1.は じ め に
将棋や囲碁で人工知能がプロの人間に勝利したニュー スを聞くと,従来は不可能と思われていた人の高度な知 的活動に人工知能が使われる未来を想像せずにはいられ ない.そのような状況から,「物理学と人工知能」とい う組合せを聞くと,普通は人工知能を使って物理学の研 究をする内容を想像するのではないかと思う*1.例えば, 膨大な観測データから未知の普遍的物理法則を発見する ことなどである.一方,人工知能の研究に物理学の知見 が何らかの示唆を与える可能性を想像する人も少しはい るだろう.本記事では,後者の視点で,将来人工知能の 理論に役に立つかもしれない非線形物理学と関連する知 見を紹介したい.特に今日の人工知能ブームを牽引する 深層学習は,ニューラルネットワークという脳を参考に つくられた数理モデルを基礎としている.ニューラル ネットワークと物理学の関係は割と親密であり,最初に 少し過去の研究の話をしよう.その後,神経科学の最近 の知見や,それと関連しそうな数理モデルの話へつなげ たい. ただ,最初に断っておきたいことは,俯瞰した視点で 物理学全般と人工知能の関係を述べる能力は我々にはな い.物理学の中でも非線形物理の視点に基づいて,神経 科学の実験的知見も織り交ぜつつ,やや偏った私的バイ アスのある記事になる点はご了承願いたい.具体的には, 大自由度の非線形力学系としてニューラルネットワーク を捉え,その知見を神経科学の実験の解釈にも生かす試 みである*2.2.ニューラルネットワークの数理
脳の情報処理を主として担っているのは神経細胞 (ニューロン)のネットワークであり,シナプス結合を 介して電気的信号をやり取りすることで高度な情報処理 を実現していると考えられている.そこからヒントを 得て,ニューロンのダイナミクスを単純化した素子を, ネットワーク状に結合して情報処理するモデルが提案さ れた.深層学習で用いられるニューラルネットワークも, 本質的には同じものである.ここでニューロン i の状態 を実数 yi(典型的な場合として発火・非発火に対応させ た±1 の 2 値に制限する場合もある)で表すと,その値 は以下の式で決定される. yi= f(ui),ui= wij xj j これは,ニューロン xjを入力層,ニューロン yiを出力 層とする 2 層のフィードフォワードネットワーク(パー セプトロン)と呼ばれるものである [Rosenblatt 58].こ こで,wijはニューロン i へ入力しているニューロン j の 状態 xjに対する結合の重みである.これをさらに,多層非線形物理学から見た
ニューラルネットワークの学習
Learning Mechanism of Neural Networks from the Perspective of
Nonlinear Dynamics
伊藤 浩之
京都産業大学情報理工学部Hiroyuki Ito Faculty of Information Science and Engineering, Kyoto Sangyo University.
[email protected], https://www.kyoto-su.ac.jp/faculty/professors/cse/ito-hiroyuki.html
青柳 富誌生
京都大学大学院情報学研究科Toshio Aoyagi Graduate School of Informatics, Kyoto University.
[email protected], http://www-np.acs.i.kyoto-u.ac.jp
Keywords:
dynamical system, nonlinear dynamics, attractor, reservoir computing, intrinsic manifold. 「物理学と AI」 *1 SF 好きの方は,人工知能が物理学の研究を自律的に行う未来 を想像するかもしれない.スタートレックのデータ少佐の晩年 のように. *2 その際,特に神経科学の知見の解釈に関しては,大胆な私的 推測も述べることになるが,学問的事実と推測は区別して記述 したつもりである.にしたマルチレイヤニューラルネットワーク(多層パー セプトロン)は深層学習で用いられている基本構造の一 つであり,以下の式で表される [Amari 67, Rumelhart 86].l 層のニューロン i の状態を xilと表すと
⎝
⎛
⎝
⎛
xil=f wijl-1 xjl-1 j のように l 層の出力は l−1 層のニューロンの状態から 決定される(図 1:左上).L 層のネットワークの場合, xi0は入力層に,xiLは出力層に対応する.多層にするメ リットとしては,ややおおざっぱな言い方をすれば,層 内のニューロン数より層自体の数を増やすほうが指数 関数的に複雑な入出力関係を学習可能である点にある [Montufar 14].言い換えると,適切な結合 wijlを学習に より獲得すれば,かなり複雑なことも実現できる可能性 がある.現実にはその学習が難しいわけであるが,深層 学習ではさまざまな工夫が功を奏して華々しい成果を上 げていると考えられる. しかし,層構造であるマルチレイヤニューラルネット ワークは,極論すれば,入力を決めると出力が決まる関 数であり,音声などの時系列処理などは難しい.例えば, 過去の履歴を参照しながら情報処理をする必要がある場 合,どこまでの履歴を情報として使うかは,そのときど きの状況に依存し,単純に時間長を固定した信号を空間 にマップして多層ニューラルネットワークで処理すれば よいというわけではない.これは,過去の情報が然るべ きタイミングで活用可能な記憶のメカニズムが必要であ ることを意味する.層状構造のネットワークは一度使っ た情報は次の層へ伝わるだけで,後で参照することはで きない.過去を必要に応じて参照可能な柔軟な記憶の機 能を備えるには,必然的に自分自身に信号を戻すフィー ドバックが必要であり,リカレントニューラネットワー クを考えることになる. 2・1 リカレント結合とアトラクタの出現 ここではまず最もシンプルなリカレントニューラル ネットワーク,すなわち,全ニューロンが相互に結合し ている以下の式に従うモデルを考えよう*3.⎝
⎛
x(ti +1)=f⎝
⎛
wij x(t)j j (1) 図 1 左下に示すように,多層構造と異なる点は,同 じ結合 wijによりニューロンの状態 x(t)が上式に従i い,繰り返し新たな状態 x(t+1)に更新される点であi る.この t を時間と見れば,次の時刻の状態 x(t+1)が, i 現在の時刻の状態 x(t)により決定される力学系の一種i と考えられる*4.初期の状態を与えれば,その後のネッ トワークの状態は自律的に時間発展する系である.マル チレイヤニューラルネットワークでは最終層の出力が結 果であるが,リカレントニューラルネットワークでは何 を出力結果と考えるべきであろうか? 話を単純にするため,初期時刻 t = 0 に適当な状態 x(t=0)を与え,その後に式(1)に従って時間発展さi せた場合を考えよう.層構造と異なる点は,繰り返し同 じ結合 wijにより状態が更新されることにある.その結 果,結合の性質がダイナミクスの結果に非線形効果とし て強く表れる.N 個のニューロンの状態 x(t)を N 次元i 空間内 X=(x1, x2, …, xN)の座標で表現しよう(状態空 間).リカレントニューラルネットワークの状態の変化 は状態空間 X 内を経めぐる軌道として表現できる.例え ば,結合を対称 wij= wjiと仮定すると,初期状態に応 じて行き先が異なる定常状態に収束させることが実現で *3 深層学習では学習がうまくいくように LSTM や GRU などの 構造化されたリカレント結合を考えている. *4 時間を連続にとれば dx(t)/dt が xi (t)により決まる微分方程i 式で表される力学系となる. ֪ҔȌǺÓȮ СݳLJЏ ֳƶ֪Ҕीઙ0
ऻඦƶ໙ǔ ऻඦƶ໙ǔ ǦǥǴǝȃȤǪǺ ǝȃȤǪǺ ȤǟȮǝȃȤǪǺ0
0
ǝȃȤǪǺƶࢽ۔ ȥǦȧȮȃȆȠÓȤȦ ȈǾȃȪÓǪ șȦǼȧǟȟ ȆȠÓȤȦȈǾȃȪÓǪ a b領域B 領域A ༫੶૮ƶੀ֬ 図 1 左:多層パーセプトロンとリカレントネットワークの情報の流れ.右上:状態空間で見た 点アトラクタと連想記憶モデルの関係.右下:さまざまなアトラクタきる.これは,情報処理の観点で見ると,領域 A にある 多様な状態をある 1 点 a に,領域 B にある別の多様な 状態を別の 1 点 b に対応付ける,一種の判断・識別を行っ ているともみなせる(図 1 右中)*5.十分時間が経過し た後のこの収束先の定常状態のことを,状態空間の 1 点 であることから点アトラクタと呼ぶ.リカレント結合に より,多様な状態を 1 点に対応付けるアトラクタと,初 期条件に応じて収束先が異なる非線形的な多重安定性が 出現したといえる. 2・2 連想記憶モデルと点アトラクタ Hopfieldがスピン系との対応で提唱した連想記憶モデ ルでは,初期に適当な入力パターン x(t=0)を与え,i その初期条件からネットワークを時間発展させて十分時 間が経ったときの x(t)の状態を出力と考えた [Amari i 72, Hopfield 82].結合 wijは,一種の相関学習である Hebbの学習則 wij= μ=1 P ξiμξjμ 1 N に従い構成する [Hebb 49].ここでξμiは,学習すべき P 個のパターンの中でμ番目のパターンにおいてニューロ ン i のとるべき状態(例えば ξiμ=±1)である.結合は 対称 wij= wjiであり,ここから時間的に定常な状態へ 収束することが保証される.初期状態として,あらかじ め学習させておいたパターンに近い状態を与える.する とネットワークの状態が式(1)に従って更新され,最 終的に学習したパターンに近いものに収束する.このモ デルでは,初期に与えたパターンから,あらかじめ学 習したいくつかの典型的パターンの一つに収束する過程 を,刺激から連想して記憶を想起する過程とみなす. この系には式(1)の更新ダイナミクスにより必ず減 少する(正確には非増加)マクロな量(磁石のシンプル なモデルであるスピン系のエネルギーに対応する) E(t)= wijx(t)xi (t)j i 1 2 が存在する.パターンの空間を横軸に,縦軸に E(t)を 表示して,想起過程のダイナミクスを概念的に表すと図 1 右上のようになる.Hebb 学習則により,与えられたパ ターンとほぼ同じ学習パターンが極小値に,想起可能な 初期刺激のパターンの範囲が谷の大きさに対応する.一 方で,記憶パターン数 P が増えるにつれて,学習すべ きパターンに関係がない極小値も出現する(偽の記憶状 態).P がさらに増加すると,この偽の記憶状態の極小 値が支配的となり,やがて学習パターンが不安定化する (スピングラス状態).この場合は,学習したパターンと 全く無関係な状態に系が収束する.このモデルは統計力 学を応用することで記憶容量の導出などのさまざまな 理論解析の結果が得られ,多数の物理学者がニューラル ネットワークの研究に参入するきっかけとなった. 非線形力学系の視点で見ると,式(1)によるダイナミ クスは離散時間の散逸力学系の一種であり,E(t)はリャ プノフ関数と呼ばれる.この関数が存在することで,十 分時間が経過したときの系の振舞いは,時間的に定常な あるパターンに収束することが保証される.E(t)を地 面の高低とみなせば,ちょうど水が局所的に低い場所へ 流れ落ちるイメージである.連想記憶モデルは,非線形 力学系における点アトラクタに記憶パターンを対応させ た情報処理のモデルといえる. 2・3 さまざまなアトラクタと情報処理 しかしながら,現実の応用や神経科学の実験事実を 考えれば,時間的に定常な状態に収束するだけでは拡張 性に乏しい.例えば,サッケードなどで視線の移動量を 一時的に記憶するため,連続的な量を符号化することを 考えよう.点アトラクタを多数用意して近似する方法は すぐに思いつくが,いかにも頭の悪い方法である.そこ で,収束先のアトラクタを点アトラクタから拡張する試 みも考えられている [Brody 03].代表的な例としては, ラインアトラクタ*6やカオスアトラクタなどである.ラ インアトラクタとは初期条件に応じてライン上の一点に 系が収束し,その値を出力とするもので,連続値の情報 を状況に応じて得ることが可能となる [Seung 00].一方 で,ノイズや系の不均一性に対して,どのように安定的 にラインアトラクタを構成するべきかは難しく,ヒステ リシスのある素子を活用するなど,さまざまな工夫があ る [Koulakov 02].カオスアトラクタは,嗅覚の情報処 理に関係しているとの実験結果や,そのフラクタル構造 などにエピソード記憶をコードするのに適しているとの 報告もある [Skarda 87, Yamaguti 11]. これらの研究では,さまざまな情報を柔軟に処理する ため,収束先のアトラクタを工夫して対応しようという 方策を用いている.一方,アトラクタへの収束に至る過 程や,刺激入力がない自律的ダイナミクスについては, あまり考察されていない.最近になって神経科学の実験 データの解析に力学系的視点が取り入れられるようにな り,次章で示すような興味深い発見が報告されている.
3.神経科学からの知見
3・1 神経ネットワークの幾何学的構造への関心 深層学習によりリバイバルしているニューラルネッ トワークの研究においては,高次元の自由度をもつ入力 データの集団に対して,比較的少数の出力層細胞の活動 で表現されるラベルとの対応関係を統計的に再現するた *5 時間や状態は離散的であるが,本質は変わらないので見やす さのため連続の場合のアトラクタを表示している. *6 角度などに対しては一周して戻るのでリングアトラクタ,さらに一般的化した連続アトラクタなどもある.めには,どのような情報圧縮を行うネットワーク構造を 構築すべきかが議論される.ニューラルネットワークの 本質は甘利が黎明期に定式化しているように [甘利 68], 入力データの自由度が張るパターン信号空間から,特徴 空間への射影を経由して,最終的に識別空間に至る写像 の幾何学的特徴付けである.ブラックボックス化してい る深層学習の内部構造に関して,写像の幾何学的構造の 解明がどの程度可能であるのかの探求は今後の重要な研 究課題であると考える. 一方,神経科学の分野でも脳神経ネットワークでの 学習における幾何学的構造に近年関心が集まっている. 1980年代初頭までの神経生理学実験では,技術的制約か ら同時に記録できる細胞数が 1 ∼ 2 個と限られていたた めに,外部刺激に対する脳の反応は個々の細胞の活動度 (発火率)の変化を中心として研究が行われていた.80 年代後半以降の測定技術の発展(多細胞活動同時記録法, 各種光学的イメージング技術など)は数十∼ 100 個程度 の細胞活動の同時記録を可能とし,外部刺激に対する反 応や運動出力という情報処理を複数細胞が構成するネッ トワークの活動として解析することが可能となった.複 数細胞の活動データは集団符号化(ポピュレーション コーディング)や同期・相関発火による細胞集団のダイ ナミックな再組織化によるバインディング問題の解決策 などの新たな情報符号化モデルの実験的検証を可能とし た [Aertsen 02, 伊藤 97, 伊藤 00, 櫻井 08].ここでは神 経ネットワークにおける細胞活動間の相関構造によって 幾何学的拘束が生じる事情を力学系の視点から説明し, Brain-Machine Interfaceでの学習に関する最近の研究 を紹介する.これらの現象は脳の神経ネットワークの活 動が単なるフィードフォワードではなく,よりダイナ ミックで力学的な描像が必要であることを示唆している. 3・2 試行間変動性と自発発火活動 神経生理実験において古くから知られながら,現在に 至ってもその機能的意味が十分に解明されていない現象 に,刺激誘発活動の試行間変動性と自発発火活動が存在 する.古典的な脳の情報処理の描像では,外部からの刺 激入力がなく,身体運動を行っていない安静状態では, 脳では何の情報処理も行われておらず,皮質の自発的な 活動は単なる無意味なノイズに過ぎないと考えられてい た.一方,刺激入力や身体運動が行われると皮質細胞は 情報処理に特異的でかつ信頼性のある活動を生じるた め,細胞活動を詳細に調べることにより,どのような情 報処理に関与しているのかを明確に同定することが可能 であるというパラダイムのもとに研究が行われてきた. 現在の神経科学では,これらの描像は複数の点で正しく ないことが認識されている.神経ネットワークにおける 細胞活動間の相関構造による幾何学的拘束と学習の議論 の前提として,まず試行間変動性と自発発火活動という 二つの現象を説明する. § 1 試行間変動性と発火数相関 まず,第一に刺激入力や身体運動に関与する個々の皮 質細胞の活動は十分な信頼性を有していない.例えば, よく知られているように視覚皮質の各細胞はその受容野 内に提示される線刺激の傾きに対して固有の方位選択性 の活動を示すが,同じ線刺激を提示する試行を複数回 繰り返しても発火数は大きくばらつくことが知られてい る.これは試行間変動性(trial-to-trial variabilities)と 呼ばれる.信号伝達で生じる一定の大きさの確率的な揺 らぎであれば,平均発火率自体が大きくなれば SN 比の 上昇が期待できるが,実験データでは試行間にわたる発 火数の分散は平均発火率に比例して増大することから, 活動を生じさせる機構と変動性の機構が同一であると考 えられている.つまり,神経ネットワークのダイナミク スという力学系固有の現象である可能性がある.この変 動性のために,データ解析では同じ刺激提示下の複数試 行にわたる平均発火率(時間平均)を用いて方位選択性 などの細胞の機能を議論している.しかし,我々の実際 の対象認識では 1 試行の刺激提示で十分であるため,信 頼性を回復する何らかの神経メカニズムが存在するはず である.この解決策として集団符号化(ポピュレーショ ンコーディング)が提案されている.これは,同一また は十分な冗長性をもって方位選択性特性を符号化する複 数の細胞が存在し,これらが統計的に独立な変動性を示 せば,1 試行の反応の細胞集団にわたる平均(アンサン ブル平均)は細胞数の平方根の逆数で標準偏差が減少す ることで信頼性が向上し(大数の法則),一つの細胞で の活動の試行平均と一致するという論理である(ニュー ロンエルゴード仮説とも呼ばれる).しかし,神経生理 実験で近接した複数の皮質細胞の活動を記録して,発火 数の細胞間相関を計算すると,独立ではなく,小さい ながらも有限の相関係数(正も負も存在する)を示すこ とがよく知られている.この相関は発火数相関(spike count correlation)と一般的に呼ばれるが,ノイズ相関 (noise correlation)という誤解を招く名称でも呼ばれる ことがある.有限の発火数相関の存在のために,集団符 号化による信頼性の向上は必ずしも単純ではないことが 認識されている [三浦 11].試行間変動性の機能的意味 に関しては現在でも統一した見解に至っておらず,活発 に研究が行われている.試行間変動性および弱い発火数 相関は情報処理にとって無益または有害な副産物である のか,それとも有効な方策であるのかすら確定していな いのが現状である.近年の研究の関心は,細胞間の発火 数相関によるネットワークの幾何学的拘束(内因性多様 体)およびその構造が Brain-Machine Interface での適 応的学習に与える影響である.この特性は深層学習にお けるネットワーク構造と学習の関係に大きく関連すると 思われるため,本概説のメインテーマとして,後に議論 を行う.
§ 2 自発発火活動と内因性多様体 D. Hebbは 1949 年に発表した著作 Organization of Behavior [Hebb 49]において今日の神経生理学での重 要な研究対象となっているテーマの多くを導入してい る.セルアセンブリ [伊藤 15] や Hebb シナプスが良く 知られているが,現在でも機能的意味が解明されていな いもう一つの現象である自発発火活動の重要性も明確 に指摘している.Hebb は末梢の感覚器からの情報が中 枢に一方的に伝搬するという当時の支配的な描像に疑 問を抱き,神経系に存在する自発発火活動を“existent excitation”と表現し,「脳は常にアクティブであり,感 覚入力で誘発される活動は自発活動に影響されるはずで ある」と議論している.その後の Hubel と Wiesel らの 機能的に特殊化した細胞の発見がもたらした研究の支配 的な流れの中で,長期にわたって自発活動に対する関心 は薄れていた.しかし,80 年代後半からの Arieli らの イスラエルグループによる光感受性色素による視覚皮質 の時空間活動パターンの光学的イメージングの実験報告 により自発発火活動に対する認識は一変した.格子(グ レーティング)刺激提示下では,格子の方位を最適刺激 とする方位選択性コラム(共通な特性を示す細胞の空間 的集まり)の活動がイメージングされるが,無刺激条件 下での自発活動の空間パターンは,刺激下での方位選択 性コラムの活動と類似した空間パターンに停留する傾向 があり,異なる方位選択性コラムの空間パターンを時間 的に遍歴しているという現象が報告された [Kenet 03]. この実験報告は,自発発火活動や皮質活動での内因性 (intrinsic)な相関構造に着目する今日の研究テーマの 根源となっている. さらに,Luczak らは麻酔下および覚醒下でのラット の聴覚皮質から記録された複数細胞のスパイク発火活動 データの解析から同様な結果を報告した [Luczak 09]. 細胞集団符号化の解析においては,各細胞の発火数を座 標軸とすることで,記録する細胞数の次元の位相空間内 の一点として一試行での細胞集団の活動を表示する.従 来の理解では,自発発火活動は構造がない雑音のような 存在であると考えられていたので,細胞集団活動は位相 空間内において,発火数の生理学的上限の範囲内にお いては一様に分布すると予想される(図 2A).しかし, Luczakらがラットの聴覚皮質から記録した自発発火活 動は細胞間の発火数相関のために,位相空間内の限定さ れた領域に拘束されていた(図 2B). 後で説明する Sadtler らにならって,この限定された 領域を内因性多様体(intrinsic manifold)と呼ぶことに する*7.さらに Luczak らは,音刺激に対する細胞集団 活動は自発発火活動が拘束される内因性多様体から逸脱 するのではなく,内因性多様体の部分集合となっている ことを報告している.異なる音刺激に対する細胞集団活 動は試行間変動性から広がりをもった異なるクラスタを 形成するが,すべてのクラスタは内因性多様体の部分集 合となっている(図 2B).これは,自発発火活動であれ, 刺激誘発活動であれ,神経ネットワークで実現され得る 活動パターンは内因性多様体上に限定されているという 描像である. この拘束は当然ながら神経ネットワークを構成する細 胞間の解剖学的結合から生じている.しかし,ここで注 意すべき点は,細胞間の発火数相関は解剖学的結合によ り 1 対 1 に決定されるものではなく,複数細胞の活動 状態に依存する力学的変数であるということである.実 際に麻酔下ネコの視覚皮質から同時記録された細胞間の 発火数相関は提示する刺激の方位により変動する(相関 の符号すら反転する)場合があることが報告されている [Maruyama 13].皮質細胞は,複数の細胞から弱い入力 を受けている分散処理系であることが知られており,一 つのシナプス前細胞からの興奮性入力だけではいくら高 頻度入力でもしきい値を超えて発火することはできな い*8.発火のためには複数の細胞から,数ミリ秒程度の 短時間に同期した入力が必要となり(同期検出器),こ の特性のために細胞間の発火数相関は 2 体問題ではな く,多体問題となる*9.解析における困難は,多体問題 に影響を与えている細胞活動の多くを実験では記録でき ていないために,これらの細胞活動状態は隠れた変数と して扱わざるを得ないということである.Aertsen らは 解剖学的な結合(anatomical connection)と区別するた めに,スパイク活動の相関に基づく関係性を機能的結合 (effective connectivity または functional connectivity)
と定義した [Aertsen 02].つまり,内因性多様体自体も, 図 2 位相空間内での細胞集団活動(概念図) A. 同時記録された 2 個の細胞の発火数の分布. 各点は一つの試行での状態を示す.B. 実際の細 胞記録では,細胞間の発火数相関のために自発 発火の状態は相空間内で限定されている(内因 性多様体).異なる刺激 A,B に対する活動を〇 と★で示している(adapted from [Luczak 09])
*7 多様体という数学的な名称が付けられているが,高次元空間 で変数間に課された拘束条件から有限な領域に閉じ込められる という程度の意味であり,その実体の数学的定式化は今後の課 題である. *8 解剖学的結合強度の分布が log ノーマルであるという報告も あり,一部の強い結合に発火は支配されるという説もある. *9 人工ニューラルネットワークにおいても同様な多体性は生じ ている.しかし,各層の状態をフィードフォワードで同期して アップデートしていくため,処理ステップとしての離散時間は 存在するが,ここで議論する力学的な連続時間は存在しない.
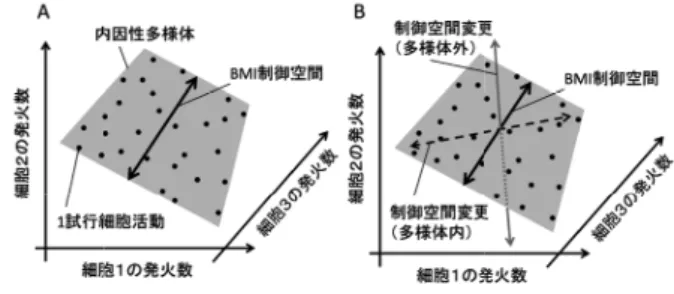
神経ネットワークでの複数細胞の活動相互作用を通じて 生じる力学的な状態であると考えられる. 3・3 内因性多様体と適応学習 神経ネットワークのダイナミクスで実現可能な状態が 内因性多様体上に限定されていることは細胞集団活動の 適応的変化・学習にも影響を与えることが予想される. Sadtlerら [Sadtler 14] は,覚醒サルの運動皮質から記 録される細胞集団を用いた Brain-Machine Interface (BMI)において,この影響を確認した.BMI はこの 20 年ほどで大きく発展を遂げた神経科学と工学との融合分 野である [Lebedev 17, Rao 13].基本的な方法論は,脳 内で同時記録された細胞集団活動から情報をコンピュー タが復号化し,筋肉などの身体機能を介さずに,直接に 外界のデバイスを制御することである.最も研究が進ん でいるのは,運動皮質の数十∼ 100 個程度の細胞の集 団活動によるモニタ上のカーソル(念力カーソル)やロ ボットアームの制御である.Sadtler らは運動皮質から 記録される細胞集団における細胞間の発火数相関から内 因性多様体を同定した.BMI では内因性多様体上に部 分空間(BMI 制御空間)を設定し,この空間内の細胞 活動状態とモニタ上のカーソルの運動を対応付けた.図 3Aの概念図では,三つの細胞の活動の内因性多様体を 平面で表し,その上に一次元部分空間を設定し,この線 上の位置とカーソルの運動との間に単調な関係を設定し たモデルを課した.この線上の状態は,細胞間の特定な 発火数相関の拘束下で各細胞の発火率を変化させること に対応する.例えば,A,B,C の三つの細胞間に A と Bは正,A と C は負の発火数相関が存在する場合には, Aの発火数の増加は B と C の発火数の増加と減少を伴 う.BMI 課題では,サルは細胞集団活動を用いてカー ソルをモニタ上のターゲットまで移動させることで報酬 がもらえる.ある程度の過渡的な状態の後に,サルは課 されたモデルに適応して細胞集団活動を制御し,適切に カーソルを制御できるようになる(ブレインコントロー ルモード). BMI研究での細胞集団活動の適応的変化における根 本的な問題は,サルは脳に存在する膨大な数の細胞の うちのどれが BMI で課されたモデルに関係しているの か,およびこれらの細胞の活動がカーソルの運動とどの ように関係付けられているのかを試行錯誤で見つけださ なくてはいけないということである.ミクロな細胞集 団の活動の変化と外部のカーソルの運動との因果関係の フィードバックを手掛かりとしていることは明らかであ るが,膨大な自由度での最適状態探索問題となるため, 多くの学習時間を要することが予想される(次元の呪い [Watanabe 17])*10.しかし,多くの BMI 実験の報告で は短時間でカーソルを制御できるようになることが報告 されている(1 時間∼数時間程度).これは,細胞集団 の活動が発火数相関による拘束により内因性多様体上に 限定されることで,最適状態探索問題が比較的少数自由 度に落ちていることが反映しているのではないかと考え られている. Sadtlerらの実験では,サルの脳活動が課せられて いる BMI モデルに適応した後にモデルを変更すること で,新たなモデルへの再適応のダイナミクスを観察し ている.モデルの変更には内因性多様体上の異なる部分 空間内の細胞集団活動を設定する多様体内変更(within-manifold perturbation)と内因性多様体から逸脱した 制御空間を設定する多様体外変更(outside-manifold perturbation)の 2 ケースが存在する(図 3B).彼らの 報告では,多様体内変更の場合は既存のネットワークの 結合構造で実現可能な細胞集団の相関活動への変更であ るため,短時間で再適応し,正答率も上昇する.一方, 多様体外変更の場合は,再適応のためにはネットワーク の解剖学的結合の変化による内因性多様体自体の変形を 行って,課された細胞集団の発火数相関を実現する必要 がある.このため,短時間での適応的変化は不可能であ り,正答率は低下したままであった. この概説では紹介しなかったが,fMRI などを用いた ヒトの脳機能イメージングでも,領域間の関係性におい て文脈依存的な相関結合(力学変数)としての effective connectivityや functional connectivity が議論されてお り,神経生理学と同様な視点での研究が活性化している [Friston 11].また,環境に対して積極的な情報処理を 行っていない安静状態でも特徴的な脳活動のダイナミク スが存在することは,脳機能イメージングでも報告され ている.Effective connectivity をもつ複数の領野間がダ イナミックなネットワークを構成し,複数の異なるネッ トワーク(resting state ネットワーク,default mode ネッ
図 3 内因性多様体と BMI 制御空間への適応(概念図) A. 同時記録された 3 個の細胞の発火数の分布.各点は一つ の試行での状態を示す.細胞間の発火数相関のために細胞 活動は相空間内で内因性多様体上に限定されている.図の 例では,BMI 制御空間として一次元軸を取り,この軸上の 位置に応じてカーソル運動を制御する.B. 設定した BMI 制 御空間に適応した後に,制御空間の変更を行い,細胞活動 の再適応を生じさせる.制御空間の変更には内因性多様体 上の異なる部分空間を設定する場合(多様体内変更)と内 因性多様体上から逸脱して制御空間を設定する場合(多様 体外変更)が存在する(adapted from [Sadtler 14])
*10 ミクロな要素の状態とマクロな出力との関係の適応学習と考 えれば,入力から出力までが多階層でつながる強化深層学習と 類似しているとも考えられる.
トワークなど)の存在が報告されている.
4.計算資源としての非線形力学系
ここで,再びニューラルネットワークの数理モデルの 話に戻ろう.前章の神経科学の知見では,特別なタスク を遂行していないときの神経活動(自発発火活動)につ いて,学習の能力や機能と関係していることを示唆する 実験結果を紹介した.ところで,工学的なニューラルネッ トワークにおいて特別なタスクをしていないときの自発 活動を考える意味はあるのであろうか? そもそも,神 経活動はエネルギーをかなり消費するので,不必要な活 動は進化論的に淘汰されて然るべきである.しかし,前 章で見たように現実には自発発火活動は存在し,雑音の ようなランダムで無意味な活動ではないとの実験的示唆 がある.最近では,内因性多様体やデフォルトモードネッ トワークの研究が盛んに行われている*11.このような自 発活動は,最適なシステムという工学的観点からは一見 むだに思える.それを覆す可能性があるという意味で興 味深いモデルであるレザバー計算機を最後に紹介したい. 4・1 レザバー計算機 知性の基盤である脳・神経系の情報処理には,学習に 時間がかかるが高度な判断や識別を行うもの*12と,ス キップなどの運動学習や環境適応など素早く学習が可能 なものがある.後者の素早い学習のモデルとして,リカ レントニューラルネットワークの一種であるレザバー計 算機(Reservoir computing)が注目されており,脳の運 動制御信号生成のモデルなどに用いられている [Laje 13, Sussillo 09].その源流は,echo state network [Jaeger 04] や liquid-state machine [Maass 02] であるが,二つの モデルの本質は同じであり,最近ではレザバー計算機 (Reservoir computing)と総称されている [Lukoševicˇius12].図 4(a)に示すように,そのアイディアの骨子は 以下に集約される. 1.レザバーと呼ばれるランダム結合したリカレント ニューラルネットワーク(大自由度の結合力学系で もよい)を用意する. 2.レザバーの全ニューロンの状態の線形和を出力と する. 3.目的の時系列を再現するように出力とレザバー間 の線形結合のみを学習する. 図 4(a)のように,周囲の環境の状態依存の入力や 出力依存のフィードバックを加えることで,複数の時系 列をより柔軟に学習することも可能になるが,必須では ない.第 1 のポイントは,系に何も入力がなくタスクを 遂行していないときでも,系は自律的に状態を更新して 常に変化している点である.第 2 のポイントはレザバー 内の結合は固定であり,タスクに依存した学習による更 新は行わない点である.学習は出力に関係する線形和の 結合のみ行う.また,出力はタスクに応じて複数あって もよい.レザバーが十分に大自由度で,各力学系の要素 の状態が多様なダイナミクスを示す場合,任意の有限の 時系列を線形和で精度良く再現でき,結合の学習も線形 計算で素早く完了するのが利点である.この話は神経系 に限らず,多様なダイナミクスを示す大自由度力学系で あれば現実の物理システムをレザバーとして活用でき る.もちろん,これらが実現するためには,レザバーに は望ましい性質がなくてはならない.その性質の条件は 以下の 2 点である. 1.時空間的に十分多様なダイナミクスを示す(多様性: カオスなど). 2.ほぼ同じ状況下で再現性が高い(安定性). この 2 点は矛盾する要請である.再現性だけ考えれ ば,周期解などの安定解が複数存在するレザバーが,ノ イズなどにロバストな再現性があるので望ましい.ただ し,生成可能な時系列の多様性は期待できない.一方で 複雑な時系列などを再現する場合は,多様な信号が生成 できるカオスが望ましい.ただ,カオスの性質上,微 少な差が指数関数的に増大して,ノイズなどにロバス トでない点が問題である.従来の研究では,カオスの 縁(edge of chaos)の状態が最も計算能力が高いとされ ている [Bertschinger 04].これを実現するのは難しそう に思うかもしれないが,レザバー内の結合を分散をコン トロールしたガウス分布でランダムにサンプルすること で,カオスの縁に系を設定できることが理論的に知られ ている [Sompolinsky 88].また,類似の指導原理として レザバー内の結合行列のスペクトル半径が 1 程度という 条件(echo state property)も広く用いられる*13.実際,
状況依存の入力 多様な時系列を素早く学習 計算能力 カオス 通常のアトラクタ Edge of chaos (a) (c) (b) フィードバック レザバーとして望ましい性質 レザバー:ネットワーク結合力学系 図 4 (a)レザバー計算機の原理.学習は出力への結合のみ行う, (b)最適なレザバーの条件,(c)情報量最大化の結果出現し たレザバーの結合構造.上位 2.6%の強い結合を表示している *11 特に何もしていないぼーっとしている時間が,実は創造的活 動には重要かもしれない,という我々の内なる声も,自発発火 活動の研究を後押ししているかもしれない. *12 この実現を目指した深層学習では,学習に熟練した技術と多 大な時間が必要である(手軽なパッケージ化したソフトの裏で). *13 おおざっぱには,時間が経過しても状態が発散せず,一方です ぐに信号が減衰せず過去の履歴が残る臨界状態を意図している.
このような条件のもとで,さまざまな事例でレザバー計 算機がうまく機能することが多数報告されている.なお, レザバー計算機はなぜ学習がうまくいくのか? 既存の 手法と比較した理論的研究として,信号を高次元の時空 間パターンに変換して線形分離を可能にするカーネル法 との類似性を指摘した研究もある [Hermans 12]. 4・2 力学系のレザバーとしての最適性 カオスの縁という状態を実現するネットワーク構造は 唯一ではなく,多数の状態があるだろう.その中のネッ トワーク構造中には,レザバーとしての優劣もあるであ ろう.レザバー内のリカレント結合は,特定のタスクに 特化した学習を必要としないことが利点ではある.しか し,もう少し欲張って結合構造を絞ることでさらに性能 を上げることはできないだろうか? その一つの可能性として,最後に我々の最近の研究を 紹介したい.ニューラルネットワークの活動の前後の状 態間の相互情報量が最大となるような結合構造は,神経 科学の実験データを統一的に説明可能であることが示唆 されている(Recurrent Infomax)[Tanaka 09].過去 の情報を記憶していないと解けない課題は,過去の入力 をできるだけ長時間蓄えられる能力が性能向上に直結す る.そこで,Recurrent Infomax の原理で生成したニュー ラルネットワークをレザバーに用いて,その性能向上を 図る方策が考えられる* 14.結果として,過去の情報を 必要とする課題に関して性能の向上が見られたが,記憶 より非線形性が必要である課題では,それほど向上は見 られなかった [Iwade 18].レザバー内の結合を最適化す る手法はすでに教師信号の有無なども含めさまざまな試 みが行われている [Lukoševicˇius 12].しかし,特に内部 結合を教師あり学習する場合は,レザバー本来の利点が 失われるジレンマもある.今後は,その点も勘案しつつ 他のレザバー設計の指導原理を探る力学系の研究が重要 に思える.
5.お わ り に
近年の神経科学では脳が固有(endogenous,intrinsic) にもっている力学的な制約による構造を前提として,感 覚刺激入力への反応や運動出力に関する脳活動を再検討 する動きが生じている.脳の固有のダイナミクスと外界 からの入力との相互作用を通じて認知が立ち上がるとい う「認知のインタラクション仮説」の実験的検証へと向 かうように思われる.個人的な見解としては,「無は無 限」とする禅の思想や「刺激のゲシュタルトは有機体そ のものによって,つまり有機体が自らを外の作用に差し 出す固有の仕方によって,創造されるのである」と主張 したメルロ=ポンティの哲学 [メルロ=ポンティ 42] に 通じるものを感じる.また,脳がネットワークのダイナ ミクスを固有の自己拘束で構造化することで,学習にお ける不良設定問題を解決しているとすれば,いわゆるベ イズ統計における事前分布に相当するものを,非線形力 学系の内部ダイナミクスを起点として構築しているよう にも思われる.このような視点を数理モデルの一般論に もち込むことは容易ではないが,次のような考え方はど うだろうか? まず,外部環境からの入力はある法則に 従った力学系から生成されており,処理するニューラル ネットワークも結合構造などに規定された制約条件のあ る力学系とみなしてよいだろう.通常であれば相互の制 約条件は学習に良い効果をもたらすとは考え難いが,環 境とニューラルネットワークの各力学系が,相互にうま く制約し合うことで,逆に結果として高次元の学習を容 易にしている可能性はないのか? この発想が正しいか どうかは今後の課題であるが,ここで述べた内容が読者 の刺激になり,新たな研究や発展につながれば幸いであ る*15. 謝 辞 レザバー計算機に関して共同研究者の中嶋浩平氏の 情報提供に感謝します.本研究の一部は文部科学省およ び JSPS 科 研 費(16K01966,16KT0019,15H05877, 15H05878)の助成を受けたものです.◇ 参 考 文 献 ◇
[Aertsen 02] Aertsen, Ad., 伊藤浩之:皮質ダイナミクスと神経計 算機構─実験・解析・モデル─,脳の情報表現,pp. 186-209, 朝倉書店(2002)
[Amari 67] Amari, S.:Theories of adaptive pattern classifiers,
IEEE Trans. on Electronic Compaters, Vo. EC-16, pp. 299-307
(1967)
[甘利 68] 甘利俊一:情報理論 II ─情報の幾何学的理論─,共立出 版(1968)
[Amari 72] Amari, S.: Learning Patterns and pattern sequences by self-organizing nets of threshold elements, IEEE Trans. on
Computers, Vol. C-21, No. 11, pp. 1197-1206(1972)
[Bertschinger 04] Bertschinger, N. and Natschläger, T.: Real-time computation at the edge of chaos in recurrent neural networks, Neural Computation, Vol. 16, No. 7, pp. 1413-1436 (2004)
[Brody 03] Brody, C. D., Romo, R. and Kepecs, A.:Basic mechanisms for graded persistent activity: discrete attractors, continuous attractors, and dynamic representations, Current
Opinion in Neurobiology, Vol. 13, No.2, pp. 204-211(2003) [Friston 11] Friston, K. J.: Functional and effective connectivity:
A review, Brain Connectivity, Vol. 1, pp. 13-36(2011) [Hebb 49] Hebb, D. O.: The Organization of Behavior, New York
John Wiley Inc.(1949)
[Hermans 12] Hermans, M. and Schrauwen, B.: Recurrent kernel machines, computing with infinite echo state networks, Neural
Computation, Vol. 24, No. 1, pp. 104-133(2012)
[Hopfield 82] Hopfield, J. J.: Neural networks and physical
*15 本解説の 1,2,4 章は青柳が,3 章は伊藤が主として担当した. 互いの内容を配慮して全体を再構成し,5 章に二人の内容をま とめている.
[Rumelhart 86] Rumelhart, D. E., Hinton, G. E. and Williams, R. J.: Learning representations by back-propagating errors,
Nature, Vol. 323, No. 6088, pp. 533-536(1986)
[櫻井 08] 櫻井芳雄:脳の情報表現を見る,京都大学出版会(2008) [Seung 00] Seung, H. S., Lee, D. D., Reis, B. Y. and Tank, D. W.: Stability of the memory of eye position in a recruitment network of conductance-based model neurons, Neuron, Vol. 26, pp. 259-271(2000)
[Skarda 87] Skarda, C. A. and Freeman W. J. : How brains make chaos in order to make sense of the world, Behavioral and
Brain Sciences, Vol. 10, pp. 161-195(1987)
[Sompolinsky 88] Sompolinsky, H., Crisanti, A. and Sommers, H.: Chaos in random neural networks, Phys. Rev. Lett., Vol. 61, No. 3, pp. 259-262(1988)
[Sussillo 09] Sussillo, D. and Abbott, L. F.: Generating coherent patterns of activity from chaotic neural networks, Neuron, Vol. 63, No. 4, pp. 544-557(2009)
[Tanaka 09] Tanaka, T., Kaneko, T. and Aoyagi, T.: Recurrent infomax generates cell assemblies, neuronal avalanches, and simple cell-like selectivity, Neural Computation, Vol. 21, No. 4, pp. 1038-1067(2009)
[Watanabe 17] Watanabe, T., Sasaki, Y., Shibata, K. and Kawato, M.: Advances in fMRI real-time neurofeedback, Trends in
Cognitive Sciences, Vol. 21, pp. 997-1010(2017)
[Yamaguti 11] Yamaguti, Y., Kuroda, S., Fukushima, Y., Tsukada, M. and Tsuda, I.: A mathematical model for cantor coding in the hippocampus, Neural Networks, Vol. 24, No. 1, pp. 43-53 (2011)
2018年 5 月 15 日 受理 systems with emergent collective computational abilities, Proc.
National Academy of Sciences, Vol. 79, No. 8, pp. 2554-2558
(1982) [伊藤 97] 伊藤浩之:脳におけるダイナミカルな情報コード─脳科 学とモデル─,神経研究の進歩,Vol. 41, No. 6, pp. 834-841,医 学書院(1997) [伊藤 00] 伊藤浩之:多細胞同時記録データの統計解析法,日本神 経回路学会誌,Vol. 7, No. 1, pp. 8-19(2000) [伊藤 15] 伊藤浩之:セルアセンブリ,脳科学辞典,DOI: 10.14931/ bsd.3845(2015)
[Iwade 18] Iwade, H., Nakajima, K., Tanaka, T. and Aoyagi, T.: Use of recurrent infomax to improve the memory capability of input-driven recurrent neural networks, arXiv:1803.05383 (2018)
[Jaeger 04] Jaeger, H.: Harnessing nonlinearity: Predicting chaotic systems and saving energy in wireless communication,
Science, Vol. 304, No. 5667, pp. 78-80(2004)
[Kenet 03] Kenet, T., Bibitchkov, D., Tsodyks, M., Grinvald, A. and Arieli, A.: Spontaneously emerging cortical representations of visual attributes, Nature, Vol. 425, pp. 954-956(2003)
[Koulakov 02] Koulakov, A. A., Raghavachari, S., Kepecs, A. and Lisman, J. E. : Model for a robust neural integrator, Nature
Neuroscience, Vol. 5, No. 8, pp. 775-782(2002)
[Laje 13] Laje, R. and Buonomano, D. V.: Robust timing and motor patterns by taming chaos in recurrent neural networks,
Nature Neuroscience, Vol. 16, No. 7, pp. 925-933(2013) [Lebedev 17] Lebedev, M. A. and Nicolelis, M. A. L.:
Brain-machine interfaces: from basic science to neuroprostheses and Neurorehabilitation, Physiological Review, Vol. 97, pp. 767-837 (2017)
[Luczak 09] Luczak, A., Bartho, P. and Harris, K. D.: Spontaneous events outline the realm of possible sensory responses in neocortical populations, Neuron, Vol. 62, pp. 413-425(2009) [Lukoševicˇius 12] Lukoševicˇius, M., Jaeger, H. and Schrauwen,
B.: Reservoir computing trends, KI-Künstliche Intelligenz, Vol. 26, No. 4, pp. 365-371(2012)
[Maass 02] Maass, W., Natschläger, T. and Markram, H.: Real-time computing without stable states: A new framework for neural computation based on perturbations, Neural
Computation, Vol. 14, No. 11, pp. 2531-2560(2002)
[Maruyama 13] Maruyama, Y. and Ito, H.: Diversity, heterogeneity and orientation dependent variation of spike count correlation in the cat visual cortex, European J.
Neuroscience, Vol. 38, pp. 3611-3627(2013) [メルロ=ポンティ 42] メルロ=ポンティ 著,滝浦静雄,木田元 訳: 行動の構造,p. 33,みすず書房(1964) [三浦 11] 三浦佳二:ポピュレーションコーディングにおけるノイ ズ相関の影響,日本神経回路学会誌,Vol. 18, No. 2, pp.67-72 (2011)
[Montufar 14] Montufar, G. F., Pascanu, R., Cho, K. and Bengio, Y.: On the number of linear regions of deep neural networks,
Proc. 27th Int. Conf. on Neural Information Processing Systems
(NIPS’14),Vol. 2, pp. 2924-2932(2014)
[Rao 13] Rao, R. P. N.: Brain-Computer Interfacing: An
Introduction, Cambridge University Press(2013)
[Rosenblatt 58] Rosenblatt, F.: The Perceptron: A probabilistic model for information storage and organization in the brain,
Psychological Review, Vol. 65, No. 6, pp. 386-408(1958)
著 者 紹 介
伊藤 浩之 京都産業大学情報理工学部教授.1988 年東京大学大 学院理学系研究科物理学専攻博士後期課程修了(理 学博士).京都大学基礎物理学研究所,理学部物理 第一学科(学術振興会特別研究員),米国ロスアラ モス国立研究所 Center for Nonlinear Studies(客 員研究員),カナダ McGill 大学生理学部 Center for Nonlinear Dynamics in Biology and Medicine(ポ スドク研究員),京都産業大学工学部情報通信工学科講師,同助教授,同 教授を経て 2008 年より現職.専門は神経科学,非線形科学,特に視覚皮 質からの多細胞記録生理実験と Brain-Machine Interface.日本神経科学 学会,日本神経回路学会,Society for Neuroscience 各会員.青柳 富誌生(正会員) 京都大学大学院情報学研究科(先端数理科学専攻) 教授.1993 年京都大学大学院理学研究科博士後期課 程修了.博士(理学).同年,京都大学工学部数理 工学科助手,同大学院情報学研究科複雑系科学専攻 講師,准教授を経て 2014 年より現職.専門は非線 形物理学,理論神経科学,特にリズム現象の解析と ネットワーク上の結合力学系の研究を行う.日本物 理学会,日本神経科学学会,日本数理生物学会,日本神経回路学会各会員.