Molecular analysis of a necrotic cell death
induced in cucumber mosaic virus-inoculated
Arabidopsis thaliana
著者
Tian Ainan
学位授与機関
Tohoku University
学位授与番号
11301甲第19321号
Molecular analysis of necrotic cell
death induced in cucumber mosaic
virus-inoculated Arabidopsis thaliana
(キュウリモザイクウイルス感染シロイヌナズナに
誘導される壊疽性細胞死に関する研究)
1
Contents
1 General introduction ... 9
1.1 Plant-pathogen interactions ... 9
1.2 Cucumber mosaic virus ... 10
1.3 Programmed cell death ... 11
1.4 HR cell death ... 12
1.5 Aims ... 13
2 Identification of necrotic cell death induced in CMV-inoculated Arabidopsis thaliana ... 14
2.1 Introduction ... 14
2.2 Materials and methods ... 14
2.2.1 Plant material ... 14
2.2.2 RNA extraction and RT-PCR ... 15
2.2.3 Construction of in vitro transcription vectors ... 16
2.2.4 Production of reassortant CMV ... 16
2.2.5 Virus inoculation and detection ... 17
2.2.6 Virus quantification by ELISA ... 18
2.2.7 Detection of cell death ... 18
2.2.8 RNA-Seq analysis ... 19
2.3 Results ... 20
2.3.1 Response of Arabidopsis thaliana ecotype Col-0 to a series of reassortant CMVs ... 20
2.3.2 Comparison of global gene expression pattern between two types of cell death in Arabidopsis leaves ... 28
2.3.3 Response of A. thaliana ecotypes to CMV(HYY) ... 34
2.4 Conclusion ... 38 3 Analysis of the determinant region in CMV RNA1 for induction of necrotic cell
2
death in virus-inoculated leaves ... 40
3.1 Introduction ... 40
3.2 Materials and methods ... 40
3.2.1 Plant material ... 40
3.2.2 RNA extraction and RT-PCR ... 40
3.2.3 Construction of in vitro transcription vectors carrying chimeric cDNA of CMV RNA1 ... 41
3.2.4 Single amino acid substitution in 1a protein encoded on RNA1 of CMV(Y) ... 42
3.2.5 Production of reassortant CMV ... 42
3.2.6 Virus inoculation and detection ... 42
3.2.7 Detection of cell death ... 42
3.3 Results ... 43
3.3.1 Analysis of viral sequence in CMV RNA1 for induction of necrotic cell death in virus-inoculated leaves ... 43
3.3.2 Analysis of single amino acid substitutions in the CMV 1a protein for induction of necrotic cell death in virus-inoculated leaves ... 52
3.4 Conclusion ... 55
4 Comprehensive discussion ... 56
5 Literature ... 61
6 Supplementary tables ... 65
3
List of Figures
Figure 1-1 A zigzag model illustrates the quantitative output of the plant immune
system. ... 9
Figure 1-2 Genome organization of CMV. ... 11
Figure 1. Schematic diagrams of RNA genomes of reassortant CMVs ... 22
Figure 2. Response of virus-inoculated leaves of Arabidopsis thaliana ecotype Col-0 to CMV(H), CMV(Y) or a series of reassortant CMVs, and virus multiplication in the inoculated leaves ... 23
Figure 3. Visualization of cell death in CMV(HYY)- or CMV(Y)-inoculated Arabidopsis thaliana Col-0 leaves ... 24
Figure 4. Accumulation of CMV coat protein in CMV(HYY)- or CMV(Y)-inoculated Arabidopsis thaliana Col-0 leaves ... 25
Figure 5. Detection of CMV RNA in virus-inoculated leaves of Arabidopsis thaliana ecotype Col-0 ... 26
Figure 6. Detection of CMV CP in non-inoculated upper Arabidopsis thaliana ecotype Col-0 leaves and systemic symptom development ... 27
Figure 7. Venn diagram of the number of DEGs ... 30
Figure 8. Gene ontology enrichment analysis for DEGs of Class I ... 31
Figure 9. Gene ontology enrichment analysis for DEGs of Class II ... 32
Figure 10. Gene ontology enrichment analysis for DEGs of Class III ... 33
Figure 11. Survey of the response to CMV(HYY)-inoculation on the leaves of 94 ecotypes of Arabidopsis thaliana ... 35
Figure 12. Schematic diagram of the CMV RNA1 encoding the 1a protein ... 45
Figure 13. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, CMV(H-Y/683) and CMV(Y-H/683) .... 46
Figure 14. Detection of CMV coat protein in extracts of virus-inoculated leaves and non-inoculated upper Arabidopsis thaliana Col-0 leaves ... 47 Figure 15. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0
4
leaves inoculated with reassortant CMVs, CMV(Y-H/343) and CMV(Y-H/344~682) ... 48 Figure 16. Detection of CMV coat protein in extracts of virus-inoculated leaves and
non-inoculated upper Arabidopsis thaliana Col-0 leaves ... 49 Figure 17. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0
leaves inoculated with reassortant CMVs, CMV(Y-H/71), CMV(Y-H/72~343), H/290), H/72~290), H/71+291~343) and
CMV(Y-H/291~343) ... 50 Figure 18. Detection of CMV coat protein in extracts of virus-inoculated Arabidopsis
thaliana Col-0 leaves and non-inoculated upper leaves ... 51 Figure 19. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0
leaves inoculated with CMVs carrying single amino acid substitutions in the 1a protein of CMV(Y) ... 53 Figure 20. Detection of CMV coat protein in extracts of virus-inoculated Arabidopsis
thaliana Col-0 leaves and non-inoculated upper leaves ... 54 Figure 21. The induction of cell death in plant-virus interactions………57
5
List of Tables
Table 1. Induction of cell death in CMV(HYY)-inoculated leaves of 94 ecotypes of Arabidopsis thaliana ... 36 Table S1. Primer list for in vitro transcribed cDNA to RNA1, RNA2 and RNA3 of
CMV(H) in this study ... 65 Table S2. Primer list for construction of chimeric cDNA to CMV RNA1 in this study. .. 66 Table S3. Template RNA and Primers used for the first RT-PCR for construction of
chimeric cDNA to CMV RNA1 in this study. ... 67 Table S4. Primer list for nucleotide substitution in cDNA to CMV RNA1 in this study. 68 Table S5. The 149 up-regulated genes in CMV(Y)-inoculated Col::RCY1 leaves showing
HR cell deathTable 6 The 149 up-regulated genes in CMV(Y)-inoculated
Col::RCY1 leaves showing HR cell death ... 69 Table S6 The 35 up-regulated genes common to CMV(HYY)-inoculated Col-0 leaves
showing necrotic cell death and CMV(Y)-inoculated Col-0::RCY1 leaves showing HR cell death ... 75 Table S7 The 17 up-regulated genes in CMV(HYY)-inoculated Col-0 leaves showing
necrotic cell death ... 77 Table S8 The 43 down-regulated genes in CMV(Y)-inoculated Col::RCY1 leaves
showing HR cell death ... 78 Table S9 The 7 down-regulated genes common to CMV(HYY)-inoculated Col-0 leaves
showing necrotic cell death and CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death ... 80 Table S10 The 12 down-regulated genes in CMV(HYY)-inoculated Col-0 leaves
showing necrotic cell death ... 81 Table S11 GO enrichment analysis of the 149 up-regulated genes in
CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death ... 82 Table S12 GO enrichment analysis of the 35 up-regulated genes common to
CMV(Y)-6
inoculated Col::RCY1 leaves showing HR cell death ... 89 Table S13 GO enrichment analysis of the 17 up-regulated genes in
7
Abbreviations
A. thaliana Arabidopsis thaliana
ANOVA one-way analysis of variance
Avr genes avirulence genes
BP biological process
bp base-pair
CMV cucumber mosaic virus
CMV(H) H strain of cucumber mosaic virus
CMV(Y) yellow strain of cucumber mosaic virus
Col::RCY1 RCY1-transformed Col-0
Col-0 Columbia-0
CP coat protein
DEGs differential expressed genes
DIG digoxigenin
DND1 dead end protein homolog 1
down-DEGs down-regulated differential expressed genes
dpi days post inoculation
EDTA ethylenediaminetetraacetic acid
ELISA enzyme-linked immunosorbent assay
ETI effector-triggered immunity
ETS effector-triggered susceptibility
GO gene ontology
GTC guanidinium thiocyanate
HEL helicase
HR hypersensitive response
kb kilo base
LSD-test least significant difference-test
MET methyltransferase
Mt-0 Martuba-0
8
NB-LRR nucleotide binding leucine-rich repeats
Nw-0 Neuweilnau-0
ORF open reading frame
PAMPs pathogen-associated molecular patterns
PCD programmed cell death
PCR polymerase chain reaction
PRRs pattern recognition receptors
PTGS post-transcriptional gene silencing
PTI pattern-triggered immunity
PVX potato virus X
R genes resistance genes
RCY1 resistance to CMV(Y) 1
Rd-0 Rodenbach-0
RdRp RNA-dependent RNA polymerase
RIN13 RPM1 interacting protein 13
RNA-Seq RNA-sequencing
rpm rotations per minute
RPM1 resistance to Pseudomonas syringae pv. maculicola 1
rRNA ribosomal RNA
Rsch-0 Rschew-0
RT-PCR reverse transcription-PCR
Stw-0 Stobowal-0
up-DEGs up-regulated differential expressed genes
UTR untranslated region
v/v volume per volume
9
1 General introduction
1.1 Plant-pathogen interactions
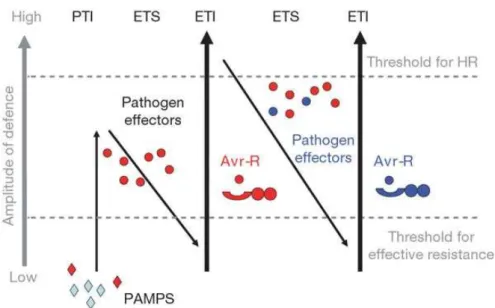
In the field, continuous attacks from numerous pathogens, including viruses, are threatening plant life. For surviving, plants evolved highly efficient defenses to defend themselves. Different from vertebrate immune systems, plant immune systems trigger both preformed defenses at the infection sites and inducible defenses at distal parts of plants. Interactions between plants and pathogens can be represented as consisting of three phases, referred to as a “Zigzag” scheme (Figure 1-1) [1].
Figure 1-1 A zigzag model illustrates the quantitative output of the plant immune system.
During the interactions between plant and pathogens, when pathogen-associated molecular patterns (PAMPs), which are often conserved pathogen molecules essential for their reproduction, were recognized by plant cell surface pattern recognition receptors (PRRs), the first layer of plant defense, pattern (or PRR)-triggered immunity (PTI) was activated. The pathogen failed in penetration of the host cell due to the activation of PTI. However, the arms race between plant and pathogen will not stop but keep evolving. The pathogen alters to produce proteins or other effector molecules under the selective pressures of PTI to suppress PTI, which is known as effector-triggered susceptibility (ETS). When PTI was defeated and pathogens kept
10
releasing their effectors to carry on invasion, the plant was exposed to a selective pressure to produce proteins for the recognition of these effector molecules and the consequently activate defense reaction. Nucleotide binding leucine-rich repeat (NB-LRR) proteins coded by resistance (R) genes in plant recognized the pathogen effectors encoded by avirulence (Avr) genes in the cytoplasm and then triggered the second layer of plant defense, which is known as R gene-mediated defense or effector-triggered immunity (ETI). ETI is a defense system which is much stronger and faster than PTI, and ETI often develops a programmed cell death (PCD) referred to as the hypersensitive response (HR). Therefore, the PCD formed in ETI is also described as HR cell death. Such interactions that fail to result in disease occurrence are described as incompatible interactions. On the other hand, interactions between plants and pathogens that result in disease are described as compatible interactions.
1.2 Cucumber mosaic virus
Cucumber mosaic virus (CMV) is an important virus which is responsible for severe agronomic losses in many crops [2]. It is transmitted by aphids in a nonpersistent
manner and through seeds in some host species [3]. CMV with a high degree of
diversity holds one of the broadest host range among plant viruses. In 1991, Drs Edwardson and Christie reported 1241 host species within 101 plant families, including monocots and dicots [4]. Different isolates of CMV exhibit various biological and
molecular features. The features facilitating experimental manipulations including mechanical transmission, strong accumulation in infected hosts and availability of infectious cDNAs to CMV RNA for a reverse genetic approach make CMV an important model to understand plant-virus interactions.
11
Figure 1-2 Genome organization of CMV.
CMV is one of the best characterized tripartite viruses that contain three positive-sense single stranded genome RNAs, named RNA1-3. RNA1 is monocistronic and encodes 1a protein, which contains two putative functional domains, 0(MET) domain in its N-terminal part, and a helicase motif (HEL) in the C-terminal part. RNA2 encodes the 2a protein, which possesses the GDD motif typical for an RNA-dependent RNA polymerase (RdRp). The 1a protein interacts with 2a protein through the HEL domain in order to form a viral replicase complex, which plays a role in virus replication. RNA2 also has a second open reading frame (ORF) overlapping 3`-terminal part of ORF 2a encoding 2b protein, which functions as a suppressor of post-transcriptional gene silencing (PTGS). Bicistronic RNA3 has two ORFs: 3a and CP. 3a encodes a cell-to-cell movement protein and CP does a coat protein. The coat protein (CP) is translated from subgenomic RNA4 generated from 3’-half of RNA3 and is essential for efficient long-distance movement of CMV. Each RNA size is slightly different among diverse strains, however, each ORF of different strains has a similar size. Each RNA has a cap structure at its 5’ end and can adopt a tRNA-like structure at its 3’-hydroxylated end.
1.3 Programmed cell death
Programmed cell death (PCD) is considered as a fundamental process of life. It is a process well organized at both genetic and biochemical levels [5]. In plant, cells and
12
example, flower development is radically affected by PCD of selected cells [6]. At early
stage of flower development, male and female flowers are indistinguishable. During flower formation, at a development stage that varies with plant species, either male or female parts cease growing and are eliminated through a cell death program [6].
Senescence of organs, including leaves, roots, flower parts, and the tissues of ripening fruits are also highly regulated processes for maximum recovery of nutrients [7].
Besides of plant growth and development, cell death also plays a crucial role in plant response to adverse environmental stimulations. Among them, the cell death triggered by infection of pathogens, including viruses, has perhaps been most intensively studied.
Backing to 1923, a study on a plant invaded by a pathogen, that was the first study to introduce the concept of PCD as a process activated by plants to defend themselves against the infection of pathogens [8]. Later, Dr Barlow reasoned regarding plants that
if cells die at a predictable time and location or if the death has some beneficial effect on tissue differentiation and is inherited in the next generation, it represents PCD [9,10].
This definition excluded necrosis as cell death due to accidental or random injuries such as exposure to some toxins or a lethal temperature [9]. However, history has
revealed that necrosis in many plant processes is programmed and meets many or all of Barlow’s criteria [10,11]. In this study, PCD resulting from symptom formation in
incompatible interactions between plant and pathogen has been described as necrotic local lesions (HR cell death), and that in compatible interactions was described as necrosis according to Dickman’s work [11].
1.4 HR cell death
RCY1, isolating from Arabidopsis thaliana (A. thaliana) ecotype C24, is a resistance gene to a yellow strain of CMV [CMV(Y)] [12]. The avirulent gene has been identified to
the CP of CMV(Y). When A. thaliana ecotype Columbia-0 (Col-0) was transformed with RCY1, it exhibits HR to the infection of CMV(Y). HR cell death is induced from 3 days
13
post inoculation (dpi) and restricts the virus to the HR lesion due to the resistance conferred by RCY1 [13]. Since HR cell death is critical to virus multiplication, the virus
replication in the inoculated leaves as well as the long-distance movement of the virus is suppressed and stopped during the induction of HR cell death [13]. However, virus
still can move into the living cells surrounding HR lesions. Recent researches indicated that cell death can be uncoupled from resistance and thus it may not be the primary barrier against pathogen proliferation. For example, Rx-mediated resistance against the potato virus X (PVX) occurs without the induction of HR cell death at the site of initial infection and thereby differs from the usual HR, was identified with extreme resistance (ER) in plants [14]. This research indicated that HR cell death is separated
from the resistance responses in plants. The pathogen arrest was thought to be due to sufficient R protein signaling. Similarly, over-expression of R protein, RPM1, interacting protein 13 (RIN13) results in abolishing HR cell death but enhancing bacterial restriction in leaves undergoing RPM1 defense response [15]. Besides, mutation of a
cyclic nucleotide-gated ion channel DND1 could also confer gene-for-gene resistance without HR cell death [16,17]. Even it has been reported that the humidity and
temperature can inhibit the HR cell death without compromising resistance [18,19].
1.5 Aims
The induction of cell death is a dominant feature between plant and virus interactions. When the resistance gene of host plant co-existed the avirulent gene of virus, HR cell death was induced in the virus-inoculated leaves and the virus was blocked to the HR lesion due to the resistance response [20]. On the other hand, when the virus escaped
from the recognition of host plant, lethal systemic necrosis was induced by the systemic spread of virus in the plants [21]. Some recent studies suggesting that systemic
necrosis could be induced through a delayed HR in the virus -inoculated plants, are accumulating, although lethal systemic cell death is categorized in compatible interaction between host plant and virus [21,22]. Thus, the role of cell death in plant
14
and virus interactions is remaining unclear. In this study, the model pathosystem of cucumber mosaic virus and A. thaliana was used to study the cell death induced in plant and virus interactions.
2 Identification of necrotic cell death induced in
CMV -inoculated Arabidopsis thaliana
2.1 Introduction
A. thaliana ecotype Col-0 exhibits different responses to the infection of CMV(H) and CMV(Y). For analyzing the virus factor that determines such different symptom developments, a series of reassortant CMVs were produced between CMV(H) and CMV(Y). In the first part, I analyzed responses of A. thaliana ecotype Col-0 to the infection of each reassortant CMV and observed cell death induced in reassortant CMV-inoculated leaves. The cell death induced in reassortant CMV-inoculated Col-0 leaves was further characterized through its comparison with RCY1- mediated HR cell death and referred as necrotic cell death. In the second part, I performed a comparative analysis of global gene expression pattern between necrotic cell death and HR cell death. In the third part, 94 ecotypes of A. thaliana were used to test if the induction of necrotic cell death is a specific phenomenon between reassortant CMV and Col-0 or a conserved one.
2.2 Materials and methods
2.2.1 Plant material
A. thaliana ecotype Col-0 and other 94 ecotypes, RCY1-transformed Col-0 (Col::RCY1)
[12], and Nicotiana benthamiana were used in the experiment. The plants were grown
15
photoperiod (14000 lux) at 25°C in a KG-201 HL-D growth chamber (Koito, Yokohama, Japan).
2.2.2 RNA extraction and RT-PCR
The collected leaf tissue was weighted and snap-frozen in liquid nitrogen. Then the frozen tissue was homogenized by Multi-Beads Shocker (Yasui kikai, Osaka, Japan) and 6-fold amount of RNA extraction buffer [4.2 M guanidinium thiocyanate: GTC, 0.5 % (w/v) Sodium N-lauroylsarcosinate, Trisodium Citrate Dihydrate, 1.6 % (v/v) 2-mercaptoethanol]. After a phenol-chloroform extraction and a 2-isopropanol precipitation, the pellet was dissolved in sterile water. 0.25-fold amount of 10M NaCl was added to the mixture and then kept it standing at 4 ◦C for 30 minutes. The mixture
was centrifuged for 20 minutes at 12000 rpm and discarded the supernatant. The pellet was washed with 70 % ethanol by spinning for 5 minutes, the supernatant was discarded, and the RNA was air-dried before being re-suspended in 30 μl of RNase-free
water and quantified using Nanodrop Microvolume Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, US).
cDNAs of CMV(H) RNA1, 2, and 3 were synthesized by RT-PCR with the following sets of primers: CMV.RNA1-5'.F and CMV.RNA1-3'.R for RNA1, CMV.RNA2-5'.F and CMV.RNA2-3'.R for RNA2, and CMV.RNA3-5'.F and CMV.RNA3-3'.R for RNA3 (Table S1). The sequence encoding T7 RNA polymerase was included in the primers CMV.RNA1-5'.F, CMV.RNA2-5’.F, and CMV.RNA3-5'.F (Table S1). RT-PCR reactions were performed using the PrimeScript™ II High Fidelity One Step RT-PCR Kit (Takara Bio, Shiga, Japan) according to the manufacturer’s instructions. 100 ng of total RNA and 1 μl of each 5 μM primer (0.4 μM final concentration) listed in Table S1 were added in a 50 μl RT-PCR reaction mixture containing 25 μl 2xOne step high fidelity buffer, PrimeScript II RT Enzyme Mix, PrimeSTAR GXL for 1 step RT-PCR and RNase-free water. The RT-PCR was programmed under the following conditions: reverse transcription at 45 ◦C for 10 min;
16
s at 60 ◦C, and 10 sec/kb at 68 ◦C.All the PCR products were purified with Wizard® SV
Gel and PCR Clean-Up System (Promega, WI, USA).
2.2.3 Construction of in vitro transcription vectors
The gel-purified cDNA of RNA1 was cloned into the HindIII and NotI sites of pUC118 (Takara Bio) and the cDNAs of RNA2 and RNA3 were cloned into the BamHI and NotI sites using the In-Fusion HD Cloning System (Takara Bio) according to the manufacturer’s instructions. The plasmid constructs containing each of the CMV(H) RNA1, RNA2, and RNA3 cDNAs were isolated according to standard protocols [23], and
were designated as pCH1-T7, pCH2-T7, and pCH3-T7, respectively. The plasmids were validated by Sanger sequencing on a CEQ8000 Automated DNA Sequencer (Beckman Coulter, Brea, CA).
In vitro transcription of infectious CMV RNA pCH1-T7, pCH2-T7, and pCH3-T7 were linearized by digestion with NotI and purified using the Wizard® SV Gel and PCR Clean-Up System (Promega). Each linearized plasmid DNA was then transcribed in vitro using T7 RNA polymerase with the standard AmpliCap-Max™ T7 High Yield Message Maker Kit (Cellscript, Madison, WI) according to the manufacturer’s instructions.
2.2.4 Production of reassortant CMV
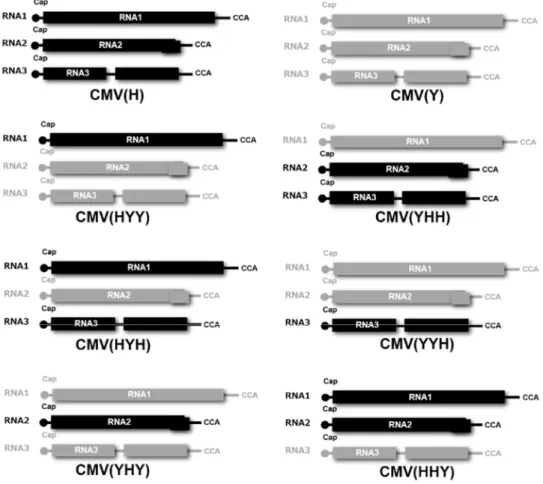
To generate the reassortant CMVs including CMV(HYY), CMV(YHY), CMV(YHH), CMV(YYH), CMV(HYH), and CMV(HHY) (Figure 1), each infectious CMV RNA1, RNA2, and RNA3 was reciprocally exchanged between CMV(Y) and CMV(H). CMV(Y) [24] and H strain of cucumber mosaic virus [CMV(H)] that was isolated from an Arabidopsis halleri plant showing no symptoms, were used for these experiments. Four-week-old N. benthamiana was rub-inoculated with a combination of infectious CMV(Y) and CMV(H) RNA1, RNA2, and RNA3 to propagate a series of reassortant CMVs [25]. At 7
17
then ground in 0.1 M phosphate-buffered saline (pH 8.0) on ice. These homogenates were used to inoculate new fully expanded leaves of 6-week-old N. benthamiana plants. At 4 dpi, the inoculated leaves were harvested and weighted. The harvested leaves were added in 4-fold amount of virus propagation buffer [0.25 M sodium citrate (pH 6.5), 50 % (v/v) chloroform, 5 mM ethylenediaminetetraacetic acid (EDTA) (pH 8.0), 0.05 % (v/v) 2-mercaptoethanol] and grounded with homogenizer for 5 min at 12000 rpm on ice. The homogenate was centrifuged for 10 min at 6000 rpm at 4 ◦C. The
supernatant was transferred to the dialysis cellulose membrane pocket and dialyzed in 5 mM borate buffer (pH 9.0) at 4 ◦C for one night. The purified virus was stocked at
-80 ◦C.
2.2.5 Virus inoculation and detection
Fully expanded leaves of A. thaliana were rub-inoculated with 100 μg/ml of purified virus (section 2.1.4) as previously described [13]. Virus was detected immunologically
by western blot analysis according to the standard protocol [23] using antibody against
the CP of CMV.
Accumulation of CMV RNA in virus-inoculated leaves of Col-0 was analyzed by northern hybridization according to the standard protocol [23]. CMV RNA-specific
cDNA probes complementary to the 3 noncoding region of all CMV RNAs were amplified from CMV(Y) RNA3 cDNA with a pair of primers: 5 -GTGAACGGGTTGTCCATCCA-3 and 5 -ACCCTGAAACTAGCACGTTGT-3 by PCR. The probe cDNA was labeled with digoxigenin (DIG)-11-dUTP using a DIG PCR labeling kit (Roche, Penzberg, Germany) according to the manufacturer’s instructions. The PCR product was purified according to the procedure of Takahashi and Ehara [25].
Ribosomal RNA (rRNA)-specific probe was obtained as described previously [13]. All
CMV RNAs were detected using an alkaline phosphatase conjugated anti-DIG antibody (Roche, Penzberg, Germany) and visualized with the CDP-Star Reagent (New England Biolabs, Beverly, MA, USA) according to the manufacturer’s protocols.
18
After A. thaliana ecotype Col-0 was inoculated with CMV containing chimeric RNA1 or RNA1 carrying a nucleotide substitution, all the chimeric RNA1 cDNAs and single nucleotide substitution RNA1 cDNAs were amplified by RT-PCR from the upper noninoculated leaves of inoculated plants. RT-PCR-amplified fragments were purified by treatment with ExoSAP-IT PCR Clean Up Reagents (Thermo Fisher Scientific, Waltham, MA, USA) according to the instruction manual, and their nucleotide sequences were confirmed by Sanger sequencing using a CEQ8000 Automated DNA Sequencer (Beckman Coulter, Brea, CA, USA).
2.2.6 Virus quantification by ELISA
For quantitative measurement of the CMV CP by ELISA, three independent virus-inoculated leaves were homogenized in a 10× volume of 0.01 M potassium phosphate buffer (pH 8.0), respectively. The protein concentrations of the homogenates were determined using the Bradford reagent [26]. The homogenates used for ELISA were
adjusted to 0.03 mg/ml total protein with 0.01 M potassium phosphate buffer. The CP quantities were measured using the method of Koenig [27] and expressed as the
absorbance at 405 nm per 0.03 mg/ml of total protein. Statistical analyses of the CP quantities were performed using one-way analysis of variance (ANOVA) and Fisher’s least significant difference LSD test for post hoc comparisons using IBM SPSS Statistics version 25 (IBM, Armonk, NY).
2.2.7 Detection of cell death
Cell death in CMV-inoculated leaves was visualized by staining with trypan blue according to a standard protocol [28]. Virus-inoculated leaves were stained by boiling
for 8 min in alcoholic lactophenol [99.5 % ethanol: phenol: glycerol: lactic acid 4:1:1:1 (v: v: v: v)] containing 0.1 mg/ml trypan blue. The stained leaves were decolorized in a 2.5 g/ml chloral hydrate solution overnight, and then held and pictured in 70 % ethanol.
19
2.2.8 RNA-Seq analysis
Three independent mock- and CMV(HYY)-inoculated Col-0 leaves showing necrotic cell death at 5 dpi and mock- and CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death at 3 dpi were used for extraction of total RNAs with the RNeasy Plant Mini Kit (Qiagen GmbH, Hilden, Germany). cDNA libraries were prepared using the TruSeq Stranded Total RNA with Ribo-Zero Plant Kit (Illumina, San Diego, CA) according to the manufacturer’s instructions. Approximately 2.7-3.9×105 paired-end reads (75-bp × 2)
were obtained for each sample using the Illumina MiSeq (Illumina). The raw sequence data was submitted to the NCBI Gene Expression Omnibus under accession number GSE137625. The sequence reads were processed using Trimmomatic version 0.38 [29]
for adaptor trimming and quality filtering. The processed reads were mapped to the genome sequences of A. thaliana ecotype Col-0, CMV(Y), and CMV(H) using STAR (version 2.7) [30] at default settings. Read counts per A. thaliana gene were retrieved
using the quantification option in STAR. Read counts were normalized and statistically tested using DESeq2 R package 3.7 [31]. Adjusted p-values were calculated [32], and the threshold adjusted p-value was set to 0.05 for the present study. Independent filtering in DESeq2 with an automatically optimized threshold was performed to filter out the genes with low mean normalized counts. Genes that passed independent filtering in both necrotic cell death versus mock and HR cell death versus mock comparisons were further analyzed for differential expression. Genes with fold-change >4 or <0.25 at an adjusted p-value of <0.05 were considered as differentially expressed genes (DEGs). DEGs with increased expression unique to HR cell death were classified as Class I, DEGs with commonly increased expression in HR cell death and necrotic cell death were classified as Class II, and DEGs with increased expression unique to necrotic cell death were classified as Class III. DEGs with decreased expression unique to HR cell death were classified as Class IV, DEGs with commonly decreased expression in HR cell death and necrotic cell death were classified as Class V, and DEGs with decreased expression
20
unique to necrotic cell death were classified as Class VI. The VennDiagram package [33]
was used to generate Venn diagrams of the sets of DEGs that overlapped between HR cell death and necrotic cell death. Gene symbols information were extracted using Metascape [34]. Gene ontology (GO) enrichment analysis was implemented using
ClusterProfiler package 3.14.0 in R software for the DEGs in each class [35].
2.3 Results
2.3.1 Response of Arabidopsis thaliana ecotype Col-0 to a series
of reassortant CMVs
Schematic structures of the reassortant CMV RNA genomes of the two parent CMV strains, CMV(Y) and CMV(H), are shown in Figure 1. When fully-expanded leaves of three independent A. thaliana ecotype Col-0 plant were inoculated with one of the reassortant CMVs [CMV(HHY), CMV(HYY), CMV(YHH), CMV(YYH), CMV(YHY) or CMV(HYH); and CMV(Y) or CMV(H) strain as a control], a cell death developed at 5 dpi in those inoculated with three of the reassortant CMVs containing CMV(H) RNA1: CMV(HHY), CMV(HYY), or CMV(HYH). However, cell death did not occur in Col-0 leaves inoculated with other reassortant CMV, CMV(Y), or CMV(H) (Figure 2A and Figure 3). At 5 dpi, cell death (which affected a much larger area in comparison with HR cell death) developed in Col-0 leaves inoculated with CMV(HHY), CMV(HYY), or CMV(HYH) (Figure 2A); whereas HR cell death developed in CMV(Y)-inoculated leaves of RCY1-transformed Col-0 (Col::RCY1) at 3 dpi (Figure 3). These results suggest that CMV(H) RNA1 might be associated with cell death development in reassortant CMV-inoculated leaves through its interaction with CMV(Y) RNA2 or CMV(Y) RNA3 and a characteristic of this cell death is to spread to a broader area around the virus primary infection site than occurs with HR cell death.
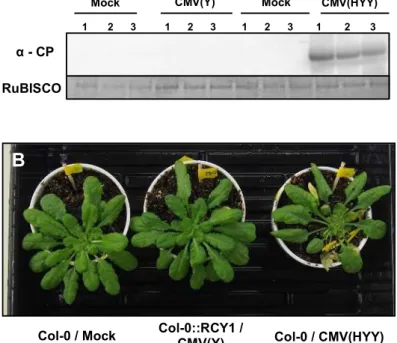
The intensities of CMV CP bands detected by western blot analysis in CMV(HYY)-, CMV(HHY)-, or CMV(HYH)-inoculated leaves of Col-0 exhibiting cell death were
21
comparable to those in CMV(Y)-, CMV(H)-, or other reassortant CMV-inoculated Col-0 leaves showing no cell death at 5 dpi (Figure 2B). The accumulated level of CP in CMV(HYY)-inoculated leaves was also quantitatively similar to that in CMV(Y)-inoculated Col-0 showing no cell death (Figure2C), but significantly higher than in CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death (Figure 4). Furthermore, comparison of the intensity of the norther blot analysis bands of CMV RNA1, RNA2, and RNA3 among the leaves inoculated with eight CMVs [CMV(H), CMV(Y), or one of six reassortant CMVs] suggests that there is no significant correlation between the induction of cell death and the accumulated level of CMV RNAs or the ratio of CMV RNA1, RNA2, and RNA3 (Figure 5). These results indicate that the cell death developing on the leaves inoculated with reassortant CMV carrying CMV(H) RNA1, seems to not suppress virus replication but instead allows it to multiply at the same level as with a susceptible interaction.
To investigate whether or not the cell death developing on the leaves inoculated with CMV(HYY) carrying CMV(H) RNA1 affects virus systemic spread to noninoculated upper leaves, CP in noninoculated upper leaves of CMV(HYY)-infected Col-0, CMV(Y)-infected Col::RCY1, or mock-inoculated Col-0 was detected by western blot analysis (Figure 6A). CP only accumulated in noninoculated upper leaves of CMV(HYY)-inoculated plants showing systemic stunting and weak yellowing symptoms, but not in upper leaves of CMV(Y)-infected Col::RCY1 or mock-inoculated Col-0. Moreover, systemic cell death was not observed in CMV(HYY)-inoculated plants and CMV(Y)-infected Col::RCY1 and mock-inoculated Col-0 (Figure 6B). Thus, the cell death developing on the leaves inoculated with reassortant CMV carrying CMV(H) RNA1 seems not to contribute to the resistance to CMV, and therefore differs from HR cell death.
22
Figure 1. Schematic diagrams of the RNA genomes of the reassortant CMVs, derived from RNA1, RNA2, and RNA3 of CMV(H) and CMV(Y) (upper part of figure), used in the present study. Black indicates the CMV(H) genomes; gray, the CMV(Y) genomes.
23 0 0.5 1 1.5 2 2.5 0 1 2 3 4 5 6 7 8 R el a ti v e a m o u n t o f co a t p ro te in
Days after inoculation
Col::RCY1#12-Mock Col::RCY1#12-CMV(Y) Col-CMV(H1Y2Y3) Cell death Cell death
CMV(YYH) CMV(YHH) CMV(YHY) RuBISCO
CMV(H) CMV(HYY) CMV(HYH) CMV(HHY) α - CP
1 2 3 1 2 3 1 2 3 1 2 3
1 2 3 1 2 3 1 2 3 Inoculated leaves of Col-0
Moc k CM V(Y ) Moc k CM V(Y ) RuBISCO α - CP C Col::RCY1 / Mock Col::RCY1 / CMV(Y) Col-0 / CMV(HYY) B A CMV(HYY) CMV(HYH) CMV(YHY) CMV(YHH) CMV(YYH) CMV(HHY) CMV(Y) CMV(HYY) CMV(HYH) CMV(YHY) CMV(YHH) CMV(YYH) CMV(HHY) CMV(Y) 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 CMV(H) CMV(H)
Figure 2. Response of virus-inoculated leaves of A. thaliana ecotype Col-0 to CMV(H), CMV(Y) or a series of reassortant CMVs, and virus multiplication in the inoculated leaves. (A) Development of cell death in leaves with a series of reassortant CMVs, or with CMV(H) or CMV(Y) as a control. Representative virus-inoculated Col-0 of 3 independent plants (plant numbers 1, 2 and 3) under bright field (left panel) and stained with trypan blue (right panel). (B) CMV CP detected immunologically by western blotting at 7 dpi in the leaves of plants inoculated with one of a series of reassortant CMVs. CMV(Y)-inoculated Col-0 leaves and mock-CMV(Y)-inoculated Col-0 leaves were used as positive and negative control. RuBISCO protein is shown as an internal reference for protein quantity. (C) Time course of virus multiplication in Col-0 leaves inoculated with CMV(HYY) carrying CMV(H) RNA1 [Col-0/CMV(HYY)], CMV(Y)-inoculated Col::RCY1 leaves [Col::RCY1/CMV(Y)], and mock-inoculated Col::RCY1 leaves [Col::RCY1/Mock]. CMV CP quantities were measured using ELISA (mean values of relative amount of CP of three independent biological samples with standard error bars).
Figure 2-2. Response of virus-inoculated leaves of Arabidopsis thaliana ecotype Col-0 to CMV(H), CMV(Y) or a series of reassortant CMVs, and virus multiplication in the inoculated leaves
24
Figure 3. Visualization of cell death in CMV(HYY)- or CMV(Y)-inoculated A. thaliana Col-0 leaves. (A) HR cell death in CMV(Y)-inoculated A. thaliana Col::RCY1 at 3 days post-inoculation (dpi) under bright field (upper panel), and with trypan blue staining (lower panel). (B) Cell death visualized by trypan blue staining: three independent CMV(HYY)-inoculated A. thaliana Col-0 leaves at 5 dpi; HR cell death in three independent CMV(Y)-inoculated A. thaliana Col::RCY1 leaves at 3 dpi; and mock-inoculated A. thaliana Col-0 leaves at 3 and 5 days.
25
Figure 4. Accumulation of CMV CP in CMV(HYY)- or CMV(Y)-inoculated A. thaliana Col-0 leaves at 5 days after inoculation. CP was quantitatively measured using ELISA. Vertical error bars indicate standard error of the mean for CP quantity from three independent samples; bars with the same letter indicate no significant difference between these treatments at p <0.05 by LSD test.
Figure 2-4. Accumulation of CMV coat protein in
26
RNA1 RNA3 RNA4
Inoculated leaves of Col-0
rRNA RNA2
Figure 5. Detection of CMV RNA in virus-inoculated leaves of A. thaliana ecotype Col-0. CMV RNA1, 2, 3 and 4 were detected by northern blot hybridization analysis of total RNA extracted from virus-inoculated leaves of Col-0 inoculated with CMV(Y), CMV(H) and a series of reassortant CMVs (as indicated) at 5 dpi. Mock-inoculated Col-0 leaves were used as a control. Total RNA was extracted from three independent samples. The position of CMV RNA is indicated at left: RNA 1, 2 and 3 represent genomic RNAs; RNA4 is subgenomic. rRNA is the loading control.
Figure 2-5. Detection of CMV RNA in virus-inoculated leaves of Arabidopsis thaliana ecotype Col-0
27
Non-inoculated upper leaves
Col::RCY1 /
CMV(Y) Col-0 / Mock CMV(HYY) Col-0 /
Col::RCY1 / Mock 1 2 3 1 2 3 1 2 3 1 2 3 α - CP RuBISCO A Col-0 / CMV(HYY) Col-0::RCY1 / CMV(Y) Col-0 / Mock B
Figure 6. Detection of CMV CP in non-inoculated upper A. thaliana ecotype Col-0 leaves and systemic symptom development. (A) CMV CP detected at 7 dpi, by western blot analysis, in non-inoculated upper leaves of CMV(HYY)-infected or mock-inoculated Col-0 [Col-0/CMV(HYY) and Col-0/Mock] and CMV(Y)-inoculated or mock-inoculated Col::RCY1 [Col::RCY1/CMV(Y) and Col::RCY1/Mock]. RuBISCO protein is an internal reference for protein quantity. Each experiment was conducted using three independent biological replicates (plant numbers 1, 2, and 3). (B) Symptom appearance observed at 14 dpi on CMV(HYY)-inoculated Col-0 [Col-0/CMV(HYY)], CMV(Y)-inoculated Col::RCY1 [Col::RCY1/CMV(Y)], or mock-inoculated Col-0 [Col-0/Mock] (control). Virus-inoculated leaves have been removed because they were already dead at this stage. Representative plants were photographed.
Figure 2-6. Detection of CMV CP in non-inoculated upper Arabidopsis thaliana ecotype Col-0 leaves and systemic symptom development
28
2.3.2 Comparison of global gene expression pattern between
two types of cell death in Arabidopsis leaves
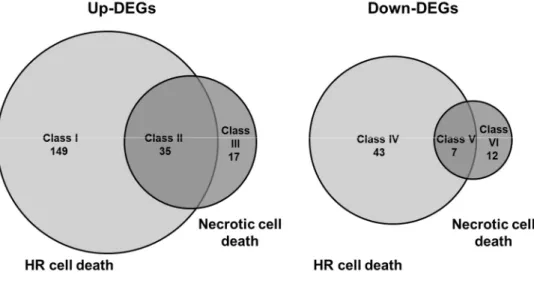
To further characterize cell death in CMV-inoculated Col-0 leaves, global gene expression patterns were compared by RNA-Seq between CMV(HYY)-inoculated Col-0 leaves showing cell death and CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death. Genes were considered DEGs during the analysis of DESeq2 (Tables S7-S12) for a >4-fold increase in expression or a <0.25-fold decrease in expression at an adjusted p-value of <0.05. Changes in transcript abundances in CMV(HYY)-inoculated Col-0 leaves showing cell death and CMV(Y)-inoculated Col::RCY1 showing HR cell death were compared against mock treatment controls for 5906 genes with sufficient read counts for statistical analyses (adjusted p-value <0.05). As shown in Figure 7, a total of 202 genes showed a >4-fold increase in transcript abundance (adjusted p-value <0.05). Of these, 35 showed significant increase in common to CMV(HYY)-induced cell death and CMV(Y)-induced HR cell death (Class II genes), while 149 (Class I genes) and 18 (Class III genes) transcripts showed significant unique increase during CMV(Y)-induced HR or CMV(HYY)-induced cell death, respectively (Figure 7; Tables S7-S9). Simultaneously, 62 genes showed a <0.25-fold decrease in transcript abundance (adjusted p <0.05) (Figure 7). Seven of these showed significant increase in common (Class V genes), while 43 (Class IV genes) and 12 (Class VI genes) transcripts were specific to CMV(Y)-induced HR or CMV(HYY)-induced cell death, respectively (Figure 7; Tables S10-S12). These results suggest that global gene expression patterns differ between CMV(HYY)-inoculated Col-0 leaves developing cell death and HR-mediated PCD coupled to RCY1-conferred CMV(Y) resistance.
The number of genes for which transcript expression increased > 4-fold or decreased < 0.25-fold in CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death was greater than that in CMV(HYY)-inoculated Col-0 leaves showing cell death (Figure 7). Furthermore, the genes (Class I genes) with increased transcript abundance in
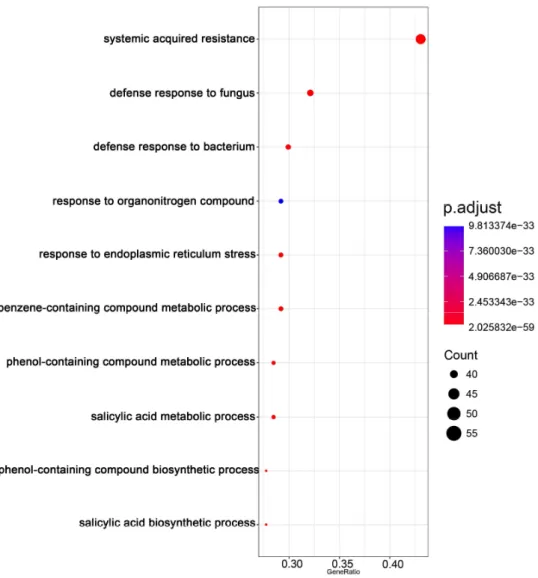
29
CMV(Y)-inoculated Col::RCY1 leaves encoded several defense-related proteins, that is, chitinase, pathogenesis-related (PR) proteins, and WRKY transcription factors; several leucine-rich repeat kinases and receptor-like proteins; calcium-binding EF-hand family proteins; and glutathione S-transferases (Table S5). Gene ontology (GO) enrichment analysis suggested that the top 3 GO enrichment term in biology process for the identified DEGs with increased expression that were specific to HR cell death were enriched into “systemic acquired resistance”, “salicylic acid metabolic process” and “salicylic acid biosynthetic process”. (Figure 8, Table S11). Transcripts encoded by overlapping sets of genes (Class II genes) were enriched to GO terms “systemic acquired resistance”, “salicylic acid biosynthetic process” and “cell death” and could therefore be associated generally with the induction of cell death (Figure 9, Table S12). In contrast, transcripts of Class III genes specific to cell death in CMV(HYY)-inoculated leaves were enriched to GO terms such as “jasmonic acid mediated signaling pathway”, “cellular response to jasmonic acid stimulus” and “response to jasmonic acid” (Figure 10, Table S13). Therefore, cell death observed in CMV(HYY)-inoculated Col-0 leaves might be a form of necrotic cell death that does not contribute to the resistance to CMV.
30
Figure 7. Venn diagram of the number of genes with increased or decreased transcript abundance in CMV(HYY)-inoculated A. thaliana Col-0 leaves showing necrotic cell death and CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death. The number of genes detected by RNA-Seq analysis with more than 4-fold increased expression and adjusted p < 0.05 in CMV(HYY)-inoculated Col-0 leaves and CMV(Y)-inoculated Col::RCY1 leaves are shown at left (Up-DEGs). Those with less than 0.25-fold decreased expression (p <0.05) are shown at right (Down-DEGs). The number of genes with increased and decreased expression in leaves showing HR cell death are shown in the light gray circles; and those showing necrotic cell death in the dark gray circles.
31
Up-regulated DEGs in HR cell death (Class I)
Figure 8. Gene ontology enrichment analysis for up-regulated 149 differentially expressed genes specific in CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death. The top 10 GO enrichment terms in BP for the DEGs were shown. The number of genes in each category is equal to the dot size. The dot color represents the p.adjust value.
32
Up-regulated DEGs in HR cell death and Necrotic cell death (Class II)
Figure 9. Gene ontology enrichment analysis for up-regulated 35 genes in both CMV(HYY)-inoculated Col-0 leaves showing necrotic cell death and CMV(Y)-inoculated Col::RCY1 leaves showing HR cell death. The top 10 GO enrichment terms in BP for the DEGs were shown. The number of genes in each category is equal to the dot size. The dot color represents the p.adjust value.
33
Up-regulated DEGs in Necrotic cell death (Class III)
Figure 10. Gene ontology enrichment analysis for up-regulated 17 genes specific in CMV(HYY)-inoculated Col-0 leaves showing necrotic cell death. The GO enrichment terms in BP for the DEGs were shown. The number of genes in each category is equal to the dot size. The dot color represents the p.adjust value.
34
2.3.3 Response of A. thaliana ecotypes to CMV(HYY)
To determine whether the cell death induced in CMV(HYY)-inoculated leaves of A. thaliana ecotype Col-0 is a general response in A. thaliana ecotypes, 94 ecotypes of A. thaliana were inoculated with CMV(HYY). Cell death developed in virus-inoculated leaves of 92 of these ecotypes from 5 dpi to 9 dpi, but not in the ecotypes Mt-0 and Stw-0 at 14 dpi (Figures 11A and B, and Table 1). The CMV CP was detected in CMV(HYY)-inoculated Mt-0 and Stw-0 at similar levels compared with the virus-inoculated leaves of other ecotypes (Figure 11C). Cell death was not observed on upper leaves of any of the 94 ecotypes systemically infected with CMV(HYY). Thus, A. thaliana ecotypes appear to generally develop cell death in response to CMV(HYY).
35
n o n - H R ( + ) n o n - H R ( - ) 92 ecotypes
2 ecotypes
B
Cell death (+) Cell death (-)
A
CMV(HYY) -inoculated leaves E c o ty p e Mt-0 Nw-0 Rd-0 Rsch-4 Stw-0 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3C
1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 RuBISCO Mt-0 Nw-0 Rd-0 Rsch-4 Stw-0 α - CP CMV(HYY)-inoculated leaves Mock CMV(Y) Col-0Figure 11. Survey of the response to CMV(HYY)-inoculation on the leaves of 94 ecotypes of A. thaliana. (A) Representative photograph of the responses: CMV(HYY)-inoculated leaves of five ecotypes randomly selected at 14 dpi. Virus-inoculated leaves under bright field (left panel) and stained with trypan blue (right panel). (B) Pie chart summary of CMV(HYY)-inoculated leaves of the 94 ecotypes. (C) CMV CP detected immunologically by western blot analysis in virus-inoculated leaves of three independent biological replicates (numbers 1, 2, and 3) of five selected ecotypes at 7 dpi. RuBISCO protein is an internal reference for protein quantity.
Figure 2-11. Survey of the response to CMV(HYY)-inoculation on the leaves of 94 ecotypes of Arabidopsis thaliana
36 Table 1. Induction of cell death in CMV(HYY)-inoculated leaves of 94 ecotypes of Arabidopsis thaliana
SASSC2)
number Ecotype
Cell Death / Viral CP1)
SASSC2)
number Ecotype
Cell Death / Viral CP1)
SASSC2)
number Ecotype
Cell Death / Viral CP1)
Inoculated leaves Non-inoculated upper leaves Inoculated leaves Non-inoculated upper leaves Inoculated leaves Non-inoculated upper leaves
JA003 Ak-1 + / + - / + JA112 Ita-0 + / + - / + JA215 Sg-2 + / + - / +
JA007 Ang-1 + / + - / + JA115 Jm-0 + / + - / + JA224 Stw-0 - / + - / +
JA008 Ba-1 + / + - / + JA117 Kä-0 + / + - / + JA230 Ts-1 + / + - / +
JA009 Bay-0 + / + - / + JA119 Kas-1 + / + - / + JA234 Ts-6 + / + - / +
JA013 Bl-1 + / + - / + JA123 Kl-0 + / + - / + JA236 Tsu-0 + / + - / +
JA014 Bla-1 + / + - / + JA126 Ko-3 + / + - / + JA237 Tu-1 + / + - / +
JA021 Br-0 + / + - / + JA131 La-1 + / + - / + JA239 Ty-0 + / + - / +
JA022 Bs-1 + / + - / + JA132 Lan-0 + / + - / + JA243 Van-0 + / + - / +
JA025 Bu-0 + / + - / + JA134 Le-0 + / + - / + JA247 Wc-2 + / + - / +
JA029 Bu-5 + / + - / + JA137 Li-3-3 + / + - / + JA248 Wil-1 + / + - / +
JA040 Bu-22 + / + - / + JA145 Lu-1 + / + - / + JA254 Wt-2 + / + - / +
JA043 Bu-25 + / + - / + JA147 Ma-0 + / + - / + JA259 X-0 + / + - / +
JA048 Cal-0 + / + - / + JA150 Me-0 + / + - / + JA263 Ze-0 + / + - / +
JA049 Can-0 + / + - / + JA156 Mrk-0 + / + - / + JA264 Zü-0 + / + - / +
JA051 Cha-0 + / + - / + JA158 Mt-0 - / + - / + JA269 Be-1 + / + - / +
JA056 Cl-0 + / + - / + JA161 Na-1 + / + - / + JA286 Co-2 + / + - / +
JA058 Col-0 + / + - / + JA165 No-0 + / + - / + JA308 Gr-6 + / + - / +
JA059 Ct-1 + / + - / + JA167 Nok-1 + / + - / + JA320 Ko-2 + / + - / +
JA060 Cvi-0 + / + - / + JA172 Nw-0 + / + - / + JA324 Li-5-3 + / + - / +
JA062 Db-0 + / + - / + JA174 Ob-1 + / + - / + JA352 Van-1 + / + - / +
37
JA075 Er-0 + / + - / + JA180 Ove-0 + / + - / + JW106 Pak-2 + / + - / +
JA076 Es-0 + / + - / + JA182 Pa-1 + / + - / + JW117 ES + / + - / +
JA078 Est-0 + / + - / + JA185 Per-2 + / + - / + JW120 TKS + / + - / +
JA088 Gd-1 + / + - / + JA193 Pr-0 + / + - / + JW121 IK + / + - / +
JA089 Ge-0 + / + - / + JA195 Ra-0 + / + - / + JW124 AK + / + - / +
JA096 Gre-0 + / + - / + JA197 Rd-0 + / + - / + JW128 Sap-0 + / + - / +
JA097 Gű-0 + / + - / + JA198 Ri-0 + / + - / + JW129 Sap-1 + / + - / +
JA098 Gy-0 + / + - / + JA202 Rsch-4 + / + - / + JW138 Kyoto-2 + / + - / +
JA099 Ha-0 + / + - / + JA203 Ru-0 + / + - / + JW142 WS + / + - / +
JA110 In-0 + / + - / + JA208 Se-0 + / + - / +
JA111 Is-0 + / + - / + JA213 Sf-2 + / + - / +
1) +/+, development of necrotic cell death and detection of viral CP in virus-inoculated leaves or non-inoculated upper leaves of each ecotype. 2) SASSC (Sendai Arabidopsis Seed Stock Center) number could be used for quick search in SASSC database (https://sassc.epd.brc.riken.jp).
38
2.4 Conclusion
CMV(H) could systemically spread in Col-0 without inducing any symptoms. In contrast, severe stunting and yellowing symptoms systemically developed in Col-0 in response to the infection of CMV(Y). For analyzing the viral factor which determined such different phenotypes, a series of reassortant CMVs were constructed as shown in Figure 1. During the analysis of the responses in Col-0 to the infection of each reassortant CMV, the induction of cell death was observed in 3 reassortant CMVs, which all contain RNA1 of CMV(H), inoculated Col-0 leaves (Figure 2A). Similar intensity of CP band and CMV RNA bands were detected in each reassortant CMV-inoculated leaves, suggesting that each reassortant CMV can accumulate in Col-0 at similar level, and in turn, the induction of cell death has nothing on virus multiplication. However, cell death is a crucial situation to the survival of virus since its reproduction is fully dependent on the host cell metabolism. Host often use cell death to defend the infection of pathogens, such as the HR cell death. Therefore, the cell death induced in reassortant CMV-inoculated Col-0 leaves was characterized through a comparative analysis with RCY1-mediated HR cell death. The cell death induced in CMV(HYY)-inoculated Col-0 was observed from 5 dpi whereas HR cell death was observed from 3 dpi in CMV(Y)-inoculated Col::RCY1 leaves (Figure 3). The virus multiplication in the virus-inoculated leaves was significantly suppressed during the induction of cell death because of the resistance conferred by RCY1 (Figure 2C). Instead, the induction of cell death in CMV(HYY)-inoculated Col-0 did not suppress multiplication of CMV(HYY), also synonymous with that in a susceptible interaction (Figure 2C and 4). Moreover, the systemic spread of CMV(HYY) was not affected by the induction of cell death. The systemic spread of CMV(HYY) induced yellowing and stunting symptoms on the non-inoculated upper leaves but not cell death. The differences to HR cell death were not just limited to the biochemical changes and symptom development. The global gene expression pattern between the cell death induced in CMV(HYY)-inoculated Col-0
39
leaves and HR cell death induced in CMV(Y)-inoculated Col::RCY1 also exhibits a massive difference. Not only the gene expression pattern between the two type of cell death, but also the GO enrichment analysis of each classes of DEGs indicated the differences existed between the two type of cell death. DEGs unique to the HR cell death were mostly enriched into defense related GO terms. However, the DEGs unique to the cell death induced in CMV(HYY)-inoculated Col-0 were mostly enriched into jasmonic acid related GO terms. Therefore, the cell death induced in CMV(HYY)-inoculated Col-0 leaves might be a form of necrotic cell death that does not contribute to the resistance to CMV. In conclusion, those results indicated that the cell death induced in the CMV(HYY)-inoculated Col-0 was a different type of cell death which is distinct to HR cell death in several aspects. It was referred to as necrotic cell death. Further, through analyzing the induction of necrotic cell death in 94 ecotypes of A. thaliana in response to CMV(HYY) infection, the result that 92 ecotypes of A. thaliana exhibit induction of necrotic cell death suggested that the induction of necrotic cell death was not specific to the interaction between CMV(HYY) and Col-0, but highly conserved in a broad range of A. thaliana ecotypes. Moreover, the result that 2 ecotypes of A. thaliana abolished the induction of necrotic cell death also suggested that putative plant factors might confer the induction of necrotic cell death through its interaction with CMV(HYY). They might be inactivated by mutation or disrupted by deletion in Stw-0 and Mt-0.
40
3 Analysis of the determinant region in CMV
RNA1 for induction of necrotic cell death in
virus-inoculated leaves
3.1 Introduction
In last chapter, the necrotic cell death was identified in the CMV(HYY)-inoculated Col-0 leaves. To further characterize it, the viral determinant for it in RNA1 of CMV(HYY) was analyzed. Since CMV(Y) did not induce cell death in Col-0, reassortant CMV between CMV(Y) and CMV(HYY) were employed to analyze the viral determinant for the induction of necrotic cell death in RNA1 of CMV(H). In the first part, the viral sequence in CMV RNA1 for induction of necrotic cell death in virus-inoculated leaves was analyzed. In the next part, single amino acid substitutions in CMV 1a protein for induction of necrotic cell death in virus-inoculated leaves were identified.
3.2 Materials and methods
3.2.1 Plant material
A. thaliana ecotypes Col-0 and Nicotiana benthamiana were used in the experiment. The plants were grown on soilless mix (Metro-Mix® 380, Sun Gro Horticulture, Agawam, MA) under a 14-h photoperiod (14000 lux) at 25°C in a KG-201 HL-D growth chamber (Koito, Yokohama, Japan).
3.2.2 RNA extraction and RT-PCR
RNA extraction and RT-PCR were performed according to the method described in the section 2.1.2
41
3.2.3 Construction of in vitro transcription vectors carrying
chimeric cDNA of CMV RNA1
In vitro transcription vectors carrying chimeric forms of the region encoding the 1a protein in the RNA1 cDNA of CMV(H) or CMV(Y) were constructed as described below to generate vectors Y-H/683, H-Y/683 (Figure 13A), Y-H/343 and Y-H/344~682 (Figure 15A) at first; and then vectors H/71, H/72~343, H/290, H/72~290, Y-H/71+291~343, and Y-H/291~343 (Figure 17A). All chimeric forms of the RNA1 cDNA region encoding 1a protein were generated by two-step PCR. First, the 5'-fragment of RNA1 cDNA and the 3'-fragment of RNA1 cDNA were amplified using CMV(H) or CMV(Y) cDNA as a template with the primers CMV RNA1-5'.FOR] (Table S2, S3) and an internal reverse primer based on the reverse-strand sequence of the junction site for chimeric constructs (Table S2, S3). Secondary PCR products were also amplified using an additional internal forward primer complementary to the reverse primer (Table S2, S4) and the CMV.RNA1-3'.REV primer (Table S2, S4) used in the primary PCR. In some instances, such as for vectors H/71, H/72~343, H/290, H/72~290, Y-H/71+291~343, and Y-H/291~343 (Figure 17A), tertiary internal fragments were also amplified by RT-PCR using another set of internal primers. The sets of internal PCR primers used are listed in Table S3.
In the second round of PCR, the resulting 5' and 3' fragments of RNA1 cDNA (and a third internal RT-PCR fragment, when necessary) amplified in the first PCR were used as templates to produce full-length chimeric RNA1 cDNA by PCR using the primers CMV.RNA1-5'.F and CMV.RNA1-3'.R (Table S1). All PCR products were purified using the Wizard® SV Gel and PCR Clean-Up System (Promega). The gel-purified RNA1 cDNA fragment was cloned into the HindIII and NotI sites of pUC118 (Takara Bio) with the In-Fusion HD Cloning System (Takara Bio) according to the manufacturer’s instructions. The nucleotide sequences of vector constructs carrying chimeric CMV RNA1 cDNAs were confirmed by Sanger sequencing on a CEQ8000 Automated DNA Sequencer
42
(Beckman Coulter).
3.2.4 Single amino acid substitution in 1a protein encoded on
RNA1 of CMV(Y)
Amino acid substitutions in 1a protein were performed by generating site-directed mutant cDNAs by nucleotide substitutions in pCY1-T7 using the GENEART Site-Directed Mutagenesis System (Thermo Fisher Scientific, Waltham, MA) with primers designed according to manufacturer’s instructions. The primers used for nucleotide substitution are shown in Table S4. All constructs were confirmed by Sanger sequencing on a CEQ8000 Automated DNA Sequencer (Beckman Coulter). In vitro transcription vectors carrying nucleotide substitutions in the 1a protein-coding region of the CMV(Y) RNA1 cDNA were designated T29A, I49V, G54S, R298Q, G299R, and H310N.
3.2.5 Production of reassortant CMV
The virus propagation was performed as described in the section 2.1.4.
3.2.6 Virus inoculation and detection
The virus inoculation and detection were performed as described in the section 2.1.5.
3.2.7 Detection of cell death
43
3.3 Results
3.3.1 Analysis of viral sequence in CMV RNA1 for induction of
necrotic cell death in virus-inoculated leaves
Induction of necrotic cell death in CMV(HYY)-inoculated Col-0 leaves but not in CMV(Y)-inoculated Col-0 leaves suggested that CMV(H) RNA1 is responsible for inducing necrotic cell death with co-infection of CMV(Y) RNA2 and RNA3 in virus-inoculated leaves (Figure 2A). There are 26 non-synonymous amino acid substitutions in CMV(H) and CMV(Y) RNA1 (Figure 12). To identify the region of 1a protein encoded by CMV(H) RNA1 responsible for inducing necrotic cell death in virus-inoculated Col-0 leaves, a series of chimeric cDNAs between CMV(Y) and CMV(H) RNA1 were generated and cloned under the control of the T7 promoter (Figures 13A, 15A, and 17A). Each infectious RNA1 was transcribed in vitro from each chimeric cDNA vector and combined with infectious RNA2 and RNA3 from CMV(Y) and used as inoculum.
Necrotic cell death developed in fully expanded Col-0 leaves inoculated with CMV(H-Y/683) containing infectious RNA1 transcribed from chimeric RNA1 cDNA vector H-Y/683, and in leaves inoculated with CMV(HYY) (Figure 13B). However, cell death did not occur in either leaves inoculated with CMV(Y-H/683), or in leaves inoculated with CMV(Y) (Figure 13B). However, CMV CP accumulated to similar amounts in CMV(H-Y/683)- and CMV(Y-H/683)-inoculated leaves (Figure 14). Systemic cell death was not observed in any upper leaves of either CMV(H-Y/683)- or CMV(Y-H/683)-inoculated plants , although the CMV CP was detected in upper leaves in similar amounts in CMV(H-Y/683) and CMV(Y-H/683) (Figure 14). These results suggest that the region of the 1a protein of CMV(H), which does not contain the helicase (HEL) domain, is necessary for the development of the necrotic cell death.
When fully expanded leaves of Col-0 were inoculated with CMV(Y-H/343) and CMV(Y-H/344~682), which contain chimeric regions from nucleotide positions 1127 to
44
2143 in the 1a protein-coding sequence due to reciprocal RNA1 cDNA fragment exchanges between CMV(Y) and CMV(H) (Figure 15A), necrotic cell death developed in the leaves inoculated with H/343), but not in those inoculated with CMV(Y-H/344~682) (Figure 15B). Systemic cell death was not observed in upper leaves of CMV(Y-H/344~682)- or CMV(Y-H/343)-inoculated plants , although the CMV CP was detected at similar levels in virus-inoculated leaves and non-inoculated upper leaves of these plants compared with CMV(Y)- and CMV(HYY)-inoculated plants as controls (Figure 16). Thus, the determinant for inducing necrotic cell death likely maps to the 5' region of RNA1, which corresponds to nucleotide positions 1 to 1126 in the 1a protein coding region and includes the methyltransferase (MET) domain.
A total of 11 amino acid substitutions remain in 1a protein between nucleotide positions 1 and 1126 of RNA1 of CMV(H) and CMV(Y) (Figure 12). Next, to further delimit the determinant that induces necrotic cell death, fully expanded leaves of A. thaliana Col-0 were inoculated with CMV(Y-H/71), CMV(Y-H/72~343), CMV(Y-H/290), CMV(Y-H/72~290), CMV(Y-H/71+291~343), or CMV(Y-H/291~343), which carries chimeric regions (Figure 17A) from nucleotide positions 1 to 310, 311 to 967, and 968 to 1126 in their 1a protein coding regions (Figure 12). Necrotic cell death developed in Col-0 leaves inoculated with H/71), H/72~343), H/290), H/71+291~343), or H/291~343), but not in those inoculated with CMV(Y-H/72~290) (Figure 17B). Systemic cell death was not observed in upper leaves of plants systemically infected with H/71), H/72~343), H/290), CMV(Y-H/72~290), CMV(Y-H/71+291~343), or CMV(Y-H/291~343) , although the CMV CP was detected in virus-inoculated leaves and non-inoculated upper leaves at similar levels (Figure 18). Thus, the development of the necrotic cell death seems to be determined by two independent regions of the 1a protein-coding region from nucleotide positions 1 to 310 or from 968 to 1126, which do not include the MET domain (Figures 12 and 17A). These two distinct 1a protein-coding regions, which equally determine the induction of the necrotic cell death, contain three amino acid differences between
45
CMV(Y) and CMV(H) (Figures 12 and 17A), respectively.
Figure 12. Schematic diagram of the CMV RNA1 encoding the 1a protein, which contains a methyltransferase (MET) and a helicase (HEL) domain. The 1a protein-coding region and corresponding 1a protein are shown as rectangles. Amino acids that differ between the 1a proteins encoded by the CMV(H) RNA1 (lower panel) and the CMV(Y) RNA1 (upper panel) and the positions of these amino acids in the 1a protein are described above each rectangle. The dotted lines connecting the two chimeric constructs indicate the amino acid and nucleotide positions of junction sites. The adjacent sequence numbers of each of these junctions are indicated above the CMV(Y) RNA1 (upper) schematic for amino acids; and above the CMV(H) RNA1 (lower) schematic for the corresponding nucleotides.
46
Figure 13. Induction of necrotic cell death in A. thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, CMV(H-Y/683) and CMV(Y-H/683), which carry chimeric 1a protein of CMV(Y) and CMV(H), and CMV(Y) and CMV(HYY). (A) Schematic diagram of the CMV RNA1 encoding the 1a protein. The 1a protein-coding region (and its corresponding 1a protein) is presented as rectangles in black for CMV(H) and in gray for CMV(Y). The dotted line indicates the junction site at amino acid position 682/683. MET, 1a protein methyltransferase domain; HEL, helicase domain. The presence (+) or absence (−) of necrotic cell death induction in virus-inoculated leaves is shown in the column on the right. (B) Responses of CMV-inoculated leaves for three independent biological replicates (plants number 1, 2, and 3). Virus-inoculated leaves to the left are under bright field and those to the right are stained with trypan blue.
Figure 3-2. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, CMV(H-Y/683) and CMV(Y-H/683)
47
Figure 14. Detection of CMV CP in extracts of virus-inoculated leaves and non-inoculated upper A. thaliana Col-0 leaves. Extracts from fully expanded leaves of three independent Col-0 plants (numbers 1, 2, and 3) inoculated with reassortant CMV: CMV(H-I/683) and CMV(Y-I/683) were analysed at 7 dpi using western blotting, as also were extracts from non-inoculated upper leaves of corresponding plants. RuBISCO protein is shown as an internal reference for protein quantity. CMV CP in CMV(Y)-inoculated leaves was quantified as a positive (P) control, and mock-inoculated leaves as a negative (N) control.
Figure 3-3. Detection of CMV coat protein in extracts of virus-inoculated leaves and non-inoculated upper Arabidopsis thaliana Col-0 leaves
48
Figure 15. Induction of necrotic cell death in A. thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, H/343) and CMV(Y-H/344~682) which carry chimeric 1a protein of CMV(Y) and CMV(H), and CMV(Y) and CMV(HYY). (A) Schematic diagram of the CMV RNA1 encoding the 1a protein (representation as in Fig.12: see legend for details). The junction sites in chimeric 1a proteins at amino acid positions 343/344 and 682/683 are indicated by dotted lines. (B) Responses of CMV-inoculated leaves (for details, see legend to Fig. 13).
Figure 3-4. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, CMV(Y-H/343) and CMV(Y-H/344~682)
49
Figure 16. Detection of CMV CP in extracts of virus-inoculated leaves and non-inoculated upper A. thaliana Col-0 leaves. Fully expanded leaves of three independent Col-0 plants (number 1, 2, and 3), which were inoculated with reassortant CMV(Y-H/343) or CMV(Y-H/344~682), were analysed at 7 dpi by western blotting, as also were extracts from non-inoculated upper leaves of the corresponding plants. RuBISCO protein is shown as an internal reference for protein quantity. Positive (P) and negative (N) controls quantified as explained in Fig. 14 legend.
Figure 3-5. Detection of CMV coat protein in extracts of virus-inoculated leaves and non-inoculated upper Arabidopsis thaliana Col-0 leaves
50
Figure 17. Induction of necrotic cell death in A. thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, H/71), CMV(Y-H/72~343), CMV(Y-H/290), CMV(Y-H/72~290), CMV(Y-H/71+291~343) and CMV(Y-H/291~343), which carry chimeric 1a protein of CMV(Y) and CMV(H), and CMV(Y) and CMV(HYY). (A) Schematic diagram of the CMV RNA1 encoding the 1a protein (representation as in Fig.12: see legend for details). Dotted lines indicate the chimera junction sites at amino acid positions 71/72, 290/291, and 343/344. (B) Responses of CMV-inoculated leaves (for details, see legend to Fig. 13).
Figure 3-6. Induction of necrotic cell death in Arabidopsis thaliana ecotype Col-0 leaves inoculated with reassortant CMVs, CMV(Y-H/71), CMV(Y-H/72~343), CMV(Y-H/290),
51
Figure 18. Detection of CMV CP in extracts of virus-inoculated A. thaliana Col-0 leaves and non-inoculated upper leaves. (A) Fully expanded leaves of three independent Col-0 plants (numbers 1, 2, and 3) inoculated with reassortant CMVs. H/71), H/72~343), H/290), H/72~290), CMV(Y-H/71+291~343), and CMV(Y-H/291~343) were analysed at 7 dpi by western blotting. (B) Non-inoculated upper leaves of the corresponding plants were analysed at 7 dpi by western blotting. RuBISCO protein is shown as an internal reference for protein quantity. Positive (P) and negative (N) controls quantified as explained in Fig. 14 legend.
Figure 3-7. Detection of CMV coat protein in extracts of virus-inoculated Arabidopsis thaliana Col-0 leaves and non-inoculated upper leaves