心 筋 細 胞 肥 大 お よ び 心 線 維 芽 細 胞 増 殖 で の Heat-shock protein 90 の 役 割
The role of Heat-shock protein 90 on cardiomyocyte hypertrophyand cardiac fibroblast proliferation
2019 年
i 目 次 緒 言 ...1 第 一 章 心 筋 細 胞 肥 大 へ の Hsp90 の 関 与 ...3 1-1 実 験 材 料 ・ 方 法 ...4 1-1-1 実 験 動 物 ...4 1-1-2 新 生 仔 ラ ッ ト 心 筋 組 織 か ら の 心 筋 細 胞 の 単 離 ...4 1-1-3 総 細 胞 数 に 占 め る 心 筋 細 胞 の 割 合 の 算 出 ...5 1-1-4 培 養 心 筋 細 胞 へ の 被 験 薬 の 投 与 ...5 1-1-5 心 筋 細 胞 面 積 の 測 定 ...6 1-1-6 培 養 心 筋 細 胞 の western immunoblotting 用 試 料 溶 液 の 調 製 ...6 1-1-7 心 筋 細 胞 培 養 液 の western immunoblotting 用 試 料 溶 液 の 調 製 ...6 1-1-8 Western immunoblotting ...7 1-1-9 統 計 方 法 ...8 1-2 結 果 ...9 1-2-1 心 筋 組 織 か ら 単 離 さ れ た 総 細 胞 数 に 占 め る 心 筋 細 胞 の 割 合 ...9 1-2-2 培 養 心 筋 細 胞 の 細 胞 面 積 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果 ... 10
1-2-3 心 筋 細 胞 培 養 液 中 の ANP 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 ... 11
1-2-4 培 養 心 筋 細 胞 の Hsp90 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果 ... 12
1-2-5 培 養 心 筋 細 胞 の c-Raf お よ び serine338 リ ン 酸 化 c-Raf 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果 ... 13
1-2-6 培 養 心 筋 細 胞 の Erk1/2 お よ び threonine202/tyrosine204 リ ン 酸 化 Erk1/2 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果 ... 15
ii
1-2-8 培 養 心 筋 細 胞 の cleaved caspase-3 含 量 に 及 ぼ す ET-1, 17-AAG
お よ び MG-132 の 効 果 ... 19 1-3 考 察 ... 20 第 二 章 心 線 維 芽 細 胞 増 殖 へ の Hsp90 の 関 与 ... 23 2-1 実 験 材 料 ・ 方 法 ... 24 2-1-1 実 験 動 物 ... 24 2-1-2 新 生 仔 ラ ッ ト 心 筋 組 織 か ら の 心 線 維 芽 細 胞 の 単 離 ... 24 2-1-3 心 線 維 芽 細 胞 へ の 被 験 薬 の 投 与 ... 24 2-1-4 心 線 維 芽 細 胞 増 殖 の 検 討 ... 24 2-1-5 心 線 維 芽 細 胞 の western immunoblotting 用 試 料 溶 液 の 調 製 ... 24 2-1-6 Western immunoblotting ... 25 2-1-7 統 計 方 法 ... 25 2-2 結 果 ... 26 2-2-1 培 養 心 線 維 芽 細 胞 の 細 胞 数 に 及 ぼ す 17-AAG の 効 果 ... 26 2-2-2 培 養 心 線 維 芽 細 胞 の Hsp90 含 量 に 及 ぼ す 17-AAG お よ び MG-132 の 効 果 ... 27
iii 略 語 表
本 文 中 で は 以 下 の 略 語 を 用 い る .
17-AAG 17-allylamino-17-demethoxygeldanamycin
ANOVA analysis of variance
ANP atrial natriuretic peptide
Ara-C cytosine--D-arabinoside
CF cardiac fibroblasts
CM conditioned medium
DAPI 4’,6-diamidino-2-phenylindole dihydrochloride
DMEM Dulbecco’s modified Eagle’s medium
DTT dithiothreitol
ECM extracellular matrix
EGTA ethylenglycol-bis(2-aminoethyl ether) -N,N,N’,N’-tetraacetic acid
ET-1 endothelin-1
Erk extracellular signal -regulated kinases
FBS fetal bovine serum
GA geldanamycin
GAPDH glyceraldehyde -3-phosphate dehydrogenase
HBSS Hanks’ balanced salt solution
HEPES 2-[4-(2-hydroxyethyl) -1-piperazinyl] ethanesulfonic acid
Hsp90 heat shock protein 90
ITS insulin-transferrin-selenium
MAP kinase mitogen-activated protein kinase
MKK MAP kinase kinase
MKKK MAP kinase kinase kinase
Mek MAPK/Erk kinase
NRVMs neonatal rat ventricular myocytes PAGE poly-acrylamide gel electrophoresis
PBS phosphate-buffered saline
PFA paraformaldehyde
PVDF polyvinilydine difluoride
S.E.M. standard error of the mean
SDS sodium dodecyl sulfate
TBST Tris-bufferd saline with Tween 20
TGF- transforming growth factor-β
iv
1 緒 言 心 臓 は ,虚 血 性 心 疾 患 や 高 血 圧 等 の 基 礎 疾 患 に 起 因 す る 負 荷 が 加 わ る と ,組 織 学 的 形 態 変 化 お よ び 心 ポ ン プ 機 能 の 代 償 機 構 を 働 か せ , 力 学 的 お よ び 代 謝 的 負 荷 に 適 応 す る 1 ).例 え ば ,心 筋 梗 塞 後 に 容 量 負 荷 あ る い は 圧 負 荷 が 持 続 す る と ,心 筋 梗 塞 を 免 れ た 生 存 心 筋 は 心 筋 細 胞 肥 大 お よ び 心 筋 組 織 の 線 維 化 を 介 し て , 心 機 能 を 維 持 す る .こ の 心 臓 の 組 織 学 的 な 再 構 築 は ,心 筋 リ モ デ リ ン グ と 呼 ば れ る 1 ).し か し な が ら ,長 期 に わ た り 心 臓 へ の 負 荷 が 持 続 す る と ,心 筋 リ モ デ リ ン グ が 過 度 に 進 行 し ,心 臓 の 代 償 機 構 は 破 綻 す る .代 償 機 構 の 破 綻 に 伴 い ,心 臓 は ,収 縮 力 低 下 に よ る 心 ポ ン プ 機 能 低 下 ,す な わ ち 心 不 全 に 陥 る 1 ).我 が 国 で は ,生 活 習 慣 の 欧 米 化 と 急 速 な 高 齢 化 で , 心 不 全 の 患 者 数 が 増 加 し て お り 2 ), 2016 年 の 厚 生 労 働 省 の 死 因 に 関 す る 統 計 で は , わ が 国 の 心 疾 患 に 起 因 す る 死 亡 者 数 197,807 人 の う ち , 約 37% の 73,508 人 が 心 不 全 で 死 亡 し て い る .し た が っ て ,心 不 全 に 対 す る 新 た な 治 療 薬 の 開 発 が 期 待 さ れ て お り , 心 機 能 の 代 償 期 か ら 心 不 全 に 陥 る 過 程 で の 心 臓 の 病 態 生 理 学 的 変 化 を 把 握 す る こ と は , 心 不 全 の 新 た な 治 療 法 を 開 発 す る た め に 重 要 な 意 義 を 持 つ . 心 筋 リ モ デ リ ン グ は ,細 胞 内 の 様 々 な 情 報 伝 達 経 路 に よ り 誘 発 さ れ る .機 械 的 刺 激 お よ び 神 経 体 液 性 刺 激 に よ り ,細 胞 膜 上 の 受 容 体 が 活 性 化 す る と ,細 胞 内 情 報 伝 達 経 路 お よ び そ れ に 引 き 続 く 転 写 因 子 が 活 性 化 さ れ る .そ の 結 果 ,タ ン パ ク 質 合 成 が 亢 進 し , 心 筋 リ モ デ リ ン グ が 進 展 す る . 細 胞 内 情 報 伝 達 経 路 の 1 種 で あ る mitogen-activated protein (MAP) キ ナ ー ゼ 経 路 は , 心 筋 リ モ デ リ ン グ の 進 展 に 重 要 な 役 割 を 果 た す と 考 え ら れ て い る .MAP キ ナ ー ゼ 経 路 は ,MAP kinase kinase kinase (MKKK), MAP kinase kinase (MKK) お よ び MAP kinase の 3 種 の プ ロ テ イ ン ナ ー ゼ に よ り 構 成 さ れ , こ れ ら 3 種 の キ ナ ー ゼ の 連 続 的 な 活 性 化 を 特 徴 と す る 3 ).
Raf/MAPK/Erk kinase (Me k)/extracellular signal -regulated kinases (Erk) 経 路 は , MAP キ ナ ー ゼ 経 路 の 1 つ で ,細 胞 増 殖 ,分 化 お よ び 細 胞 生 存 に 寄 与 す る 3 ).MKKK の
Raf は ,低 分 子 量 G タ ン パ ク 質 フ ァ ミ リ ー お よ び 他 の リ ン 酸 化 酵 素 と の 相 互 作 用 で 活 性 化 さ れ る .Raf は セ リ ン /ス レ オ ニ ン キ ナ ー ゼ で ,そ の 下 流 に 位 置 す る MKK の Mek1 お よ び Mek2 を リ ン 酸 化 す る . リ ン 酸 化 さ れ る こ と で 活 性 化 さ れ た Mek1 お よ び Mek2 は , Erk1/2 を リ ン 酸 化 反 応 を 介 し て 活 性 化 す る . こ の 活 性 化 さ れ た Erk1/2 は ,最 終 的 に 核 内 の 転 写 因 子 や 他 の プ ロ テ イ ン キ ナ ー ゼ の 活 性 化 を 制 御 す る 3 ). 培 養 心 筋 細 胞 で は , Erk1/2 が 受 容 体 刺 激 あ る い は 伸 展 刺 激 で 活 性 化
さ れ る 4 ).加 え て ,圧 負 荷 で 生 じ る 肥 大 心 で も Erk1/2 の 活 性 化 が 報 告 さ れ て い る
4 , 5 ). こ れ ら の 知 見 か ら , 心 臓 で の Raf/Mek/Erk 経 路 は , 心 筋 リ モ デ リ ン グ に 関 与
す る と 推 測 さ れ て い る .
4 1-1 実 験 材 料 ・ 方 法
1-1-1 実 験 動 物
本 研 究 で は , Wistar 系 新 生 仔 ラ ッ ト (哺 乳 2-3 日 齢 , Japan SLC Inc., Hamamatsu, Japan) を 用 い た .実 験 操 作 は ,動 物 の 保 護 お よ び 管 理 に 関 す る 法 律 (昭 和 48 年 法 律 第 105 号 ) お よ び 動 物 の 飼 育 お よ び 保 管 等 に 関 す る 基 準 (昭 和 55 年 総 理 府 告 示 第 6 号 ) に 従 っ た . 本 実 験 計 画 は 東 京 薬 科 大 学 動 物 実 験 委 員 会 に て 審 査 お よ び 認 可 後 , 実 施 さ れ た . 1-1-2 新 生 仔 ラ ッ ト 心 筋 組 織 か ら の 心 筋 細 胞 の 単 離 心 筋 細 胞 の 単 離・培 養 は Maki ら の 方 法 1 4 ) 従 っ て 行 っ た .新 生 仔 ラ ッ ト を 70% エ タ ノ ー ル で 消 毒 後 ,心 臓 を 摘 出 し た .摘 出 心 臓 か ら 心 房 を 除 去 し ,氷 冷 し た Ads buffer (116 mM NaCl , 20 mM 2-[4-(2-hydroxyethyl) -1-piperazinyl] ethanesulfonic acid [HEPES], 5.36 mM KCl, 0.83 mM MgSO4・ 7H2O, 12.5 mM NaH2PO4・ 2 H2O, 5.55 mM
glucose, pH 7.40) 中 に て 眼 科 用 ハ サ ミ で 心 室 筋 を 細 切 し た . 細 切 さ れ た 心 室 筋 を 0.08% (w/v) collagenase type II (Worthington Biochemical Co ., Lakewood, NJ, USA) を 含 む Ads buffer 中 で 37℃ ,6 分 間 イ ン キ ュ ベ ー ト し ,心 筋 細 胞 お よ び 非 心 筋 細 胞 を 分 散 さ せ た . こ の 行 程 は , 細 胞 が 分 散 し 組 織 が 原 形 を 留 め な く な る 6 回 目 ま で 行 っ た . そ の 後 , 細 胞 懸 濁 液 に 氷 冷 し た fetal bovine serum (FBS; Biowest, Nuqillé, France) を 加 え ,collagenase の 反 応 を 停 止 さ せ た 後 ,遠 心 分 離 (1,000 rpm, for 5 min, at 25℃) を 行 い , 細 胞 懸 濁 液 か ら collagenase を 除 去 し た . 次 に , PercollⓇ 密 度 勾 配 遠 心 法 に て 心 筋 細 胞 と 非 心 筋 細 胞 を 分 離 し た . Ads buffer で 希 釈 し た 58.5% PercollⓇ (比 重 1.082) (GE Healthcare UK Ltd. , Little Chalfont, UK) 中 に 細 胞 を 懸 濁 さ せ た 後 , 細 胞 懸 濁 液 を 40.5% PercollⓇ ( 比 重 1.060) の 下 に 積 層 し , 遠 心 分 離 (3,000 rpm, for 30 min, at 25℃ ) し た . こ の 遠 心 分 離 に よ り 非 心 筋 細 胞 は 上 層 に , 心 筋 細 胞 は 下 層 に 分 離 さ れ る (心 筋 細 胞 画 分 ). 遠 心 後 , 得 ら れ た 下 層 を 回 収 し た . 細 胞 懸 濁 液 に Ads buffer を 加 え た 後 , 遠 心 分 離 (1,000 rpm, for 5 min, at 25℃) を 行 い , PercollⓇ
を 洗 浄 し た . こ の 洗 浄 操 作 は 2 回 行 っ た .
次 に , 心 筋 細 胞 中 に 混 入 し た 非 心 筋 細 胞 を 除 去 す る た め , 心 筋 細 胞 を 100 mm culture dish (Corning Inc., Corning, NY, USA) に 播 種 し , 30 分 間 CO2 イ ン キ ュ ベ ー
タ (MCO-18AC-PJ, Panasonic, Osaka, Japan, 37℃ , 95% air + 5% CO2, 飽 和 水 蒸 気 ) 内
5
Glass Co. LTD., Shizuoka, Japan), 35 mm dish (Corning Inc.) お よ び 60 mm dish (Corning Inc.) に そ れ ぞ れ 4.0×104 cells/well, 4.5×105 cells/dish お よ び 1.0×106
cells/dish と な る よ う に 播 種 し た . な お 心 筋 細 胞 は CO2 イ ン キ ュ ベ ー タ 内 に て
10% FBS を 含 む Dulbecco’s modified Eagle’s medium (DMEM) (89% DMEM [ Sigma-Aldrich, St. Louis, MO, USA ], 10% FBS, 1% PenstrepⓇ [penicillin; 10,000 単 位 /mL, streptomycin; 10,000 g/mL, Thermo Fisher Scientific Inc. Waltham, MA USA]) 中 で 培 養 し た . さ ら に 非 心 筋 細 胞 を 排 除 す る た め , 心 筋 細 胞 単 離 後 24 時 間 目 に 1M cytosine--D-arabinoside (Ara-C; Sigma-Aldrich) を 培 地 に 添 加 し , 48 時 間 イ ン キ ュ
ベ ー ト し た . そ の 後 , 培 地 交 換 に よ り Ara-C を 除 去 し た .
1-1-3 総 細 胞 数 に 占 め る 心 筋 細 胞 の 割 合 の 算 出
単 離 操 作 後 の 培 養 さ れ た 総 細 胞 数 に 占 め る 心 筋 細 胞 の 割 合 は , 総 細 胞 に 対 す る -actinin 陽 性 細 胞 の 割 合 を 算 出 す る こ と に よ り 求 め た . 4’,6-diamidino-2-phenylindole dihydrochloride (DAPI; Thermo Fisher Scientific Inc.) に よ る 核 染 色 陽 性 細 胞 を 総 細 胞 と し ,筋 細 胞 マ ー カ ー で あ る -actinin の 免 疫 蛍 光 染 色 陽 性 細 胞 を 心 筋 細 胞 と し た .免 疫 蛍 光 染 色 に は 0.1% gelatin で コ ー テ ィ ン グ し た 8 well Chamber Slide 上 に 播 種 し た 細 胞 を 用 い た . Ara-C 除 去 6 時 間 後 の 心 筋 細 胞 を 0.1 M phosphate-buffered saline (PBS; 0.1M Na2HPO4, 0.1M KH2PO4, 0.1M NaCl) で 洗 浄 後 ,
2% paraformaldehyde (PFA) 中 で 室 温 ,60 分 間 固 定 し た .PFA を 除 去 し ,PBS で 洗 浄 後 ,0.5% TritonX-100Ⓡ 含 有 PBS で 室 温 ,30 分 間 浸 透 化 し た .PBS で 洗 浄 後 , 20% Blocking One HistoⓇ (Nacalai Tesque, Inc., Kyoto, Japan) 含 有 PBS で 室 温 , 30 分 間 ブ ロ ッ キ ン グ を 行 っ た . そ の 後 , 一 次 抗 体 と し て , mouse monoclonal anti- -actinin (EA-53, Sigma-Aldrich) を 5% Blocking One Histo 含 有 PBS で 100 倍 に 希 釈 し ,4℃ で 一 晩 反 応 さ せ た .反 応 終 了 後 ,PBS で 洗 浄 し ,二 次 抗 体 と し て ,Alexa FluorⓇ 555 donkey anti-mouse IgG (
Thermo Fisher Scientific Inc.) を PBS で 100 倍 に 希 釈 し 室 温 で 60 分 間 反 応 さ せ た . 核 染 色 に は 300 nM DAPI を 用 い , 室 温 で 10 分 間 反 応 さ せ た . 染 色 操 作 終 了 後 , PBS で 洗 浄 し , DAKO fluorescent mounting medium (DAKO Co., Carpinteria, CA) を 用 い て 封 入 し , 落 射 式 蛍 光 顕 微 鏡 (BX-52, Olympus,Tokyo,Japan) に て 観 察 を 行 っ た .細 胞 数 の 測 定 は well 内 の 5 視 野 を 無 作 為 に 選 択 し , そ の 細 胞 数 の 平 均 を 1 例 と し た .
1-1-4 培 養 心 筋 細 胞 へ の 被 験 薬 の 投 与
6
ア ソ ー ム 阻 害 薬 で あ る MG-132 (Peptide Institute Inc., Osaka, Japan) を そ れ ぞ れ 1 M お よ び 0.1 M と な る よ う に 投 与 し た .さ ら に 18 時 間 後 ,心 筋 細 胞 肥 大 を 惹 起 す る た め ,endothelin-1 (ET-1; Peptide Institute Inc.) を 1 nM と な る よ う に 添 加 し た .
1-1-5 心 筋 細 胞 面 積 の 測 定
心 筋 細 胞 面 積 は , 細 胞 膜 を 染 色 し た 心 筋 細 胞 の 細 胞 表 面 積 を 測 定 す る こ と に よ り 算 出 し た . 心 筋 細 胞 は , 0.1% gelatin で コ ー テ ィ ン グ し た 8 well Chamber Slide 上 に 播 種 し た 細 胞 を 用 い た . ET-1 添 加 24 時 間 後 の 心 筋 細 胞 を PBS で 洗 浄 後 , 2% PFA 中 で 室 温 , 60 分 間 固 定 し た . PFA を 除 去 し , PBS で 洗 浄 後 , fluorescein-conjugated wheat germ agglutinin (WGA -FITC, Vector Laboratories Inc., Burlingame, CA, USA) を 0.1% CaCl2 含 有 PBS で 200 倍 に 希 釈 し ,室 温 で 60 分 間 反 応 さ せ
た . な お , WGA-FITC は , 細 胞 膜 を 染 色 す る . 核 染 色 に は 300 nM DAPI を 用 い , 室 温 で 10 分 間 反 応 さ せ た . 染 色 操 作 終 了 後 , PBS で 洗 浄 し , DAKO fluorescent mounting medium を 用 い て 封 入 し ,落 射 式 蛍 光 顕 微 鏡 に て 観 察 を 行 っ た .撮 影 し た 画 像 の 解 析 に は , 画 像 解 析 用 ソ フ ト ウ ェ ア ImageJ (National Institutes of Health, Bethesda, MD, USA) を 使 用 し た . 心 筋 細 胞 面 積 は , 各 群 の 150-210 cells の 細 胞 面 積 を 測 定 し , そ の 平 均 を 1 例 と し た .
1-1-6 培 養 心 筋 細 胞 の western immunoblotting 用 試 料 溶 液 の 調 製
ET-1 添 加 24 時 間 後 に 氷 上 に て 培 地 を 除 去 し , 氷 冷 し た Homogenate buffer (20 mM HEPES, 0.25 M sucrose, 1 mM dithiothreitol [DTT] , 1 mM ethylenglycol -bis(2-aminoethyl ether) -N,N,N’,N’ -tetraacetic acid [EGTA] , protease inhibitor cocktail [cOmpleteⓇ; Roche, Mannheim, Germany], phosphatase inhibitor cocktail [PhosSTOP; Roche], pH 7.4, 4℃) を 加 え , cell scraper で 細 胞 を 回 収 し た . ポ ッ タ ー ホ モ ジ ナ イ ザ ー (Digital homogenizer, Iuchi, Osaka, Japan) で ホ モ ジ ナ イ ズ (700 rpm, for 2 min, 2 strokes) 後 , タ ン パ ク 質 量 を Bradford 法 で 測 定 し た . タ ン パ ク 質 の 最 終 濃 度 が 1.0 g/L と な る よ う に sample buffer (312.5 mM 2-amino-2-hydroxymethyl-1,3-propanediol [Tris]-HCl, 10% sodium dodecyl sulfate [SDS], 25% -mercaptoethanol, 50% glycerol, 0.005% bromophenolblue, pH 6.8) を 加 え , 沸 騰 水 浴 中 で 5 分 間 , 2 回 イ ン キ ュ ベ ー ト し western blotting 用 の サ ン プ ル と し た .
1-1-7 心 筋 細 胞 培 養 液 の western immunoblotting 用 試 料 溶 液 の 調 製
7 1-1-8 Western immunoblotting
心 筋 細 胞 c-Raf, Erk1/2, GATA4, Hsp90 お よ び caspase-3 タ ン パ ク 質 含 量 , お よ び 心 筋 細 胞 培 養 液 中 の atrial natriuretic peptide (ANP) 量 の 変 化 を 把 握 す る た め western immunoblot 法 を 用 い た .
SDS ポ リ ア ク リ ル ア ミ ド ゲ ル 電 気 泳 動 (SDS-PAGE) で 調 製 し た 各 画 分 中 の タ ン パ ク 質 を 分 離 し , 転 写 装 置 (KS-8460, Marysol, Tokyo, Japan) を 用 い て polyvinilydine difluoride (PVDF) 膜 (Immobilon-P, Millipore, Billerica, MA, USA) に 転 写 し た . 転 写 後 , PVDF 膜 を Tris-bufferd saline with Tween 20Ⓡ
(TBST) で 2 分 間 洗 浄 し た 後 , 50% BlockingOneⓇ
(Nacalai Tesque, Inc.) 含 有 TBST 溶 液 を 用 い て 室 温 で 30 分 間 ブ ロ ッ キ ン グ を 行 っ た . 5% Blocking OneⓇ 含 有 TBST 溶 液 で 5 分 間 , 3 回 洗 浄 し た . そ の 後 , 一 次 抗 体 を 5% Blocking OneⓇ 含 有 TBST 溶 液 で 希 釈 し て PVDF 膜 に 添 加 し , 低 温 室 4℃ で 一 晩 イ ン キ ュ ベ ー ト し た . 使 用 し た 一 次 抗 体 を Table 1 に 示 す .
Table 1 Details of the primary antibodies used for western immunoblotting.
一 次 抗 体 反 応 後 ,5% Blocking OneⓇ
含 有 TBST 溶 液 で 5 分 間 ,3 回 洗 浄 し た . 二 次 抗 体 に , c-Raf, Hsp90 お よ び GAPDH を 検 出 す る た め 3000 倍 と な る よ う に 5% Blocking OneⓇ 含 有 TBST 溶 液 で 希 釈 し た horseradish peroxidase conjugated goat anti-mouse IgG (Sigma -Aldrich) を , ANP, p-c-Raf (Ser338), Erk1/2, p-Erk1/2 (Thr202/Tyr204), GATA4, p-GATA4 (Ser105) お よ び cleaved caspase -3 (Asp175) に は
Primary antibody Clone
number Manufacture Dilution
Secondary antibody Rabbit polyclonal anti-ANP - Abcam, Cambridge, UK 1000 Rabbit
Mouse monoclonal anti-c-Raf 53/c-Raf-1 BD Biosciences,
Franklin Lakes, NJ, USA 2000 Mouse Rabbit monoclonal anti-p-c-Raf
(Ser338) 56A6
Cell Signaling Technology, Inc.,
Beverly, MA, USA 1000 Rabbit Rabbit polyclonal anti-Erk1/2 - Cell Signaling Technology, Inc. 2000 Rabbit Rabbit polyclonal anti-p-Erk1/2
(Thr202/Tyr204) - Cell Signaling Technology, Inc. 1000 Rabbit Rabbit monoclonal anti-GATA4 D3A3M Cell Signaling Technology, Inc. 1000 Rabbit Rabbit polyclonal anti-p-GATA4
(Ser105) - Abcam 1000 Rabbit
Mouse monoclonal anti-Hsp90 68/Hsp90 BD Biosciences 1000 Mouse Rabbit monoclonal anti-cleaved
8
3000 倍 と な る よ う に 5% Blocking OneⓇ 含 有 TBST 溶 液 で 希 釈 し た horseradish peroxidase-conjugated goat anti -rabbit IgG (Sigma-Aldrich) を 用 い , 室 温 で 1 時 間 イ ン キ ュ ベ ー ト し た . 二 次 抗 体 反 応 後 , TBST 溶 液 で 5 分 間 , 3 回 洗 浄 し , 検 出 に は Chemi-Lumi OneⓇ
(Nacalai Tesque, Inc.) を 用 い て 分 離 し た タ ン パ ク 質 を 発 光 さ せ , ル ミ ノ ・ イ メ ー ジ ア ナ ラ イ ザ ー (LAS3000, Fuji Photo Film Co. Ltd., Tokyo, Japan) に よ り 検 出 し た .現 像 さ れ た PVDF 膜 上 の タ ン パ ク 質 は ,ImageJ を 用 い て 半 定 量 化 し た .
1-1-9 統 計 方 法
9 1-2 結 果
1-2-1 心 筋 組 織 か ら 単 離 さ れ た 総 細 胞 数 に 占 め る 心 筋 細 胞 の 割 合
蛍 光 免 疫 染 色 に よ り 心 筋 細 胞 の 総 細 胞 数 に 占 め る 割 合 を 求 め た .DAPI を 用 い た 核 染 色 像 お よ び 筋 細 胞 マ ー カ ー の -actinin 染 色 像 を Figure 1 に 示 す . DAPI 陽 性 細 胞 に 対 す る -actinin 陽 性 細 胞 の 割 合 は 93.6 ± 2.0% (n=4) で あ っ た .
Figure 1 Ratio of cardiomyocytes to total cell s. Representative photomicrographs of cells stained
10
1-2-2 培 養 心 筋 細 胞 の 細 胞 面 積 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果 培 養 心 筋 細 胞 の 細 胞 面 積 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 2 に 示 す .
培 養 心 筋 細 胞 の 細 胞 面 積 は ,ET-1 投 与 に よ り Control 群 の そ れ の 約 120% と な り ,ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 80% と な っ た .ET-1, 17-AAG お よ び MG-132 投 与 で は , ET-1 お よ び 17-AAG 投 与 の そ れ と 同 様 の 値 と な っ た (Figure 2B).
Figure 2 Change in cell size of neonatal rat ventricular myocytes (NRVMs). (A) Representative
11
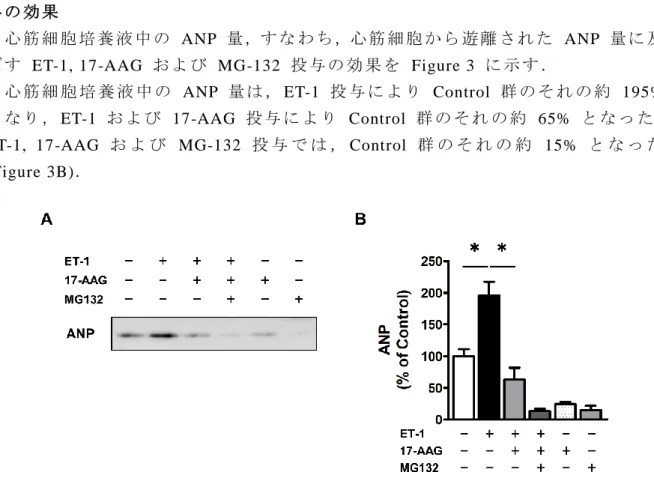
1-2-3 心 筋 細 胞 培 養 液 中 の ANP 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果
心 筋 細 胞 培 養 液 中 の ANP 量 ,す な わ ち ,心 筋 細 胞 か ら 遊 離 さ れ た ANP 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 3 に 示 す .
心 筋 細 胞 培 養 液 中 の ANP 量 は , ET-1 投 与 に よ り Control 群 の そ れ の 約 195% と な り , ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 65% と な っ た . ET-1, 17-AAG お よ び MG-132 投 与 で は , Control 群 の そ れ の 約 15% と な っ た (Figure 3B).
12 1-2-4 培 養 心 筋 細 胞 の Hsp90 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果 培 養 心 筋 細 胞 の Hsp90 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 4 に 示 す . 培 養 心 筋 細 胞 の Hsp90 含 量 は , ET-1 投 与 で は Control 群 の そ れ と 同 程 度 の 値 と な っ た . ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 180% に 増 加 し ,ET-1, 17-AAG お よ び MG-132 投 与 で は ,Control 群 の そ れ の 約 235% ま で 増 加 し た (Figure 4B).
Figure 4 Change in Hsp90 contents of NRVMs treated with or without ET-1, 17-AAG or MG-132. (A,B) Western blot analysis of Hsp90 proteins. The cells were pre -incubated with 17 -AAG (1 μM) and MG-132 (100 nM) followed by ET-1 (1 nM) stimulation for 6 h. Values are expressed as percentages compared with the non -stimulated group (incubated in the absence of ET-1, 17-AAG, and MG-132). Each value represents the mean ± S.E.M. of 7 independent experiments. * p < 0.05
vs. Control group .
13
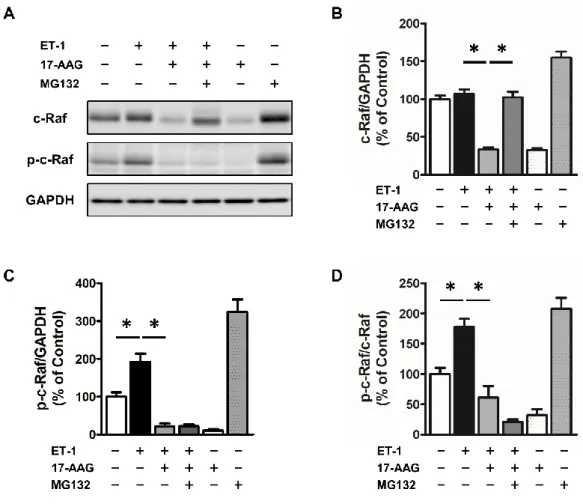
1-2-5 培 養 心 筋 細 胞 の c-Raf お よ び serine338 リ ン 酸 化 c-Raf 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果
培 養 心 筋 細 胞 の c-Raf 含 量 お よ び Ser338 リ ン 酸 化 c-Raf 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 5 に 示 す .
培 養 心 筋 細 胞 の c-Raf 含 量 は ,ET-1 投 与 で は Control 群 の そ れ と 同 程 度 で あ っ た も の の ,ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 35% ま で 減 少 し た . ET-1, 17-AAG お よ び MG-132 投 与 で は , Control 群 の そ れ と 同 程 度 ま で 回 復 し た (Figure 5B).
培 養 心 筋 細 胞 の Ser338 リ ン 酸 化 c-Raf 含 量 は ,ET-1 投 与 に よ り Control 群 の そ れ の 約 190% と な り , ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 20% と な っ た .ET-1, 17-AAG お よ び MG-132 投 与 で は ,ET-1 お よ び 17-AAG 投 与 の そ れ と 同 程 度 の 値 と な っ た (Figure 5C).
14
15
1-2-6 培 養 心 筋 細 胞 の Erk1/2 お よ び threonine202/tyrosine204 リ ン 酸 化 Erk1/2 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果
培 養 心 筋 細 胞 の Erk1/2 含 量 お よ び Thr202/Tyr204 リ ン 酸 化 Erk1/2 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 6 に 示 す .
培 養 心 筋 細 胞 の Erk1/2 含 量 は , 全 群 で 同 様 の 値 と な っ た (Figure 6B).
培 養 心 筋 細 胞 の Thr202/Tyr204 リ ン 酸 化 Erk1/2 含 量 は , ET-1 投 与 に よ り Control 群 の そ れ の 約 150% と な り ,ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 80% と な っ た .ET-1, 17-AAG お よ び MG-132 投 与 で は ,ET-1 お よ び 17-AAG 投 与 の そ れ と 同 程 度 の 値 と な っ た (Figure 6C).
16
17
1-2-7 培 養 心 筋 細 胞 の GATA4 お よ び serine105 リ ン 酸 化 GATA4 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果
培 養 心 筋 細 胞 の GATA4 含 量 お よ び Ser105 リ ン 酸 化 GATA4 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 7 に 示 す .
培 養 心 筋 細 胞 の GATA4 含 量 は ,ET-1 投 与 で は Control 群 の そ れ と 同 程 度 と な り ,ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 130% と な っ た .ET-1, 17-AAG お よ び MG-132 投 与 で は , ET-1 お よ び 17-AAG 投 与 の そ れ と 同 様 の 値 と な っ た (Figure 7B).
培 養 心 筋 細 胞 の Ser105 リ ン 酸 化 GATA4 含 量 は , ET-1 投 与 に よ り Control 群 の そ れ の 約 135% と な り , ET-1 お よ び 17-AAG 投 与 に よ り Control 群 の そ れ の 約 115% と な っ た .ET-1, 17-AAG お よ び MG-132 投 与 で は ,ET-1 お よ び 17-AAG 投 与 の そ れ と 同 様 の 値 と な っ た (Figure 7C).
18
19
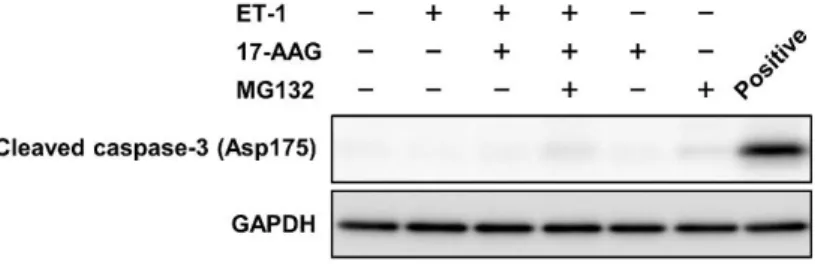
1-2-8 培 養 心 筋 細 胞 の cleaved caspase-3 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 の 効 果
培 養 心 筋 細 胞 の cleaved caspase-3 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 8 に 示 す .
培 養 心 筋 細 胞 の cleaved caspase-3 含 量 は , ET-1, 17-AAG お よ び MG-132 投 与 で 変 化 し な か っ た .
20 1-3 考 察
本 章 で は ,ま ず ,心 筋 細 胞 面 積 へ の Hsp90 阻 害 薬 17-AAG の 効 果 を 検 討 し た . 17-AAG は , ア ン サ マ イ シ ン 系 化 合 物 で あ る geldanamycin (GA) の 誘 導 体 で , GA の 肝 毒 性 を 軽 減 し な が ら , GA と ほ ぼ 同 等 の Hsp90 阻 害 活 性 を 発 揮 す る 誘 導 体 と さ れ る 1 5 ). 17-AAG は , Hsp90 の ATP 結 合 ポ ケ ッ ト で ATP と 競 合 す る こ と で ,
Hsp90 の シ ャ ペ ロ ン 機 能 を 阻 害 す る . 本 実 験 で は ,心 筋 細 胞 へ の 17-AAG 投 与 に よ り , ET-1 が 誘 発 し た 心 筋 細 胞 面 積 の 増 大 お よ び 培 養 液 中 の 肥 大 応 答 遺 伝 子 産 物 の ANP 量 の 増 加 が 抑 制 さ れ た .こ れ ら の 結 果 か ら ,Hsp90 が 心 筋 細 胞 肥 大 の 進 展 に 寄 与 す る 可 能 性 が 示 さ れ た . な お , 17-AAG を 投 与 し た 心 筋 細 胞 の 細 胞 面 積 は , 未 処 置 心 筋 細 胞 の そ れ よ り も 減 少 し た も の の , 17-AAG を 投 与 し た 心 筋 細 胞 で は caspase-3 が 活 性 化 さ れ て い な い こ と を 確 認 し た .Caspase-3 は ,切 断 さ れ て 活 性 化 さ れ る と ,細 胞 内 の タ ン パ ク 質 を 分 解 し ,ア ポ ト ー シ ス を 実 行 す る エ フ ェ ク タ ー カ ス パ ー ゼ で あ る 1 6 ).し た が っ て ,本 研 究 で は 17-AAG 投 与 後 の 細 胞 面 積 の 減 少 は , 心 筋 細 胞 で の ア ポ ト ー シ ス 誘 導 に よ る も の で は な い と 考 え ら れ た . 次 に ,Hsp90 が 心 筋 細 胞 肥 大 に 寄 与 す る 機 序 に つ い て 検 討 し た .本 研 究 で は ,細 胞 内 情 報 伝 達 経 路 と し て ,Raf/Mek/Erk 経 路 に 着 目 し た .Erk1/2 は ,c-Raf の 細 胞 内 情 報 伝 達 系 の 下 流 に 位 置 し て お り , c-Raf の 活 性 化 は Erk1/2 の 活 性 化 を 導 く . 17-AAG 投 与 に よ り ,心 筋 細 胞 の c-Raf 含 量 は 減 少 し ,ET-1 に よ る c-Raf お よ び Erk1/2 の リ ン 酸 化 の 促 進 が 抑 制 さ れ た .こ れ ら の 結 果 か ら ,Hsp90 は c-Raf の 安 定 化 に 寄 与 し て お り , c-Raf の 安 定 化 を 介 し て Raf/Mek/Erk 経 路 の 活 性 化 を 維 持 す る こ と が 示 さ れ た . 活 性 化 さ れ た Erk1/2 は , 細 胞 質 お よ び 核 内 の 基 質 タ ン パ ク 質 を リ ン 酸 化 す る . GATA4 は ,成 体 の 心 臓 で 恒 常 的 に 発 現 し て い る 転 写 因 子 で あ り ,Erk1/2 に よ り リ ン 酸 化 さ れ る こ と が 示 さ れ て い る 1 7 , 1 8 ). 心 肥 大 で 発 現 が 誘 導 さ れ る 肥 大 応 答 遺 伝 子 は , そ の プ ロ モ ー タ ー 領 域 に GATA4 結 合 部 位 を 保 持 し て お り , 心 臓 特 異 的 に GATA4 を 過 剰 発 現 さ せ る と 心 肥 大 を 呈 す る こ と か ら ,GATA4 の 活 性 化 は ,心 肥 大 の 進 展 に 寄 与 す る と 考 え ら れ て い る 1 9 ).GATA4 の 活 性 は ,そ の タ ン パ ク 質 発 現 お
よ び 翻 訳 後 修 飾 に よ り 制 御 さ れ る . GATA4 の Ser105 部 位 の リ ン 酸 化 は , GATA4 の DNA 結 合 能 お よ び 転 写 活 性 を 上 昇 さ せ る と 考 え ら れ て お り ,Erk1/2 は ,GATA4 の Ser105 部 位 を リ ン 酸 化 す る 1 8 ). 本 実 験 で は , ET-1 が 誘 発 し た GATA4 の
21 パ ク 質 含 量 の 減 少 が 回 復 す る が ,リ ソ ソ ー ム 阻 害 薬 ,カ ス パ ー ゼ 阻 害 薬 あ る い は カ ル パ イ ン 阻 害 薬 の 投 与 で , ク ラ イ ア ン ト タ ン パ ク 質 含 量 の 減 少 は 回 復 し な い こ と が 報 告 さ れ て い る 2 0 – 2 2 ).加 え て ,17-AAG 投 与 に よ り ,ユ ビ キ チ ン 化 さ れ た ク ラ イ ア ン ト タ ン パ ク 質 が 増 加 す る こ と も 示 さ れ て い る 2 1 , 2 2 ).そ こ で ,17-AAG を 投 与 さ れ た 心 筋 細 胞 の c-Raf の プ ロ テ ア ソ ー ム で の 分 解 に つ い て 検 討 す る た め , 心 筋 細 胞 に プ ロ テ ア ソ ー ム 阻 害 薬 で あ る MG-132 を 作 用 さ せ た . そ の 結 果 , 心 筋 細 胞 の c-Raf 含 量 は ,MG-132 存 在 下 で 17-AAG に よ る 減 少 が 抑 制 さ れ た .し た が っ て , 17-AAG 投 与 後 の 心 筋 細 胞 で の c-Raf 含 量 の 減 少 に は , プ ロ テ ア ソ ー ム で の 分 解 が 関 与 し て い る 可 能 性 が 示 さ れ た .さ ら に ,Hsp90 阻 害 薬 の 投 与 に よ り ,ク ラ イ ア ン ト タ ン パ ク 質 の タ ン パ ク 質 含 量 は 減 少 す る 一 方 で ,そ れ ら の mRNA 含 量 は 増 加 す る こ と が 報 告 さ れ て い る こ と か ら 9 , 2 3 ), 17-AAG 投 与 時 の c-Raf 含 量 の 低 下 は , c-Raf の 合 成 抑 制 で は な く , 分 解 促 進 に 起 因 す る と 推 察 さ れ た . 前 述 し た よ う に , c-Raf は ,Hsp90 の ク ラ イ ア ン ト タ ン パ ク 質 で あ り ,適 切 な フ ォ ー ル デ ィ ン グ に よ る 機 能 的 構 造 の 維 持 に Hsp90 と の 結 合 を 必 要 と す る 9 , 1 0 ).本 実 験 で は ,MG-132 投
与 は ,17-AAG が 減 少 さ せ た c-Raf 含 量 を 回 復 さ せ た が ,c-Raf の リ ン 酸 化 レ ベ ル は 変 化 さ せ な か っ た . 加 え て , MG-132 は 17-AAG が 抑 制 し た Erk1/2 お よ び GATA4 の リ ン 酸 化 と 心 筋 細 胞 面 積 の 増 大 に は 影 響 し な か っ た .こ れ ら の 結 果 か ら , 心 筋 細 胞 で は c-Raf が 機 能 す る た め に は , Hsp90 と の 相 互 作 用 が 必 要 で あ る と 考 え ら れ た .さ ら に ,17-AAG 投 与 時 に は c-Raf と Hsp90 の 相 互 作 用 が 阻 害 さ れ た 結 果 ,c-Raf は 機 能 的 構 造 を 形 成 で き ず ,そ の 細 胞 内 情 報 伝 達 系 の 下 流 シ グ ナ ル の 活 性 化 が 抑 制 さ れ る 可 能 性 が 推 察 さ れ た . 本 研 究 で は , 心 筋 細 胞 肥 大 に 寄 与 す る 細 胞 内 情 報 伝 達 経 路 と し て , Raf/Mek/Erk 経 路 に 着 目 し た が ,心 筋 細 胞 肥 大 に は ,心 筋 細 胞 内 の 種 々 の 情 報 伝 達 経 路 が 関 与 す る . Phosphoinositide 3-kinase (PI3K)/Akt 経 路 2 4 ) お よ び nuclear factor-B 経 路 2 5 )
の 活 性 化 は ,心 肥 大 の 進 展 に 寄 与 す る と さ れ る .こ れ ら の 情 報 伝 達 系 の 情 報 伝 達 因 子 で あ る Akt お よ び IB kinase (IKK) は ,Hsp90 の ク ラ イ ア ン ト タ ン パ ク 質 で あ る こ と が 知 ら れ て い る 8 , 2 6 ). し た が っ て , 本 実 験 で の 17-AAG に よ る 心 筋 細 胞 肥
23 第 二 章 心 線 維 芽 細 胞 増 殖 へ の Hsp90 の 関 与 第 一 章 で は , Hsp90 が c-Raf の 機 能 制 御 を 介 し て 心 筋 細 胞 肥 大 に 寄 与 す る 知 見 を 得 た .前 述 し た よ う に ,心 室 リ モ デ リ ン グ の 形 態 的 な 特 徴 は ,心 筋 細 胞 肥 大 お よ び 間 質 組 織 の 線 維 化 で あ る 1 ). 組 織 線 維 化 は , 細 胞 外 マ ト リ ッ ク ス (extracellular matrix; ECM) が 組 織 に 過 剰 に 沈 着 し た 状 態 を 主 徴 と し て お り , 傷 害 を 受 け た 組 織 で の 創 傷 治 癒 の 一 過 程 で あ る も の の , 過 度 の 線 維 化 の 進 行 は 当 該 組 織 の 機 能 障 害 を 惹 起 す る .心 線 維 化 は ,心 筋 細 胞 の 脱 落 を 伴 う 修 復 性 線 維 化 (reparative fibrosis) と , 心 筋 細 胞 の 脱 落 を 伴 わ な い 病 態 で 血 管 周 囲 か ら 緩 徐 に 進 行 す る 反 応 性 線 維 化 (reactive fibrosis) と に 大 別 さ れ ,心 線 維 芽 細 胞 は 両 者 に 関 与 す る .過 度 の 心 線 維 化 の 進 行 は , 左 室 拡 張 末 期 の stiffness を 上 昇 さ せ て 左 室 拡 張 機 能 障 害 を 誘 発 し , 線 維 化 に 起 因 す る 伝 導 障 害 お よ び 酸 素 お よ び 栄 養 物 の 拡 散 障 害 は , 左 室 収 縮 機 能 障 害 の 誘 因 と な る 2 7 ). 心 線 維 芽 細 胞 は ,ECM の 主 な 産 生 細 胞 で あ り ,心 臓 で の 主 な ECM は I お よ び III 型 コ ラ ー ゲ ン で あ る . 加 え て , オ ス テ オ ポ ン チ ン , ペ リ オ ス チ ン , テ ネ イ シ ン C な ど の ECM は , ス ト レ ス 後 に 発 現 が 顕 著 に 増 大 し , 心 筋 組 織 の 細 胞 の 活 性 化 , 増 殖・肥 大 お よ び 遊 走 能 の 惹 起 と そ れ ら の 促 進 に 関 与 す る と さ れ る 2 8 ).ECM の 代
謝 回 転 に は , ECM を 分 解 す る matrix metalloprotease (MMP) お よ び そ れ を 阻 害 す る tissue inhibitor of metalloprotease (TIMP) の 活 性 化 も 関 与 し て い る 2 8 ). 心 線 維 芽
細 胞 は こ れ ら の 酵 素 の 発 現 も 制 御 す る こ と か ら 2 7 ), 心 線 維 芽 細 胞 は 心 線 維 化 の 中
心 的 な 役 割 を 担 っ て お り ,心 線 維 芽 細 胞 の 増 殖 制 御 は ,心 線 維 化 の 軽 減 に 有 効 で あ る と 推 測 さ れ る .
24 2-1 実 験 材 料 ・ 方 法 2-1-1 実 験 動 物 1-1-1 と 同 様 に 実 験 動 物 を 使 用 し た . 2-1-2 新 生 仔 ラ ッ ト 心 筋 組 織 か ら の 心 線 維 芽 細 胞 の 単 離 心 線 維 芽 細 胞 の 単 離・培 養 は ,1-1-2 と 同 様 の 方 法 で 行 っ た .PercollⓇ 密 度 勾 配 遠 心 法 に て 分 離 し た 非 心 筋 細 胞 は , 遠 心 分 離 (1,000 rpm, for 5 min, at 25℃) を 行 い PercollⓇ を 洗 浄 し た 後 , 10% FBS (NICHIREI BIOSCIENCES INC., Tokyo, Japan) を 含 む DMEM で 再 懸 濁 し た . 非 心 筋 細 胞 中 に 混 入 し た 心 筋 細 胞 を 除 去 す る た め , 100 mm culture dish に 播 種 し , 30 分 間 CO2 イ ン キ ュ ベ ー タ 内 に 静 置 し た . こ の
dish の 底 面 に は ゼ ラ チ ン が コ ー テ ィ ン グ さ れ て い な い の で 心 筋 細 胞 は 接 着 で き な い が , 非 心 筋 細 胞 は 接 着 で き る . 30 分 後 , 培 養 皿 を Hanks’ Balanced Salt Solution (HBSS; Sigma-Aldrich) で 洗 浄 し , 新 た な 10% FBS を 含 む DMEM を 加 え た 後 , CO2 イ ン キ ュ ベ ー タ 内 で 培 養 し た .
2-1-3 心 線 維 芽 細 胞 へ の 被 験 薬 の 投 与
本 実 験 に は 2 継 代 目 の 培 養 心 線 維 芽 細 胞 を 用 い た . コ ン フ ル エ ン ト 前 の 心 線 維 芽 細 胞 を HBSS で 洗 浄 し , TrypLET M Express (Thermo Fisher Scientific Inc.) で 培 養
皿 か ら 剥 離 さ せ た 後 , 10% FBS を 含 む DMEM を 加 え て 反 応 を 停 止 さ せ た . 回 収 し た 細 胞 を 1000 rpm で 5 分 間 遠 心 分 離 し た 後 , 上 清 を 除 き , 下 層 の 細 胞 層 を 10% FBS を 含 む DMEM で 再 懸 濁 し た . 細 胞 計 数 盤 を 用 い て 細 胞 数 を 計 測 し , 60 mm culture dish に 3.5×106 cells/dish で 播 種 し , CO
2 イ ン キ ュ ベ ー タ 内 で 培 養 し
25 2-1-6 Western immunoblotting
1-1-8 と 同 様 の 方 法 で 行 っ た .
2-1-7 統 計 方 法
実 験 結 果 は , す べ て 平 均 値 ±標 準 誤 差 (standard error of the mean; S.E.M.) で 表 し た . 統 計 学 的 検 定 法 と し て , 群 間 比 較 の 解 析 お よ び 統 計 に は 1-way analysis of variance (ANOVA) あ る い は 2-way ANOVA 法 を 用 い た . ANOVA に よ る 有 意 差 が 認 め ら れ た 場 合 ,さ ら に 多 群 間 比 較 を 行 う た め post-hoc test と し て Tukey-Kramer 法 を 用 い た .測 定 値 の 統 計 的 比 較 に は ,95% の 信 頼 度 (p<0.05) を も っ て 有 意 と み な し た .統 計 処 理 に は ,StatViewⓇ
26 2-2 結 果 2-2-1 培 養 心 線 維 芽 細 胞 の 細 胞 数 に 及 ぼ す 17-AAG の 効 果 培 養 心 線 維 芽 細 胞 の 細 胞 数 に 及 ぼ す 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 10 に 示 す . 17-AAG 投 与 後 24 時 間 目 で の 培 養 心 線 維 芽 筋 細 胞 の 細 胞 数 は , Vehicle 群 で は 投 与 時 の そ れ の 約 155% に 増 加 し た も の の , 17-AAG 群 で は 投 与 時 の そ れ の 約 85% で あ っ た . 17-AAG 投 与 後 48 時 間 目 で の 培 養 心 線 維 芽 筋 細 胞 の 細 胞 数 は , Vehicle 群 で は 投 与 時 の そ れ の 約 195% に 増 加 し た も の の , 17-AAG 群 で は 投 与 時 の そ れ の 約 70% で あ っ た .
27 2-2-2 培 養 心 線 維 芽 細 胞 の Hsp90 含 量 に 及 ぼ す 17-AAG お よ び MG-132 の 効 果 培 養 心 線 維 芽 細 胞 の Hsp90 含 量 に 及 ぼ す 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 11 に 示 す . 培 養 心 線 維 芽 筋 細 胞 の Hsp90 含 量 は ,17-AAG 投 与 に よ り Vehicle 群 の そ れ の 約 185% に 増 加 し , 17-AAG お よ び MG-132 投 与 で は , Vehicle 群 の そ れ の 約 320% ま で 増 加 し た (Figure 11B).
Figure 11 Change in Hsp90 contents of CF treated with or without 17 -AAG or MG -132. (A,B) Western blot analysis of Hsp90 proteins. The cells were incubated with 17 -AAG (30 nM) and MG-132 (300 nM) for 48 h. Values are expressed as percentages compared with the Vehicle group (incubated in the absence of 17 -AAG and MG-132). Each value represents the mean ± S.E.M. of 4 independent experiments. *p < 0.05 vs. Vehicle group . (For interpretation of the references to colour in this figure legend)
28
2-2-3 培 養 心 線 維 芽 細 胞 の c-Raf お よ び serine338 リ ン 酸 化 c-Raf 含 量 に 及 ぼ す 17-AAG お よ び MG-132 の 効 果
培 養 心 線 維 芽 細 胞 の c-Raf 含 量 お よ び Ser338 リ ン 酸 化 c-Raf 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 12 に 示 す .
培 養 心 線 維 芽 細 胞 の c-Raf 含 量 は , 17-AAG 投 与 に よ り Vehicle 群 の そ れ の 約 35% ま で 減 少 し , 17-AAG お よ び MG-132 投 与 で は , Vehicle 群 の そ れ の 約 60% ま で 回 復 し た (Figure 12B).
培 養 心 線 維 芽 細 胞 の Ser338 リ ン 酸 化 c-Raf 含 量 は , 17-AAG 投 与 に よ り Vehicle 群 の そ れ の 約 55% に 減 少 し た .17-AAG お よ び MG-132 投 与 で は ,Vhicle 群 の そ れ の 約 40% ま で 減 少 し た (Figure 12C).
29
30
2-2-4 培 養 心 線 維 芽 細 胞 の Erk1/2 お よ び threonine202/tyrosine204 リ ン 酸 化 Erk1/2 含 量 に 及 ぼ す 17-AAG お よ び MG-132 の 効 果
培 養 心 線 維 芽 細 胞 の Erk1/2 含 量 お よ び Thr202/Tyr204 リ ン 酸 化 Erk1/2 含 量 に 及 ぼ す ET-1, 17-AAG お よ び MG-132 投 与 の 効 果 を Figure 13 に 示 す .
培 養 心 線 維 芽 細 胞 の Erk1/2 含 量 は ,17-AAG 投 与 に よ り Vehicle 群 の そ れ の 約 80% と な っ た .17-AAG お よ び MG-132 投 与 で は ,Vehicle 群 の そ れ の 約 65% と な っ た (Figure 13B).
培 養 心 線 維 芽 細 胞 の Thr202/Tyr204 リ ン 酸 化 Erk1/2 含 量 は ,17-AAG 投 与 に よ り Vehicle 群 の そ れ の 約 75% と な り , 17-AAG お よ び MG-132 の 投 与 に よ り Vehicle 群 の そ れ の 約 60% と な っ た (Figure 13C).
31
32 2-3 考 察 本 実 験 で は ,新 生 仔 ラ ッ ト か ら 単 離 し た 培 養 心 線 維 芽 細 胞 を 用 い た .心 線 維 芽 細 胞 の 単 離 操 作 で は ,心 筋 細 胞 お よ び 内 皮 細 胞 が 混 入 す る 可 能 性 が あ る .心 筋 細 胞 は 分 裂 ・ 増 殖 能 を 持 た な い こ と か ら , 本 実 験 で は 培 養 細 胞 を 2 継 代 す る こ と で , そ の 混 入 を 排 除 し た .血 管 内 皮 細 胞 は ,内 皮 細 胞 増 殖 因 子 の 非 存 在 下 で は 増 殖 し 難 い . し た が っ て , 本 実 験 で 用 い た 細 胞 は ほ と ん ど が 心 線 維 芽 細 胞 で あ る と 考 え ら れ る . 本 研 究 で は ,前 述 し た 方 法 で 単 離・培 養 し た 非 心 筋 細 胞 を 心 線 維 芽 細 胞 と し ,実 験 に 使 用 し た . ま ず , 心 線 維 芽 細 胞 の 細 胞 数 に 及 ぼ す Hsp90 阻 害 薬 17-AAG の 効 果 を 検 討 し た . 心 線 維 芽 細 胞 へ の 17-AAG 投 与 に よ り , 心 線 維 芽 細 胞 の 細 胞 増 殖 は 抑 制 さ れ た . こ の 結 果 か ら , Hsp90 の 心 線 維 芽 細 胞 の 細 胞 増 殖 へ の 寄 与 が 示 さ れ た . 次 に ,Hsp90 の 心 線 維 芽 細 胞 増 殖 へ 寄 与 す る 機 序 に つ い て 検 討 し た .第 一 章 と 同 様 に ,Raf/Mek/Erk 経 路 に 着 目 し た .そ の 結 果 ,17-AAG 投 与 に よ り ,心 線 維 芽 細 胞 の c-Raf 含 量 は 減 少 し , か つ c-Raf お よ び Erk1/2 の リ ン 酸 化 が 抑 制 さ れ た . つ ま り ,Hsp90 は c-Raf の 高 次 構 造 の 維 持 に 寄 与 し て お り ,こ の c-Raf の 安 定 化 を 介 し て Raf/Mek/Erk 経 路 の 活 性 化 を 維 持 す る こ と が 示 さ れ た . 第 一 章 で は , 17-AAG 投 与 後 の 心 筋 細 胞 の c-Raf 含 量 の 減 少 に は , プ ロ テ ア ソ ー ム で の 分 解 が 関 与 す る こ と が 示 さ れ た . そ こ で , 心 線 維 芽 細 胞 で の 17-AAG 投 与 後 の c-Raf の プ ロ テ ア ソ ー ム に よ る 分 解 に つ い て 検 討 す る た め , 心 線 維 芽 細 胞 へ プ ロ テ ア ソ ー ム 阻 害 薬 MG-132 を 作 用 さ せ た . そ の 結 果 , 17-AAG 投 与 後 の 心 線 維 芽 細 胞 の c-Raf 含 量 減 少 は ,MG-132 に よ り 軽 減 さ れ た .し た が っ て ,17-AAG 投 与 後 の c-Raf 含 量 の 減 少 に は , プ ロ テ ア ソ ー ム で の 分 解 の 関 与 が 示 さ れ た . そ の 一 方 で , MG-132 は 17-AAG に よ り 減 少 し た c-Raf 含 量 を 回 復 さ せ た も の の , c-Raf お よ び Erk1/2 の リ ン 酸 化 の 抑 制 は 軽 減 さ せ な か っ た . こ れ ら の 結 果 か ら , 心 筋 細 胞 と 同 様 に ,心 線 維 芽 細 胞 で も c-Raf は ,機 能 す る た め に Hsp90 と の 相 互 作 用 が 必 要 で あ る と 推 察 さ れ た .加 え て ,17-AAG 投 与 に よ り c-Raf と Hsp90 の 相 互 作 用 が 阻 害 さ れ , c-Raf が 機 能 的 構 造 を 形 成 で き な か っ た 結 果 ,c-Raf の 下 流 シ グ ナ ル の 活 性 化 が 抑 制 さ れ た と 考 え ら れ た . 前 述 し た よ う に ,リ ン 酸 化 さ れ る こ と で 活 性 化 さ れ た Erk1/2 は ,核 内 の 転 写 因 子 の 活 性 化 を 制 御 す る . Fos, Jun お よ び ATF family の ヘ テ ロ 二 量 体 で 構 成 さ れ る activating protein -1 (AP-1) は ,細 胞 増 殖 を 促 進 さ せ る 転 写 因 子 で あ る 3 0 ).活 性 化 さ
れ た Erk1/2 は , 核 内 に 移 行 し Elk な ど の 転 写 因 子 の 活 性 化 を 介 し て Fos, Jun, Myc お よ び Egr な ど の タ ン パ ク 質 発 現 を 増 加 さ せ る こ と が 示 さ れ て い る 3 0 ).本 実
33
心 線 維 芽 細 胞 は ,種 々 の サ イ ト カ イ ン お よ び 増 殖 因 子 を 産 生 し ,オ ー ト ク リ ン ま た は パ ラ ク リ ン を 介 し て 分 泌 細 胞 自 身 お よ び そ の 周 囲 の 細 胞 の 増 殖 を 促 進 す る 3 1 ).
Transforming growth factor- (TGF-) は , 線 維 芽 細 胞 の 増 殖 を 促 進 す る と さ れ て お
り 3 2 ), 心 線 維 芽 細 胞 は TGF- を 分 泌 す る こ と が 報 告 さ れ て い る 3 3 ). TGF- 受 容 体 を 介 し て 活 性 化 さ れ る 細 胞 内 情 報 伝 達 経 路 は 多 岐 に わ た り , そ れ ら の 中 に は Raf/Mek/Erk 経 路 も 含 ま れ る 3 4 ). す な わ ち , TGF- 受 容 体 の 活 性 化 に 続 い て Raf/Mek/Erk 経 路 が 活 性 化 さ れ る と 考 え ら れ る .圧 負 荷 心 肥 大 モ デ ル 動 物 へ の 17-AAG 投 与 は , TGF- 受 容 体 の 下 流 に 位 置 す る 情 報 伝 達 経 路 の 活 性 化 を 減 弱 さ せ , 心 線 維 化 を 抑 制 す る こ と が 示 さ れ て い る 3 5 ). こ れ ら の 知 見 か ら , 本 実 験 で の 17-AAG に よ る Raf/Mek/Erk 経 路 の 活 性 化 抑 制 に は , TGF- 受 容 体 を 介 す る シ グ ナ ル の 減 弱 も 寄 与 す る と 考 え ら れ た .
Figure 14 Effects of 17 -Allylamino -17 -Demethoxy- geldanamycin (17 -AAG) treatment on
Raf/Mek/Erk signaling pathway in CF.
36 引 用 文 献
1) Cohn JN, Ferrari R, Sharpe N. Cardiac remodeling -concepts and clinical implications: A consensus paper from an international forum on cardiac remodeling. J. Am. Coll. Cardiol., 35, 569–582 (2000).
2) Okura Y, Ramadan MM, Ohno Y, Mitsuma W, Tanaka K, Ito M, Suzuki K, Tanabe N, Kodama M, Aizawa Y. Impending epidemic. Circ. J., 72, 489–491 (2008).
3) Widmann C, Gibson S, Jarpe MB, Johnson GL. Mitogen -activated protein kinase: conservation of a three -kinase module from yeast to human. Physiol. Rev., 79, 143–180 (1999).
4) Bueno OF, Molkentin JD. Involvement of extracellular signal -regulated kinases 1/2 in cardiac hypertrophy and cell death. Circ. Res., 91, 776–781 (2002). 5) Rapacciuolo A, Esposito G, Caron K, Mao L, Thomas SA, Rockman HA.
Important role of endo genous norepinephrine and epinephrine in the development of in vivo pressure -overload cardiac hypertrophy. J. Am. Coll. Cardiol., 38, 876– 882 (2001).
6) Lai BT, Chin NW, Stanek AE, Keh W, Lanks KW. Quantitation and intracellular localization of the 85K hea t shock protein by using monoclonal and polyclonal antibodies. Mol. Cell. Biol., 4, 2802–2810 (1984).
7) Picard D. Heat-shock protein 90, a chaperone for folding and regulation. Cell. Mol. Life Sci., 59, 1640–1648 (2002).
8) Citri A, Harari D, Shohat G, Ra makrishnan P, Gan J, Lavi S, Eisenstein M, Kimchi A, Wallach D, Pietrokovski S, Yarden Y. Hsp90 recognizes a common surface on client kinases. J. Biol. Chem., 281, 14361–14369 (2006).
9) Schulte TW, Blagosklonny M V., Ingui C, Neckers L. Disruption of the Hsp90 molecular complex results in destabilization of Raf -1 and loss of Raf-1-Ras association. J. Biol. Chem., 270, 24585–24588 (1995).
10) Soga S, Kozawa T, Narumi H, Akinaga S, Irie K, Matsumoto K, Sharma S V, Nakano H, Mizukami T, Hara M. Radicicol leads to selective depletion of Raf kinase and disrupts K -Ras-activated aberrant signaling pathway. J. Biol. Chem., 273, 822–828 (1998).
11) Soonpaa MH, Field LJ. Survey of studies examining mammalian cardiomyocyte DNA synthesis. Circulation Research, 83, 15-26 (1998).
37
Neonatal rat cardiomyocytes - a model for the study of morphological,
biochemical and electrophysiological characteristics of the heart. Biomed. Pap., 145, 49–55 (2001).
14) Maki T, Kawahara Y, Tanonaka K, Yagi A, Takeo S. Effects of tanshinone VI on the hypertrophy of cardiac myocytes and fibrosis of cardiac fibroblasts of
neonatal rats. Planta Med., 68, 1103–1107 (2002).
15) Schulte TW, Neckers LM. The benzoquinone ansamycin 17
-allylamino-17-demethoxygeldanamycin binds to HSP90 and shares important biologic activities with geldanamycin. Cancer Chemother. Pharmacol. , 42, 273–279 (1998).
16) Porter AG, Jänicke RU. Emerging roles of caspase -3 in apoptosis. Cell Death Differ., 6, 99–104 (1999).
17) Arceci RJ, King AA, Simon MC, Orkin SH, Wilson DB. Mouse GATA -4: a retinoic acid-inducible GATA-binding transcription factor expressed in
endodermally derived tissues and heart. Mol. Cell. Biol., 13, 2235–2246 (1993). 18) Liang Q, Wiese RJ, Bueno OF, Dai YS, Markham BE, Molkentin JD. The
transcription factor GATA4 is activated by extracellular signal -regulated kinase 1- and 2-mediated phosphorylation of serine 105 in cardiomyocytes. Mol. Cell. Biol., 21, 7460–7469 (2001).
19) Liang Q, De Windt LJ, Witt SA, Kimball TR, Markham BE, Molkentin JD. The transcription factors GATA4 and GATA6 regulate cardiomyocyte hypertrophy in vitro and in vivo. J. Biol. Chem., 276, 30245–30253 (2001).
20) Dou F, Yuan L Di, Zhu JJ. Heat shock pro tein 90 indirectly regulates ERK activity by affecting Raf protein metabolism. Acta Biochim. Biophys. Sin. (Shanghai)., 37, 501–505 (2005).
21) Basso AD, Solit DB, Chiosis G, Giri B, Tsichlis P, Rosen N. Akt forms an intracellular complex with heat shock p rotein 90 (Hsp90) and Cdc37 and is destabilized by inhibitors of Hsp90 function. J. Biol. Chem., 277, 39858–39866 (2002).
22) Grbovic OM, Basso AD, Sawai A, Ye Q, Friedlander P, Solit D, Rosen N. V600E B-Raf requires the Hsp90 chaperone for stability and i s degraded in response to Hsp90 inhibitors. Proc. Natl. Acad. Sci. U. S. A. , 103, 57–62 (2006).
23) Cao X, Bloomston M, Zhang T, Frankel WL, Jia G, Wang B, Hall NC, Koch RM, Cheng H, Knopp M V, Sun D. Synergistic antipancreatic tumor effect by
simultaneously targeting hypoxic cancer cells with HSP90 inhibitor and glycolysis inhibitor. Clin. Cancer Res., 14, 1831–1839 (2008).
38
hypertrophy. J. Biol. Chem., 275, 4693–4698 (2000).
25) Purcell NH, Tang G, Yu C, Mercurio F, DiDonato JA, Lin A. Activation of NF -κB is required for hypertrophic growth of primary rat neonatal ventricular
cardiomyocytes. Proc. Natl. Acad. Sci., 98, 6668–6673 (2001).
26) Lee KH, Jang Y, Chung JH. Heat shock protein 90 regulates IκB kinase complex and NF-κB activation in angiotensin II -induced cardiac cell hypertrophy. Exp. Mol. Med., 42, 703 (2010).
27) Fan D, Takawale A, Lee J, Kassir i Z. Cardiac fibroblasts, fibrosis and
extracellular matrix remodeling in heart disease. Fibrogenesis Tissue Repair , 5, 15 (2012).
28) Manabe I, Shindo T, Nagai R. Gene expression in fibroblasts and fibrosis involvement in cardiac hypertrophy, Circulation Research, 91, 12, 1103-1113 (2002).
29) Zak R. Cell proliferation during cardiac growth. Am. J. Cardiol., 31, 211–219 (1973).
30) Shaulian E, Karin M. AP-1 in cell proliferation and survival. Oncogene, 20, 2390–2400 (2001).
31) Bouzegrhane F, Thibault G. Is angiotensin II a proliferative factor of cardiac fibroblasts? Cardiovasc. Res., 53, 304–312 (2002).
32) Salazar KD, Lankford SM, Brody AR. Mesenchymal stem cells produce Wnt isoforms and TGF-β 1 that mediate proliferation and procollagen expression by lung fibroblasts. Am. J. Physiol. Cell. Mol. Physiol. , 297, L1002–L1011 (2009). 33) Campbell SE, Katwa LC. Angiotensin II Stimulated Expression of Transforming
Growth Factor-β1in Cardiac Fibroblasts and Myofibroblasts. J. Mol. Cell. Cardiol., 29, 1947–1958 (1997).
34) Leask A, Abraham DJ. TGF-β signaling and the fibrotic response. FASEB J., 18, 816–827 (2004).