ウイルソン病の動物モデルである
LEC ラット肝組織の微小領域元素分析
はじめに
肝臓は、銅や鉄の遷移性微量元素の恒常性にとって最 も重要な臓器である。過剰に摂取された銅は肝臓から胆 汁中に排泄される。その排泄能の低下した銅過剰症は肝 臓を障害する [1]。生体にとって最も重要な金属元素であ る鉄には生理的な排泄系がない。消化管での鉄吸収は、 肝臓で産生されるペプシジンにより調節される。この調 節ペプチドの産生が極度に低下した遺伝性鉄過剰症では、 鉄の貯蔵臓器である肝臓が主に障害される [2]。このよう に遷移元素の過剰な蓄積は酸化ストレスを介して肝毒性 を発揮する [3]。その解毒機序として、それぞれヘモジデ リンや銅チオネインとして遷移性を軽減し、さらに肝細 胞ライソゾームに貯え、他の細胞内小器官への障害性を 制御している [4]。このライソゾームは、酸性条件化で活 性化される加水分解酵素に富む小器官として、ミトコン ドリアから分離され、透過型電子顕微鏡(TEM)での観察 も可能になった [5]。 ウイルソン病は、家族性のレンズ核変性症、肝・レンズ 核変性症、ATP7B 欠損症と、時代とともにその病因の理解 も深まった [1]。この病気では、肝細胞の膜系に銅を取り込 むトランスポーターである ATP7B に遺伝的欠陥があり [6]、 銅の胆汁排泄が完全に遮断される。過剰な銅は、まず肝臓 に溜まり、次いで中枢神経系を含む肝臓以外の臓器に沈着 する。細胞内では動員されたライソゾームに銅の蓄積して いる像が顕微鏡で確認できる。本症は、ヘモクロマトーシ スと同じように、遺伝的障害がライソゾーム以外の輸送タ ンパクにあるため、二次ライソゾーム病に分類される [7]。 最近の研究では、動員されたライソゾームには銅のほかに 鉄の蓄積がしばしば観察されると報告されている [8]。それ は、本症に特徴的な低セルロプラスミン血症と関連してい る。血中のセルロプラスミンは、主なフェロキシダーゼで あるため、その活性低下時には鉄の輸送が制限される。特に、 長期間の銅キレート剤治療後には、ライソゾームに蓄積し ていた銅は鉄によって置き換わる [9]。 Long−Evans Cinnamon(LEC)ラットは、ATP7B 変異 と低セルロプラスミン血症を持つウイルソン病の動物モデ ルである [10]。銅と鉄は LEC ラットの劇症肝炎から慢性肝 炎に至る種々の肝障害に関与している [11、12]。これらの 遷移元素は最終的に解毒化されて肝細胞ライソゾームに貯 えられるため、ウイルソン病の動物モデルである LEC ラッ ト肝臓組織の超微形態的研究は、ウイルソン病の患者で、 慢性肝炎から肝硬変、さらに肝細胞癌の発生する過程で、 肝毒性のある遷移元素に対して細胞がどのように自らを防 御しているのかの情報を与えるであろう。 光学顕微鏡は現代病理学の基本的な機器であるが、鉄過 剰症であるヘモクロマトーシスや銅過剰症のウイルソン病 においては毒性金属の検出感度が悪いので注意が必要であ る。それゆえ、銅過剰症に対する低セルロプラスミン血症や、 鉄過剰症に対する高フェリチン血症のような金属過剰の臨 床情報のある場合に限って、毒性金属を検出するため特殊 〒 464-8650 名古屋市千種区楠元町 1-100 E-mail: [email protected]古川 実穂
†、 鈴木 瑛子
†、 林 久男
†、 藤田 芳和
††、 服部 亜衣
†、
巽 康彰
†、加藤 宏一
† †愛知学院大学薬学部薬物治療学講座
††名古屋大学医学部附属医学教育研究支援センター
電子顕微鏡とエネルギー分散型X線分析装置を搭載した JEM− 2010 と JEM− 2800 による微小領域元 素分析は Long− Evans Cinnamon ラット肝組織の銅と鉄の過剰状態を可視化した。二つの遷移性微量 元素は肝細胞とクッパー細胞のライソゾームにタンパク体として蓄積していることが確認された。従って、 この動物は、ウイルソン病における遷移性微量元素による進行性の肝臓を含めた多臓器障害の機序を研究 するのに良いモデルである。な染色方法が指示される。これら特殊な染色法には、銅に 対してロダニンあるいはルベアン酸、鉄に対してベルリン ブルー染色がある。それでも、これらの検出感度は低いので、 特殊染色が陰性結果となっても、過剰による障害、すなわ ち遷移元素の毒性は否定できない [13]。その理由は光学顕 微鏡レベルでは、特殊染色であっても金属元素を貯蔵して いるライソゾームが数多く集合した時に限り、染色される からである。 一方、TEM も病理学的診断や研究で広く使われている。 TEM により判明する細胞内小器官の構造上の変化は、人 や動物の病気のメカニズムを理解するために重要な情報 である。加えて、TEM とエネルギー分散型 X 線元素分析 (EDX)を組み合わせた微小領域元素分析(MA)は、微量 元素の細胞小器官内での沈着だけではなく、金属タンパ ク複合体の組成もおおよそ判明する [4、14]。この論文で は、LEC ラットの肝組織を対象に、遷移元素である銅と 鉄への毒性の制御システムについて超微形態学的検討を した。
材料と方法
本研究に使用した 2 匹の 16 週令の雄性 LEC ラットは 中部科学資材株式会社(名古屋)から購入した。肝切除術 は麻酔下に行った。肝組織を 0.1M カコジル酸で緩衝した 2%グルタールアルデヒドで 3 日間固定した。後固定は 0.1% オスミウム酸で 45 分実施した。また組織切片上でライ ソゾームを確認するため Novikoff の方法に従い酸性ホス ファターゼ反応を加えた [5]。肝組織の一部を 0.1M カコジ ル酸で pH7.2 に緩衝した 4%パラホルムアルデヒドで、4℃、 60 分間、固定した。検体を 4℃で、pH7.2 に緩衝した 0.1M カコジル酸、0.25M スクロースで洗浄した後、寒天培地に 包埋し薄く切った。この薄切片を、硝酸鉛、スクロース、 β−グリセロリン酸を加えた 0.05M 酢酸緩衝液(pH5.0-5.2) 中で、37℃、10 分間反応させた。最後に蒸留水で短時間 の洗浄をした。いずれの肝組織も、最終的に TAAB812 エ ポキシ樹脂に包埋し、超薄切片を作成し金グリッドにのせ た。無染色の切片と 2%ウラン液で染色した切片を、EDX を装備した日本電子社製 TEM である JEM− 2010 と JEM − 2800 で調べた。 は じ め に、JEM− 2010 を 使 用 し て 予 備 検 索 を し た。 TEM による観察では肝細胞とクッパー細胞の電子密度の 高い顆粒の細胞内分布を確認した。次いで EDX を使って、 肝細胞とクッパー細胞の高電子密度の顆粒に短時間、無作 為に電子線を当て、銅と鉄の沈着の有無をスクリーニング した。鉛は電子密度の高い顆粒でいつも同定された。銅 や鉄が個々の顆粒で同定された時には、その顆粒から発 生した全ての X 線を 200 秒間測定した。銅と鉄の同定は、 EDX に装備された自動分析法に従った。銅と鉄の共存性 は、銅単独、銅・鉄共存、鉄単独として表現した。JEM− 2010 によるスクリーニングで確認した銅と鉄の共存性に ついて、JEM− 2800 を使用して元素の二次元の分布を調べ た。その EDX により、金属を含む顆粒の分散する視野(細 胞質)を 480 秒間、走査して元素の分布情報を集め、二次 元画像を得た。JEM−2800 には二次元の走査終了後にも局 所から発生したすべての元素の X 線スペクトラムが再現 できる記録能がある。結果
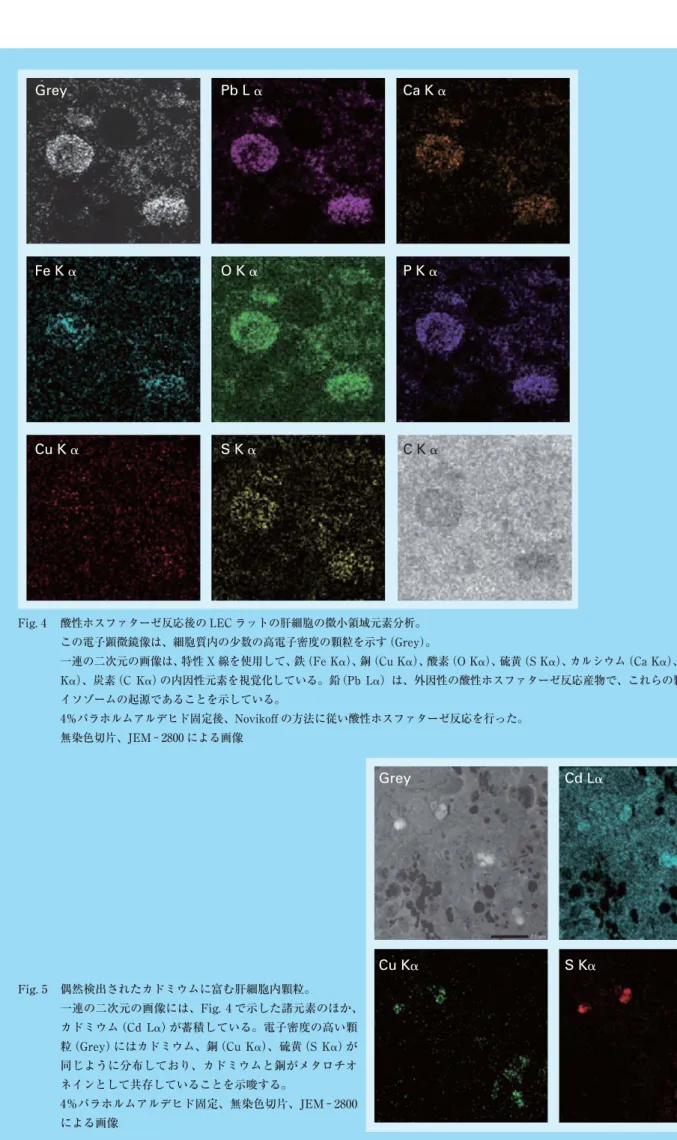
TEM を 使 用 し て 0.1% オ ス ミ ウ ム 酸 で 固 定 し た LEC ラットの肝組織超薄切片を観察すると、肝細胞とクッパー 細胞に存在する核、ミトコンドリア、高電子密度の顆粒 などの細胞内小器官の確認は容易であった。酸性ホスファ ターゼを加えたパラフォルムアルデヒドの単固定・無染 色切片では、ライソゾームは鉛沈着があるため、容易に 同定できた。EDX と組み合わせると、細胞内の銅と鉄をFig. 1 LEC ラットの肝細胞の微小領域元素分析。 この電子顕微鏡像は、肝細胞内のさまざまは高電子密度の顆粒を示す(上)。 2 つのサブタイプがあり、銅に富む顆粒と鉄を持つ顆粒は EDX により同定された(下)。 鉄顆粒は様々な形をしており、基質内に微粒子を含んでいる。一方、銅顆粒の基質は均一で、び漫性である。 0.1%オスミウム酸固定、ウラン染色 持ったライソゾーム顆粒の特徴が視覚化された。二重固 定し、電子染色した標本では、肝細胞の銅に富む顆粒の 基質はび慢性で、鉄を持つ顆粒の基質は電子密度の高い 粒子に富んでいた(Fig. 1)。クッパー細胞は、様々な形態 を示す高電子密度の顆粒を含有していた。銅と鉄の分布 は、これらの顆粒内で明らかに違っていた(Fig. 2)。酸 性ホスファターゼ反応を加えた標本では、肝細胞とクッ パー細胞の一部のライソゾーム顆粒内に内因性元素の銅・ 鉄と外因性の鉛の共存が確認された(Fig. 3)。顆粒以外 の細胞質の鉛沈着は軽微であった。二次元の鉄と他の元 素の分布はライソゾーム内ではっきり視覚化された(Fig. 4)。鉄は、リンと酸素と共存し、一方で銅は硫黄と同じ 部位に現れた。ライソゾーム内の銅と鉄の局在性は少し 違っていた。一部の高電子密度の顆粒には、鉄、銅に加 えカドミウムが検出された(Fig. 5)。
考察
ウイルソン病は肝細胞の銅輸送タンパクである ATP7B の異常による原発性銅過剰症であり、肝細胞内のゴルジ 体内へ銅分子を輸送できない。その結果、銅は胆汁中に 分泌されないため、肝細胞質に蓄積する [1]。しかし、最 近の研究ではウイルソン病の銅キレート療法後、特に男 性患者では銅と鉄の共存することが示されている [9]。ウ イルソン病患者とよく似た観察結果がウイルソン病動物 モデルであるこの LEC ラットで得られた。鉄複合体は、 リンと酸素から構成される一方で、銅は硫黄と複合体を 形成していた。肝細胞とクッパー細胞の高電子密度の顆 粒は酸性ホスファターゼに陽性で、銅や鉄が肝実質細胞 と網内系細胞のライソゾームに銅チオネインあるいはヘ モジデリンとして、蓄積していることが判明した。人のFig. 3 酸性ホスファターゼ反応後の LEC ラットの肝細胞の微小領域元素分析。 この電子顕微鏡像は、肝細胞質内に散在している高電子密度の顆粒を示す(左)。 この顆粒から得られたスペクトラムには、内因性のカルシウム、鉄、銅の3元素、そして外因性の鉛と金の2元素が見える。 従って、銅と鉄はライソゾームに蓄積していることが明らかになった。 4%パラホルムアルデヒド固定後、Novikoff の方法に従い酸性ホスファターゼ反応を行った。 無染色切片、JEM−2010 による画像 400 300 200 100 0 0 2 4 6 8 10 keV O P Pb Cl Pb Ca Fe Cu Au Pb
Fig. 2 JEM−2010 による LEC ラットのクッパー細胞の微小領域元素分析。 この電子顕微鏡像は、様々な高電子密度の顆粒を示す(左)。 EDX を併用した二次元の画像では、顆粒とそれらの基質内で銅(黒色化)と鉄(白色化)の異なる分布を示す。 0.1%オスミウム酸固定、ウラン染色 ウイルソン病での鉄過剰には複数の原因が挙げられる [8]。 このモデル動物の鉄過剰の主な原因は低セルロプラスミ ン血症によると思われる。このラットは、ウイルソン病 患者と同様に低セルロプラスミン血症を持つ [10]。セル ロプラスミンの持つフェロキシダーゼ活性は鉄の体内輸 送に必須である。また、動物では、餌に鉄を添加すると、 より多産になるのでしばしば実施されている。LEC ラッ トでは、フェロキシダーゼ欠損と食品への鉄添加が原因 で二次的に鉄過剰となるのであろう。 原発性銅過剰症にも関わらず、16 週令くらいの若い LEC ラットの肝組織は、ロダニンとルベアン酸のような 組織化学的銅染色に対して陰性である。毛細胆管周囲に 多数の銅陽性ライソゾームの集合することが、組織化学 的な染色に必須である。組織化学的な銅染色反応はその 感度が悪いために、肝組織の生化学的な銅測定が LEC ラッ トの多くの調査で広く利用されている [10]。実際、ウイル ソン病に対する国際的な診断法として生化学的な銅測定 が強く推奨されている [15]。組織化学的な検査は補助的基 準である。この研究に使用された MA は、総合病院など で一般に行われる検査ではないが、TEM による細胞内小 器官の確認と EDX により得られた全ての X 線がエネル ギーのスペクトラムとして保存されるので、ライソゾー ム内部での複数の金属蓄積が二次元で明らかとなる。そ のため、MA は多数元素の生体内での相互作用を研究す るための道を切り開いた [4、13、14]。JEM − 2800 を使用 すると、LEC ラットの肝臓からライソゾーム内の銅と鉄 の分布情報と他の元素との共存情報が短時間で同時に得 られた。そのうえ、肝細胞とクッパー細胞では銅沈殿物 の異なる傾向が得られた。JEM−2010 と JEM−2800 では、 元素の検出感度は違わないが、一つの分析に対する時間
Fig. 4 酸性ホスファターゼ反応後の LEC ラットの肝細胞の微小領域元素分析。 この電子顕微鏡像は、細胞質内の少数の高電子密度の顆粒を示す(Grey)。
一連の二次元の画像は、特性 X 線を使用して、鉄(Fe K␣)、銅(Cu K␣)、酸素(O K␣)、硫黄(S K␣)、カルシウム(Ca K␣)、リン(P K␣)、炭素(C K␣)の内因性元素を視覚化している。鉛 (Pb L␣)は、外因性の酸性ホスファターゼ反応産物で、これらの顆粒がラ イソゾームの起源であることを示している。 4%パラホルムアルデヒド固定後、Novikoff の方法に従い酸性ホスファターゼ反応を行った。 無染色切片、JEM−2800 による画像
Grey Pb
L
␣ Ca
K
␣
Fe K
␣ O
K
␣ P
K
␣
Cu K
␣ S
K
␣
C K
␣
Fig. 5 偶然検出されたカドミウムに富む肝細胞内顆粒。 一連の二次元の画像には、Fig. 4 で示した諸元素のほか、 カドミウム(Cd L␣)が蓄積している。電子密度の高い顆 粒(Grey)にはカドミウム、銅(Cu K␣)、硫黄(S K␣)が 同じように分布しており、カドミウムと銅がメタロチオ ネインとして共存していることを示唆する。 4%パラホルムアルデヒド固定、無染色切片、JEM− 2800 による画像Cu K
␣ S
K
␣
Grey Cd
L
␣
消費が全く異なる。前者では1イメージの取得に数時間 かかるのに対して、後者では 10− 20 分で終了する。二次 元の分析において蓄積した X 線の情報量もまた異なる。 分析領域内の任意の部位から産出された X 線は、コン ピュータで保存され、分析後に再生可能である。私たち は、20 年前、古い型の EDX を使用して、男性ウイルソ ン病患者の肝臓に銅と鉄の共存する未知の情報にはじめ て出会った。現在、ウイルソン病では、血清セルロプラ スミン低値、つまりフェロキシダーゼ欠乏のため、鉄も 肝臓に蓄積するとの理解が一般化している。この仮説は、 ウイルソン病の動物モデルである LEC ラットでの銅と鉄 の 2 重過剰の確認 [12]、Miyajima et al. により発見された 無セルロプラスミン血症が中枢神経と肝臓を含めた全身 性の鉄過剰症である [16] との 2 つの事実により支持され ている。無セルロプラスミン血症は、銅の病気ではなく、 遺伝性の鉄過剰症に分類されている [17]。 ライソゾームの鉄化合物と銅チオネインの沈着は、細胞 障害像を見ているわけではなく、細胞の持っている防御機 構による解毒された過程を見たものである。残念ながら現 在の技術では、細胞内小器官の周りの毒性の強い(活性の ある)遷移元素の存在を定量することはできない。蓄積し たタンパク体からその前駆体の毒性を推測しているにすぎ ない。胆道系に生理学的排泄ルートのある銅では、銅輸送 タンパクの欠損したウイルソン病では銅過剰になるが、他 の病的状態での銅過剰は稀である [1]。これらは、原発性 胆汁性肝硬変と非アルコール性脂肪肝炎を含む。原発性胆 汁性肝硬変における銅過剰は、有効なウルソデオキシコー ル酸治療で解決された [18]。対照的に、鉄は生理学的な排 泄ルートがなく、鉄過剰が様々な病的状態で生じる。C 型 慢性肝炎において肝細胞のライソゾームに溜まっている少 量の鉄は、C 型肝炎ウイルスに感染した肝細胞に対する鉄 誘導酸化ストレスのあることを暗示している。その根拠は、 ライソゾーム内の鉄を完全に除去すると難治性のC型慢性 肝炎が鎮静化する事実である [19]。しかし、ライソゾーム 内に多量の鉄を持つ無セルロプラスミン血症の肝細胞は鉄 の悪影響を受けない。歳をとった患者でも、その肝組織は、 鉄沈着以外の形態・機能はほぼ正常である [16]。無セルロ プラスミン血症の患者が肝臓に多くの鉄蓄積があるにも関 わらず、肝臓障害の軽微であることの背景は今後の研究課 題である。 カドミウムは汚染された水から米などに吸収されヒトに 神経障害を来たすことが知られている。肝臓ではライソ ゾーム分画に集積する [20]。我々はこの動物の肝臓で電子 密度の高い顆粒における銅と鉄の共存をスクリーニングし ている過程で偶然にカドミウムが硫黄を伴って蓄積してい る像を得た。このことは、カドミウムが、銅と同様に、メ タロチオネインとして蓄積していることを示唆する。
結論
LEC ラットの肝組織に対し微小領域元素分析を行った。 JEM− 2010 による予備調査では、肝細胞とクッパー細胞 の電子密度の高い顆粒に、銅と鉄の蓄積が確認され、基 質内の分布にも差異のあることが明らかにされた。次い で、酸性ホスファターゼ反応を施行した肝組織を JEM− 2800 により観察した。銅と鉄は酸性ホスファターゼ反応 陽性の顆粒、すなわちライソゾームに蓄積していること、 銅は硫黄と、鉄はリンと酸素と共存していることが確認 された。JEM− 2800 では短時間に詳細な二次元画像が正 確に取得できた。参考文献
[1] Gitlin J. D., Gastroenterology, 125, 1868−1877(2003). [2] Pietrangelo A., N Engl J Med, 350, 2383−2397(2004). [3] Britton R. S., Bacon B. R., and Ricknagel R. O.,Chem Phys Lipids, 45, 207 − 239(1987).
[4] Hanaichi T., Kidokoro R., Hayashi H., and Sakamoto
N., Lab Invest, 51, 592 − 597(1984).
[5] Novikoff A. B., Beaufay H., and De Duve C., J
Biophysic Biochem Cytol, 2, supl 179(1956).
[6] Yamaguchi Y., Heiny M. E., and Gitlin J.D., Biochem
Biophys Res Commun, 197, 271 − 277(1993).
[7] Hayashi H., Wakusawa S., Yano M., et al., Hepatol Res, 37, 777 − 783(2007).
[8] Walshe J. M., and Cox D.W., Lancet, 352, 150(1998). [9] Shiono Y., Wakusawa S., Hayashi H., et al., Am J
Gastroenterol, 96, 3147 − 51(2001).
[10] Mori M., Hattori A., Sawaki M., et al., Am J Pathol, 144, 200 − 204(1994).
[11] Togashi Y., Li Y., Kang J., et al., Hepatology, 15, 82 − 87(1992).
[12] Kato J., Kobune M., Kohgo Y., et al., J Clin Invest, 98, 923 − 929(1996).
[13] Hayashi H., Fujita Y., Tatsumi Y., et al., Hepatol Res, 39, 563 − 568(2009).
[14] Motonishi S., Hayashi H., Fujita Y., et al., Ultrastruct
Pathol, 30, 409 − 414(2006).
[15] Ferenci P., Caca K., Loudianos G., et al., Liver Int, 23, 139-142(2003).
[16] Miyajima H., Nishimura Y., Mizoguchi K., et al.,
Neurology, 37, 761 − 767(1987).
[17] Kaneko Y., Miyajima H., Piperno A., et al., J
Gastroenterol, 45, 1163 − 1171(2010).
[18] Poupon R., Chretien Y., Poupon R.E., et al., Lancet, 2, 834 − 836(1987).
[19] Hayashi H., Takikawa T., Nishimura N., et al., Am J
Gastroenterol, 89, 986 − 8(1994).