
メチル化を行う(Feldman et al., 2006; Epsztejn-Litman et al., 2008).
さらに,動原体周囲のサテライトリピートにおけるヘテロクロマチン化においても, このような ヒストン修飾と
DNA
メチル化の関係性が知られている.これらのサテライト配列ではSET domain
を含むヒストンメチルトランスフェラーゼであるSUV39H1
とSUV39H2
がH3K9
のトリメチル化 を行い,ヘテロクロマチン化に関与する.SET domainとは高等生物において保存されているおよ そ150
アミノ酸残基からなるモチーフ配列であり, クロマチン構造の調節に関係し, それを通し て転写の活性化, もしくは抑制化に関与することが示唆されている(Dillon et al., 2005).これら
のタンパク質はサテライト配列内のCpG
をメチル化するためにDNMT3A
とDNMT3B
を誘導す図
5: DNA
メチル化とヒストン修飾の関係性DNA
メチル化とヒストン修飾の関係性の例を示した.赤の線は染色体
DNA
を表し,その上の小 さな丸はそれぞれ,修飾可能な塩基を表している.上図では,DNMT3L
がヒストンに結合し,DNMT3
を誘導してDNA
のメチル化を行っている.下図ではヒストンにメチル化が起きており,DNMT3L
が結合できず,DNA
のメチル化が阻害されていることを表している.る
(Fuks et al., 2003; Lehnertz et al., 2003).このヘテロクロマチン化のプロセスは RNA
の二本 鎖構造を認識するDicer
を仲介して開始されている.RNA-induced silencing complex (RISC) がSUV39H1
とSUV39H2
を誘導すると思われる動原体周辺の領域を標的にしていることが報告されている
(Fukagawa et al., 2004; Kanellopoulou et al., 2005; Sugiyama et al., 2005). ヒストン修飾
がどのようにDNA
メチル化に影響を及ぼすかということを見てきたが,これからDNA
メチル化 がヒストン修飾パターンの維持にどのような働きをしているかということについて見ていく.DNA 複製の際, メチル化されていたCpG
は複製後, 娘鎖がメチル化されていない状態になる. これ をヘミメチル化状態という. これは複製複合タンパク質と関連して働くDNMT1
によって娘鎖も メチル化されることで,両鎖がメチル化されている状態が維持される(Leonhardt et al., 1992). 最
近の研究で,DNMT1はSRA domain
というヘミメチル化状態のCpG
に結合するドメインを持つUHRF1
とともに複製時にヘミメチル化DNA
となっているCpG
を探し,メチル化状態を維持することが示されている
(Bostick et al., 2007; Sharif et al., 2007; Achour et al., 2008).一方,ヒス
トンはDNA
複製の際,複製フォークによってクロマチン構造が崩されるため,そのあとでもう一 度再構築する必要がある. このときDNA
メチル化による目印がその再構成の際に役立つことが知 られている.DNA メチル化とヒストン修飾の関係はMECP2
やMBD2
といったメチルシトシン 結合タンパク質によってまず認識され,これらはDNA
メチル化領域にヒストンアセチルトランス フェラーゼを誘導する(Jones et al., 1998; Nan et al., 1998). また DNA
メチル化は直接H3K9
の ジメチル化と関係していることが報告されており,G9aと複製複合体に付随しているDNMT1
が相 互作用することにより,DNAメチル化が起きている領域で同時にH3K9
のジメチル化を起こすこ とができることが報告されている(Esteve et al., 2006).他にも DNA
メチル化がH3K4
のメチル 化を阻害することが示されており(Lande-Diner et al., 2007), 植物の DNA
メチル化が起きている 領域では,H2A.Z がヌクレオソームから除去されることが示されている(Zilberman et al., 2008).
H3K4me
もH2A.Z
も転写活性に関係する事が知られており, これらからもDNA
メチル化が転写抑制に関係することが示唆されている.
DNA
メチル化とヒストン修飾の関係性については高等生 物における複雑なエピジェネティクスな制御を明らかにするためにも, 今後ますます力が入れられ る研究だろう. 現在は修飾の開始における関係性に過ぎないが, このような組み合わせを複雑な ヒストン修飾のネットワークに組み込んだときにどうなるかということは非常に興味深い. また,原核生物から存在する
DNA
メチル化と真核生物において獲得されたヒストン修飾がどのように邂 逅し, お互い相互作用するようになったのか,またそれによってどのような制御が可能になったの かということも興味深い問題である.4 small RNA
とヒストン修飾真核生物では多様な
non-codig RNA
の発見が続いているが,その代表例としてRNAi
に関連す るshort interfering RNAs (siRNAs)
やmicroRNAs (miRNAs)
などのsmall RNA
が挙げられる.これらの