Amphibienlarven
Von
Tatsuo Fukai
(Aus dem Anatomischen Institut der Kaiserlichen Universität zu Kyoto :
Prof. Dr. Ch. 0 ga w a) Mit 70 Textfiguren. Inhalt. belle Vorwort ...632
I. Eliminationsversuche mit den Blutgefässen . . . ... 633
Einleitung ...633
Eigene Versuche ... . . ... 633
A. Teilelimination der Schwanzgefässe in etwas späterem Stadium. 634 1. Normales Verhalten der Schwanzgefässe ...634
2. Einfache Elimination der V. caudalis... 634
3. Elimination der V. caudalis an zwei Stellen ...637
4. Elimination der A. caudalis ...638
5. Elimination der Vasa caudalia bei der Metamorphose . . 640
B. Teilelimination der Vasa caudalia in möglichst frühem Stadium. 640 C. Totale Elimination der Vasa caudalia...642
D. Teilelimination der Rumpfgefässe...644
1. Einseitige Elimination der V. jugularis externa. . . . . 644
2. Beiderseitige Elimination der Vv. jugulares externae . . 646
3. Elimination der A. carotis externa ...647
Diskussion ...649
A. Über die Wiederherstellungsformen der Blutgefässe . . . . 649
.B. Über die Wiederherstellungsprozesse der Blutgefässe . . . . 651
Zusammenfassung ...654
II. Transplantationsversuchemitden Organanlagen...657
Einleitung...657
632 Tatsud Fukai,
A. Hornoioplastiscbe Transplantation der Schwanzknospe . . 657 1. Die normale Entwicklung der Schwanzgefässe ...658
2. Transplantation in den Seitenbauch... . 659 3. Transplantation in die Kiemen gegend ...669
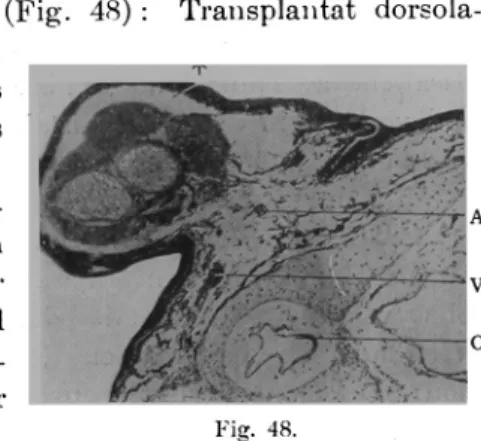
4. Transplantation in die Schwanzknospe...676
B. Autoplastisch dorsoventral umgekehrte Transplantation der Schwanzknospe in loco...678
C. Xenoplastische Transplantation der Schwanzknospe . . . . 680 D. Hornoioplastische Transplantation der Hinterbeinanlage...683
1. Transplantation in die Ohrgegend ... . . . . .683 2. Transplantation in den Seitenbauch... . . . .687
3. Transplantation in die Schwanzknospe...691
Diskussion ...694
A. Über die Genese der Blutgefässe im Transplantat ...694
B. Über die VerbindungsartderBlutgefässezwischenWirtund
Transplantat...698
C. Über das Verhalten der Blutgefässe im Transplantat . . . . 700 D. Über den Blutkreislaufim'Transplantat, ...702
Zusammenfassung... . . .703 III. VariationsversuchemitdenBlutgefässen. .... . . . .707 Einleitung... . ...707 Eigene Versuche...707 . A. Einige Schwanzgefässvarietäten ...707
B. Die experimentell erzeugte „äussere Schwanzvene" bei folarven ...708
C. Verhalten der BlutgefässeimregenerieeenSchwanzvon
Amphibienlarven...710
D. Die Variation der Blutgefässe, entwicklungsmechanisch be- trachtet ...710
Zusammenfassung ...713
Literatur. . . . • . . . . . 714
Vorwort.
Experimentelle Arbeiten bezüglich der Transplantation der
Blutge-fässe oder Herstellung der Kollateralbahnen usw. sind schon mehrfach
veröffentlicht worden. Die betreffenden Versuche wurden jedoch
haupt-sächlich an ausgewachsenen Exemplaren der höheren Wirbeltierklasseii
vorgenommen. Und die Untersüchungen wieder, die am Etnbryo,
insbe-sondere in möglichst frühem Entwicklungsstadium, gemacht wurden,
scheinen mir, was die Entwicklungsmechanik der Blutgefässe anbetrifft,
noch nicht allen wissenschaftlichen Ansprüchen zu genügen.
Darum hat Prof. Dr. Ch. Ogawa mir vorgeschlagen, die
Gefässbil-dung experimentell zu studieren. Dieses Thema zielt darauf, die Art
und Weise der Potenzäusserungen der embryonalen Blutgefässe bei
Entwick-lungsvermögen eines Ei- oder Embryoteiles ist um so grösser, je früher
das Entwicklungsstadium ist, und in der Regel äussert sich bei normaler
Entwicklung niemals das gesamte Vermögen, sondern dies bleibt bis zu
einem bestimmten Grade stets latent. Demnach können wir sagen, der
Zweck meiner Untersuchungen ist der, das latente Entwicklungsvermögen
der embryonalen Blutgefässe künstlich ans Licht zu stellen.
Somit erzwang ich durch verschiedenartige Eingriffe, die den noch
in der Entwicklung befindlichen Blutgefässen zugefügt wurden,
Varia-tionen im Wachstumsverlauf derselben, um diese von verschiedenen
Gesichtspunkten aus zu betrachten. Als Material verwandte ich
Am-phibienlarven, weil bei diesen der gesamte Entwicklungsprozess der
Blutgefässe ziemlich leicht zu beobachten ist. Die Experimente währten
von 1933 bis 1935.
I. Eliminationsversuche mit den Blutgefässen.
Einleitung.
Es ist eine schon allgemein bekannte Tatsache, dass bei erwachsenen
Wirbeltieren eine Unterbrechung des Blutkreislaufes meistens durch die
Kollateralbahnbildung wiederhergestellt werden kann. Über ebendiese
Verhältnisse wissen wir jedoch nichts beim Embryo. Durch die folgen
den Experimente soll darum der Wiederherstellungsprozess der
- fässe nach ihrer Elimination beim Embryo geklärt werden.
Findet bei diesem die Regeneration auch in der gleichen Weise wie beim Erwachsenen oder andernfalls z. B. durch eine Umgestaltung der Kapillaren zur Hauptbahn oder eine Neubildung der Blutgefässe u.a. statt? Beim Experiment habe ich besonders Differenzen im Entwick-lungsstadium, die Arten der Blutgefässe, das topische Verhalten derselben und den Umfang der Elimination berücksichtigt.
Eigene Versuche.
Die Experimente gingen wie folgt vor sich. Als Material benutzte ich hier Bufo- und Rhacophoruslarven. Die Operation wurde unter
Binokularlupe mit Hilfe einer scharfen Nadel ausgeführt. Die operierten Larven wurden in mit Leitungswasser gefüllten Glasschalen gezüchtet. Sie sind teils lebend, teils als Schnittpräparat unter dem Mikroskop beobachtet worden. Genauere Angaben bringen die einzelnen Kapitel.
634 A.
Tatsuo Fukai,
Teilelimination der Schwanzgefässe in etwas späterem Stadium.
1. Normales Verhalten der Schwanzgefässe.
Der Schwanz von Amphibienlarven besteht aus zwei Teilen, näm- lich aus der Muskelschichtregion, die die Breite der Muskelschicht ein- nimmt und zwischen den beiderseitigen Muskelschichten Chorda und Rückenmark enthält, und den beiden Flossen (dorsal und ventral) , die durchsichtig sind.
Der Schwanz wird durch Vasa caudalia versorgt. A. caudalis ver-läuft den unteren Rand der Chorda entlang und V. caudalis zwischen den beiden unteren Rändern der Muskelplatte fast parallel mit A . caudalis. Diese Gefässe senden unterwegs in bestimmtem Abstande zahlreiche Seitenäste in beide Flossen. A. caudalis ist eine Fortsetzung der Aorta dorsalis; V. caudalis geht direkt in V . cardinalis post. über. Dieses Verhalten gilt für die Larven aller Ainphibienarten (Bufo vulgaris , Rana temporaria, Rana nigrotnaculata und Rhacophorus Schlegelii arborea).
Ausser diesen Hauptgefässen ist bei der Rana- und Rhacophoruslarve eine dic'zere Nebenvene d.h. „äussere Schwanzvene" vorhanden . Diese
liegt, und zwar je eine zu
Fig. 1.
Schema der Schwanzgefässe von Racophorus-larve. M Muskelschichtregion, Fd dorsale
Fv ventrale Flosse, ac A. caudalis, vc
V. caudalis, asv äussere Schwanzvene .
beiden äusseren Seiten der Muskelschicht, im Wurzelstück des Schwanzes und verläuft rumpfwärts in der mittleren Höhe der Muskelschicht, fast parallel mit der Schwanzachse. An der Grenze von Schwanz und Rumpf wendet sie sich etwas nach unten und ergiesst
sich dann sofort in V. cardinalis post .. Das Blut aus der dorsalen Flosse
des Wurzelstückes wird von diesen Nebenvenen aufgenommen . Nur bei
der Bufolarve fehlt diese Vene; hei der Rhacophoruslarve entwickelt sie
sich am besten (Fig. 1).
2. Einfache Elimination der V. caudalis.
a) Bufo vulgaris.
Bei der Operation betrug die Länge der Larve ca. 24 mm (Schwanz 17 mm). Im Schwanz sind die Vasa caudalia und ihre Seitenäste schon unter der Binokularlupe bemerkbar.
Zuerst habe ich dies Blutgefäss in der Mitte durchgeschnitten, so dass in der kaudalen. Hälfte des Schwanzes Stauung auftrat. Als sich aber die beiden Schnittenden nach einigen Tagen direkt miteinander verbanden, wurde der Kreislauf wieder vollkommen hergestellt.
Ich eliminierte ein Stück (1 mm) von V. caudalis an derselben Stelle. Die Operation geschah Anfang April. Die durch die Operation verur-sachte Blutung war nach mehreren Minuten vollkommen gestillt. Die Stauung trat auch in der kaudalen Hälfte auf. Am Tage nach der Operation fliesst das durch die direkt kaudal von der Operationsstelle liegenden. Arterienäste in die Flosse gesandte Blut möglichst weiter proximalwärts in der Flosse und kehrt dann durch die nahe der Opera-tionsstelle vorhandenen proxinaalen. Venenäste zur V. caudalis zurück, so dass in den beiden Flossen der betreffenden Region der Blutfluss ziemlich lebhaft wird. Die Stauung dauert aber noch an. Am 2. Tag nach der Operation wird das in der Kaudalhälfte gestaute Blut nunmehr
durch V. caudalis proxirnalwärts gesandt und fliesst an der Operations-stelle wieder durch einige Venenäste in die Flosse. Dann nimmt es durch die Kapillaren, besonders durch die randständigen, den gleichen Verlauf wie oben, zur V. caudalis zurück. Hier beginnt der ganze Kreislauf im Schwanz sich wiederherzustellen.
Anfangs benutzt das Blut einige feinere Bahnen. Allmählich aber nimmt es stets denselben, bestimmten Weg, der möglichst nahe der
Die nach Teilelimination der V. caudalis ausgApildete Seitenbahn (s) hei Bufolarve
(schematisch).
Schwanzachse liegt. Es werden
nämlich die axial befindlichen
Astehen bevorzugt, wodurch der
neue Weg kürzer wird als der
frühere.
4-5 Tage nach der Operation
ist die neue Bahn fertig, deren
Dicke der der Hauptbahn entspricht d.h. der Defekt nach der
Elimina-tion ist nunmehr vollkommen
ersetzt. Vorläufig spreche ich hier von einer „Seitenbahn" (Fig. 2).
Doch ist damit selbstverständ-lich etwas anderes gemeint als beim
erwachsenen Tier. Diese Seitenbahn verläuft oberflächlich und anfangs schlängelnd, wird aber allmählich mehr gerade, und besteht, ist sie erst einmal vollkommen ausgebildet, endgültig.
bemer-636 Tatauo Fukai,
kenswert, worüber in der Diskussion eingebend gesprochen werden soll. An der Operationsstelle bemerkte ich Neubildung der Kapillaren. Bei 39 von 43 beobachteten Exemplaren verläuft die neue Bahn in der yen-tralen Flosse, bei einem an der Aussenfläche der Muskelschicht und bei den übrigen zeigt sie einen doppelten Verlauf (in jeder Flosse je eine Bahn). Als verursachende Momente dafür, dass die Seitenbahn meistens in der ventralen Flosse entsteht, dürften die Wirkung der Schwerkraft, die Dickendifferenz der Seitenäste u.a. in Frage kommen.
b) Rhaconhorus Schlerelii arborea.
Bei der Operation betrug die Länge der Larve 20 mm (Schwanz 15 mm). Die Gefässanordnung im Schwanz ist unter der Binokularlupe wegen des geringeren Pigmentgehaltes besonders gut bemerkbar. Ein Stück (1 mm) von V. caudalis wurde in dpr Gegend der Schwanzwurzel d. 11. in der Nähe der „äusseren Schwanzvene" eliminiert. In diesem Fall stellte sich der Kreislauf in zwei Formen wieder her.
Erstens: Das durch V. caudalis proximalwärts gesandte Blut floss an der Operationsstelle dorsalwärts durch den dorsalen dickeren Ast, der sich fast direkt kaudal von der Operationsstelle verzweigte, und ergoss sich dann in die „äussere Schwanzvene" auf der einen Seite durch die in der Aussenfläche der Muskelschicht vorhandenen Kapillaren. Am Tag nach der Operation entstand hier eine dicke Vene (die ich vorläufig „Verbindungsbahn" nenne), so dass der ganze Kreislauf vollständig wiederhergestellt war. Dieses Verhalten dürfte ebenfalls sehr beachtens-wert sein (Fig. 3 A).
Zweitens: In diesem Fall wurde der Kreislauf durch die
Seiten-Die nach Teileliminatiön der V. caud. ausgebildeten neuen 1 tahnen hei Bam-- phoruslarve (schematisch). vb Verbin-dungsbahn, ov obere Schwanzvene, s
Seitenbahn.
bahnbildung, wie bei Bufolarven, wiederhergestellt.
Bei 26 von 30 beobachteten Fällen wurde die Verbindungsbahn und bei den übrigen die Seitenbahn gebildet. Hieraus erkennt man, dass bei der Wiederherstellung das topische Verhalten der Blutgefässe eine grosse Rolle spielt.
Beinerkeilswert ist es auch, dass die „obere Schwanzvene" bei diesem Experiment in einigen Fällen entstand, ungeachtet dieser Wiederherstellungsformen. Das
aus der dorsalen Flosse zurückkehrende Blut sammelte sich nämlich im Oberrand der Muskelschicht und floss weiter denselben entlang proximalwärts zur äusseren Schwanzvene (Fig. 3 A, B). Dieses Ergebnis beruht darauf, dass die Richtung des Blutflusses infolge der durch die Elimination verursachten Stauung des Blutes sich veränderte.
Weiter eliminierte ich ein Stück (1 mm) von V. caudalis im Mittel-stück des Schwanzes d.h. an einer von der äusseren Schwanzvene entfernten Stelle. Diesmal wurde der Defekt nur durch Seitenbahnbil-dung wiederhergestellt (Fig. 3 B). Diese fand aber immer in der dorsalen Flosse besonders längs des Oberrandes der Muskelschicht statt. Bei der Rhacophoruslarve sind einige dickere Äste vorhanden, die sich nur in die dorsale Flosse verzweigen und zwar sich in einem bestimmten Abstaude einstellen. Meistens werden bei der Seitenbahnbildung nur diese dickeren Äste verwendet. Deshalb ist es verständlich, dass die dickeren Äste für die Seitenbahnbildung zunächst in Betracht kommen.
3. Elimination der V. caudalis an zwei Stellen.
Ich beabsichtigte hauptsächlich durch dieses Experiment, das
Schick-sal des zwischen beiden Eliminationsstellen vorhandenen Gefässstückes
zu erforschen. Die Elimination geschah so, dass einige Äste zwischen
den beiden Stellen derselben bestehen blieben. Der Blutkreislauf wurde
bei der Bufolarve durch Seitenbahnbildung an beiden Stellen ergänzt;
das zwischen den beiden Stellen zurückgelassene Stück wurde bei der
Wiederherstellung benutzt. Die Seitenbahnen entstanden an beiden
Stellen fast gleichzeitig.
Bei der Rhacophoruslarve wählte ich die Wurzelgegend des Schwanzes,
d.h. die nächste Umgebung der äusseren Schwanzvene als erste Opera-
tionsstelle und die Kaudalgegend als zweite. Die Wiederherstellung
geschah in der Mehrzahl der Fälle wie folgt : An der ersten Stelle wurde
eine Verbindungsbahn zwischen V. caudalis und der äusseren
Schwanz-vene, und an der zweiten eine Seitenbahn gebildet. Das zwischen den
beiden Stellen vorhandene Stück wurde bei der Wiederherstellung meist
benutzt (Fig. 4 A). In einigen Fällen wurde es jedoch als Hauptbahn
niemals verwendet. Das Blut floss nämlich an der zweiten Stelle
dor-salwärts durch den dorsalen Ast zum Oberrand der Muskelschicht und
längs desselben weiter proximalwärts in die äussere Schwanzvene, indem
diese neue Bahn fast alles aus der dorsalen Flosse zurückkehrende Blut
aufnahm. Es handelt sich um eine typische obere Schwanzvene (Fig.
4 B).
638 Tateuo Fukai,
in diesem Fall ? In bezug auf diese Frage wurden zwei Formen be-obachtet. Erstens: Es nimmt das aus der ventralen Flosse zwischen den
beiden Stellen zurückkehrende Blut auf und dieses fliesst proximalwärts
Die nach Teilelimination der V. cand. an zwei Stellen wiederhergestellte Gefässanordnung bei Racophoruslarve (schematisch). Nb neue
Bahn.
zur äusseren Schwanzvene durch eine etwas feinere Ver-bindungsbahn hin (Fig. 4 B). Zweitens: Das aus der ven- traten Flosse betreffender Re-gion zurückkehrende Blut fliesst umgekehrt kaudalwärts in dem Zwischenstück und an der zweiten Operationsstelle dorsalwärts durch den dickeren Ast in die neugebildete obere Schwanzvene (Fig. 4 C). Dies stete Zurücklenken durch die Hauptbahn scheint mir sehr bemerkenswert zu sein. Die neue Bahn verläuft in ver-schiedener Weise, aber der Wiederherstellungsprozess ist ganz ähnlich wie bei der
ein-fachen Elimination d.h. sie entsteht ausnahmslos durch eine rasche
Umgestaltung der feineren Äste oder der Kapillaren zur Hauptbahn.
4. Elimination der A. caudalis.
Zuerst sei die einfache Elimination beschrieben. Material und
Entwicklungsstadium waren ebenso wie beim vorhergehenden
Experi-mente. Die Operation geschah Anfang April. In dem Mittelstück des
Schwanzes exstirpierte ich ein Stück (1 mm) von A. caudalis. Nach der
Operation trat Stauung in der kaudalen Hälfte auf. Am Tage nach der
Operation verdickten sich die direkt proximal von der Operationsstelle
liegenden beiden Arterienäste, und das Blut floss durch diese in die
beiden Flossen ab.
Das in die Flosse gesandte Blut wird grösstenteils durch die
Venen-äste zur V. caudalis, zu einem Teil aber durch die randständigen
Kapillaren möglichst kaudalwärts getrieben. Ich habe bisweilen
kon-statiert, dass das in die V. caudalis zurückgekehrte Blut umgekehrt
kaudalwärts fliesst und durch Venenäste in die Flosse des kaudaleu
Wiederherstel-lung des Blutkreislaufes, indem das Blut durch die in der Oberfläche der Muskelschicht vorhandenen Kapillaren zur A. caudalis im Kaudal-stück strömt.
Anfangs fliesst das Blut sehr langsam und zwar mehrere Wege benutzend. Allmählich aber wird der Blutfluss lebhafter und die
Die nach Teilelimination der A. caud. ausgebildete Seitenbahn (s) bei Bufolarve (schematisch).
Kapillaren verdicken sich sehr rasch für einen bestimmten Weg. Nach 7-8 Tagen ist eine neue Seitenbahn entstanden, deren Verlauf möglichst kurz ist und zwar kürzer als bei der Vene (Fig. 5). Diese ersetzt den Defekt nach der Elimination endgültig und vollständig. Ihre Lage ist immer etwas mehr nach der Oberfläche hin abweichend. Das Venensystem im Schwanz ist niemals von der Seiten-bahnbildung der Arterie wesentlich be-einflusst. An der Operationsstelle ent-wickeln sich später einige Kapillaren.
Für die Neubildung der Seitenbahn werden sogar feinere Äste, wenn sie
sich in der Nähe der Operationsstelle befinden, mitherangezogen. Es ist
dies etwas anders als bei der Vene. In der Mehrzahl der Fälle verläuft
die Seitenbahn in der Muskelschichtregion (17 Fälle) , bei 2 Fällen in
der dorsalen Flosse und bei 5 Fällen in der ventralen . Im ganzen
beob-achtete ich nämlich 24 Exemplare. Als Grund dafür , dass die Seitenbahn
meistens in der Muskelschichtregion entstand, lässt sich anführen , dass
in der Aussenfläche der Muskelschichtregion die Kapillaren ziemlich
reichlich sind und bei der Bildung der Arterie sogar feinere Ästchen gut
brauchbar werden. Bei der Rhacophoruslarve geschieht die Wiederher
-stellung fast in der gleichen Weise wie bei der Bufolarve .
Zweitens habe ich Stücke (1 mm) der A . caudalis an zwei Stellen
entnommen. Die Wiederherstellung fand auch hier an beiden Stellen
durch Seitenbahnbildung und zwar zuerst (am 2. Tag nach der Operation)
an der proximalen Stelle und dann (am 4. Tag nach der Operation)
an der distalen statt. Auch in diesem Fall wurde das Zwischenstück
benutzt.
Die zeitliche Differenz in der Seitenbahnbildung bei beiden Elimina
-tionsstellen wurde nur bei der Arterie nachgewiesen , während sie bei der
Vene gleichzeitig an beiden Orten stattfand. Deshalb lässt sich offenbar
schliessen, dass für die Seitenbahnbildung der Druck des Blutstromes
640 Tatsuo Fuksi,
Weiter habe ich die gleichzeitige Elimination (exstirpiertes Stück 1 mm) der Vasa caudalia vorgenommen. Die operierte Stelle der A. caudalis liegt mehr proximal als die andere, Die Seitenbahn wurde hierbei zuerst bei der Arterie und dann bei der Vene gebildet. Es ist also deutlich, dass auch bei der Seitenbahnbildung der Vene der Blut. stroin nötig ist.
5. Elimination der Vasa caudalia bei der Metamorphose.
Wird der Defekt der Blutgefässe auch während ihres Rückbildungs-prozesses ersetzt? Vorausgesetzt nun, dass dies der Fäll ist, in welcher Weise dann geht die Wiederherstellung vor sich? Das soll hier unter-sucht werden. Ich benutzte 23 mm lange Bufolarven, deren Schwanz die beginnenden Rückbildungszeichen d. h. Pigmentzunahme oder Knickung des Spitzenteils zeigte. Das Stück (1 min) der A. oder V. caudalis wurde im Mittelstück des Schwanzes eliminiert.
Bei der Vene ist der Defekt schon am Tag nach der Operation durch Seitenbahnbildung vollkommen ersetzt. Der Wiederherstellungsprozess ist ähnlich wie beim vorigen Experimente. Bei der Arterie fand am Tag nach der Operation keine Wiederherstellung statt. Am 2. Tag nach der Operation schritt die Rückbildung des Schwanzes hochgradig vor, so dass eine eventuelle Neubildung der Bahn nicht mehr festgestellt werden konnte. Als Grund dafür, dass bei der Arterie keine Seitenbahnbildung stattfand, kann man darauf verweisen, dass die Ernährung unnötig ge-worden und infolgedessen mit einer Abnahme des arteriellen Blutes zu rechnen ist. Dagegen lässt sich denken, dass bei der Vene das erhöhte Bedürfnis des Blutflusses wegen der Resorption des zerfallenen Gewebes im Schwanz die rasche Bildung der Seitenbahn betrieb.
13. Teilelimination der Vasa caudalia in möglichst frühem Stadium.
Ich beabsichtigte in diesem Experiment, die Wiederherstellung der Blutgefässe in ei nein sehr frühen Entwicklungsstadium zu untersuchen. B. min lange Bufolarven (Schwanz 2 mm) wurden verwendet, bei denen Stücke (0.5 rnm) der Vasa caudalia gleichzeitig in der Mitte des Schwanzes eliminiert wurden. Im Schwanz waren nur die Hauptgefässe, aber mit Andeutung der Seitenastbildung, im Schnittpräparat nachweisbar. Die Operation geschah Ende April.
Die Wunde schloss sich am Tage nach der Operation. Erst nach einer Woche wurden die Blutgefässe am lebenden Material unter dem Mikroskop nachgewiesen. Der durch Elimination erzeugte Defekt war vollkommen durch die Neubildung ersetzt. Die Gestaltung derselben
fand wie folgt statt : I. A. caudalis verläuft in der operierten Region etwas dorsal abbiegend d.h. es entsteht die Seitenbahn. V. caudalis zieht sich, in der betreffenden Region, in der mittleren Höhe der ven-tralen Flosse (20 Exemplare) hin oder in dem Rand derselben (8 Ex-emplare). Es ist dies auch eine Seitenbahn. Ihr Verlauf ist bei der Arterie sehr kurz, bei der Vene aber nicht immer so kurz. Bei 28 von -30 Exemplaren trat die neue Bahn in dieser Weise auf (Fig . 6 A). 2. A. caudalis verläuft wie in den obigen Fällen etwas dorsal abbiegend in der operierten Region, V. caudalis verläuft in der Kaudalhälfte des Schwanzes in der mittleren Höhe der ventralen Flosse und sammelt das aus derselben zurückkehrende Blut. Das Blut aus der dorsalen Flosse sammelt sich in einem dickeren dorsalen Ast, der sich in der Wurzel in die V. caudalis ergiesst. In der kaudalen Hälfte beobachtet man keine Hauptvene, die normal verläuft. Es lässt sich deshalb schliessen, dass die früher dort vorhandene Vene nach der Operation atrophierte. Die obige neue Hauptvene verläuft ziemlich gewunden (Fig. 6 B).
Ferner habe ich ein Stück (0.5 mm) der Muskelschichtregion mit ihren Gefässen ausgeschaltet. Nach einigen Tagen wurde das fehlende
Fig. 6.
Die nach Teilelimination der Vasa caudalia in möglichst frühem Stadium wiederherge-stellte Gefässanordnung bei Bufolarve (schematisch). L
Lücke.
Stück der Muskelschichtregion durchi
Regeneration des Mesenchyms ersetzt und
die eliminierte Blutgefässpartie ebenfalls
durch Seitenbahnbildung (Fig. 6 C, D).
Diese vollzog sich in sehr verschiedener Form.
Die Seitenbahn der Vene verläuft in
der ventralen Flosse (13 Exemplare) oder
in dem regenerierten Achsenteile (7
Ex-emplare); die der Arterie in der dorsalen Flosse (13 Exemplare) oder in der ventralen (5 Exemplare) oder in dein regenerierten
Achsenteile (2 Exemplare). Die
Seiten-bahn ist bei der Arterie sehr kurz, bei
der Vene dagegen manchmal länger.
Die oben beschriebene neue Bahn
muss durch eine rasche Umgestaltung der
Kapillaren oder der feineren Äste, die
aber meist erst nach der Operation ganz
neu entstanden sind, sich gebildet haben.
In der Diskussion werde ich hierauf
642 Tatsuo Fukai,
C. Totale Elimination der Vasa caudalia.
Die Untersuchung betrifft die Wiederherstellung der Blutgefässe nach ihrer totalen Eliinination. Zuerst habe ich das Experiment an Bufolarveii von 8 mm Körperlänge vorgenommen. Der Schwanz betrug 2 mm, in ihm sind fast nur die Hauptgefässe wahrnehmbar. Ich habe die ventrale Hälfte des Schwanzes, von dem Unterrand der Chorda ab, fortgeschnitten. Die Vasa caudalia sind natürlich durch diese Operation
Inaausgeschaltet.
Einige Tage nach der Operation wurden wieder Blutgefässe gesehen. Es hatte nämlich eine totale Neubildung derselben stattgefunden. Die ventrale Hälfte war regeneriert. 43 Larven wurden zur Beobachtung gebracht. A. caudalis verläuft ausnahmslos den Unterrand der Chorda entlang und zwar etwas schlängelnd und oberflächlich von der Wurzel zum Schwanzende hin (Fig. 7 A—G). Die Verzweigung ist fast normal.
Fig. 7.
Die nach totaler Elimination der Vasa cand. wiederhergestellte Gefässanordnung bei Bufolarve (schematisch). Frv regenerierte
ventrale Flosse.
V. caudalis zeigte dagegen, was Lage und Verlauf betrifft, mannigfache Gestaltungen : 1. (Fig. 7 A) In Wurzel und Spitze verläuft sie normal, in der übrigen Region, die den grössten Teil des Schwanzes ausmacht, in der dorsalen Flosse und zwar in der mittleren Höhe derselben (15
verläuft sie normal. In der übrigen Region zieht sie sich längs des dorsalen Randes der Muskelschicht zur Spitze hin (12 Exemplare). Dies kann man von der typischen oberen Schwanzvene behaupten. 3. (Fig. 7 C) In der Wurzel verläuft sie normal. In der anderen Region zieht sie sich in der mittleren Höhe der dorsalen Flosse hin (2 Exemplare). 4. (Fig. 7 D) In der Wurzel und Spitze verläuft sie normal, in der übrigen Region den Oberrand der Muskelschicht entlang (3 Exemplare). 5. (Fig. 7 E) In Wurzel und Spitze verläuft sie normal und im übrigen in der mittleren Höhe der Muskelschichtregion (3 Exemplare). 6. (Fig. 7 F) Zwei gleichdicke Venen entstanden, von denen die eine in der mittleren Höhe der dorsalen Flosse und die andere in der mittleren Höhe der regenerierten ventralen Flosse verlief. Die beiden Venen vereinigten sich miteinander in der Wurzel (2 Exemplare).
Die neugebildete Vene verläuft auch sehr gewunden und etwas oberflächlich. In der dorsalen Flosse fliesst das Blut durch zahlreiche
feinere Ästchen direkt in die neugebildete Hauptvene. Das aus der regenerierten ventralen Flosse zurückkehrende Blut wird in folgenden zwei Formen zentralwärts gesandt : 1. Es wird von einer hier neuge-bildeten dickeren Vene aufgenommen und ergiesst sich bei der Wurzel in die Hauptvene (Fig. 7 G). 2. Das Blut fliesst in der Hauptsache in zwei Richtungen d.h. proximalwärts und kaudalwärts; das letztere fliesst durch einen etwas dickeren Ast in die Hauptbahn, die in der dorsalen Flosse entstand und wird in der Spitzengegend aufgenommen, während das erstere durch einen dickeren Ast proximalwärts zur Hauptbahn in der Wurzelgegend gesandt wird. Die oben erwähnten Venen müssen alle durch die rasche Umgestaltung der nach der Operation ganz neuge- bildeten Kapillaren oder feineren Äste entstanden sein. Siehe Diskus- sion !
Ferner habe ich die A. caudalis an 20 mm langen Bufolarven total eliminiert. Doch trat in diesem Falle niemals eine Wiederherstellung der Gefässe ein und bald war der Schwanz wegen mangelnder Ernährung abgestorben. Bei der totalen Elimination der V. caudalis an ebensolchen Larven entstand dagegen in günstigen Fällen eine neue Hauptvene nach einigen Tagen und zwar ebenfalls durch die Umgestaltung von Ästen oder Kapillaren.
Bei der totalen Elimination der äusseren Schwanzvene der einen Seite bei der Rhacophoruslarve (20 mm) floss das Blut durch feinere Ästchen, teils in die V. caudalis, teils in die äussere •Schwanzvene der anderen Seite, und eine Neubildung der äusseren Vene wurde nirgends
644 Tatsilo Fukai,
D. Teilelimination der Rumpfgeesse.
Es sei hier das eine ziemlich komplizierte Anordnung aufweisende Rumpfgefäss zur Untersuchung gebracht. Als Material benutzte ich 20 min lange Rhacophoruslarven. Beobachtung der Gefässanordnung schon unter der Binokularlupe. Operation Anfang Juni.
1. Einseitige Elimination der V. jugularis externa.
V. jugularis externa entsteht durch den Zusammenhang von V.
lingualis und V. mandibularis interna, sie verläuft den lateralen Rand
des M. sternohyoideus entlang und ventral von Truncus arteriosus
rück-wärts zur V. cava ant.. Ihr Verhalten ist aus FW. 8 zu ersehen. Ich
lig. 8.
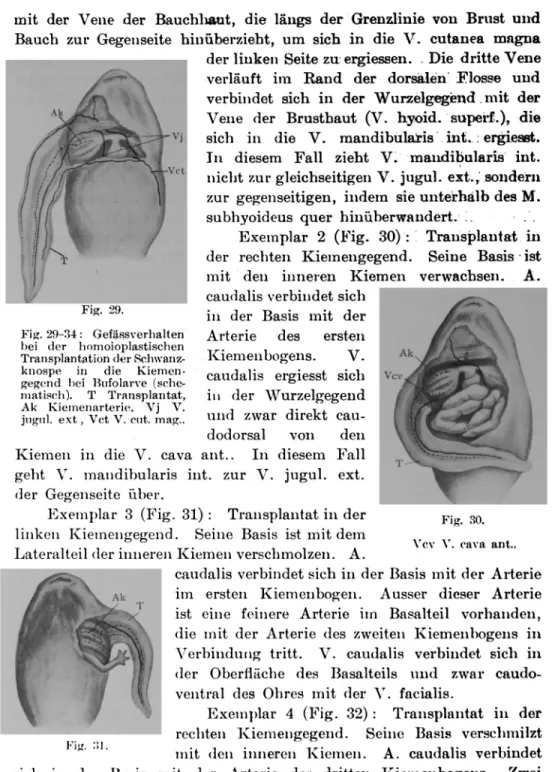
( ;efässanordnung der Brustregion von Racophoruslarve (schema-tisch). H Herz, T Truncus arte-riosus, K Kiemen, msx M. sub-maxilJaris, msh M. suhhyoideus, acr A. carotis ext., VIII V. mandi-hularis int., vl V. lingualis, vj V. jugidaris ext., vhh
vene.
v ernaiten ist aus r ö zu ersenen. Len eliminierte ein Stück (1 mm) der V. jugul.
ext. der linken Seite in der Nähe des
Zusammenflusses obiger zwei Venen, indem
ich die Brusthöhle öffnete.
Am Tag nach der Operation schloss sich die Wunde vollständig und in der
betreffenden Region trat Stauung auf. Am
2. Tag nach der Elimination fing das
staute Blut an, durch Verzweigungen oder
Kapillaren in verschiedener Richtung z. B.
in die jenseitige V. jugul. ext. oder in die
gleichseitige V. facialis oder in die Vene
der Bauchhaut (gleichseitig), die sich an
der Grenze von Brust und Bauch in die V.
cava ant. ergiesst, u.a. zu fliessen. Vor allein war der Abfluss zur Gegenseite sehr beträchtlich. Am 4. Tag nach der
tion fand die vollkommene
lung des Blutflusses statt, indem sich die
Verbindungsbahn oder die Seitenhahn bildete. Die Seitenbahn entstand
dadurch, dass das Blut von dem kranialen Stück der V. jugul. ext. durch
die in der Operationsgegend vorhandenen Kapillaren zum kaudalen Stück
derselben floss (Fig. 1) A). Diese Seitenbahn war sehr dick und ersetzte
den Defekt gänzlich. Nur bei 3 von 50 beobachteten Exemplaren trat diese
Seitenbahn auf, sonst bildete sich die Verbindungsbahn, wobei das von
der Operationsstelle zentral liegende Stück der V. jugul. ext. atrophierte.
Lage und Verlauf der Verbindungsbahnen sind sehr mannigfaltik.:
seifig), die normal das Blut aus der Bauchhaut sammeln und in die V. cava aut. sich ergiessen, indem das Blut durch die Zweige der V. man-dibularis int. (meistens V. hyoid. superficialis) und die Kapillaren in der Brusthaut zur soeben erwähnten Vene der Bauchhaut fliesst (10 Exemplare) (Fig. 9 B). 2. solche zwischen beiden Vv. jugul. ext.,
Fig. 9.
Die nach Teilelimination der V. jugul. ext. ausgebildeten neuen Bahnen bei Racophoruslarve (schematisch) . s Sei-
646 Tatsuo Fukai,
wobei das Blut durch die Zweige der V. maudibularis int. oder der V. liugualis und die in der Brustregion vorhandenen Kapillaren zur gegen- seitigen V. jugul. ext. fliesst (21 Exemplare) (Fig. 9 C). 3. solche zwischen V. jugul. ext. und V. jugul. int. der gleichen Seite, bei denen das Blut durch die Zweige der V. maudibularis int. oder durch die Kapillaren, und zwar das Mundhöhlendach durchbohrend, in die V. jugul. int. fliesst (1 Exemplar) (Fig. 9 D). 4. solche zwischen V. jugul. ext. und V.. facialis der gleichen Seite, bei denen das Blut in der V. mandibularis int. oralwärts rückströmt und dann durch die Zweige der V. facialis (meistens V. nasalis ext.) zur letzteren fliesst (15 Exemplare) (Fig. 9 E, E'). Die Rückströmung des Blutes, die stets und zwar durch die dickere Vene stattfand, ist besonders beachtenswert. 5. Das Blut verteilt sich teils in die Gegenseite, teils in die Vene der Bauchhaut und teils in die V. facialis, so dass hierbei drei Verbindungsbahnen entstehen (I Exemplar). Die Verbindungsbahn hat die gleiche Dicke wie die Haupt-bahn. Der Entwicklungsprozess der Seitenbahn oder der Verbindungs-bahn ist ebenfalls ganz derselbe wie bei den Schwanzgefässen d.h. die Bahnen entstehen durch die rasche Umgestaltung der Zweige und Kapillaren.
2. Beiderseitige Elimination der Vv. jugulares exteriiae.
Die Elimination der V. jugul. ext. wurde diesmal auf beiden Seiten vorgenommen. Die Operationsstelle ist dieselbe wie bei der einfachen Elimination. Die Wiederherstellung geschah fast in der gleichen Weise wie bei der einfachen Elimination d.h. durch die Bildung der Verbin-dungsbahn oder der Seitenbahn, die durch die rasche Umgestaltung der Zweige und der Kapillaren zur Hauptbahn entstanden. Das Verhalten der neugebildeten Bahnen war etwas kompliziert, weil diese gleichzeitig auf beiden Seiten auftraten:1. Auf jeder Seite entstand die Verbin- dungsbahn zwischen V. jugul. ext. und der Vene der Bauchhaut (5 Exemplare). 2. (Fig. 10 A) Die beiden Verbindungsbahnen flossen zu einer sehr starken Bahn in der Brusthaut zusammen, verliefen dann weiter in derselben, um sich mit der Vene der Bauchhaut zu vereinigen (4 Exemplare). 3. (Fig. 10 13) Auf der rechten Seite entstand die Ver-bindungsbahn zwischen V. jugul. ext. und V. facialis und auf der linken zwischen V. jugul. ext. und der Vene der Bauchhaut (1 Exemplar). 4. Links entstanddie Seitenbahnund rechts die Verbindungsbahn zwi- schen V. jugul. ext. und V. facialis (2 Exemplare). 5. Links entstand die Seitenbahn. Rechts die Verbindungsbahn zwischen V. jugul. ext. und V. jugul. int. der gleichen Seite (2 Exemplare). 6. (Fig. 10
Das Blut in der rechten Seite floss durch die Verbindungsbahn in 'die V, jugul. ext. der anderep Seite und es entstand dort die Seitenbahn (6 Exemplare). 7. Beiderseits Bildung der Seitenbahn (6 Exemplare). 8. Auf beiden Seiten entstand die Seitenbahn, dazu bildete sich zwischen beiden Seiten eine Verbindungsbahn (1 Exemplar). 9. (Fig, 10 D) Das
Fig. 10.
Die nach beiderseitiger Elimination der Vv. jugul. ext. wiederhergestellte Gefässanordnung hei Racophoruslarve
(schematisch). s Seitenbahn, vb Verbindungsbahn.
Blut in der linken Seite floss durch zwei Verbindungsbahnen in die V .
jugul. ext. der anderen Seite und es entstand die Verbindungsbahn zur
Bauchhautvene (5 Exemplare). In jedem Fall ist der Verlauf der neuen
Bahn sehr mannigfaltig. Die gesamte Zahl der beobachteten Fälle betrug
39.
3. Elimination der A. carotis externa.
A. carotis externa geht aus dem Kiemengefäss des ersten Bogens
648 Tatsuo Fukai,
der Rhacophoruslarve ist diese Arterie während der Operationszeit stark pigmenthaltig. Ein Stück (1 mm) dieser Arterie wurde in dorsokranialer Gegend des M. subliyoideus ausgeschaltet.
Nach einer Woche war der Blutfluss vollkommen wiederhergestellt, indem sich die Seitenbahn gebildet hatte. Die Verbindungsbahn wurde in diesem Fall nirgends nachgewiesen. Die Seitenbahn entstand dadurch, dass sich die feineren Zweige der A. carot. ext. oder die Kapillaren rasch zur Hauptbahn umgestalteten. Diese Seitenbahn ist pigmentarm, so dass sie deutlich von dem alten Stück zu unterscheiden ist und zieht sich nur eine ziemlich kurze Strecke hin. Der Verlauf dieser Seitenbahn ist ebenfalls ein mannigfacher. Im ganzen wurden 31 Fälle beobachtet. 1. (Fig. 11 A, B) Sie zieht in der Gegend des Oberrandes von M.
sub-Fig. 11.
Die nach Teilelimination der A. carotis ext. ausgebildete Seitenhahn (s) bei Racophoruslarve (schematisch). mgh M.
gen iohyoideus.
hyoideus etwas ventromedianwärts und dann unter der Brusthaut
vor-wärts, um sich in der Gegend des Unterrandes des M. submaxillaris
mit dein alten Stück der A. carot. ext. zu vereinigen (22 Exemplare).
2. (Fig. 11 C) Sie verläuft in der dorsalen Gegend des M. subhyoideus
dorsomedianwärts und dann fast längs des M. geniohyoideus vorwärts,
um sich mit dein alten Stück der A. carot. ext. in der dorsokaudalen
Gegend des M. submaxillaris zu vereinigen (7 Exemplare). 3. (Fig.
denen. die eine ähnlich wie bei Fall 1. und die andere wie bei Fall 2. verlief (2 Exemplare). Die Seitenbahnbildung scheint auf die andere Arterie oder Vene keinen beträchtlichen Einfluss auszuüben.
Diskussion.
Beim Embryo werden die Blutgefässe in mannigfacher Weise wie-derhergestellt. Die Regeneration ist immer von verschiedenen
Momen-ten z. B. den Arten der Blutgefässe, ihrem topischen Verhalten, von der Eliminationsstelle, dem Entwicklungsstadium usw. abhängig. Ich gehe hierauf sogleich im Kapitel über die Wiederherstellungsformen und Wiederherstellungsprozesse der Blutgefässe näher ein.
A. Über die Wiederherstellungsformen der Blutgefässe.
Der durch die Elimination erzeugte Defekt der Blutgefässe wurde in meinem Fall durch die Bildung einer neuen Bahn ersetzt. Ich unterschied bei dieser neugebildeten Bahn drei Formen d. h. die Bildung einer Seitenbahn, einer Verbindungsbahn oder eines selbständigen Blutge-fässes.
1. Unter der „Seitenbahn" ist eine neue Bahn zu verstehen, die beide Enden des künstlich unterbrochenen Gefässes verbindet. In meinem Fall hat diese die gleiche Dicke wie die Hauptbahn und ersetzt den Defekt vollkommen. Deshalb behaupte ich, dass die Seitenbahn in diesem Fall, d.h. beim Embryo, sicher ein Stück des Hauptgefässes ausmacht, während sie beim ausgewachsenen Tier nur als Nebenbahn auftritt. Ihre Bildung fand in folgenden Fällen statt : a. Teilelimination der Schwanzgefässe bei 24 mm langer Bufolarve (alle Fälle). b. Teilelimination der Schwanz-gefässe von 20 min langer Rhacophoruslarve in der Kauda]hälfte (alle Fälle). c. Teilelimination der A. carot. ext. von 20 mm langer Rhaco-phoruslarve (alle Fälle). d. Teilelimination der Vasa caudalia von 8 mm langer Bufolarve (in der Mehrzahl der Fälle). e. Teilelimination der V. caudalis von 20 min langer Rhacophoruslarve in der Wurzel (einige Fälle). f. Teilelimination der V. jugul. ext. von 20 min langer Rhacophoruslarve (einige Fälle) u. a. in..
Es trat also die Seitenbahn bei Teilelimination auf. Bei der Arterie wird der Teildefekt ausnahmslos durch Seitenbahnbildung ersetzt. Da-gegen ist die letztere bei der Vene, wie oben gesagt, eine Form der Wiederherstellung. Dies beruht vielleicht darauf, dass bei Venen ziem-lich stark Anastomose stattfindet, während dies bei Arteriell wenig der Fall ist. Bei. der Rumpfvene, die eine etwas komplizierte Anordnung
650 Tatsuo Fukai,
zeigt, war die Seitenbahnbildung seltener als bei der Schwanzvene.
Daraus lässt sich entnehmen, dass die Wiederherstellung auch von dem
topischeii Verhalten abhängen muss. Bei der Rhacophoruslarve, deren
Schwanz eine äussere Schwanzvene besitzt, wird der Defekt durch
Seiten-bahiibildung, wenn die Elimination in der Kaudalhälfte geschah, und
in einer anderen Form (Verbindungsbahnbildung), wenn sie in der
Wurzel stattfand, wieder ausgeglichen. Hieraus sieht man, dass die
Elirninationsstelle für die Art der Wiederherstellung von Wichtigkeit ist.
Sowohl bei 24 mm als auch bei 8 mm langen Bufolarveii entstand die
Seitenbahn. Das beweist, dass die Seiten bahn sowohl in frühem wie
auch in späterem Stadium entstehen kann. Diese Seitenbahn verläuft
für gewöhnlich möglichst kurz, besonders bei der Arterie, in sehr frühem
Stadium dagegen nicht immer so kurz. Dies beruht wohl darauf, dass
der Prozess der Seitenbahnbildung von dem Entwicklungsstadium
ab-hängig ist. Im Kapitel über die Wiederherstellungsprozesse der
Blutge-fässe wird des Genaueren hierüber gehandelt.
2. Eine dicke Bahn, die die zwei Hauptgefässe verbindet, nenne
ich hier vorläufig „Verbindungsbahn". Diese Verbindungsbahn besitzt
natürlich die gleiche Stärke wie die Hauptgefässe. Das Blut aus einem
Gefäss wird durch die neugebildete Verbindungsbahn, wegen der
Unter-brechung des Blutstromes in dem betreffenden Hauptgefäss selber, zum
anderen Hauptgefäss gesandt. Solche Bahnen traten bei mir in folgenden
Fällen auf : a. bei der Teilelimination der V. caudalis von 20 mm langer
Rhacophoruslarve in der Wurzelgegend zwischen V. caudalis und der
äusseren Schwanzvene (mehrere Fälle). b. bei der Teilelimination der
V. jugul. ext. derselben Larven zwischen V. jugul. ext. und entweder
der gegenseitigen V. jugul. ext. oder der V. facialis oder der V. jugul.
iiit. oder der Vene der Bauchhaut (mehrere Fälle).
Sie trat also bei Teilelimination und zwar hei Venen auf. Dies
scheint, wie oben gesagt, darauf zu beruhen, dass die Anastomose im
Venensystem verhältnismässig ausgiebig ist. Sie entstand auch dort, wo
die Gefässanordnung ziemlich kompliziert, war. Wenn das Blut durch
solch eine Verbindungsbahn zum anderen Hauptgefäss fliesst, atrophiert
meistens das zentral von der Operationsstelle liegende Stück des Haupt-
gefässes. Inwenigen Fällen aber entstanden Seitenbahnund Verbin-
dungsbahn gleichzeitig. Das bedeutet, dass die Zirkulation durch die
Mitarbeit noch einer zweiten neuen Bahn wiederhergestellt wird. Die
einmal neugebildete Verbindungsbahn besteht auch endgültig.
Bezüg-lich ihres Verlaufes ist zu sagen, dass sie verschiedene Wege einschlägt,
3. Ein neues selbständiges Hauptgefäss entstand in folgenden
Fällen : a. Bei der totalen Elimination der Vasa caudalia bei 8 mm
langer Bufolarve fand die Wiederherstellung der Vasa caudalia d.h. die
vollkommene eneration derselben nach mehreren Tagen statt (alle
Fälle). h. Bei der totalen Elimination der V. caudalis von 24 min langen
Bufolarven trat eine ganz neue Hauptvene auf (im günstigen Fall). c.
Bei der Teilelimination der Vasa caudalia von S mm langen Bufolarven
entstand ebenfalls eine neue Vene, wobei die alte Vene atrophierte (einige
Fälle). Die totale Neubildung eines Gefässes findet sich demnach meistens
bei dem totalen Defekt und selten bei Teilelimination. Diese Neubildung
tritt im früheren Stadium leicht, im späteren dagegen ziemlich selten
auf, insbesondere im Arteriensystem. Mit anderen Worten, diese
Neu-bildung des Blutgefässes ist nur beim Embryo möglich, weil in so jungem
Stadium das Gefässbildungsvermögen sehr gross ist. Man kann diese
Erscheinung bei erwachsenen Tieren durchaus nicht beobachten. Uber
den Neubildungsprozess ist in einem späteren Kapitel die Rede.
4. Anschliessend seien hier einige weitere Befunde besprochen.
Bei der Teilelimination der Schwanzgefässe von 24 mm langen
Bufo-larven au zwei Stellen trat meist die Seitenbahnbildung an beiden Stellen
auf. Das zwischen beiden Stellen vorhandene Stück des Hauptgefässes
wurde bei wieder aufgenommener Zirkulation, und zwar nach mehreren
T: en, wie vorher benutzt. Deshalb kann man schliessen, dass der
längere Zeit fast untätig gebliebene Gefässteil sich gut als
Hauptgefäss-stück verwenden lässt. Ferner beobachtete ich, dass die Unterbrechung
des Gefässverlaufes, wie sie durch einen einfachen Schnitt hervorgerufen
wurde, durch die direkte Verbindung der Schnittstumpfe des Gefässes
leicht geheilt werden kann. Es scheint dies ein nur beim Embryo
auftretendes und wohl beachtenswertes Phänomen zu sein.
B. Uber die Wiederherstellungsprozesse der Blutgefässe.
Oben habe ich die Wiederherstellung der Blutgefässe heim Embryo
nach ihrer dreifachen Art der Neubahnbildung unterschieden und
be-sprochen. Es ist jedoch kurz zu sagen, dass beim Embryo die
Regenera-tion der Blutgefässe immer durch die Neubildung der Hauptbahn
statt-findet. Der Wiederherstellungsprozess dieser neuen Hauptbahn ist in
der Regel in jedem Falle bei den obigen drei Formen ganz der gleiche,
abgesehen von dem Entwicklungsstadium der Larve. Deshalb sollen
diese drei Fälle gemeinsam erklärt werden.
Es scheint mir, dass beim Wiederherstellungsprozess der Blutgefässe
652 Tatauo Fukai,
1. Zunächst der Wiederherstellungsprozess in etwas späterem Stadium. Bei 24 min langen Bufolarven oder bei 20 mm langen Rha-cophoruslarven wird die neuentstandene Anordnung der Blutgefässe untersucht. In diesem Falle wird das unterbrochene Blut infolge der Elimination der Blutgefässe durch die Verzweigungen oder Kapillaren zur Peripherie (bei der Arterie) oder zum Zentrum (bei der Vene) ge-sandt. Zu Beginn der Wiederherstellung fliesst das Blut in feineren Äderchen und zwar auf verschiedenen mehrfachen Wegen. Allmählich aber wird eine grössere Menge Blut auf einen mehr und mehr bestimmten Weg beschränkt, der meistens verhältnismässig kurzwegig das Blut transportiert, so dass sich der betreffende Weg infolgedessen verbreitert
und nach einigen Tagen zur neuen Hauptbahn entwickelt. Je nach der Lokalisation der Verzweigungen oder Kapillaren, die bei der
Wieder-herstellung verwendet werden, entsteht entweder eine Seitenbahn oder eine Verbindungsbahn oder ein selbständiges Blutgefäss. Daraus erkennt man, dass in dem etwas späteren Stadium die neue Hauptbahn durch die rasche Unigestaltung der schon existierenden Verzweigungen oder Kapillaren entstand.
Warum die neugebildete Hauptbahn meist nur einen kurzen Ver-lauf zeigt, trotzdem anfangs das Blut auf mehreren Wegen umfloss? Dies mag wohl auf die grössere Zweckmässigkeit eines kurzen Weges zurückzuführen sein. Ausserdem muss man aber auch wohl mechani-sche Bedingungen z. B. den Verzweigungswinkel der Blutgefässe, die Blutströmung usw. in Betracht ziehen. Doch verlangt diese Frage noch eine weitere Untersuchung.
Die neue Hauptbahn verläuft anfänglich sehr gewunden und wird erst allmählich mehr gerade. Sie ist gleichsam eine Zusammensetzung aus mehreren Verzweigungen oder Kapillaren, daher die anfänglichen Krümmungen. Nach und nach wird sie aber wohl dadurch glatt, dass die Knickungen durch die Umgestaltungskraft der Gefässe oder durch den Druck des Blutstroms verschwinden.
Bei Venen sind die dickeren Zweige leicht zur Hauptbahnbildung verwendbar, bei Arterien dagegen werden sogar ziemlich feine Ästchen häufig benutzt. Die Wiederherstellung findet bei Venen früher als bei Arterien statt. Als diese Erscheinung veranlassende Momente kann man die Anastoniose der Gefässe, den Blutdruck, die Umgestaltungsfähigkeit usw. in Rechnung ziehen.
Bei gleichzeitiger Teilelimination der Vasa caudalin trat die Wieder-herstellung der Arterie früher als die der Vene auf, und zwar fand bei der Teilelimination der A. caudalis an zwei Stellen die Wiederherstellung,
an der proximal liegenden Stelle früher als an der anderen, statt. Aus
diesen Befunden ist zu schliessen, dass für die Wiederherstellung der
Blutgefässe der Blutstrom nötig ist.
Der Regeuerationsprozess wird von dem Klima ziemlich stark
be-einflusst. In meinem Fall geschah die vollkommene Wiederherstellung
im Juni am 1. oder 2. Tag nach der Operation, dagegen im April ca.
1 Woche nach derselben. Diese Verzögerung wird natürlich bei normalen
Entwicklungen ebenfalls beobachtet.
Eine totale Elimination der Blutgefässe wurde in diesem
Entwick-lungsstadiutn nur bei Venen durch die Regeneration der Hauptvene
ausgeglichen, bei der Arterie dagegen niemals. Es mag dies wie folgt
zu erklären sein : Im Fall der Arterie sterben Verzweigungen oder
Kapillaren ab, bevor eine neue Bahn entsteht, weil das
Umgestaltungs-vermögen der Verzweigungen oder Kapillaren ziemlich schwach geworden
und die Nahrungszufuhr infolge der Elimination fast gänzlich
unter-brochen ist. Bei der Vene wird jedoch im günstigen Falle die neue
Bahn gebildet, weil währenddein Nahrung durch die Arterie übermittelt
wird, trotzdem das Umgestaltungsvermögen ebenfalls geringer geworden
sein mag.
Im Stadium der Metamorphose wird der Defekt der Schwanzgefässe
auch nur bei der Vene ersetzt. Die Erklärung dürfte wohl ungefähr
dieselbe sein wie im vorigen Fall.
2. Der Wiederherstellungsprozess nun in früherem Stadium. Form
und Verlauf der neugebildeten Bahn sind in diesem Falle ebenfalls ganz
dieselben wie im späteren Stadium. Darum ist anzunehmen, dass die
neue Hauptbahn auch diesmal durch die rasche Umgestaltung der
Ver-zweigungen oder Kapillaren entstand. Bei der Operation war aber die
Blutgefässbildung noch eine ganz ungenügende, es waren nämlich nur
die Hauptgefässe im Schwanz; Verzweigungen oder Kapillaren waren
noch nicht sicher nachweisbar. Deshalb möchte ich annehmen, dass die
bei der Bildung der neuen Hauptbahn verwandten Verzweigungen oder
Kapillaren grösstenteils nach der Operation neugebildet wurden . Einige
deutliche Beispiele mögen dies erweisen. Bei der Ausschaltung eines
Stückes der Muskelschichtregion des Schwanzes von 8 mm langen
Bu-folarven wurde der Defekt des Achsenteiles durch Regeneration des
Mesenchyms und der Defekt der Blutgefässe durch Neubildung der
Hauptbahn ersetzt. Hierbei verlief die neue Hauptbahn in dein
rege-nerierten Achsengebiet. Bei der Ausschaltung der ventralen Hälfte des
Schwanzes derselben Larvenart wurde der ganze den Blutgefässen
654 Tatou° Fukai,
beobachtete ich, dass in einigen Fällen das Hauptgefäss in der rege-
nerierten ventralen Flosse verlief.
Jedenfalls werden im früheren Entwicklungsstadium zuerst die
feineren Ästchen oder Kapillaren gebildet, um sich dann ziemlich rasch
zum Hauptgefässe umzugestalten, das den Defekt der Blutgefässe
voll-ständig ersetzt.
Zusammenfassung.
1. Ich habe teilweise oder totale Elimination der Blutgefässe in verschiedenen Körpergebieten bei Amphibienlarven vorgenommen, um die Art der Regeneration der embryonalen Blutgefässe zu untersuchen.
2. Ich trennte Vasa caudalia durch einfachen Schnitt bei 24 mm laugen Bufolarven ; der unterbrochene Blutfluss war einige Tage nach der Operation durch selbsterzeugte direkte Verbindung der beiden Schnittstumpfe wiederhergestellt.
3. Ich eliminierte ein Stück der V. caudalis von Bufolarven (24 mm). Der Defekt war einige Tage nach der Operation durch Seiten-bahnbildung vollkommen ausgeglichen. Diese Seitenbahn verlief häufig in der ventralen Flosse.
4. In der Schwanzwurzel von Rhacophoruslarven (20 mm) eliminierte ich ein Stück der V. caudalis. Es entstand meistens eine Verbindungs-bahn zwischen der V. caudalis und der äusseren Schwanzvene. Geschah die Elimination in der kaudalen Gegend, so wurde eine Seitenbahn ge-bildet, die meist in der dorsalen Flosse verlief. Bisweilen sah man gleichzeitig eine obere Schwanzvene entstehen, ausser der Seitenbahn-oder Verbindungsbahnbildung.
5. Ich eliminierte an zwei Stellen ein Stück der V. caudalis bei Bufolarven (24 mm) oder hei Rhacophoruslarven (20 mm). Bei der Bufolarve entstand die Seitenbahn, das zwischen den beiden Stellen liegende Stück wurde bei der Wiederherstellung für die Hauptbahn wieder benutzt. Bei der Rhacophoruslarve entstanden Verbindungsbahn und Seitenbahn. Das Zwischenstück wurde in einigen Fällen für die
Hauptbahn nicht benutzt, sondern atrophierte mehr oder weniger. 6. Ein Stück der A. caudalis wurde bei Bufolarven (24 mm) und bei Rhacophoruslarven (20 mm) eliminiert. Der Defekt wurde durch die Seitenbalinbildung ersetzt. Die Seitenbahn verläuft meistens in der
Aussenfläche der Muskelschicht.
7. Ich eliminierte bei Bufolarven (24 mm) ein Stück der A. caudalis an zwei Stellen. An jeder Stelle entstand eine Seitenbahn, das
sehenstück wurde auch in diesem Falle für die Hauptbahn wieder ver-wendet.
8. Bei der Elimination der V. caudalis an zwei Stellen entstand die Seitenbahn gleichzeitig an beiden Stellen. Bei der Elimination der
A. caudalis an zwei Stellen wurde die Seitenbahn zuerst an. mehr proxirnal liegender Stelle und dann an kaudal liegender gebildet. Bei gleichzeitiger Elitnination der Vasa caudalia entstand zuerst die Seitenbahn der Arterie
und dann_ der Vene.
.9. Bei der Metamorphose wurde der Defekt der V. caudalis nach der Teilelimination durch die Seitenbahnbildung ersetzt. Bei der A. caudalis trat diese jedoch nicht mehr in Erscheinung.
10. Die Seitenbahn entstand bei obigen Experimenten in allen Fällen in folgender Weise : Das unterbrochene Blut floss in die Seiten-äste oder in die Kapillaren und durch diese fing es von der Peripherie zum Zentrum (Vene) oder von dein Zentrum zur Peripherie (Arterie) zu fliessen an. Anfangs floss das Blut auf mehreren Wegen. Allmählich aber wurde eine grössere Menge Blut durch einen bestimmten Weg getrieben, der verhältnismässig kurz ist. Die für diesen Weg benutzten Zweige oder Kapillaren entwickelten sich nach und nach zur Hauptbahn. Die Seiten bahn besitzt die gleiche Stärke wie die Hauptgefässe und stellt ganz und gar ein Stück des Hauptgefässes dar. Die einmal entstandene Seitenbahn besteht endgültig.
11. Ich eliminierte je ein Stück der Vasa caudalia bei Bufolarven von 8 nun Körperlänge. Die Wiederherstellung fand in folgender Weise statt : a. In den meisten Fällen entstanden die Seitenbahnen. b. In wenigen Fällen entstanden eine neue selbständige Vene und die Seiten-bahn der Arterie. Auch in diesem Stadium ist die SeitenSeiten-bahn meistens kurz, bei der Vene manchmal etwas länger.
12. Ein Stück der Muskelschichtregion von 8 mm langen Bufolarven mit dem Blutgefässe wurde elimiert. Dabei entstand ebenfalls eine Seitenbahn, die meistens in der Flosse oder in dein aus dein Meseiichym regenerierten Achsenteile verlief.
13. Ich eliminierte die A. caudalis total bei 24 mm langen Bufo-larven. Es zeigte sich niemals eine Wiederherstellung. Bei totaler Elimination der V. caudalis dagegen entstand eine neue Vene. Bei Rhacophoruslarven eliminierte ich die äussere Schwanzvene einer Seite . Das Blut wird in solchem Falle von der äusseren Schwanzvene der anderen Seite und von der V. caudalis aufgenommen. Es entsteht deshalb keine neue dicke Bahn ausserdem.
656 Tatsuo Fukai,
Bufolarven mit Vasa caudalia. Dabei entstand ein neues .selbständiges Blutgefäss. Die neugebildete Arterie verlief etwas gewunden in der Muskelschichtregion, die neugebildete Vene in der dorsalen Flosse oder den Oberrand der Muskelschicht entlang. In einigen Fällen entstand in der regenerierten ventralen Flosse eine Hauptvene.
15. Ich eliminierte ein Stück der V. jugul. ext. von 20 min langen Rhacophoruslarven. Der Blutkreislauf wurde einige Tage nach der Operation meist durch Bildung der Verbindungsbahn wieder hergestellt. Diese Verbindung geschah zwischen der V. jugul. ext: und der Vene eines anderen Körpergebietes (V. jugul. ext. der Gegenseite, V. facialis,
V. j ugul. iiit. oder Vene der Bauchhaut). In wenigen Fällen entstand aber die Seitenbahn.
16. In der Verbindungsbahn zwischen V. jugul. ext. und V. faci-alis fliesst das Blut umgekehrt. Es handelt sich um eine endgültig
dauernde Rückströmung des venösen Blutes.
17. Ich eliminierte ein Stück der V. jugul. ext. auf beiden Seiten voll Rhacophoruslarven (20 mm). Es entstand auf jeder Seite ebenfalls eine Verbindungsbahn oder Seitenbahn.
18. Der Entwicklungsprozess der Verbindungsbahn ist ganz ähnlich wie bei der Seitenbahnbildung.
19. Ein Stück der A. carot. ext. wurde hei 20 min langen Rhaco-phoruslarven eliminiert. Dabei wurde der Defekt durch die
Seitenbahn-bildung wieder aufgehoben.
20. Bei Bildung der Seitenbahn oder Verbindungsbahn sind die dickeren Äste leicht verwendbar, besonders bei Venen.
21. Aus den einzelnen Befunden schliesse ich wie folgt : Die Wiederherstellung der „ embryonalen Blutgefässe" findet durch rasche Bildung einer neuen Hauptbahn statt. Bei dieser Hauptbahn kann es sich um eine Seitenhahn, Verbindung,bahn oder um ein selbständiges
Blutgefäss handeln. Diese drei Möglichkeiten ergeben sich in Abhän-gigkeit von der Art des Blutgefässes, dein topischen Verhalten, der Eliminationsstelle, der Art der Elimination usw.. Der Entstehungsprozess dieser neuen Bahn ist von dem Entwicklungsstadium abhängig. In einem etwas späteren Stadium entsteht die neue Hauptbahn durch. rasche Umgestaltung der Zweige oder Kapillaren, die schon bei der Operation existierten. In einem möglichst frühen Stadium entsteht sie dadurch, dass zuerst die Kapillaren oder Zweige sich entwickeln und dann zur
Hauptbalin uingestalten. Jedenfalls ist die. Neubildung eines Hauptge-fässes im embryonalen Stadium ziemlich leicht ; hierbei spielen die
II. Transplantationsversuche mit den Organanlagen.
Einleitung.
Als zu den wichtigeren Untersuchungsmethoden gehörig, wurden schon verschiedene Transplantationsversuche in der Entwicklungsmechanik vorgenommen. Doch scheinen, trotz vieler Veröffentlichungen, Ver-suche, die eine Untersuchung der Entwicklung der Blutgefässe bezwecken, nur sehr selten gemacht worden zu sein.
In welcher Art und Weise finden die morphogenetischen Verän-derungen der Blutgefässe bei der Transplantation d. h. in einer abge-änderten Situation statt? Um diese Frage zu lösen, unternahm ich hier Transplantationsversuche mit den Organanlagen.
Ich wählte. die Schwanzknospe oder die Hinterbeinanlage für das Transplantat. In bezug auf die Transplantation der Schwanz- oder
Beinknospe finden sich schon viele ausgezeichnete Arbeiten veröffent-licht. Sonderbarerweise hat aber das Verhalten der Blutgefässe bei dieser Transplantation bisher wenig Beachtung gefunden und ist noch kaum geklärt, obwohl einige kurze Beschreibungen z. B. von Born, Braus,
Gräper, Weiss, Suzuki, Ogawa u. a. vorhanden sind. Darum habe ich meine Aufmerksamkeit bei diesem Experiment besonders der Ent-wicklung der Blutgefässe, die bei der Transplantation das Transplantat
versorgen sollen, zugewandt.
Eigene Versuche.
Als Material benutzte ich Amphibienlarven, besonders Bufo vulgaris
und Diemietylus pyrrhogaster. Die Operation geschah unter der
Bino-kularlupe mit Hilfe einer scharfen Nadel. Genauere Angaben folgen in
den einzelnen Kapiteln. Die Beobachtung wurde teils an lebenden
Präparaten, teils an Schnittpräparaten unter dem Mikroskop gemacht.
Fixierung in Formol (20%), Färbung mit Hämalaun und Einbetten in
Paraffin.
A. Homoioplastische Transplantation der Schwanzknospe.
Das Material liefert die Bufolarve. Bei der Operation betrug die ganze Länge der Larve ca. 4.5 mm und die Schwanzknospe 0.5-1 mm. Als Transplantationsstelle wählte ich Seitenbauch, Kiemengegend und Schwanzknospe. Ich habe so viel Ektoderm, als der Basis der
Schwanz-658 Tatsuo Fukai,
knospe entsprach, an der betreffenden Stelle entfernt und die Schwanz-knospe in diese ektodermlose Stelle eingefügt. Die operierten Larven wurden in einer mit Leitungswasser gefüllten Glasschale gezüchtet.
1. Die normale Entwicklung der Schwanzgefässe.
Was die Entwicklung der Schwanzgefässe von Amphibieularven anbetrifft, ist unsere Kenntnis noch als gering zu bezeichnen. Deshalb habe ich hier entsprechende Untersuchungen bei Bufolarven vorgenommen und den Entwicklungsprozess der Schwanzgefässe besonders in den früheren Stadien zu erklären versucht. Die Beobachtung geschah an Schnittpräparaten.
Bei Larven, deren Körperlänge ca. 4.5 mm (Schwanz 0.5-1 mm) betrug, wurden zwei Gefässanlagen d. h. die der Arterie und die der
Anlage der Schwanzge-fässe von Bufolarve. Aag Anlage der A. cand., Vag Anlage der V.
cand., X 50.
Vene in dem Achsengebiet und zwar unterhalb der Chorda nachgewiesen (Fig. 12). Die Arterien-anlage liegt direkt unter der Chorda und zeigt eine sehr dichte Anordnung, die Venenanlage be-findet sich weiter unter der Arterienanlage und ist sehr locker angeordnet.
Bei Larven, deren Körperlänge 5 mm (Schwanz 1-1.5 mm) betrug, erkannte man einige Lakunen an der Stelle, wo sonst die Venenanlage lokalisiert zu sein pflegte, doch war im locus der Arterien-anlage keine Lakune erweisbar.
Bei Larven von 5.5-6mm Körperlänge (Schwanz 1-1.5 mm) wurde die Lakune auch am Ort der Arterienanlage nachgewiesen, an dem der
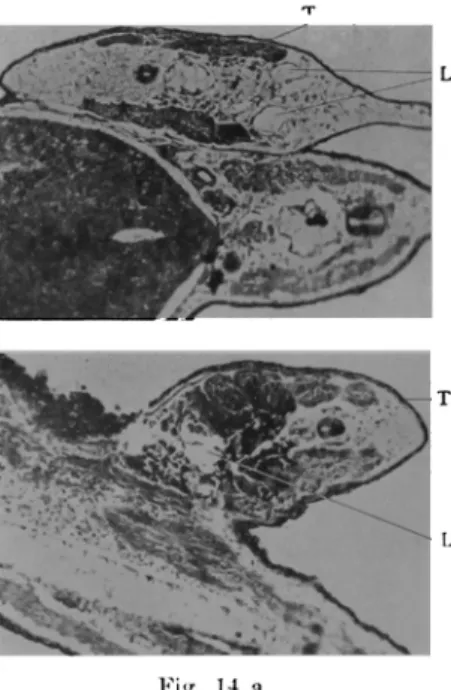
Venen-anlage beobachtete man eine Zunahme der Lakunen nach Zahl und Grösse (Fig. 13 A). Form und Grösse der Lakunen ist sehr mannigfaltig. Sie
entstehen im allgemeinen zuerst in der Wurzel des Schwanzes . In einigen Fiillen werden sie aber schon in verhältnismässig kaudaler Ge-gend nachgewiesen, während sie iii der Wurzel noch nicht erkennbar
sind. Die im locus der Arterienanlage entstandene Lakune ist im allgemeinen klein, gering und zirkumskript. Dagegen sind die Lakunen der Vene fast immer zahlreich, disseminiert und sehr mannigfaltig, was
ihre Grösse betrifft. Ein Zusammenhang zwischen Lakunen und Rumpf-gefässen wurde noch niemals festgestellt. Diese Lakunen enthalten kein
Blut.
Fig. 13.
Lakunenbildung im Schwanz von Bufolarve. L Lakunen,
x 50.
führen die oben erwähnten Lakunen Blut und verbinden sich mit dem Blutgefässsystem des Rumpfes (Fig. 13 B).
Obigen Befunden nach möchte ich bezüglich der Entwicklung der Schwanz-gefässe schliessen : Bei der Bufolarve entsteht das Schwanzgefäss zuerst in seiner Anlage an der bestimmten Stelle und dann entwickelt sich aus dieser Anlage die Lakuue. Bald verbinden sich die Lakunen miteinander und mit dem Zentrum, so dass zum ersten Mal der Blutkreislauf im Schwang auftritt. Mit anderen Worten handelt es sich um eine lokale und lakunäre Entwick-lung.
2. Transplantation in den Seitenbauch.
Bufolarven. Operation Anfang Mai. Länge der Larve ca. 4.5 mm bei der Operation. Am Tag nach der Operation war das Transplantat vollkommen eingeheilt.
Zunächst wurde der Entwicklungsprozess der Blutgefässe im Trans-plantat vor allein in den Anfangsstadien beobachtet. Am lebenden Material konnte ich erst am 5. Tag nach der Operation, wo die Larve ca. 14 mm lang (Schwanz 9 mm, Transplantat 4 mm) war, die Kapillar-netze in der Wurzel des Transplantates und zwar in der ventralen Flosse nachweisen. Bei dieser Larve schnitt ich das Transplantat in der Mitte ab, so dass das Blut in dein Achsengebiet und zwar entweder in zwei Strahlen (aus A. u. V. caudalis) oder nur in einem Strahl (aus A. caudalis) ausfloss. Am 7.-8. Tag nach der Operation konnte ich den Verlauf der Hauptgefässe des Transplantates an dem lebenden Material beobachten. Dabei betrug die Länge der Larve ca. 17 min (Schwanz 12 mm, Trans-plantat 7 mm).
35 Larven wurden nun in Schnittpräparaten beobachtet und folgende Befunde notiert.
Am Tag nach der Operation : Die Larvenlänge 8 mm (Schwanz 2 mm, Transplantat 2 mm). In dein Schwanz des Wirtes sind Blutge-fässe und Zirkulation nachweisbar. Im Achsengebiet des Transplantates sind zwei Gefässanlagen (Arterien- und Venenanlage) beobachtbar. Die