The Japanese Society for Plant Systematics
NII-Electronic Library Service
The Japanese Sooiety for Plant Systematios
ISSN OOOI;−6799 植物分類, 地 理
50
(1):101−138
(1999)ミ ズ
ニラ属
の自 然 誌 と 分 類
高宮正 之
〒 860−8555熊 本 県 熊 本 市 黒 髪2−39−1熊 本 大 学理学 部 環 境理学 科
MASAYUKI TAKAMIYA :Natural history and taxonomy of lsoetes(Isoetaceae).− Acta Phytotax. Gθo加ム50ω ’101−13&
Depa 厂tnzent of Environmental Sci召nce , Faculり’qf Science, Kuma 脚 ‘o Universめ’, Kurok翩 i 2・
39−1,K“snamoto 860−8555, Japan
Abstract. On the bas重s of a review of earlier studies inclu〔Hng tbose of the author, our current knowledge on morphology , palynology, reproduction mode , cytology and metabolism of the genus’soe 榔 (Isoetaceae;Ptehdophyta )were summarized . Morphological and anatomical
features of corms , roots , leaves, sporangia , mega −and microspores and spore germination, as
well as their taxonom畫cal values , were discussed. All chromosomal data ava孟lable throughout the
world show the presence of a pGlyploid series from 2兀(2n=22)to 12x(2n=132)and eight species
to have intrasp if菫c polyploidies. Out of 81 taxa whose chK 〕1nosome numbers were reported ,27
are diploids,30 tetraploids, and l3 natural hybrids. Isoetes山us shows a reticulate evolution through hybridization and po】yploidization, A variety of reproduction mode , i,e., sexual ,
apomictic and vegetative reproduction was explained . A diagllostic key to Japanese taxa
comprising four species , one hybrid (∬soetes ×michinokuan の, and two varieties was also provided, and in addition phylogenetic relationships among them was discussed.
Key words :anatomy , chromosome number , hybrid, lsoetes, morphology , polyl〕10idy, reproduction , spore, taxonomy .
Reeeived Februaりy 12 ,1999’accepted ルfaY 10, ig99
ミ ズニ ラ属 (∬Soetes
L
.)は 変 わっ た形 態 を もつ シダ植物である。 植物体は比較的 小さ く, 極度に短縮さ れ た茎,束生する葉と根を も ち, ある種の単 子葉植物に よ く似てい る (図1)。この類 似には Linnaeus(1751)す らだ ま さ れて,ミズニ ラ属を種子植物 に含み,雌雄の花
ば か りか果 実 と種 子 まで記 載 し て しまっ た、日本名の ミズニ ラは水辺 のニ ラ を 意味 し,
中 国で も水韮をあて る。 欧 米で は Merllyn’s Grass や quillwort が 用 い ら れる。 属 名の Isoetesは ISO(ギ リ シャ語の iSOS由 来で同じの意 味 )+ etOS (年 )で常 緑 植 物 を意 味 してい る
が, 多くの場 合 常 緑ではない 。 すべ て多年草である。
現在で は ほ と ん ど有用 性は無い が,ヨ ーロ
ッ パで は飢 饉 時に食 用 とさ れて い た記 録 も ある (May ,1978)。 植 物 体 には多量の で んぷ ん と油 脂 が 含 ま れてい る。野鳥,あるい は ア
ヒ ル, 豚, 牛な どの家 畜類も生草として食べ る可能性 も考 えら れて い た (Pfeiffer, 1922)。
脾 臓や肝 臓の治療薬 として用 い ら れ た記録も あ る (May ,1978)。 観 賞 用 水 草 として稀に水 槽で栽培さ れてい る。
現 在,ミ ズニ ラ属に は
150
種 以 上 が ある と考え られ てい る が ,種数は 1(Linnaeus , 1754),49(Baker,1887),62 (Sadebeck ,1902),64(Pfeiffer,1922),100(Reed,1953),130(Jerrny,N工 工一Eleotronio Library
102 Acta Phytotax. Geobot. Vol.50 1990),150(Taylor and Hickey,1992)と年々 増加し て きた。 中に は400 種以 上 と考 える立場
もある (Srivastava et at.,
1993
)。 ミズニ ラ属はア メ リ カ, ヨーロッ パ ,アフ リカ, イン ド,
アジ ア , オース トラ リ アなど 汎 世界 的に分布してい るが, 集 団がそれぞれ孤立 し てい る
た め固有種が多い 。 アフリ カ,オース トラリア
, イ ン ド, 南ア メ リカの もの は 類 似 性が 高い こ と か ら, ミズニ ラ属 をゴ ン ドワナ大 陸 起 源とする 立場もある (Marsden ,1976a;
Taylor and Hickey, 1992)。 多くの もの は湿地, 湖沼,あるい は流水に生 育 し, 陸生あるい
は水生の抽水また は沈水植物であ る (図
1
)。 しか し,年 間の ど こ か に乾期がある よう な 環 境に適 応 した 種 も あ り, 乾季に葉は枯れ て塊茎と して休眠 する。 系統学的には, 白亜紀に水生植 物か ら陸生植物に適 応 放 散 し, その後 様々 な 地域で 二次的に水生 植 物に逆 戻 り し た とも考えら れてい る (Hickey,1986a;Taylor and Hickey,1992 )。
Stytites Amstutzは, AmstUtz(1957)とRauh and Falk(1959a, b)と に よっ てペ ール の アン
デ ス の 同一場所か ら別々 に発 見 され た ミズニ ラ科の 新属で ,S. andic ・
1
αAmstutz
と ∫.g翩 配蜘rαRauh &Falkの 2 種か ら なっ てい た。し か し, 他の研 究者たちは,これ ら を他の 南 米種の極端な変異 型 と考え,ミズニ ラ属 に 含め て お り(
Kubitzki
andBorchert
,1964; G6mez ,1g80;Hi
。key
,1984
),本 稿で もそ れ に従い ミズニ ラ属を ミズニ ラ科の 唯一の 現 生 属と し て扱 う。 汎 世界的 な分 布 を する ミズニ ラ属 だが, これ まで に世 界 的 なモ ノ グラ フ ィ
ッ クな整理 (Baker,1887;Sadebeck,1902;Pfeiffer,1922; Reed,1953;Fuchs,1962)や 以下の よ うな各地 域ごとの分 類 学 的 報 告がある:北米 (Reed,1965;
BoGm
,1982;Taylor et al.,1993a;Musselman and Knepper,1994),南 米 (Fuchs,1982),アフ リカ (
Wanntorp
,1970;Ha11,1971),イン ド (Panigrahi,198 1; Srivastava et at.,1993),オース トラリア (Marsden,1976b;Johnson,1984;
Chinnook ,1998),ニ ュ ーギニ ア (Croft.1980),オース トラ リア ・イン ド・東南ア ジ ア ・中 国 ・日本 (
Marsden
, 1979),日本(Takamiya et al.,1997)。本稿で は, これ まで邦 文で ほ とん ど紹 介 さ れた こ との な か っ た ミズニ ラ属の形 態 学,
細胞学, 生活史等につ い て レビュ ーし,日本の ミ ズニ ラ属の分 類につ い て述べ る。
形 態
塊茎
ミ ズ ニ ラ属の軸は短く直立 する構造で塊茎 (corm )と呼ば れ, 肥 大成長を行 う(図
2A
,D
,E)。 毎 年 新 しい 組織が内側か ら 追 加 され るこ と に より, 外側の 古い 部 分は剥 離 してい く
(図 3B)。 通 常 直 径は 0 .5 〜数cm だが, 日本の ミ ズ ニ ラ で は 8cm に も な ることが ある
(
Makino
,1go4)。上部は多数の細 長い 葉を叢生し,その中 央は くぼ ん でい て,ぎっ しり重 な り合っ た葉に覆 われ た くぼ みの底に成 長 点がある (図2A
,3A
)。 下部は多数の根を叢生 し,1条 ま たは中 心 より放射状に でた3条の溝によっ て浅 く2分 ま た は3分 (まれに 4分 ) する (図2A ,3A , B)。 塊 茎は通 常 分枝し ない が , 稀 に分 枝 するこ ともある。上 述 した Isoetes andicola (Amstutz)L .D . Gomez ← Stylites andicota )で は, 常に数 回の分枝が見ら れ る(図2B)。 唯一の
例だ が L tegetiformans Rury の ように短 く匍匐して根 茎 状に な る場合 (図 2C)も ある (Rury,1978)。 塊 茎はこれ まで に,直 立 した根 茎,茎, あるい は上 部の葉 が生 じ る とこ ろ は茎で 下部は担 根 体な ど,様々 に記 載 されて きた (Scott and Hill, 1900;Stokey,
1909;
Lang
,1915;West and Takeda,1915;武 田,1916;Osborn ,1922;Stewart,1947:Bhambie,1962;Palillo,1963 ;Ogura ,1972)。 塊 茎の解 剖 学的特徴や担根 体として の解釈は Palillo
The Japanese Society for Plant Systematics
NII-Electronic Library Service
The Japanese Sooiety for Plant Systematios
August l999 高宮 ミ ズニ ラ属の 自然 誌と分 類 103
図 1.ミズニ ラ属の生 育 状 況。 A.ミズ ニ ラモ ドキ (lroetes pseudojaponica),葉は抽水し てい る (1993 年 9月,岡 山 県 )[コB,C.ミズニ ラ(L
japonica
)。干魃の た め 干 上 がっ た 池の底に 生育してい るのが発 見さ れた。陸生の状 態 だが,例年は一
年中 沈水してい て ,ミズニ ラ が 生育してい る こ と は知 ら れて い な かっ た。 C の群 落は数 万 個 体か ら構 成さ れて い た(1994年 8月, 香JII県)。
N工 工一Eleotronio Library
104 Acta Phytotax. Geobot. Vol.50
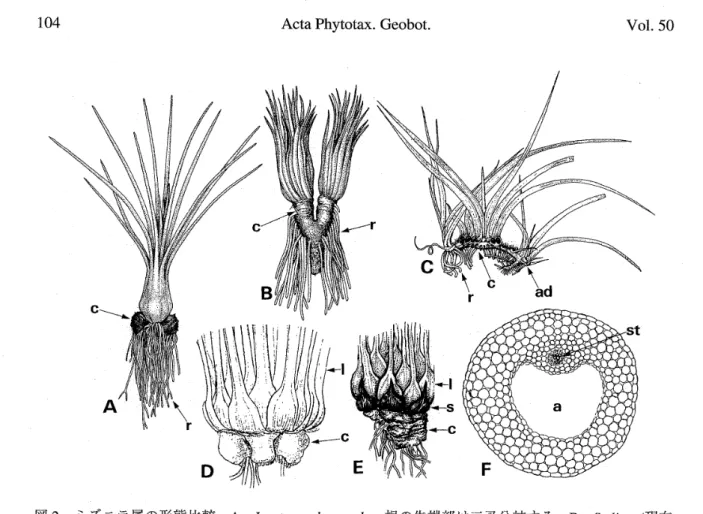
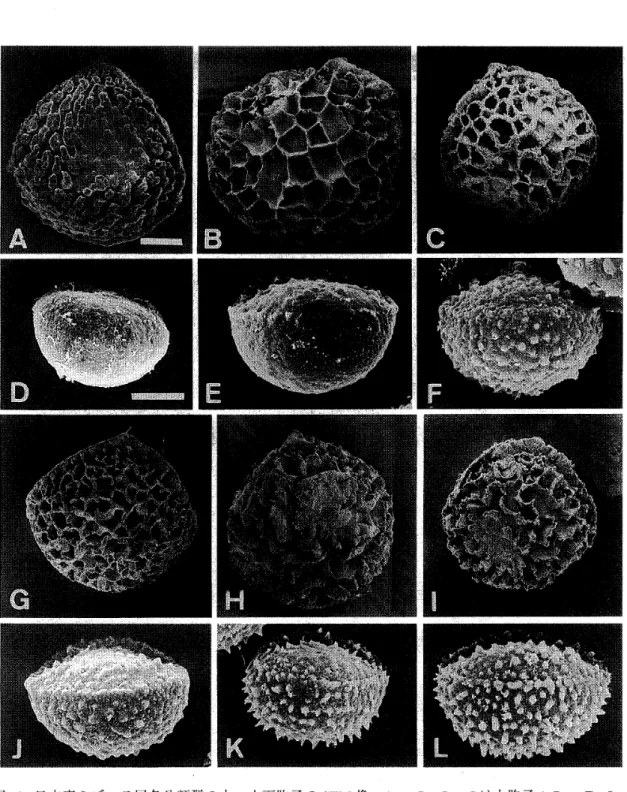
図2.ミ ズニラ属の形態比較。 A. Isoetes melanopoda 。 根の先 端 部は二叉 分 岐 する。 B. Stylites(現在 は lsoetes)の 1種, 塊 茎 が 伸 長 して二 分 し, 分岐しない 根が両 側か ら 生 じ て い る。 C.∬. tegettfqnnans ,塊 茎 は根 茎 状に なっ て不定芽を持ち,葉は2列に生 じる。 根は分 岐しない。 D.ミ ズニ ラ (L
japonica
)の塊 茎 付 近。 E.∬. howelliの塊 茎 付 近,鱗 片 葉 を 持つ 。 F,ミ ズ ニ ラ (∬.japenica
)の根の横 断 面,上 側が塊 茎の 巾 央部。 aは空 隙, ad は不定芽, c は塊 茎, 1は葉, r は根,sは鱗片 葉,st は中心柱を示 す。 A, B は Stewart and Rothwell(1993) を, C はRury (1978)を, D はWest and Takeda(1915)を, E はTaylor et al.(1993乱)を, Fは Ogura(1972)を改 変し た。
(t963, 1982), Karrfalt and Eggert (1977a, b,1978), Karrfart(1984a, b), Stewart and Rothwell
(1993)に詳しくs 今市(1996)は イワヒ バ属を含め た担根体につ い て総 説 して い る。
ミズニ ラ属の塊 茎の比 較 研 究か ら, 古 生 物 学 的に は中 生 代 石 炭 紀の木 本 性 リンボク類 から
1
槓次 縮 小 して草本性の ミズニ ラ属に なっ た として, 日本 語の解 説に もしば し ば引 用 さ れ てい る (岩槻,1977,1988;西 田,1977;戸 部,1994)。 しか しな が ら最 初 か ら草 本として存 在 して い た との見 解 も紹 介 さ れて い る (加 藤,1997;西田,正998)。 近年巾 生 代三畳紀か らミズニ ラ属その もの の化石が発見さ れ (Wang ,1991;Retallack, 1997),縮 小 説は塗 り替えら れ
ようと してい る。 現生 の ミズニ ラ属に関 係 する化 石 種の比 較は Pig (1992)や Skog and Hi1玉
(1992)が総 説してい る。
塊 茎の裂 片 数 はミ ズニ ラ属 分 類の指 標 形質とし て重要視さ れて きた (Braun,1863;Baker,
1887;Pfeiffer,1922)。塊 茎の裂 片数は種に よっ て ほ ぼ一
定で はあるが, 同 種 内の 変 異
(Clute,1905;Marsden,1976b;Takainiya et al.,1997)や (図3B),個 体 発 生 過 程の変 化 (Karrfalt
and Egger,1977a)も報 告 されてい る。
The Japanese Society for Plant Systematics
NII-Electronic Library Service
The Japanese Sooiety for Plant Systematios
August
l999
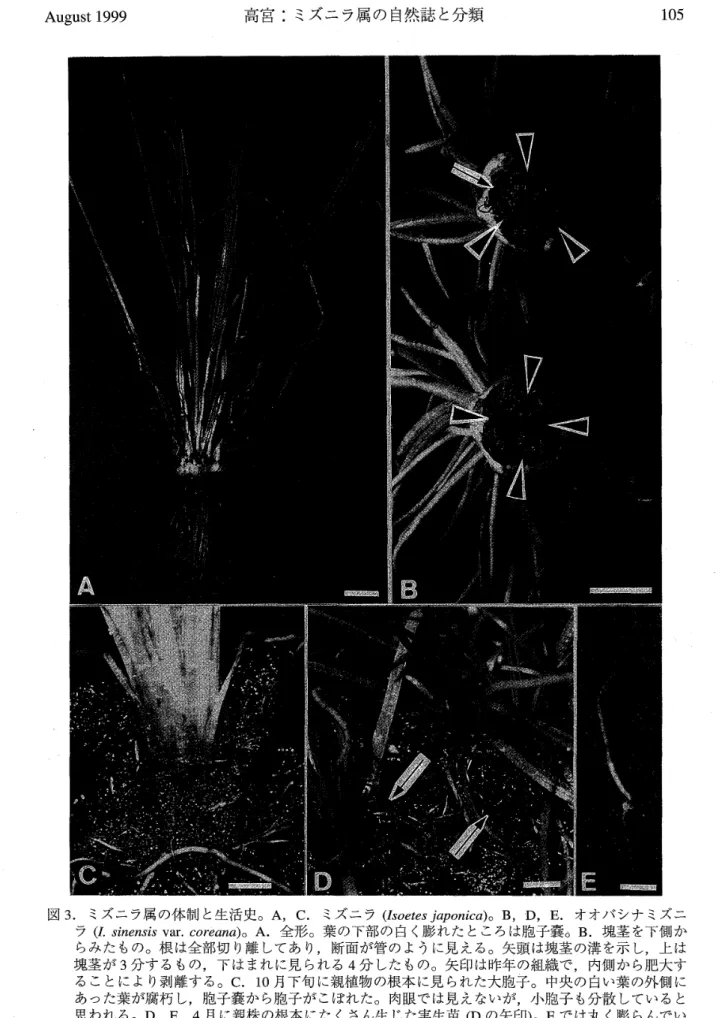
高 宮 :ミ ズニ ラ属の自然誌と分類 105図 3.ミズニ ラ属の体 制と生 活 史。 A, C,ミズニ ラ (Lyoe tes J’
apon ica) o B, D, E.オ オバ シ ナ ミ ズニ ラ (Lsinensis yal・. coreana )。 A .全形。 葉の下部の 自 く膨れ た とこ ろ は胞子嚢。 B.塊 茎を 1・.
側か
ら み たもの。根は全 部 切り離し てあり,断面 が管の ように見え る。 矢頭は 塊茎の溝を示 し,上 は 塊 茎 が 3分 す る もの,下 は ま れに見ら れ る4分 し た もの 。 矢 印は昨 年の組 織で , 内側か ら肥 大 す る こ とに より剥 離 する。 C.1 月 下 旬に親 植 物の根 本に見 られた大 胞.」 中 央の 白い 葉の外 側に あっ た葉 が 腐 朽 し,胞子嚢 から胞子がこ ぼ れた。肉眼で は見 えない が,小 胞子も分散し てい る と 思われ る。D , E.4月に親株の根本に たくさ ん 生 じ た実生苗 (D の矢 印)。 E で は丸 く膨ら ん でい
るのが大 胞 子壁の殻で, h側が葉, 下側は根。 ス ケール はA − D =lcm
, E =2 。
N工 工一Eleotronio Library
106 Acta Phytotax, Geobot , Vol.50
B C
図4・ミズニ ラ属の葉の模 式 図 と解 剖 図. A .ヒメ ミ ズニ ラ (・s・e・、s
.
a、i。tica). B − E.ミ ズニ ラ (・. .iaponica)。 A − C は葉の付け根 を 向
軸側 か ら見た図。 A, B は小胞予嚢, C は大 胞 子 嚢を描い て
い る。 1は小 舌,m は膜 質に なっ た葉 縁, s は胞 予 嚢, se は隔 壁, v は蓋 膜,矢頭はD の横 断 面
の位 置 をし めす。 スケール は 5 。D, E.葉の横断面 と その拡 大で, 上側が向輌 。 1は鱒
で,大 き な4個の空 隙 が 維 管束を囲む。 p は周辺 繊 維 束, stは 中 心 柱 を示 す。ス ケール は lmm 。
E は中心 樹 寸近の拡 大,iは軋 ・柱 空 隙, xy は木 部, phは師部。 ス ケー]vはO.2mm 。
根
根は塊 茎内部で発生 し, 塊 茎 下 部か ら多数現れ,二叉 分 岐を繰 り返 す (図
2A
,3A , B)。 た だし,Isoetes andicot α や L tegetifonnans では分 岐 し ない (図 2B , C)。 根は葉より も長 く 宿 存 し, 皮層は木化して外観上褐 色 となる。 横断面で は1
個の大 きな空 隙が あ り(図2F),断 面は管の ように見 える (図3B)。維 管束は中央にな く,空隙に向かっ て突 出 した 皮層の
…部にある (図 2F)。 木部と師部は単 尤 型で , 内 側に師部がある。 根の構造は ミズニ ラ属 全体で均一であ り,リン ボ ク類の担 根 体
Stigmaria
か ら生じ る棋に極似 す る (Stewart andRothwell,1993)。
葉
葉は螺旋 状 に配 列 して い て, 葉 柄は 無 く先端は尖る (図 1, 2A , B , 3A )。 Iseete,s’ tegenf・rmans の葉は 2列に生 じる (図2C)。 断面は,向軸 側が広い台 形 となる (図4D)。多
い もの で は 1 株 か ら200 本近 くの葉が 生 じ る。 葉 脚は匙状に広 くな り,その葉 縁 は一層
の細胞層か ら な る薄い 翼 状 となる (図4A −
C
)。 葉の 下部は,し ば し ば地 面の中にあるの で クロ ロ フ ィ ル を欠 き 白 色である。 葉 は通常長さ 2 〜 30cnユ,幅は 1 〜5mm だ が,ミ ズニ ラ では長さ 1m 幅 lcm に な る ことも ある。 陸生の ミ ズ ニ ラ属には保護器官と して鱗 片 葉
(scale )とphyllopodiaが知ら れ る (図 ゴE)。 鱗 片 葉は葉 全 体の細胞が厚膜化して休 眠 期に茎
The Japanese Society for Plant Systematics
NII-Electronic Library Service
The Japanese Sooiety for Plant Systematios
August 1999 高宮 ミ ズニ ラ属の 自然誌と分 類 107
頂 を保 護 する もの,phyllopodiaは葉の一
部が厚膜化しその 一一
部が残っ た もの で,地 中海, 南アフ リカ, オース トラ リア
, イン ド, 南 米, 北 米 中 央 部の ミズ ニ ラ属か ら知ら れて い る (Braun,1864,1868;Pfeiffer,1922;Duthie,1929a;Wanntorp,1970; Hickey,1 986a; Srivastava
and Wagai ,1996)。 Hickey (1986a)は こ れ らの保護葉の有 無を系統上重 要 視 してい る。 日本
の ミズニ ラ属に は どちらも ない 。
解剖学的に は葉に は縦につ な がっ た 4つ の気道 (lacuna, air chamber )がある (図4D)。 気 道は ミ ズニ ラ属 植 物の全て が もつ 共 有 形 質である こ と か ら, ミズニ ラ属は基 本 的に水 生 植物であっ た と考えられ てい る (Hickey,1986a;Taylor anf Hickey ,1992)。 気道は浮力を 生 じ る と ともに,水生植物の通気に関 する機能を持つ (Keeley,1982)。 気 孔 を持つ 種で は,
気孔と気道がつ ながっ てい る。 とこ ろ どこ ろ に気道を横 切る星 状の 柔細 胞か らなる隔膜
(transverse septa, diaphragm) がある (図4B)。維管束は葉の 中央部に 1 本通っ てい て,ど ん
な長い 葉で も分岐する こ と は ない (図4D)。 葉の解剖 学 的特徴は 生育環 境の影響 を受 けに くい た め, 安 定 した 指 標 形 質 と考え ら れてい る (Braun,1863; Pfeiffer,1922;Duthie,1929a;
Hall,1971;Marsden ,1979)。
気 孔の有無は分 類上重要と さ れ たり (Pfeiffer, 1922;Marsden , 1979),生育条件に影響さ れ 不安定とさ れた り (Kott and Britton,1985)する。 日本 産の ミズニ ラ属で は沈 水 性の ヒ メ ミ ズニ ラ に は気孔が無 く, 他の抽水 性の分 類群には ある と記載さ れ て きた (Makino , 1904;
Braun, 1862a;West and Takeda , 1915;武田,1916;Pfeiffer,1922;Palmer,1927;L6ve ,1962;
DeVol ,1972;Marsden ,1979;中 池,1992)。し か し,栽培品と各研究機関の標 本の観察か ら,
ヒ メ’
ミ ズニ ラ に も葉の先 端 部に気 孔 が 僅 か な が ら あるこ と が 判 明した (Takamiya et al., 1997)。 他の 日本 産 ミズニ ラ属で は,気孔は葉の上部 1〆3 に多 数存在する。 後述 する よう
に水 性の ミ ズニ ラ属には CAM 代謝
経
路がある こ と が知ら れてい る の で , 本 属の気 孔の機能につ い て は機能 面か ら検 討 する必 要 が あるだろ う。
ミズニ ラ属の葉に は, 表 皮 細 胞の 下部に厚膜 細胞か ら な る器械組織である周辺繊維束
(peripheral fibre strands )が見ら れ るこ と がある (図4D)。 周 辺 繊 維 束の有無, 位 置, 数は指 標 形 質とされ た り (Braun,1863;Pfeiffer,
1922
;Duthie
,1929a;parker, lg43;Wanntorp,1970;Marsden ,1979),否定的だ っ た りする (West and Takeda,
1915
;武 田,1916;Hall,1971
;Kott−amdBritton,1985)。 日本 産の ミズニ ラ属では, ヒ メ ミズニ ラ に は無 く,他の 分類群で は葉の向 軸 側に 3箇 所,背 軸 側に 1 箇 所規 則 的に観 察 さ れる (Takamiya et at., 1997)。 ミズニ ラ で は,
稀に 6 箇 所 (左 右の側方に 1 箇所 ずつ )見ら れ るこ とも知ら れ る が (West・and・Takeda,1915;
武田,1916),我々 の観察で は 6 箇所の例はなかっ た。 周 辺 繊 維 束は葉の 強 化の 機能か ら陸
生へ の適 応 と考 えら れて い るが (Hickey,1986a),水生種に現れ る 理由は不 明で ある。
葉は大 き な
4
個の気道以外に,維管束 内に空 隙を もち (図4E), 中 心 柱 内 空 隙 (intrastelar
canal )と呼ばれてい る (Han, 1971)。 日本 産の ミズニ ラ属で は,シナ ミ ズニ ラ と オ オバ シナ
ミズニ ラ には空 隙 が
3
箇 所 あるが 他の分 類 群で は 1個で,良い 指 標 形 質 となる (Takamiya
etal .,1997)Q 胞 子 葉 と胞 子 嚢
幼 植 物を除い て,基 本 的に ミズ ニ ラ属の 全て の葉は胞 子嚢をつ け 胞 子 葉である。 た だ しシ ーズ ン の 最初に 生 じ る葉 (一香 外 側 に着 く葉 )は稔 性 を 持たない こ と
が多い 。 ミ ズニ
ラ属植物は大胞子と小 胞 子の 2 種の 胞 子 を持つ 異 形 胞子シ ダ植物で, 大胞子は大 胞 子嚢
N工 工一Eleotronio Library