Tel81972322125.Fax81972322293.Emailnariaki23@affrc.go.jp
クエ Epinephelus bruneus 稚魚の成長に及ぼす飼育塩分の影響
井 上 誠 章,
1岩 崎 隆 志,
2嶋 田 幸 典,
1佐 藤
純,
1西 岡 豊 弘
1 (2015 年 4 月 15 日受付,2015 年 6 月 17 日受理) 1国立研究開発法人水産総合研究センター増養殖研究所上浦庁舎 2国立研究開発法人水産総合研究センター西海区水産研究所八重山庁舎EŠect of salinity on the growth of juvenile longtooth grouperEpinephelus bruneus in tank culture
NARIAKIINOUE,1TAKASHIIWASAKI,2YUKINORISHIMADA,1
JUNSATOH1AND TOYOHIRONISHIOKA1
1Kamiura Laboratory, National Research Institute of Aquaculture, Fisheries Research Agency, Saiki, Oita 872
2602,2Yaeyama Laboratory, Seikai National Fisheries Research Institute, Fisheries Research Agency, Ishigaki,
Okinawa 9070451, Japan
Commercially viable farming of the longtooth grouper Epinephelus bruneus has been impeded by the fact that it takes at least 3 years for cultured groupers to attain a marketable size. This is due to growth stagnation at low temperatures in winter. It has been reported that the growth and condition of some marine ˆsh is enhanced at low salinities; therefore, we evaluated the eŠect of low salinity on the growth of tank-reared juvenile groupers using a generalized linear model. The results demonstrated that growth increased at 17 psu during the feeding period, and body weight loss during the non-feeding period was lower at a salinity of 10 psu compared to 34 psu.
キーワードEpinephelus bruneus,GLM,クエ,浸透圧,成長,低塩分海水 クエ Epinephelus bruneus は,本州中部以南から東シ ナ海までの沿岸の岩礁域に分布する大型のハタ類であ る。1,2)クエの商品価値はきわめて高く,種苗生産技術は 確立されつつあることから本種は新たな養殖対象魚とし て注目されている。3)しかし,クエは水温が 1820°C 以 下に低下する冬季に成長停滞が起こり,4)そのため出荷 までに少なくとも 3 年以上の飼育期間を必要とし,こ れがクエ養殖の普及を妨げる要因の一つとなっている。 この問題の解決には,種苗期以降の飼育水温,飼育塩分 および摂餌量と成長量との関係といった成長に関連する 飼育条件の知見は不可欠である。しかし,これまでクエ 養殖に関連する研究は,親魚養成や種苗生産技術に関す る研究が中心に進められており,5)種苗期以降の中間育 成の期間の飼育条件に関する知見は少ない。 種苗生産の対象とされる海産魚において,種苗生産や それに続く中間育成は多くの場合,希釈等を行わない通 常海水で行われる。6)しかし,自然環境下では海産魚は 一般にその生活史の初期において干潟域や河口域などを 育成場とし,そのため稚魚期の成長は淡水と海水の中間 的な塩分で最大となる場合が多い。7)天然海域における クエの初期生活史は詳しくは明らかにされていないが, クエ稚魚は塩分変動が激しい沿岸の漁港内や岩礁のタイ ドプールなどに出現することが知られている。2)種苗生 産や中間育成における低塩分海水による飼育の有効例と して,カサゴ Sebastiscus marmoratus では日齢 1540 ま でに生じる大量減耗が軽減されることや,8)トラフグ Takifugu rubripesでは多くの場合,成長量の増大が起 こるという報告がある。6,911) またクエと同じハタ科魚類であるキジハタ Epinephe-lus akaaraの稚魚についても1926 psu の低塩分海水に
よる飼育で成長量が増大することが報告されている。12) 本研究ではクエ稚魚の成長量の改善を目指し,成長に及 ぼす飼育塩分の影響を明らかにすることを目的とした。 材料と方法 予 備 飼 育 2013 年 6 月 6 日 に メ ス 3 個 体 か ら 採 卵 し,これにオス 6 個体から採精した精子を人工授精さ せて作出した日齢 77 のクエ稚魚を,8 月 23 日に室内
Fig. 1 Daily food consumption (DFC) of juvenile Epinephelus bruneus in relation to water temperature and salinity during the feeding period (from 16 May to 13 June 2014). Solid line with open circles, water temperature; gray, dotted and solid lines, average DFC at 10, 17 and 34 psu, respectively; areas with and without shadows, periods at which water temperature was below and above 20°C, respectively.
60 t 水槽に収容して日齢 335 まで予備飼育した。飼育 海水には砂ろ過海水を電解殺菌処理したものを,温度調 整を行わず注水量 750 L/h の流水条件で用いた。飼料 はクエ稚魚の口径に合わせた市販のドライペレット(日 清丸紅飼料,おとひめ EP14)を用いて週 5 日,体重 の 12程度を一回給餌した。 馴致飼育 飼育試験を行う前に以下の手順でクエ稚魚 を 3 段階の塩分に馴致した。まず 500 L ポリエチレン 水槽 3 基にろ過殺菌海水を 300 L(水温 17.4°C,塩分 34.2 psu)を注水し,これに日齢 335 のクエ稚魚(体重 25.8 ± 6.8 g , 全 長 120.8 ± 10.9 mm ) を 30 個 体 ず つ 2014 年 5 月 8 日に収容した。つぎに,3 水槽のうち 1 水槽の飼育水の塩分は変化させず,残り 2 水槽には水 道水を少量ずつ注入して 5 月 12 日まで 4 日間かけて徐 々に両水槽の塩分をそれぞれ塩分 17 psu および塩分 10 psuまで低下させ,その後 5 月 16 日まで馴致した。馴 致の期間中,すべての水槽の水温は室温として換水は行 わず,通気はセラミック製エアストーン(いぶきエアス トーン,23×100 mm)1 個を用いて通気量 6 L/min に 調整し,飼料は市販のドライペレット(日清丸紅飼料, おとひめ EP4)を用いて体重の 12 程度を毎日一回 給餌した。 飼育試験 まず,ろ過殺菌海水を水道水で希釈するこ とにより塩分 34, 17 および 10 psu に調整した海水を各 塩分区 3 水槽ずつ,合計 9 基の 100 L ポリエチレン製 水槽に 80 L ずつ注水した。次に,馴致飼育したクエ稚 魚から各塩分それぞれ任意の 15 個体を選別し,FA100 (DS ファーマアニマルヘルス社)を用いて麻酔し全長 と体重を計測した後に PIT タグ(BIOMARK 社)を装 着して個体識別した。最後に,各塩分区の水槽に個体識 別したクエ稚魚を 5 個体ずつ,すなわち 3 塩分区×3 水 槽×5 個体の合計 45 個体を 5 月 16 日に収容し 7 月 22 まで飼育した。飼育試験中,各水槽はセラミック製エア ストーン(いぶきエアストーン,30F 丸)1 個を用いて 通気し(0.91.5 L/min),室温で塩分 34, 17 および 10 psuに調整した海水を用いて日間 15 の換水(12 L/ day)を行った。また,本試験の事前検討により各水槽 の水温は室温とほぼ一致することがわかっており,その ため室温をもって飼育水温とした。 飼育試験中,5 月 16 日から 6 月 13 日までの 28 日間 は各水槽に給餌を行い,6 月 14 日から 7 月 22 日までの 38 日間は無給餌とした。給餌期間中の飼料にはおとひ め EP4 を用いて,各水槽に毎日 40 粒(0.05 g/grain) を給餌し,翌日にサイフォンを用いて底掃除をするとと もに残餌を計数して各水槽の日間摂餌量[g/(ˆsh・day)] を求めた。給餌期間中は 5 月 16 日(日齢 343),5 月 23 日 ( 日 齢 350 ), 5 月 30 日 ( 日 齢 357 ), 6 月 8 日 (日齢 366)および 6 月 13 日(日齢 371),無給餌期間 中は 6 月 20 日(日齢 378),6 月 27 日(日齢 385),7 月 4 日(日齢 392),7 月 11 日(日齢 399)および 7 月 18 日(日齢 406)の合計 10 回,全長と体重の計測を FA100 を用いた麻酔下で行った。 7 月 18 日の計測後さらに 4 日間,上記同様の無給餌 条件で飼育を続け,7 月 22 日に以下の方法によりクエ 稚魚の全血の浸透圧を測定した。まず,各水槽のクエ稚 魚 5 個体のうち任意の 23 個体,すなわち各塩分区そ れぞれ 89 個体ずつ選別する。次に,これらのクエ稚 魚からそれぞれ,ヘパリン処理した 25 ゲージの注射器
Table 1 Body weight (BW) and total length (TL) of juvenile Epinephelus bruneus during the feeding and non-feeding periods at three salinity levels
Date Rearingdays Tank 10 psu 17 psu 34 psu
BW (SD) TL (SD) BW (SD) TL (SD) BW (SD) TL (SD) Feeding 16 May 0 1 31.7 5.9 130.8 6.8 23.4 7.5 115.6 12.6 20.9 6.9 112.6 13.0 2 25.4 8.7 120.2 11.7 27.1 7.0 122.0 9.2 24.6 6.0 119.4 12.1 3 26.5 4.5 125.0 3.8 27.8 6.0 123.8 8.2 24.6 7.5 118.0 13.5 Ave. 27.9 125.3 26.1 120.5 23.4 116.7 23 May 7 1 32.3 6.3 131.6 7.1 24.2 7.3 117.4 14.9 20.7 6.9 115.4 14.0 2 25.7 8.4 123.8 12.5 28.2 7.1 124.6 12.0 24.8 6.3 120.2 11.7 3 26.2 3.9 125.0 3.8 27.8 5.9 125.2 7.8 24.1 6.1 120.0 14.1 Ave. 28.1 126.8 26.7 122.4 23.2 118.5 30 May 14 1 34.2 6.7 136.4 7.8 26.0 7.4 123.6 13.9 20.5 6.5 116.2 12.4 2 27.3 9.7 137.8 21.5 29.2 7.7 126.8 10.2 26.0 6.9 123.6 12.4 3 27.4 4.0 129.0 5.7 29.2 5.6 125.4 9.2 24.5 7.4 120.8 14.4 Ave. 29.6 134.4 28.1 125.3 23.7 120.2 8 Jun 23 1 36.8 7.4 140.4 9.1 27.8 7.6 124.8 13.1 23.2 6.2 122.2 11.3 2 29.1 10.6 130.2 14.5 31.6 8.0 133.4 13.1 28.3 7.3 126.0 11.0 3 29.0 3.7 130.6 4.3 32.1 5.5 131.8 6.5 26.8 7.2 124.2 12.5 Ave. 31.6 133.7 30.5 130.0 26.1 124.1 13 Jun 28 1 37.6 7.9 140.6 8.6 28.8 7.8 126.8 12.5 24.0 6.4 123.2 11.5 2 29.8 11.1 131.0 14.6 33.0 8.0 132.8 12.0 28.9 7.2 126.8 10.7 3 29.5 4.2 130.0 5.1 32.8 5.6 133.8 7.8 26.8 7.9 125.2 13.4 Ave. 32.3 133.9 31.5 131.1 26.6 125.1 Non-feedting 20 Jun 6 1 36.7 6.7 140.6 9.0 27.9 7.4 127.0 12.2 23.1 6.2 122.6 11.4 2 29.0 10.9 131.2 14.6 31.9 7.8 132.6 11.1 27.8 7.0 127.4 10.1 3 28.8 4.3 131.8 5.0 31.4 5.2 133.6 6.8 25.7 7.7 126.2 13.4 Ave. 31.5 134.5 30.4 131.1 25.5 125.4 27 Jun 13 1 35.4 6.7 140.2 8.2 26.7 7.3 128.4 12.4 22.0 6.6 118.8 12.9 2 27.9 11.1 131.6 14.6 30.9 7.7 134.8 10.6 27.0 7.5 127.4 10.6 3 27.6 4.2 132.2 5.1 30.6 5.5 134.2 6.7 25.0 7.6 124.8 13.5 Ave. 30.3 134.7 29.4 132.5 24.7 123.7 4 Jul 20 1 34.9 6.5 137.8 7.2 26.3 6.9 128.2 12.7 21.1 6.2 121.2 10.5 2 27.2 10.8 130.2 16.2 30.6 7.6 134.2 10.8 26.1 7.4 104.6 52.2 3 26.7 4.1 130.8 5.9 29.8 5.3 134.4 6.6 24.2 7.4 123.6 12.4 Ave. 29.6 132.9 28.9 132.3 23.8 116.5 11 Jul 27 1 34.3 6.7 143.0 8.6 24.9 6.9 131.2 9.5 20.5 5.9 123.2 12.0 2 26.9 10.6 133.6 16.6 30.2 7.7 136.6 11.2 25.3 7.0 128.0 11.5 3 26.3 4.3 133.2 6.5 29.5 5.3 137.0 6.2 23.5 7.1 127.0 12.9 Ave. 29.2 136.6 28.2 134.9 23.1 126.1 18 Jul 34 1 33.5 6.6 144.6 8.5 25.1 7.2 129.2 13.3 19.8 5.6 121.4 10.7 2 25.9 10.4 133.0 17.3 29.2 7.6 134.0 11.0 25.2 7.0 128.2 10.8 3 25.5 4.8 132.8 5.9 29.8 4.6 136.8 7.4 23.0 7.0 125.8 13.3 Ave. 28.3 136.8 28.0 133.3 22.7 125.1 SD, standard deviation
Fig. 2 Fold changes in body weight of juvenile Epinephe-lus bruneus during the feeding (from 16 May to 13 June 2014) and non-feeding (from 14 June to 18 July 2014) periods in relation to three salinity levels of 10 (ˆlled circles ), 17 (open circles ) and 34 psu (open trian-gles ). を用いて尾 柄部大動脈よ り 610 mL 採血する。最 後 に,各個体の全血の浸透圧を VAPRO オズモメーター 5520 型(WESCOR)を用いて蒸気圧法により計測した。 また,計測は各個体とも 2 回ずつ行い,その平均値を 各個体の全血の浸透圧とした。これに加えて,飼育に用 いたのと同様のろ過殺菌海水を水道水で塩分 34.20.0 psuまでの 9 段階の濃度に希釈し,上記同様の方法でこ れらの浸透圧を計測した。 一般化線形モデル(GLM)解析 給餌および無給餌 期間中の体重変化に対する飼育日数,飼育塩分および各 期間の初期体重とそれぞれの交互作用の影響を評価する ために,各期間の開始時データを除外したデータセット を用いて一般化線形モデル(GLM)解析を行った。給 餌または無給餌期間ともに,対数変換した体重[log (BW)]を応答変数とし,飼育日数(Day ),飼育塩分 (Sal )および対数変換した初期体重[log(IniBW )]を 説明変数として二次の交互作用を考慮した GLM を仮定 した。また,応答変数の確率分布は正規分布を仮定し, 飼育塩分の効果については塩分 34 psu 区での飼育を基 準(b3=0)とした。両期間の採択モデルの選択は以下 の手順によって行った。まず,説明変数の組み合わせ全 てを用いたモデルを作成し,それぞれのモデルにおける 赤池情報量基準(Akaike's Information Criterion: AIC) を算出する。それらのモデルを AIC が小さい順に並び 替え,AIC が最小になるモデルから AIC が急増するま でのモデルまでを採択モデルの候補とし抽出した。最後 に,それらのモデルの中で常に選択される説明変数を持 つモデルを採択モデルとし,各要因によってどのように 体重が変化するか解析した。本研究で使用したすべての 統計検定,モデル推定および各モデルの AIC の算出に は Windows 版の R3.0.1 を用いた。12) 結 果 給餌期間中,各水槽の水温は実験開始時(5 月 17 日) の水温 18.5°C から上昇し 5 月 25 日に水温 20°C を超え て 6 月 10 日以降は水温 22.0°C 以上に(Fig. 1),その 後の無給時期間中も飼育水温は上昇し続け飼育試験の終 了時の 7 月 18 日には水温 24.0°C 以上となった。水温が 20°C 以上となる 5 月 25 日以降の 3 塩分区の日間摂餌量 の変動様式は類似するが(Fig. 1),一方で水温が 20°C 以下の期間中(5 月 1625 日),塩分 34 psu 区の日間摂 餌量は両塩分区と比較して有意に小さかった(One-way ANOVA with Tukey's test, p<0.01; Fig. 1)。
飼育試験中の各水槽の平均体長および平均体重を Ta-ble 1に,給餌および無給餌期間中の体重変化を Fig. 2 に示した。給餌期間中,塩分 17 psu 区の体重は開始直 後から増加し始め,31 日間の給餌期間で初期体重の 1.22 倍となった。一方で,塩分 34 psu 区および塩分 10 psu 区では給餌期間中に体重の増加は認められたもの の,増加率はそれぞれ初期体重の 1.14 および 1.15 倍と 塩分 17 psu 区と比較して小さかった。また,塩分 34 psu区では,飼育試験開始から 5 月 23 日までの水温 20 °C 以下の期間中には,他の 2 塩分区と異なって,体重 増加がほとんど認められなかった。給餌期間中の体重変 化に対する飼育日数,飼育塩分および初期体重とそれぞ れの交互作用の影響を評価するため GLM 解析を行っ た。その結果,採択モデル候補として 4 つのモデルが 抽出され,これらのモデルの中で常に選択される説明変 数は飼育日数,飼育塩分,初期体重および初期体重と飼 育塩分の交互作用であったことから,これらを含むモデ ルを採択し,各要因による体重変化を検討した(Table 2)。採択モデルの残差の正規性は QQ プロットにより 確認し(図省略),説明変数の係数および分散分析によ る尤度比検定の結果をそれぞれ Table 3 および Table 4 に示した。採択モデルに用いられたすべての説明変数と 交互作用はワルド検定により有意と判定され(Table 3),また尤度比検定よるヌルモデルとの比較において もすべて有意と判定された(Table 4)。得られた採択モ
Table 2 Factor selection for generalized linear models (GLM) for changes in the body weight (BW) of juvenile Epinephelus bruneus during the feeding and non-feeding periods
GLM model df AIC DAIC Intercept (b0) Day (b1) log(IniBW ) (b2) Sal (b3) Day:log(IniBW ) (b4) Day:Sal (b5) log(IniBW ):Sal (b6) Feeding Model 1
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b4Day:log(IniBW )
+b6log(IniBW ):Sal+e
9 926.51 0.00 + + + + + +
Model 2 (Optimal model)
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b6log(IniBW ):Sal
+e
8 926.61 0.10 + + + + +
Model 3
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b5Day:Sal
+b6log(IniBW ):Sal+e
10 927.51 1.00 + + + + + +
Model 4(Full model)
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b4Day:log(IniBW )
+b5Day:Sal
+b6log(IniBW ):Sal+e
11 927.71 1.20 + + + + + + +
Model 5
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+e
6 942.31 15.80 + + + NULL model 179 1518.74 592.23
Non-feeding Model 1
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b4Day:log(IniBW )
+b6log(IniBW ):Sal+e
9 782.20 0.00 + + + + + +
Model 2 (Optimal model)
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b4Day:log(IniBW )
+e
7 783.50 1.30 + + + + +
Model 3(Full model)
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b4Day:log(IniBW )
+b5Day:Sal
+b6log(IniBW ):Sal+e
11 785.00 2.80 + + + + + + +
Model 4
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b4Day:log(IniBW )
+b5Day:Sal+e
9 786.40 4.20 + + + + + +
Model 5
log(BW)=b0+b1Day+b2log(IniBW )
+b3Sal+b5Day:Sal
+b6log(IniBW ):Sal+e
10 793.30 11.10 + + + + + +
NULL model 223 1834.70 1052.50 +, Using factors for GLM analysis. Shaded models are optimal models.
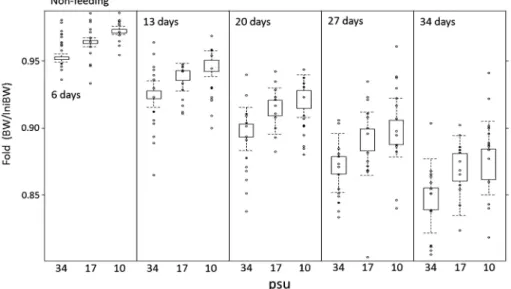
デルを用いて体重の予測値を計算して初期体重からの変 化率(BW/IniBW)を算出し,これと実際の観測値の 結果を合わせて Fig. 3 に示した。採択モデルによる各 塩分区の予測値は実際の体重変化を妥当に表現できてお り(Fig. 3),各説明変数の係数より以下の点を推察で きる。すなわち,給餌期間中,クエ稚魚の体重は飼育日 数(Day; b1=0.007)にともない増加するが,体重増加 に対して初期体重[log(IniBW); b2<1]は負の効果が (初期体重の効果は一般的な係数の解釈とは異なり,1 未満であれば負の効果を持つ),また塩分 34 psu 区と比 較して塩分 17 psu 区での飼育は正の効果が(Sal 17;b3 =0.313),一方で塩分 10 psu 区での飼育は負の効果を 持つ(b3=-0.280)。加えて,初期体重と飼育塩分の交 互作用[log(IniBW ): Sal]より,体重増加に対して初 期体重が及ぼす効果は飼育塩分によって異なる。 無給餌期間中,各塩分区の体重は無給時開始の直後か ら減少し始め,6 月 14 日~7 月 18 日までの 34 日間で 塩 分 34, 17 お よ び 10 psu 区 で は そ れ ぞ れ 初 期 体 重 の 0.85, 0.87 および 0.88 倍に減少した。給餌期間中と同 様の方法によりモデル選択を行った結果,飼育日数,飼 育塩分,初期体重および飼育日数と初期体重の交互作用 を説明変数に持つモデルが採択モデルとなった(Table
Table 3 Parameter estimates and standard errors (SE) in the optimal generalized linear models (GLM) ˆtted to the data for juvenile Epinephelus bruneus during the feeding and non-feeding periods
Estimate SE Feeding (Intercept) 0.122 0.075 Day 0.007 0.000 log(IniBW) 0.940 0.024 Sal17 0.313 0.110 Sal10 -0.280 0.122 log(IniBW):Sal17 -0.078 0.034 log(IniBW):Sal10 0.093 0.038 Non-feeding (Intercept) 0.3969 0.1227 Day -0.0124 0.0023 log(IniBW) 0.9124 0.0363 Sal17 0.0114 0.0045 Sal10 0.0200 0.0045 Day:log(IniBW) 0.0025 0.0007 p<0.05, p<0.01, p<0.001 via Wald's test
Table 4 Likelihood ratio test by ANOVA of the optimal generalized linear models (GLM) for changes in the body weight (BW) of juvenile Epinephelus bruneus during the feeding and non-feeding periods
df SS F Feeding Day 1 0.618 237.508 Sal 2 1.439 276.663 log(IniBW) 1 10.344 3977.504 log(IniBW):Sal 2 0.052 9.974 Residuals 173 0.003 Non-feeding Day 1 0.327 478.69 Sal 2 2.244 1641.92 log(IniBW) 1 14.277 20889.96 Day:log(IniBW) 1 0.009 13.43 Residuals 218 0.149
df, Degree of freedom; SS, sum of squares; F, F-value; p<0.001
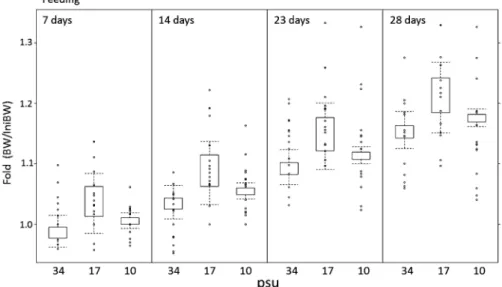
Fig. 3 Observed (open circles ) and predicted (box-plots) changes in body weight of juvenile Epinephelus bruneus during the feeding period (from 16 May to 13 June 2014) in relation to three salinity levels. Predicted changes of body weight were simulated us-ing an optimal GLM model. Open columns of box-plots indicate middle 50 of the predicted dataset, and the upper-and lower-most areas of the columns indicate 75 and 25 of the predicted dataset, respectively. Upper and lower vertical lines (whiskers) on the columns present minimal and maximal values equivalent to 1.5 times the inter-quartile range, respectively. 2)。採択モデルに選択されたすべての説明変数と交互 作用はワルド検定により有意と判定され(Table 3),ま た尤度比検定によるヌルモデルとの比較においてもすべ て有意と判定された(Table 4)。得られた採択モデルを 用いて体重の予測値を計算して初期体重からの変化率 (BW/IniBW)を算出し,これと実際の観測値の結果と を合わせて Fig. 3 に示した。採択モデルによる各塩分 区の予測値は実際の体重変化を妥当に表現できており (Fig. 4),各説明変数の係数より以下の点が推定でき る。すなわち,クエ稚魚の体重は無給餌での飼育日数 (Day;b1=-0.0124)にともない減少し,塩分 34 psu 区 と比較して塩分 17 psu 区(Sal 17; b3=0.0114)および 塩分 10 psu 区(Sal 10; b3=0.02)での飼育は体重増加 に対して正の効果(=体重減少を抑制する)を持つが, その効果は塩分 10 psu 区のほうが大きい。また無給餌 飼 育 の 開 始 時 は 体 重 増 加 に 対 し て 初 期 体 重 [ log
Fig. 4 Observed and predicted changes in body weight of juvenile Epinephelus bruneus at three salinity levels during the non-feed-ing period (from 14 June to 18 July 2014). Predicted changes of body weight were simulated usnon-feed-ing an optimal GLM model. Symbols are the same as in Fig. 3.
(IniBW ): b2<1]は負の効果(=体重減少を増大する) を持つが,一方で飼育日数と初期体重の交互作用[Day: log(IniBW );b4=0.0025]より,飼育日数が増加するに つれて初期体重の体重増加に対する正の効果が増大す る。すなわち,無給餌飼育の開始直後は初期体重の大き な個体ほど体重減少が起こるが,一方で無給餌期間が長 くなるにつれてこの関係は逆転し,初期体重の大きな個 体ほど体重減少は小さくなる。 塩分 34, 17 および 10 psu 区の飼育魚の全血の浸透圧 はそれぞれ 356.6±33.2 mOsm/kg, 305.4±16.3 mOsm/ kg および 295.2±21.3 mOsm/kg であり,塩分 17 psu 区および塩分 10 psu 区と比較して塩分 34 psu 区は有意 に高かった(One-way ANOVA with Tukey's test, p< 0.01)。また,塩分 34.20.0 psu の希釈ろ過海水の浸透 圧は次式y=28.68x+10.30 (R2=0.99, p<0.01)で直 線回帰でき,本回帰式と各塩分区の全血の測定結果か ら,塩分 10 psu 区の飼育魚の全血と飼育海水の浸透圧 (297.1 mOsm/kg)はほぼ一致した。 考 察 給餌期間中,塩分 34 psu 区と比較して,塩分 17 psu 区での飼育はクエ稚魚の体重の増加量を増大させる効果 があり,一方で塩分 10 psu 区での飼育は体重の増加量 を減少させる効果が認められた(Table 3, Fig. 3)。本 研究と同様に,低塩分条件下で飼育することにより,ト ラフグ稚魚の成長量の増大が多くの場合に起こることが 報告されている。6,911)またヒラメ Paralichthys olivaceus 稚魚についても,通常海水での飼育と比較して 50 海 水での飼育を行うことにより摂餌量および餌料効率を増 大させ,その結果,成長量を増大させることが報告され ている。14)一般に,海産魚では環境水の浸透圧が体液の 約 3 倍であるため,魚体内は脱水されると同時に塩が 流入する。そのため海産魚は多量の海水を飲み,腸から Na+お よ び Cl-な ど の 1 価 イ オ ン と と も に 水 を 吸 収 し,過剰となった塩は主に鰓から排出して体液の浸透圧 を海水の約 1/3 の一定の範囲に保つが,これには他の 代謝と同様にエネルギー消費を伴う。15)Kikuchi et al.16) はトラフグ稚魚が低塩分で高成長を得られる機構の仮説 として浸透圧調節にかかわるエネルギー量の節約を挙げ ているが,これまでにこの機構を検証した例はほとんど なく,飼育海水の塩分濃度が成長に影響する機構は明ら かになっていない。10) 無給餌期間中の飼育結果より,塩分 17 psu 区および 塩分 10 psu 区での飼育は,塩分 34 psu 区での飼育と比 較して無給餌による体重減少を抑制する効果があり,さ らにその効果は塩分 17 psu 区よりも塩分 10 psu 区のほ うが大きい(Table 3, Fig. 4)。加えて,塩分 34 psu 区 のクエ稚魚の全血の浸透圧は塩分 10 psu 区および塩分 17 psu 区よりも有意に高く,塩分 10 psu 区において飼 育海水の浸透圧と飼育魚の全血の浸透圧がほぼ一致し た。これらの結果は,体液の浸透圧に近い低塩分海水を 用いて飼育を行うことが浸透圧調節にかかわるエネル ギーの節約につながるという上記の Kikuchi et al.16)の 仮説を部分的に支持する。しかし,浸透圧調節のエネル ギー節約のみでクエ稚魚の成長量の増大が説明できるの であれば,給餌期間中の体重の増加量は塩分 10 psu 区 のクエ稚魚が最も大きくなるはずである。しかし,実際 は体 重 増 加 に 対 す る効 果 は 塩 分 17 psu 区 が 最 も大 き
く,塩分 10 psu 区での飼育は塩分 34 psu 区よりも体重 の増加量を減少させる効果があった(Table 3, Fig. 3)。 キジハタ稚魚(体重 190 g)の場合,12)塩分 1926 psu での飼育により成長量の増大が起こるが,恐らくはこの 場合においても飼育塩分の浸透圧は飼育魚の全血より高 い。これらの点からみれば,低塩分飼育における成長量 の増大の機構には,浸透圧調節にかかわるエネルギー節 約以外の要因も含まれると推察できる。低塩分海水によ る飼育が成長量や摂餌量等に与える影響は魚種や発育段 階によって大きく異なることが示されており,7,17)さら に同一の魚種および成長段階においても飼育水槽の形状 やサイズ,収容密度や飼育水温,飼育期間などの各種飼 育条件の違いにより低塩分海水での飼育が成長に与える 影響は異なる。10)今後,クエ稚魚において低塩分飼育に より成長量が増大された理由を明らかにするためには, エネルギー収支を考慮したより詳細な飼育実験が必要で あろう。 クエ稚魚の日間摂餌量は飼育水温に影響され,水温 1820°C 以下では極端にその量が減少することが知られ ている。3,4)本研究では,水温 20°C 以上の期間において は 3 塩 分 区 と も に 日 間 摂 餌 量 の 変 動 様 式 は 類 似 し た が,一方で水温 20°C 以下の場合,塩分 10 psu 区および 塩分 17 psu 区の日間摂餌量は塩分 34 psu 区と比較して 有意に大きかった(Fig. 1)。この結果より低塩分での 飼育は低水温期における日間摂餌量を増大させる効果を もつ可能性が考えられた。 井上ら3)によれば,クエ養殖の効率化には冬季の稚魚 の成長量の向上が重要であり,そのためには種苗期以降 の低水温期までの期間になるべく魚体重を増加させるよ うな飼育手法の開発が必要である。本研究では,塩分 17 psu という低塩分での飼育が種苗期以降の成長量お よび冬季の摂餌量の増大につながる可能性を示唆でき た。またクエ稚魚は水温 14°C以下となる冬季の低水温 期では摂餌が認められなくなり体重減少が起こるが,3) 塩分 10 psu で飼育することにより低水温期の体重減少 を抑制できる可能性がある。これらの結果は種苗期以降 のクエ稚魚の成長促進につながる知見であり,今後クエ 養殖における飼育技術の改良を目指すにおいて重要であ ろう。 謝 辞 本研究を遂行し論文としてまとめるに当たり,ご尽力 いただいた国立研究開発法人 水産総合研究センター増 養殖研究所 上浦庁舎の職員各位に厚くお礼申し上げ る。本研究の GLM 解析を行うに当たり,有益なご助言 をいただいた東京農業大学 生物産業学部 アクアバイオ 学科の金岩稔准教授に感謝の意を表する。 文 献 1) 瀬能 宏.ハタ科.「日本産魚類検索」(中坊徹次編)東 海大学出版会,東京.1993; 601632. 2) 小島純一.マハタ属クエ.「日本産稚魚図鑑」(沖山宗雄 編)東海大学出版会,東京.1988; 416417. 3) 井上誠章,岩崎隆志,加治俊二.低水温期におけるクエ Epinephelus bruneus と マ ハ タ E. septemfasciatus の 成 長 量および摂餌量の変化.日本水産学会誌2014;80: 56 58. 4) 岡田貴彦,米島久司,向井良夫,澤田好史.人工孵化ク エ稚魚の環境ストレス耐性について.近畿大学水産研究 所報告1996;5: 139146. 5) 中川雅弘,堀田卓朗,吉田一範,服部圭太.配合飼料の 給餌量の違いがクエEpinephelus bruneus 種苗の成長と生 残に及ぼす影響.水産増殖 2011;59: 579584. 6) 韓 慶男,荘 恒源,松井誠一,古市政幸,北島 力. トラフグ幼稚魚の成長,生残,および飼料効率に及ぼす 飼育水塩分の影響.日本水産学会誌 1995;61: 2126. 7) Bœuf G, Payan P. How should salinity in‰uence ˆsh
growth? Comp. Biochem. Physiol. Part C 2001;130: 411 423. 8) 御堂岡あにせ.低塩分飼育法による種苗生産技術の開 発初期減耗の低減に有効,漁獲魚の蓄養にも.アクア ネット2012;15: 4245. 9) 神谷直明,辻ヶ堂諦.トラフグ仔稚魚に対する塩分の影 響.栽培漁業技術開発研究 1995; 23: 113115. 10) 多賀 真,山下 洋.トラフグ仔稚魚の成長における低 塩分の有効性とその要因.水産増殖 2011;59: 225233. 11) 片山貴士,森田哲男,今井 正,山本義久.閉鎖循環シ ステムを用いた低塩分条件下でのトラフグ量産飼育.水 産技術2013;5: 165169. 12) 森田哲男.キジハタの循環式陸上養殖事業化への可能 性.月刊養殖ビジネス 2014;12: 1416.
13) R Development Core Team: R. A language and environ-ment for statistical computing. In: R Foundation for Statistical Computing. Vienna. 2009.
14) 斎藤節男,佐々木睦子,李 海,清水幹博,山田寿 郎.ヒラメ稚魚の成長と代謝に及ぼす低塩分環境の影 響.北海道水産試験場研究報告 1990; 34: 18. 15) 岩田宗彦,平野哲也.浸透圧調節.「魚類生理学」(板沢 靖男,羽生 功編)恒星社厚生閣,東京.1991; 125 150.
16) Kikuchi K, Furuta T, Ishizuka H, Yanagawa T. Growth of tiger puŠer, Takifugu rubripes, at diŠerent salinities. J. World. Aqua. Soc. 2007;38: 427434.
17) Yamashita Y, Tanaka M, Miller JM. Ecophysiology of juvenile ‰atˆsh in nursery grounds. J. Sea Res. 2001;45: 205218.