象鼻虫3種の単為生殖に関する生態学的研究 (英文)
44
0
0
全文
(2) Vol. 10, No. 2 Journal of Hokkaido Gakugei University Dec. 1959. Some Oecological Observations of Three Species of Curculionid Weevils, With Special Reference to Parthenogenetic Reproduction Yasushi TAKENOUCHI Biological Laboratory, Sapporo College, Hokkaido Gakugei University. w^i ^: a»A3@®-i|i^^m{c:nT^^i^6^j^^ CONTENTS Introduction. Material. and. ..................................... methods. Observations. 298. ................................ .................................... 299. 299. A. About Listroderes costirostris Schonherr ................. 299 1. Activities of the adult female ..................... 299 2.. The. 3.. 4.. esg............ Oviposition. Duration. of. the. ...................... 300. ............................... egg. stage. 300. ........................ 300. 5. Duration of the larval stage ...................... 300. B. About Catapionns gracilicornis Roelofs .................. 300 1.. Activities. of. the. adult. .......................... 300. 2. Parthenogenetic reproduction ..................... 303 3.. Theegg.... 4.. Oviposition. 5.. Duration. 6.. The. of. .............................. records. the. larva. egg. 303. ............................ stage. 303. ........................ ................................ 310. 311. 7. Duration of the larval stage ...................... 312 8.. Larval. 9.. The. 10.. development. pupa. Length. of. ........................... ................................ the. pupal. stage. 312. 314. ........................ 314. C. About Pseitdocneorhimis bif'asciatiLS Roelofs ................ 314 1.. Activities. of. the. adult. .......................... 314. 2. Parthenogenetic reproduction ..................... 316 3.. 4. 5.. Theegg.. ............................... Oviposition Duration. 6.. The. of. records. the. larva. egg. 317. ........................... stage. 317. ........................ ................................ 323. 324. 7. Duration of the larval stage ...................... 325 8.. Larval. Discussion Summary. ........................... .................«.......•-.-.-•-••••. Literature. Explanation Plates.. development. ......................-...•..••-•••••. ..................................... of. plates. 327 327. ................................ ....................................... 326 326. 328. 330. A grant from the Scientific Research Expenditure of the Ministry of Education is gratefully acknowledged here.. —297—.
(3) Yasushi Takenouchi. Introduction It has been known that curculionid weevils belonging to the subfamilies Otiorrhynchinae and Brachyderinae include many species which propagate parthenogenetically. There occur among many species of several genera belonging fco these subfamilies both parthenogenetic and bisexual races. Cytological investigation has made it clear that the reproduction occurring in these curculionid weevils is to be considered as apomictic parthenogenesis (Seller. 1947; Suomalainen 1940, 1947, 1949, 1950, 1954, 1955). The great majority of parthenogenetically reproducing weevil species in these subfamilies are polyploid, while all the bisexual weevils so far known are diploid. By reference to the current cytological knowledge of. these weevils accumulated exclusively by Suomalainen (1954, 1955), Seller (1947), and Mikulska (1953), one learns that the total number of parthenogenetic weevil species and races so far studied cytologically is thirty-one. The localities of these parthenogenetic weevils are restricted to Northern, Central and Southern Europe. a Based on the results of breeding experiments and of examination of cytological features, the present author reported that Catapionus gracilicornis Roelofs was parthenogenetic (Takenouchi 1957a). Further, it was suggested from the results of the cytological investigations that other two species of Japanese weevils, Listroderes costirostris Schonherr and Pseudocneorhimis bifasciatus Roelofs, also reproduce parthenogenetically (Takenouchi 1957b). The vegetable weevil, Lisiroderes cosfirostris, a member of the subfamily Cylindrorrhininae, is known as a serious vegetable pest in South America, Australia, the U. S. A., and South Africa. This species has established its remarkable distribution in various provinces along the Pacific coast of southwestern parts of Japan, such as southern parts of Okayama Prefeature, and in Kyushu, Shikoku, Chugoku, Kinki, Tokai, and Kanto Districts. In early summer of 1940, the first adult specimen of this species was captured in Okayama Prefeature. Since the pest damaged carrots in Okayama Prefecture in 1942, its occurrence in Japan has attracted the attention of many entomologists (Yasue 1952). Dr. Hiroharu Yuasa once suggested that the vegetable weevil seemed to propagate parthenogenetically on the basis of the fact that there was no male in population. Catapionu3 gracilico'/nis, one of the native weevils in Japan, belongs to the tribe Dermatodini of the subfamily Brachyderinae. It has a wide range of distribution in Hokkaido, though no accurate accounts of its ecological features have been published. In Hokkaido, the weevils first appear on the surface of the ground from the beginning to the middle of IVIay. The seasonal maxima in occurrence were observed from the latter part of May to the latter part of June, showing very low frequency in both July and August ; the animals disappeared after August. The third species, Psendocneorhimis bifasciatus is a member of the subfamily Eremninae, and one of the native. curculionid weevils in Japan. It is widely distributed throughout Japan, from Hokkaido to Kyushu, inhabiting gardens, orchards, and hillocks with Lespedeza bicolor near human dwellings. It has been said that activity of adult beetles is associated with the hot season, from July to August, in Hokkaido. On the basis of the chromosome study of Japanese curculionid weevils, the suggestion has been made that Psendocneorhinus bifasciatus may be a parthenogenetic weevil, since the female of this species is tetraploid ; all the specimens known to date by the author were female. So far as the author is aware, the details of reproduction of these weevils have remained almost unknown. This enhanced the author's interest in these species and prompted his 298—.
(4) Some Oecological Observations of Three Species of Curculionid Weevils attempts to observe the breeding habits. Mention should be made here that two of three curculionid weevils here considered are members of the subfamily Cylindrorrhininae and Eremninae, respectively, in striking contrast to the fact that the European parthenogenetic weevils are members of the Otiorrhynchinae and Brachyderinae only. In the present paper, the author wishes to report the results in detail of breeding experiments of these weevils with special concern for their parthenogenetic reproduction. Before proceeding further, it is the author's pleasant duty to acknowledge here his great. indebtedness to Professor Sajiro Makino, Hokkaido University, for his kind guidance with valuable advice in the course of the study and for the improvement of the original manuscript of this paper. The author's thanks must be also extended to Assistant Professor Eizi Momma for his helpful suggestions and encouragement; to Professor Michio Chujo, Kagawa University, and Dr. Yasunobu Yasue, Ohara Institute for Agricultural Research, Okayama University, who generously placed the material at the author's disposal ; and to Messrs Seki. Oyamada, Takaaki Nara, Akio Takauji, Yutaka Nakatsuji, Kei Sakai, Shuichi Sohma, Yuji Araoka, Kuniyasu Yoshimoto and Miss Yanagi Kimura who all have lent kind cooperation. in collecting the material used for this study and kindly helped in breeding animals. Many thanks are also due to Professor Hiromichi Kono for giving him opportunities for consulting the literature; and to Mr, Masayasu Konishi, Entomological Institute Hokkaido University, not only for identification of the species employed but also for valuable advice.. Material and methods The species with which the present study has been carried out are Listroderes costirostris Schonherr, Catapionus gracilicornis Roelofs and Pseudocneorhinus bifasciatzts Roelofs. A number of specimens of the first species were collected in Okayama, and Kagawa Prefectures,. and kindly sent to Sapporo in February 1957 and IVtarch 1958 by Drs. Mjchio Chujo and Yasunobu Yasue whose generous assistance is acknowledged. Coldness prevents success of breeding of this species in the laboratory. Many specimens of the second species collected. in Hokkaido, at the foot of Mt. Moiwa (Sapporo), at an altitude of 860 m on M:t. Daisetsu (Kamikawa), and at the foot of Mt. Esaomantottabetsu (Hidaka) provided the material for this study. The localities where specimens of this species were collected are listed in Table 1. The data herein presented are based upon the research work covering three seasons in the. years 1955, 1956, and 1957. Many individuals of Pseudocneorhinus bifasciatus were obtained in the vicinity of human dwellings in the City of Sapporo and used as material for study (Table 15). Breeding experiments were carried out with adult females of the above named three species, A single virgin female was caged separately in each rearing container with great care ; the eggs deposited were collected in glass vessels. The eggs were then kept at room temperature. Larvae were reared in the containers with a food supply. The method of rearing is described in detail in another chapter. Observations A. About Listroderes costirostris Schonherr. 1. Activities of the adult female Twenty-five virgin females were caged separately for two years (1957 and 1958) in —299—.
(5) Yasushi Takenouchi. petri dishes which were covered with glass lids of the ordinary type. Throughout the entire experiment, a piece of leaf of Pe-Tsai (Brassica pekinensis) was kept in each container. During egg-laying period the beetles were actively on the move in the glasses and fed freely on cabbage leaflets. All beetles deposit a few eggs every day for about two weeks and then all cease from egg-laying thereafter. 2. The egg. The egg (Fig. 2) is approximately 0.85 mm in length, by 0.72 mm in width ; it is spherical or in some cases oval. The original color of the freshly deposited egg is shiny milky white; the color changes to dull yellow one to three days later, and then to light brown as further development takes place. The covering of the egg is elastic and almost smooth in texture. Shortly before hatching, the tips of the mandibles of the larvae are visible through the egg covering. 3. Oviposition All virgin weevils placed in the author's stove-heated living room deposit a few eggs every day for about two weeks and then all cease from egg-laying afterward. During the experiment the daily variation in room temperature was from 5°C to 20"C. The beetle deposited one or two eggs together with a drop of clear mucus on a leaf of the host plant or on the bottom of the container through her ovipositor (Fig. 3) in a short time. When the mucus dried up, the eggs adhered lightly to the leaf or bottom of the petri dish. Because of the abnormality of egg laying, oviposition records were not noted. 4. Duration of the egg stage For the breeding experiment in 1957, use was made of eggs which were deposited in the bamboo-containers for sending by post to Sapporo. The eggs were removed from the bamboo-containers and transferred to incubation petri dish containing a circular pieces of blotting paper which were always kept moistened. In order to avoid injury by fungus the eggs were carefully watched and from time to time moved to new containers. During the incubation period the temperature fluctuated between 5°C and 20°C every day. On February 18, a total of 109 eggs were placed in the incubation container, and from this lot 96 larvae were hatched on the 5th of JVEarch. The hatch ability is 87 per cent in frequency. 5. Duration of the larval stage One of the comparatively larger petri dishes was used as a rearing container for the larvae. Small pieces of Pe-Tsai {Brassica pekinensis) leaf were placed on the bottom of the petri dish. Ninety-six newly hatched larvae were removed from the incubation dish and transferred to the rearing container. The newly hatched larvae fed lightly on the surface of the leaf. The temperature rose from 3°C to about 18°C. During the entire rearing work, fungus attack caused a high percentage of mortality in the larvae. Forty-four of the larvae survived the first instar but during the second instar 42 of them died of injury of mycelia. Only two larvae of the third-instar were raised in the rearing experiment, but they died on April 30. Malting was observed to have taken place in the feeding cells on the pieces of the leaf. No fully mature larva and pupa were obtained in the present experiment. It is evident that the vegetable weevils reproduce parthenogenetically because most of their eggs hatch successfully, notwithstanding the fact that they had been laid by virgin creatures. B. About Catapionus gracilicornis Roelofs. 1. Activities of the adult —300.
(6) Some Oecological Observations of Three Species of Curculionid Weevils Observations in Sapporo: Seasonal changes in the activities of the beetles are apparent. It seems that weather conditions, primarily soil and air temperature, influence the time of emergence. In Sapporo, as the ground begins to thaw and warm up, the beetles climb upward in the soil, making tunnels to the surface first to emerge on the surface of the ground in the middle of May. The first adults were recorded on May 16, in 1956 and on May 14 in 1957. In 1958, eight weevils were first recorded on May 13 at an average daytemperature of 18.5°C and soil temperature of 13.9°C at a depth of 10 centimeters. On May 14, three animals were collected in the same place at an average day-temperature of 22"C and soil temperature of 15.5°C. On May 15, when the day-temperature showed 22.1°C and the soil temperature 14.5 C at a depth of 10 centimeters, sixteen specimens were obtained. The largest number of the beetles is observed during the latter part of May and June. With a sudden decrease during July and August, one rarely could see the weevils unless careful search was made for them around the crown of food plants. Late in August, no individuals were obtained anywhere in any of the three years. Migration of the weevil is quite a little in the field; in addition, the beetle occurs always in a small area, about 50 square meters, at any one locality, so the author was put to great trouble in finding this beetle in the field. Such habits were ascertained in every place where the animals were found by the author. Beetles are few in number which emerge in an area in one season. During five successive years a total of 369 weevils were collected in several localities of Hokkaido, as shown in Table 1. It seems likely that the beetles found in these areas have been maintaining themselves Table 1.. Collection records from several parts of Hokkaido from. May to August in 1953, 1954, 1955,>, 1956 and 1957. No.. Sex. Year. Date. Locality. 1953. July 12-16. Ikomanbetsu, on Mt. Daisetsu. 22. ?. 1954. May 15-July. Sapporo, foot of Mt. Moiwa. 45. ?. 1955. At the middle of May. 20. ?. 1955. May-August. 1955. June-August. Sakae-machi (Sapporo). 1955. August 22. Jozankei. specimens. collected. 113. 9. 45. ?. Food plants Cirsium pect'inellu.m. 1. ?. Petasites jap^nic'us. 16. ?. Cirsiwn pectinellmn. 1956. May 16. Sapporo, foot of Mt. Moiwa. 1956. May 21. Teine-machi, Ishikami-gun. 6. ?. Petasites japonzcns. 1956. July 6. Sapporo, foot of Mt. Moiwa. 2. ?. Cirsiwn pectinellum. 1956. July 26. Lake Okotan, Chitose-gun. 1. ?. 1956. August. Sapporo, foofc of Mt. AIoiwa. 5. ?. 1957. May 14. Maruyama, Sapporo. 1. ?. 1957. May 15. Sapporo, foot of Mt. Moiwa. 3. 9 9. 1957. May 16. 3. 1957. May 17. 29. ?. 1957. June 29. Esaomantottabetsu, Hidaka. 4. ?. 1957. July 16-29. upper Ikomanbetsu, Mt, Daisetsu. 11. ?. July 16-20. lower Ikomanbetsu, Mt. Daisetsu. 42. ?. 1957. 301—. Petasites japonwzis.
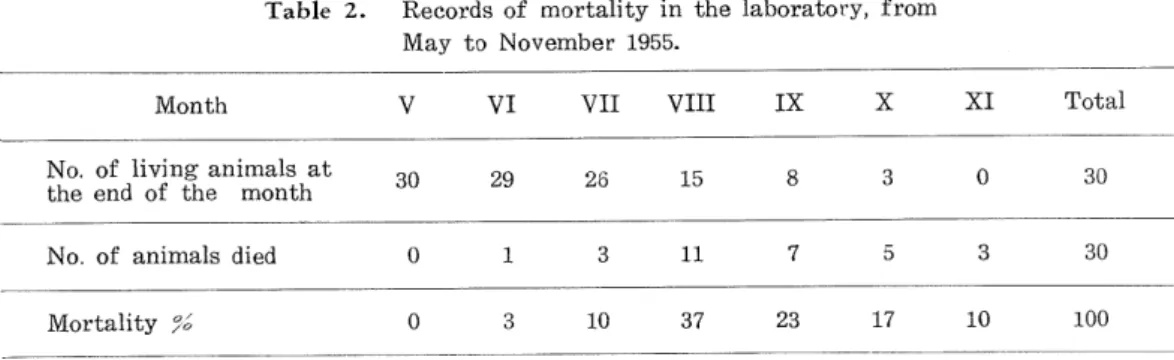
(7) Yasushi Takenouchi for many years on the wild host plants, Cirsium pectinellum, and Petasites japonicus. which are their favored hosts. Distinctions in food-habits among different local types of the beetle were apparent in these localities. Although the host plants as mentioned above are present in every locality, the insects occurring in Jozankei, Teinemachi, and the lower area of Ikomanbetsu feed on Petasites japonicus, whilst those of the remaining localities exclusively attack Cirsium pectinellwn. However, in the laboratory all animals coming from the lower area of Ikomanbetsu eat C. pectinellum. Solitary beetles were found on scattered host plants in a narrow localized area in every locality. Following their first appearance in the middle part of MEay at the foot of Mt. Moiwa, many adult beetles were collected every day during the season throughout three years. The last living adult was taken on August 23 in Jozankei.. Attention should be called to the fact that dissections of 224 specimens among 269 adults collected in the above localities showed that all of the individuals were females ; further, the remaining females laid numerous eggs under captivity without males. No male has been found anywhere, so far as the field collection has been done. The migration of beetles from one area to another has never been found; the insects emerge regularly in a narrow strip of land every year. During May, June, and July the beetles feed on the foliage of the host plants. When disturbed, these feeding adults feign death and drop to the ground. The adults of this species avoid either very strong bright sunlight, or very weak illumination and usually move about by trial until the most favoura-. ble condition is found. Vassiliev (1914) and Palm (1935) reported that bright sunlight is harmful to the adults of alfalfa snout beetle, Brachyrhimis ligustici. The most favourable air temperature for animals is from 10°C to approximately 25°C. It is observed that in hot weather, some of the beetles hide in the debris and soil beneath the host plants. Although all of these adults were inactive in their hiding quarters, they immediately became active when handled and placed together in a collecting bottle. Phases of adult activity which were observed in the laboratory provided the following information. A -few animals were observed feeding on thistle leaves until the last part of. October, 1955; by September 10 in 1956, all the beetles died; the last living animal was seen on August 21, 1957. Table 2 shows the mortality of thirty beetles collected in the. middle of May, 1955. Table 2. Records of mortality in the laboratory, from May to November 1955.. Month. v. VI. VII. VIII. IX. x. XI. Total. of living animals at the end of the month. 30. 29. 26. 15. 8. 3. 0. 30. of animals died. 0. 1. 3. 11. 7. 5. 3. 30. No.. No.. Mortality % 0 3 10 37 23 17 10 100. The seasonal maximum of mortality in the laboratory was observed in August. In general, beetles cease from both oviposition and feeding activity before their death. Usually, every animal in the egg-laying period, when kept in a petri dish without leaf of host plants, dies in a few days. —302—.
(8) Some Oecological Observations of Thres Species of Curculionid Weevils. Generally, egg laying had begun by the end of May. During the oviposition period the beetles feed actively on leaves of the host plants. 2. Parthenogenetic reproduction Dissections of many newly emerged adults collected in the foot of Mt. Moiwa in the. middle of May in each year of 1955, 1956, and 1957 showed that they were all females with numerous developing eggs in their ovaries. After emergence, the fresh beetles, begin egg-. deposition within 7 to 10 days. As described later, a total of 78 beetles (among which 20 were collected on May 18, 1955 ; 22 were obtained on May 16-21, 1956 ; 36 were secured on. IVIay 14-17, 1957 at the foot of Mt. IVtoiwa) were placed separately in petri dishes with a little soil and thistle leaf. All of these individuals began to deposit eggs after a few days ; most of the eggs that were deposited developed normally. 3. The egg. The freshly deposited egg is shiny milky white in color ; either end of the egg become translucent three to five days later and then its color changes to light green as further development takes place. The covering of the egg is elastic and just deposited egg is. slightly pliant; about one hour later it becomes comparatively firm. The egg (Fig. 20) is bilaterally symmetrical, and is approximately 1,23mm (from 1.10 to 1.50mm) in length by 0.53 mm (from 0.39 to 0.63 mm) in width. Table 3 shows the results of investigations of the proportions between length and width in 54 eggs of this species; mean value is 0,44. Table 3. Proportions between length and width in 54 eggs of Catapiomis gracilicorms.. Length. Width. No.. of individuals. 0.30-0.33. 0.34-0.37. 0.38-0.41. 0.42-0.45. 0.46-0.49. 0.50-0.53. 2. 4. 9. 19. 14. 6. Since the weight of the egg is very slight the eggs were weighed by a chemical balance. It is evident by referring to Table 4 that an egg is 25 milligrams on the average. 4. Oviposition records Table 4. Weight of the egg in miUigrams, In order to obtain some detailed information on oviposition, breeding experiments were ]^a of eggs. Whole weight. Mean weight. performed respectively for three seasons. In ^Q | ^0 : O^g every year, freshly emerged beetles were col- ^g | gg | 0^5 lected at the foot of Mt, JVIoiwa, and were - - ' - - - -. caged separately in petridishes which were covered with ordinary glass lids (Figs. 5-6). Narrow gaps were made by small iron needles in containers (Fig. 7). Twenty fresh beetles were collected in the field on May 18, 1955, twenty-two on JVIay 16 to 21, 1956, and thirty-. six on May 14 to 17, 1957; they were put separately into petridishes with about an inch of sifted moist soil. Throughout the entire experiment, fresh thistle leaf was kept in each container. By soaking the leaf before placing it in the petridishes, fresh state is kept for several hours. JVtoderate humidity and suitable objects to which the animal may cling and insert her ovipositor are indispensable to induce egg laying. Usually, egg-laying takes place on the leaves of host plants. Conversely no egg are deposited in dry air. It was observed, however, that a few females deposited eggs on moistened absorbent cotton or in fissure of a piece of wet straw. Generally, well-fed females show high egg-laying activity. The —303—.
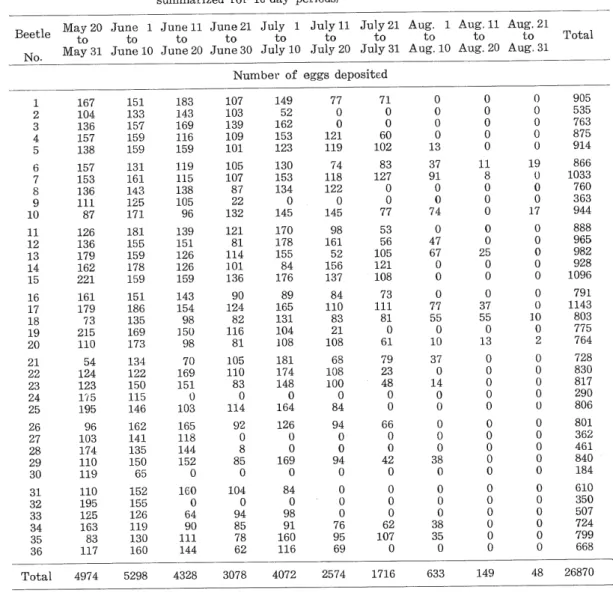
(9) Yasushi Takenouchi mother beetle proceeds about her egg-laying in the following manner (Pig. 8-19). In an ordinary way, (1) the female beetle stands on a portion near the apex of the leaf in parallel. with its edge, (2) turns her back towards the reverse of the leaf, (3) three legs of the one side stretched by turns to catch the tip, when the tip is caught by the legs, (4) the leaf is drawn horizontally inwards and doubled around the distal end of the beetle from both sides of the fold by the power which is concentrated on her legs, inside that fold egg-laying is. performed; (5) the beetle extends her flexible cylindric ovipositor (Fig. 21) and primarily ejaculates a drop of clear mucus which sticks opposite surfaces of the leaf to each other,. (6) then, deposits eggs gradually one by one in order, and (7) finally secretes a drop of the mucus to paste the eggs on the leaf, and then she goes away. When the transparent adhesive mucus is exposed to air it can dry rapidly, being transformed into a fibrous pellicle Table 5. Oviposition records for Catapiomis gracilicornis at Mt. Moiwa, Sapporo, during May, June, July and August, 1957.. (The data are based upon daily records from 36 beetles and are summarized for 10-day periods). No.. May 20 June 1 June 11 June 21 July 1 July 11 to to to to to to May 31 June 10 June 20 June 30 July 10 July 20. July 21 Aug. 1 Aug. 11 Aug. 21 to to to to July 31 Aug. 10 Aug. 20 Aug. 31. Total. Number of eggs deposited. 1 2. 167 104 136 157 138. 151 133 157 159 159. 183 143 169 116 159. 157 153 136 Ill. 131 161 143 125 171. 119 115 138 105 96. 126. 139 151 126 126 159. 121. 179 162 221. 181 155 159 178 159. 161 179 73 215 110. 151 186 135 169 173. 143 154 98 15U 98. 90 124 82 116. 21 22 23 24 25. 54 124. 70 169 151 0 103. 105. 175 195. 134 122 150 115 146. 26 27. 96 103 174 110 119. 162 141 135 150 65. 165 118 144 152. 92. 110. 0. 85 0. 160. 104. 64 90 Ill. 94 85. 78. 4328. 3078. 3. 4 5 6 7 8. 9. 10 11 12 13 14 15 16. 17. 18 19. 20. 28. 29. 30. 87. 136. 123. 31 32 33 34 35 36. 195 125 163 83 117. 152 155 126 119 130 160. Total. 4974. 5298. 0. 144. 107 103 139. 109. 101. 105. 107. 87 22 132. 81. 114 101 136. 81. no. 83 0 114. 0 8. 0. 62. 149 52 162 153 123. 121 119. 130 153 134. 74 118 122. 145. 145. 170 178 155 84 176. 98. 161 52 156 137. 89. 84. 0. 77 0. 0. 0. 71 0 0 60 102 83. 127 0 0 77 53 56 105 121. 108 73. 0. 0 0. 0 13 37 91. 0 0. 74. 0. 0. 0. 77. 0 61. 55. 0 37 55. 10. 13. 37 0. 0. 68 108 100 0. 79. 126 0. 94 0. 66. 0 0. 0. 0 42 0. 38 0. 0 0. 0 0 0. 0 0 0. 98 91. 0. 0 0. Ill. 181 174 148 0 164. 94 0. 0 0 0. 0 25. 110 83 21 108. 0 169 0 84 0. 8 0. 47 67 0. 165 131 104 108. 84. 0 0 0 0 0 11. 81. 23 48 0 0 0. 0. 14 0 0. 0. 0. 0. 0 0 0 0 0 0 0. 0 0. 888 965. 0 0. 0. 0 0 0. 0 10 0 2. 0 0. 0. 0. 0. 982 928 1096 791 1143 803 775 764 728. 830. 817 290 806. 0. 0 0. 610 350 507 724. 799. 48. 26870. 0. 0. 160. 0. 0 0. 4072. 2574. 1716. 633. 149. —304—. 17. 866 1033 760 363 944. 0 0 0. 0. 38 35. 0. 0 19 0. 801 362 461 840 184. 62 107. 69. 0 0. 905 535 763 875 914. 0 0. 76 95. 116. 0 0. 0. 0 0 0. 0. 668.
(10) Some Oecological Observations of Three Species of Curculionid Weevils colored white. A similar process was observed in egg-laying in Otiorrhyn^hus snlcatus. (Seller 1947). A summary of oviposition records obtained in 1957 is given in Table 5. Reference to Chart 1 will show that air temperature seems to affect the activity of the beetles.. cWWV}MSW.:TWKW. <".Nal:,w.pv(^18EEmE$7^.'^l^;:':::1^-. -m['vwfy:sws^!1 -/w^w;sa,sr{i: ?t,By»lj>eg »W. n:1;. 9*MHH^m<Ht*v^^,. W^^^ilmliJSh •^•^?l:.:^i!::s''!^^. ^¥^: ^ :f ;-t;^ ?;;!: '^ ^ '\I^W^ t;v 1^. '••;'^-.-A'^^^:^^^ ^^. t?ffn<i. .^B^::..:,^:ll: :1111'; ll:':i;:^'..' 11'1'11 ''1'. *y^1 11 11 :1 • : ..ii-.i.' ' s*fi>. '•',. Chart 1. Graphs showing a close correlation between tempsrature and egg-laying activities observed in 38 specimens of Catapionus gracilwornis during their oviposition period.. As shown m Table 6 and Table 7, a total of 9,973 eggs were deposited by 22 females, showing a variation in number of eggs ranging from 55 to 763 per beetle with an average of 453.3 in 1956, and ranging from 506 to 901 eggs per beetle with an average of 427.0 in. 1955. Table 6. Egg-laying activities in 32 specimens of Catapiomt.s gracilicornis (1955).. Maximum. Minimum. Number of eggs per day per living beetle. 50. 0. Total number of eggs deposited per bestle. 901. 506. Length oviposition period (days) Number of days beetles lived after beginning to lay eggs - 305 —. Average 12.3. 427. 77. 35. 44.3. 100. 46. 61.4.
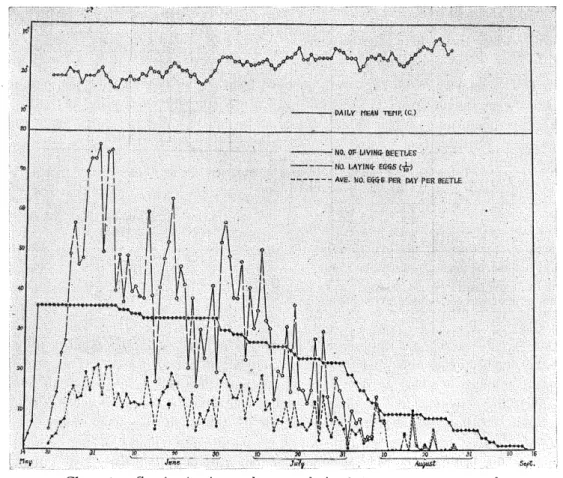
(11) Yasushi Takenouchi Table 7. Egg-laying activities in 22 specimens of Catapiomi.s gracilicornis (1956). Maximum. Minimum. Average. Number of egg's per day per living beetle. 57. 0. 16.3. Total number of eggs deposited per beetle. 763. 53. 453.3. 76. 13. 54.4. 91. 18. 69.7. Length ovi position period (days) Number of days beetles lived after beginning to lay eggs. In 1957, a total of 26,870 eggs were obtained from 36 females, showing the variation in number of eggs ranging from 115 to 1,143 eggs per beetle with an average of 746.4 (Table 8). The maximum number laid by any one beetle in one day was 57, with a daily average of 12.8-16.27 eggs per beetle, Table 8.. Egg-laying activities in 36 specimens of Catapiomcs gracil.icornis. (1957).. ]Vtaximum Minimum Number of eggs per day per living beetle Total number of eggs deposited per beetle Length of oviposition period (days) Number of days beetles lived after beginning to lay eggs. Average. 54. 0. 15.9. 1,143. 115. 746.4. 100. 11. 64.5. 116. 13. 70.3. Daily records during the entire egg-laying period in 1957, with the number of beetles alive, the number ovipositing, and the average number of eggs oviposited per day per living beetle, are graphed in Chart 1. The evidence presented in Chart 1 shows that air temperature affects the activity of the beetles. A similar characteristic was observed in the alfalfa. snout beetle, Brachyrhinus ligzistici by Palm (1935). Although egg-laying under insectary conditions extended from May 16 until August 26, the greatest number of eggs were depos-. ited during the last part of May and the beginning part of June (Charts 1-3). Individual females deposited their eggs regularly for several days, and then rested for a day or longer. MAY Chart 2. A summarized record on oviposition data from Catapionus grac'Hicorn'is obtained in 1956. The curve shows the percentage of total number of eggs deposited, in ten-day intervals, during an entire period of oviposition.. —306—.
(12) Some Oecological Observations of Three Species of Cu.rculionid Weevils. •^. i-ic n-so. n-36. JUNE. 'i-w n-w. JULY:. im". mOT. II-A5. Chart 3. A summarized record on oviposition data from Catapiomis gracilicornis given in Table 5. The curve shows the percentage of total number of eggs deposited in ten-day intervals, during an entire period of oviposition.. before resuming their egg laying activities. Favourable air temperature for egg-laying ranges between 16°C and 25°C on the average. There appears to be a short period after egg-laying is completed in which the adults remain active and feed, up to the time of their death (Tables 6, 7, 8, and Chart 1\ Generally a female deposited eggs from one to a few times. •PEtil.EtlTN-l.al' l.V'i'.IHif.&TiS. .?»:C£h't:Nft;?:l;WW8£(-TliES1: —-Aw.mi.csas pr's at f.ns:. w:./. 'AM. ;;/. •'I/:. 2AM,|4AM.p.n.|SAf1.| ]OA.>iJK/1,M.|2f>M,|4P.ri,l6 W.\ 8RM.|10|?rt.|KP(1, Chart 5. A summarized record on oviposition data from Catapiomis gracilicornis given in Table 9. The curve shows the number of .'KM. •wn. e m. sw- wm \'<w. wv w- w s^.M >S'A"1' M'w. egg-laying in two-hour intervals, observed from May 29 to June 4, 1957.. Chart 4. Graphs showing a close correlation between time and egg-laying. activities in weevils specimens of Catapionus gracilicornis observed in May 29 to June 4, 1957. — 307 -.
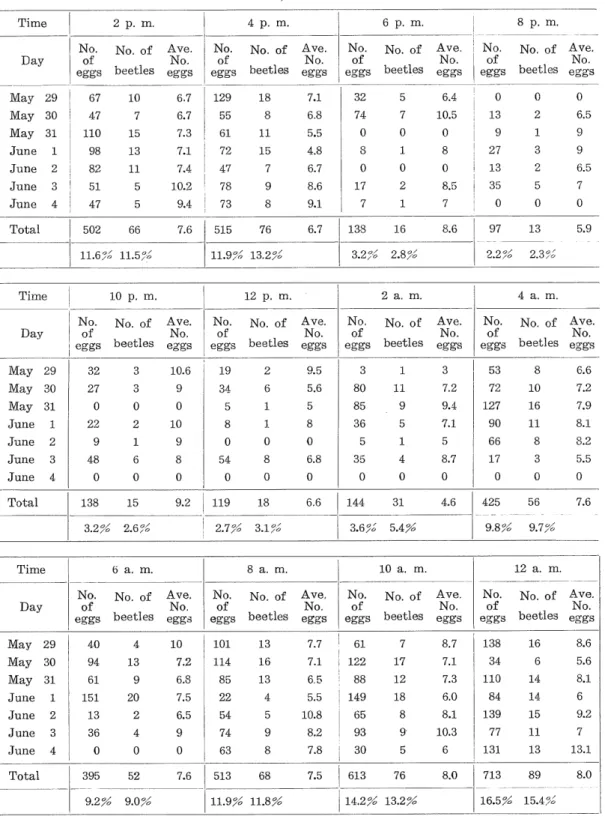
(13) Yasushi Takenouchi Table 9.. Oviposition records for Catapiomis gracilicornis in Sapporo, from May 29 to June 4, 1957.. (The data are based upon records obtained at intervals of two hours from 36 beetles).. Time Day. 2 p. m, No.. 10 7. No.. eggs. No.. of eggs. No.. No. of. Ave.. eggs. of eggs. beetles. 32. 5. 6.4. 7. 10.5. 0 1 0. 0 8 0. No.. No.. eggs. 7.1. 8. 6.8. 15. 7.3. 61. 11. 5.5. 13. 7.1. 72. 15. 4.8. 11. 7.4. 47. 7. 6.7. 51. 5. 10.2. 78. 8.6. 17. 2. 8.5. 47. 5. 9.4. 73. 9 8. 74 0 8 0. 9.1. 7. 1. 7. 502. 66. 7.6. 515. 76. 6.7. 138. June 3 June 4. 11.6^. Time. 11.9%' 13.2^. 11.5°». 10 p. m. No.. of eggs. JVtay 29. 32. JVTay 30. 27. No. of. Ave.. beetles. No.. 2. 9.5. 3. 1. 34. 6. 5.6. 0. 80. 5. 1. 5. 85. 1 0 8. 8 0. 36. 5. 9. 8 0. 11 9. 5. 6.8. 35. 0. 1 4 0. 144. 31. 4.6. 8. 54. June 4. 0. 0. 0. 0. 138. 15. 9.2. 119 2.7^. o/. I/'O. M:ay 31. 61. June 1. 151. June 2. 13. June 3. 36. June 4. No. of. beetles. 4 13. No.. 0. 0 18. 6.6. 0 1 0,. of eggs. 3.6^. .7°. Ave. No,. eggs. No.. of eggs. No. of. beetles. Ave. No.. eggs. No.. of eggs. 101. 13. 7.7. 61. 7.2. 114. 16. 7.1. 122. 6.8. 85. 13. 6.5. 7.5. 22. 4. 5.5. 88 149. 10. No. of. 0 13. 0. 0. 2. 6.5. 9. 1. 27. 3. 9. 13. 2. 6.5. 35. 5. 0. 0. 7. 97. 13. 0 5.9. 2.3%. No.. of eggs. No. of. Ave.. beetles. eggs. No.. 3. 53. 8. 6.6. 7.2. 72. 10. 7.2. 9.4. 127. 16. 7.9. 7.1. 90. 11. 8.1. 5. 66. 8. 8.2. 8.7. 17. 3. 5.5. 0. 0. 0. 425. 56. 0. 9.8%. 7.6. 9.7^. 12 a. m. Ave.. No.. No. of. Ave,. beetles. eggs. 7 17 12. 8.7. 138. 16. 8.6. 7.1. 34. 6. 5.6. 7.3. 110. 14. 8.1. 18. 6.0. 84. 14. 6. 8.1. 139. 15. 9.2. 10.3. 77. 11. 7. 6. 131. 13. 13.1. 8.0. 713. 89. 8.0. 5. 10.8. 65. 74. 93. 63. 9 8. 8.2. 0. 9 0. 7.8. 30. 8 9 5. 395. 52. 7.6. 513. 68. 7.5. 613. 76. —308—. 9. eggs. 54. 11.9% 11.8^. No.. of eggs. beetles. 6.5. 9.0%. No.. 5.4^. 9 20 2 4 0. 9.2^. eggs. 10 a. m.. 8 a. m.. 6 a. m.. 94. eggs. 19. 6. 4.0. beetles. 9. 48. May 30. Ave.. 10.6. June 3. May 29. No. of. 3. 9. of eggs. No.. eggs. June 2. Ave.. 4 a. m.. beetles. 10. No, of. beetles. 2 a. m.. eggs. 0. of eggs. No.. 2.2 %. Ave.. 22. No.. 2.8^. No. of. May 31. Time. ,0. 8.6. of eggs. No.. June 1. 3.2^. 16 <y/. 12 p. m.. 3 0 2 1. Total. Ave.. 18. 82. Day. beetles. 55. June 2. Total. No. of. 129. June 1. Day. Ave.. 6.7. 67. 47 110 98. Total. beetles. 6.7. May 29 May 30 May 31. No. of. of eggs. 8 p. m.. 6 p. m.. 4 p. m.. 14.2^ 13.2^. No.. 16.5^. 15.4,%. No..
(14) Some Oecological Observations of Three Species of Curculionid Weevils. ®. ^ s s &. cu ro. ^2. si CD c! cd 1=! T3. ^ Sb co. So w. t)-l. 0. s-. <u. •<s>. 0 e. s> &1. I <%. I 0. § 1->. c.. u-i. ro. 0^. w '^. ^ 0. 1^ 1-1. G. 4-1. c!. (U. +-•. s-1. ®. d. m 0. c?. +J. rt. b. CH. 0. rt. cti. 4-'. ra. fl. +->. bfl .° .^'. 3. T3. x. "s (B. X!. rt. CTi. •^. s:. ^ 0. ;§. ^. 4-> u ^•s g m ^ 8 '0 >> s t". ^ in ^ rt g. y. ^ m !-1 0). —309—. c3. T3 § ,,5 0 'p '<-! -U. ?T * 3. &;••»'!S/X^':X.S ;.A. a>. a. Ill. bE bfl ;>,. ,K^C!; R':li^:'a'itS^FtlS3. <u. f-<. +->. '0. & o? v^K. m. &. +-'. <D'. t. !-l. <3. c s? 0. p. "i. 6. 0 a. ;§. i^ g ?-1. ra. fe ft '0. ,J=i. E-i. 0. &s. v ^ 4->. K]. ?. (D e 1-A. ^^5 ,8.
(15) Yasushi Takenouchi. per day. According to Seller (1947), egg-laying of Otiorrhynchus sulcatus takes place throughout the night. In order to obtain detailed data on the egg-laying habits in Catapionus gracilicornis, direct observations were repeated at intervals of two hours throughout one week of maximum egg production, from May 29 until June 4, in 1957. Summarized oviposition records from this period are given in Table 9. Charts 4-5 show a close correlation between the time and egg-laying activities observed. in 36 beetles from May 29 to June 4, 1957. Although individual females deposited their eggs irrespective of whether it was day or night, the maximum frequency of egg-laying was recorded at noon. Generally speaking, the frequency of egg-laying by day was higher than that at night; the maximum average number of eggs per deposited beetle was recorded at 10 p.m. (Chart 4). The productivity increased with nightfall, reached a maximum at 12 noon, declined slowly thereafter and decreased rapidly with daybreak (light at 4 a.m.). Frequency of oviposidon in a single individual per day was observed in 36 females for a week. As given in Chart 6, the largest frequency was found twice and the smallest frequency five times. An egg-mass which is made in one egg-laying contains one to twenty-eight eggs (Charts 7-8). Through two years' observations, an egg-mass containing six eggs was most frequent in occurrence, with a frequency of 13.0 per cent (1,700 specimens). The egg-mass containing eight eggs ranked second in frequency. In general, females deposited 3-14 eggs per egg-laying. 5. Duration of the egg stage After having been laid the eggs were removed from the leaves (Fig. 7), and then placed on circular pieces of blotting paper in petri dishes (9 cm in diameter). The blotting paper was kept sufficiently moistened. From time to time, fungus grows over the eggs unless they are carefully watched and transferred to new containers as soon as mycelia begin to appear. Even if the eggs were kept on the water they easily hatched. The dry condition exerts an injurious effect on eggs, A well-developed larva bites off the chorion of the egg herself and emerges. During the incubation period the room temperature increased slowly from 10.7°C to approximately 23.4°C. Under this condition almost all eggs hatched in 11 to 15 days. The number in days of the incubation period of the eggs deposited in the laboratory was observed. The data are as shown in Table 10. Table 10.. Days No.. of larvae hatched. Length, in days, of the incubation period of eggs. 11. 12. 13. 14. 15. 16. 1. 7. 20. 50. 20. 0. The eggs were kept in incubation dishes which were placed in the laboratory. During the work the temperature in the room gradually increased from 19°C at the beginning of the experiment to 21CC at the time when the last eggs hatched on July 28. From 100 eggs used in this experiment, 98 larvae emerged. Hatching takes place through a narrow range of temperature. Both low and high temperatures obstruct the development of the eggs. At the very end of the oviposition period (October 1955), a small lot of eggs kept at room temperature (7°C-16°C) showed no hatchability. The eggs kept in incubator at 30°C also failed to hatch. Results of similar observations in —310—.
(16) Some Oecological Observations of Three Species of Curculionid Weevils. 1957 on 300 eggs are shown in Table 11. It has been shown that the eggs incubated at 16°-24°C are possessed of excellent hatchability. On this basis it seems apparent that the proper range of temperature for incubation is from 16°C to 24°C. It is certain that not only humidity but also temperature may be a factor affecting emergence. The incubation period become shorter with an increase of temperature. Table 11. Data regarding relationship between emergence of eggs and temperature. 7- 15°C. Temperature No, of eggs observed. 25-30°C. 16-24°G. 100. 100. 100. 0. 99-100. 0. % of no. of eggs hatched. Hatchability of eggs was examined fifteen days after the egg-laying. Table 12 shows. the hatchability of the eggs deposited by seven mother beetles : eggs deposited by every mother beetle show a high hatchability in 83 to 96 per cent without exception. Table 12. Hatchability of eggs observed in seven mother beetles. Serial no. of mother beetles. 1. 2. 3. 4. 5. 6. 7. Total. No. of eggs. 20. 49. 54. 51. 46. 26. 41. 285. No. of hatched eggs. 19. 44. 52. 47. 38. 21. 40. 261. °/o of hatched eggs. 95. 94. 96. 92. 83. 81. 98. 91. Under field conditions the hatchability seems not to be always good. Sixteen egg-masses were observed ,for their hatchability on leaves of the thistle in the bush at Sakae-machi, Sapporo, during a period from June 29 to July 11. The data are presented in Table 13. Table 13. Results of the observations of egg hatchability in field. 9 10 11 12 13 14 15 16. Serial no. of egg-masses. Hatched larvae + No. of developing esgs 0. +-++--++-++-++ 07002900 12000407. No. of dead eggs —. 812 66458 11 4 12 3321. % of hatched eggs 100. 10047- -2575 - - 52- - 0 - - 88. As shown in Table 13, two egg-masses showed high hatchability, and the remaining 14 masses contained developing eggs and dead eggs. In general, the eggs placed in the middle of the egg-mass may develop favourably, while those at the periphery died as a result of drying. Sometimes the eggs in field were attacked by ants. It should be mentioned that the injury by enemies is not always negligible. Furthermore, the eggs are largely influenced. by the weather, especially by temperature and humidity. It is therefore likely that the hatchability of eggs in the laboratory is significantly superior to that of eggs in the field. All the features presented above are sufficient to show that the animals under consideration propagate parthenogenetically. 6. The larva The full-grown larva (Fig, 22) averages 12 mm in length (range 10.5 to 14 mm). The. head is slightly convex and is brownish in color with lighter streaks on the sides and the 311.
(17) . Yasush i Takenouchi di te l lowi s sParselyr The body i eshy, more or less curved sh whi s ye r , legless ,a , an h i h f i f h l d d , t i l t 1 2 h h t d i h - n 〔 l e n c a r e s e a s 1 1 【 コ L uc o e Seg consss o p rated into Covere w t s ort ars ,w ,. ddl 1 ni e .. f- 、hal fthe body, The dorsal hal tudinal grooves running the length o f‐ ts by two longi l segmel i di l b h l ides lgi segn 〔 lents are bu ng; the ventralones are aat . 0n the s , a ove t e ongtu na separa‐ r l lseg北Ients exceptthe 2nd les are present on a d spi l lr ing grooves c o u n t r a s t n a ,the 3rd , and , A i f h h l 1 fthe t h . t 靴 tt t mos e en re sur ace o las the .2 t th( ) Those on t e st segmen are e arge .. ・. lowi inized yel ight ly chi t sh f .st dorsal hal shaped, l -segment i s covered by a narrow, bow- , plate , i 7 f the larval stage on o . Durat Just ha t ched outlarvae were removed from the incubation dishes and transferred to the les in and wooden boxes in which thist t l contained in several kinds of cans( fsoi surface o ts in every conにalner Thi ig ・ng process produced good resul s rear were growing (F . . . 23) f the thistle and develop favoura‐ l apart fro Larvae dig the ]m the root o 1 1 ・ selves into the soi fthe food plant they appear l whi h ti t the soi h bly ch has no root o . However , w en t ey ge n o b f f i h d d lin a few days an fthe soi e o unger again on the surface o . At 五rst , or a out ten days the larvae were found at depths ranging f ‐ rom 8 to 12clm at solne distance froln the , 1 f d t f h i t n s a l t t t o l d i t h i l o l d b o r o o s o r Th t e P m a e o i t a t we e s n g r v a e v o e s o e a , e y roo n g . th the roots of the host The wor ]ns eighteen days of age were found to be in contact wi i l h th d the leaves h d h i b ions Under 五eld condi t P1ants , t was o serve t at t e an【nas a c e on . lp ofthe ste]m then burrowed fthefood plant bythe he descended to the ground atthe root o i h k h t ti l th the rootofthe thistle h d l h o n a ct wi t n into t we r e e c N l he soi w c r a a a e p c e v ew y . ld gnawed 1 h 1 Th t 1 i d t i nty‐one days o b t d d r e we i e a v a beca ight rown n cooura on an e s or y. ]me1 l ing beneath the outer ‐ i f the roo 七 l 1 L lnet es found tunne deeper into the surface o , and were so ls ttakes place in the feeding cel l i layer o fthe cortex. M【 t ng was rarely observed because i o i 丘 d h l d i i h t i l t b t n i hi n h n r h a e e r r n c n a e f t t e e a o t n g e roo s n t e so , o o . 8 . Larval development A newly ha tched larva averages l.8-2.o m m in length,. t reaches to Five days later i. ltri t measures ap‐ d 25 t i d about 2,2 mm , seventeen days later to . m m, an twen y‐sx ays a e d i l t f h h l h b d t s some t a s old i l Th s i r e x 0 a va l 4 4 5 t o e y i e n y e o g y prox・nate y , - , lnm n eng ,. fthi l leasure- fthe body length in e×cess o b le growth o i i sl 13一14n s no v s [ lm, Because there i ・ l he ful tage are t -grown larvae ht that the animal hi i i h s at t ss lnent . ln general , , t s t oug 5 to 24 cm in the s。iL ・nature larvae were formed at the 7‐loth week at depths from 1 0 0 Dur ing the course o fthi s observation the atmosphere temLperature variedfrom 20 c to 24C. h h le fa fa snout beet l i l iv ( Vass 1914 )concluded that the larva ofthe al , passes t roug seven d f a larva after each 江Iolt ing t l d by measur hel instars ength o , lt was observe , as re▽ea e f おかαじカタγ彪Z”“s Zig拶sZZGZ Passed through 五ve instars of develop‐ eld larvae o also that the 6 r ion. lnent in the arst season, before hibernat ly part of october feeding dur ing the ear ,. ia the larvae were found to cease ln Russ. ing lowing spr Feeding was resu]med in the fol ,. ion in june and July. i lopment before pupat r deve and the larvae completed the. i l iv Vass. l d notstop feeding in the oswego area unti i 1914 ty of the larvae di t j or ( )repor ed that the ma i l d l b h t t i i h h r ma u r a v a 3 4 f t t late i y e overw n erng gru s reac e ] [ lost o n Decelnber, 19 , and t at ・ According to Jab 1909 lonowski( )the ion for the winter ing condi t before going into a rest , larva o f the a1falfa snout beet1e in Hungary feed throughout the winter in areas where the i l does not freeze. The above observations indicate thatthe stage of developmentreached so f the feeding period during ion i th the length o by the larvae before hibernat s correlated wi. 一312一.
(18) . I i r ionid 頓 S i Some 。eco 1 i eevi 1 obServat es of curcul ogica onS of Three Spec l ing the fal i l temperature favours prolonged feeding dur 1arval stage and that the so , nths .. f The present author observed the number of larvalinstars based on the measureTnent o l f the arvae occur- [ lent o dth ofthe head capsule. ln order to determine the developn wi f h d h i t c a h h d f d ヒ e Psule of grouPs o i i l h t e a t h w ma e o n t s e o t me r n I e n wa asu e e so , e g n h l f 仁 t e 4 h om 1 t s r e s u i i l e r T b i h d A larvae at 7 day interval s s seen n t e ata g ven n a e , . Z Z Z 一 α じ γ i cの s αかの2鋭 g ix to seven larvalinstars n cate thatthere ares Ve measurementsindi fl ike ly that since therei 期s s a considerable Variation in the development o arvae sl , エti fthe head capsu1e on1y dth o d 1i b l th i i f h every stage , t e Countng o arva nstars ase upon e w not always proper . Tabl e 14.. Z Z I C0γれ槌 c l o児錫s 9γα idth of head caPsu 斬7 ′ ein larvaeof CαtαのZ ) Sapporo l iouss in var tagesof deve opmen七( , ,1957. 8 23 / 8 / 16 9 / 8 / 8 / 7 2 7 26 / / 12 7 19 / / 7 / 6 5 14 6 28 / 6 / 6 21 7 9th loth llth 8th 7th th 6th 5 3rd 4th lst 2nd t jus i depo ted week week week week week week week week week Week week s l l imeters ) d Wi th of head capsule (mi. dth M [ imum wi ax Minimum wi dth Totallarvae. 2 00 . 00 2 . 1 50 .. 80 1 . 2 00 . 2 03 .. 2 00 . 2 00 . 2 03 .. 00 2 . 00 2 . 90 1 。. 1 46 . 1 04 .. 1 75 ‐ 1 50 .. 2 00 . 1 90 .. 00 2 . 00 2 .. 90 1 . 2 00 .. 0 44 . 0 46 .. 0 62 . 0 62 .. 0 62 . 0 6 .0. 68 0 . 0 94 .. 1 10 . 1 20 .. 0 36 . 0 38 .. 0 46 , 0 48 .. 16 1 . 1 04 .. 0 46 . 0 44 .. 0 60 . 0 60 . 0 60 .. 10 1 . 0 86 .. 0 38 , 0 38 ,. 0 42 . 0 4 .2 0 42 .. 1 10 . 0 70 .. 1 10 . 10 1 .. 0 36 , 0 36 .. 0 46 . 0 48 .. 0 62 .. 0 62 .. 0 86 . 0 78 .. 00 1 . 04 1 .. 04 1 . LO0. 0 38 . 0 38 ,. 0 46 . 0 46 .. 0 42 , 0 42 ,. 0 61 .. 0 76 .. 0 62 .. 80 0 .. 00 1 . 00 1 .. 1 10 . 1 00 .. o 38 . o 36 .. 0 43 . 0 44 ,. 0 62 . 0 42 .. 0 64 . 0 58 ,. 1 10 . 0 68 .. 20 1 . 00 1 .. 46 374 0 o . . lo lo. M [ ean value. 20 1 . 1 16 。. 1 10 .. 0 38 ‐ 38 0 .. 2 00 . 2 00 . 2 10 . 2 00 . 90 1 . 0 2 .0. 75 1 . 75 1 .. 90 1 . 2 0コ ,. 2 00 . 2 12 .. 00 2 . 00 2 .. 00 2 . 1 50 .. 00 2 , 90 1 .. 2 10 . 04 2 .. 2 00 . 00 2 .. 1 75 .. 00 2 ,. 00 2 .. 2 00 .. 2 00 , 2 0 .O. 46 1 . 00 1 .. 00 2 . 1 50 .. 2 00 . 1 80 .. 2 12 . 00 2 .. 2 00 . 1 90 .. 2 1O . 1 9O .. 12 674 1 858 0 0 609 0 86 0 ‐ . . . .4 lo lo lo lo lo. 1 75 . lo. 95 1 , lo. 98 026 1 2 . . o o l l. 2 20 . lo. 0 42 . 0 48 .. 0 58 , 0 64 .. 1 10 .. 00 2 . 00 2 .. ion o Pupat fthi , The , 1957 s sPecies was 五rst observed in the laboratory on August 16 iner Whi ing Conta ch contained 20 grubs ion エ laboratory exatnina仁 ー ] Lade for l。 weeks in a rear ‐ hi l h em‐ d f a s i i l i d d l h pupae 957 revea ed t attwo n v uas were oun hatched on June 7 1 , w et er. ,. ,. th day Then, three o f that re. r 工 lainder were found as pupae on the 84 hi h vere hatched on August l9 and kept in another i id l 12th week) ( , Twenty ind v uas w c v found i s b n thi 33th day(19th weeks’ container were examined on the 1 , ut no pupae were i l h t i l l a h t a s o t n e t n c e d ly lnature larva ceased feeding and for・ne a nea ear e case, Ful ’ f t l l℃ idf a er 9℃ to Fig depth of 20cm ( ) , The temperature of the laboratory vare rom ,24 t ion made i i n l956 ・ninary examinat ddl the mi 、200larvaejus e part of october , ln the prel ‐ 0 3 i 0 i 2 j un e 3 5 n o 2 5 × n s z e b cm d × cm i cm i l n a woo en ox, , , o emerged were removed to s i le [ longst 86larvae observed, wh on exannination made on August 30,there were no pupae an th of24Cnl ・ ] Lature larvae and 4 pupae atthe dep i on made on october 6 showed 8 1 examinat They pupate in l t nt l t in the container . The bodies of nnature arvae are par y ranspare , l l Fig ) ly formed earthen ce s ( neat . .25 ainder were larvae,. 3- -31.
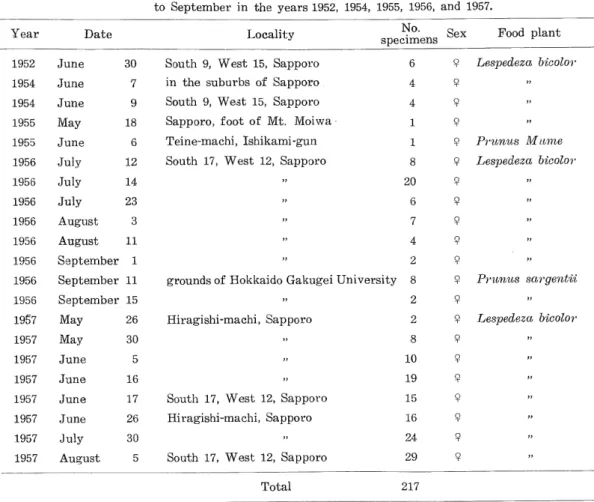
(19) Yasushi Takenouchi 9. The pupa. The pupa (Fig. 26) is approximately 11 mm in length (range 10 to 13 mm), and it is yellowish white in colour; the eyes are black. There are 14 spines on the head — 4 in a semicircle on the vertex, 2 on the front between the eyes, 2 near the apex of the beak, and 3 on each side near the apex of the beak above and to the out side of the basal attachment of the epistoma. There are 16 symmetrically arranged spines on the dorsal surface of the prothorax. On the abdominal segments, a single row of spines is present on the dorsal fold; the spines on the ventral surface are few and inconspicuous, except on the posterior segments. Two outgrowths of the last abdominal segment extend backward and diverge slightly. The tip of each of these processes is dark brown and sharply pointed.. 10. Length of the pupal stage In order to determine the length of the pupal stage, examination was made of the pupae in the soil in several containers throughout the season, but none of the observations yielded information due to a high percentage of mortality caused by fungus and some other artificial causes. From numerous newly hatched grubs placed in an oil-can on June 13, 1956, two pupae were metamorphosed into adults by September 22. Cultures continuously carried out since 1957 are now being kept in several containers to observe the details of the pupal stage. The examination made on January 31, 1958, showed that there were fully mature larvae at hibernation. It is doubtless a fact that soil temperature exercises a significant influence on the rate of pupation and metamorphosis. C. About Pseudocneorhinus bifasciatus Roelofs. 1. Activities of the adult Observations in Sapporo The beetles were found in several places in Sapporo as shown in Table 15. In late spring the beetles climb upward through the soil, making tunnels to the surface. In 1955, beetles were first recorded on May 15. In 1957, some field observations were carried out by the author around the habitats of the beetle in early spring. The first adults were found in one of the places in Sapporo on May 25, whilst in another place newly emerged adults were first observed on June 17. It is most probable that weather conditions and soil and air temperature largely influence the time of emergence of the beetle. Numerous beetles of this species are observed from the middle part of June until early September. It was during this period of activity that most of the feeding injury to various plants was observed. The occurrence of beetles is restricted to several small areas in Sapporo. A total of 217 weevils were collected in several parts of Sapporo during the past six years. Table 15 lists data on beetles so far collected. Examination by dissection was performed on one hundred and thirty-nine beetles of this species out of 217 specimens collected; it was clearly found by this examination that all specimens under study were female. The remaining seventy-eight specimens were observed in captivity; they were females, since without exception they laid eggs. It is then evident that there has been found no male amongst the specimens collected by the author in the past five years. Although Lespedeza bicolor is the preferred host plant of this species, the beetles feed upon a large variety of other species widely distributed among the plant families. The plants upon which the beetles feed are Malva sylvestus, Pnmus sargentii Rehder, Prwws Mume Sieb. et Zucc., Rumex obiusifolitis, Rumex longifolius, Rtimex crispus, Actinidia callosa Lindl. var. argttta Makino, Artemisia montana (Nakai) Pampan, and M.orus bombicis 314—.
(20) Some Oecological Observations of Three Species of Curculionid Weevils Table 15. Collection records from several regions of Sapporo, from May. to September in the years 1952, 1954, 1955, 1956, and 1957. Date. Year. Locality. No.. Sex. ?. specimens. Food plant. 1952. June. 30. South 9, West 15, Sapporo. 6. 1954. June. 7. in the suburbs of Sapporo. 4. ?. 1954. June. 9. South 9, West 15, Sapporo. 4. ?. 1955. May. 18. Sapporo, foot of M±. JVtoiwa. 1. 9. 1955. June. Teine-machi, Ishikami-gun. 1. ?. Prztnus M nine. 1956. July. 12. 8. ?. Lespedeza bicolor. 1956. July. 14. 20. 9. 1956. July. 23. 6. ?. 1956. August. 3. 7. ?. 1956. August. 11. 4. ?. 1956. September. 1. 2. ?. 1956. September 11. 8 grounds of Hokkaido Gakugei University 8. ?. 1956. September 15. 2. ?. 1957. May. 26. 2. 9. 1957. May. 30. 8. ?. 1957. June. 5. 10. ?. 1957. June. 16. 19. 9. 1957. June. 17. South 17, West 12, Sapporo. 15. 9. 1957. June. 26. Hiragishi-machi, Sapporo. 16. ?. 1957. July. 30. 24. ?. 1957. August. South 17, West 12, Sapporo. 29. ?. 6. 5. South 17, West 12, Sapporo. Hiragishi-machi, Sapporo. Total. Lespedeza bicolor. Prunus sargentu. Lespedeza bicolor. 217. Koidz. With the exception of 11 individuals which were obtained on leaves of cherry-trees (Prunus sargentii) and a plum-tree (Prtmus M.ume), specimens were collected exclusively from leaves and branches of Lespedzza bicolor in every locality. In 1957, the first adult was obtained on May 25 in Hiragishi-machi, while at South 17, West 12, Sapporo, adults were. first collected on June 17. In the previous year (1956) beetles were obtained on July 12 in the latter place. It seems likely that the beetles are living in a small group in any given locality. The last living adult was obtained on September 15. The animals live on the host plants on the naked soil without any grass. During a feeding period from May to September the beetles were observed on the foliage of the host plants. When disturbed by enemies,. the feeding adults immediately feigned death and dropped to the ground. Bright sunlight is harmful to the adults, as was found in Caiapionus gracilicornis and Brachyrhinus ligustici (Vassiliev 1914; Palm 1935). In general the beetles live among the foliage or on the leaves of the host plants in the shade. It was found that the most favourable air temperature for this species was from 20°C to approximately 30°C. When the beetles were handled and placed together in a collecting bottle, they instantly became active though they were usually inactive in their hiding places. A comparatively long duration of time seems to be needed to deposit eggs. Numerous adults of various ages were dissected before the last part of June, but the author has failed to find eggs in most beetles, furthermore their ovaries were —315.
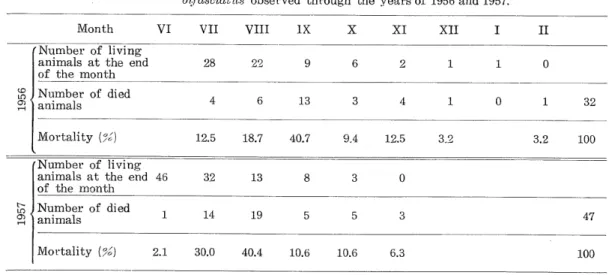
(21) Yasushi Takenouchi too small to detect. A total of 109 beetles were used in the rearing experiment carried out over the past two years. Thirty specimens from among them were caged in 1956, and the remaining animals were reared in 1957. The beetles were placed separately in petri dishes with a little sifted soil and a piece 01 a leaf of Rtimex obtttsifolius, or some foliage of Lespsdzza bicolor. (Fig. 29). The following information describes the adult activity which was observed in the laboratory. The beetles fed on the leaves of the host plant during their active period from emergence to the middle of September. In the active period the beetles died in a few days when they were caged in a container without food, while when the active period was once over, the animals survived for a comparatively long period without food. Table 16 shows the mortality of 79 adult females observed within the past two years. Table 16. Comparative records of mortality of Psetidocneorhinus. bifasciatus observed through the years of 1956 and 1957. Month. IX. x. XI. 22. 9. 6. 4. 6. 13. 12.5. 18.7. 46. 32. 1 2.1. VI. Number of living animals at the end of the month <c. m. oi T-l. Number of died animals. Mortality {%} Number of living. animals at the end. of the month tin. ai. Number of died animals. Mortality {%}. VII. VIII. 28. XII. I. II. 2. 1. 1. 0. 3. 4. 1. 0. 1. 32. 40.7. 9.4. 12.5. 3.2. 3.2. 100. 13. 8. 3. 0. 14. 19. 5. 5. 3. 47. 30.0. 40.4. 10.6. 10.6. 6.3. 100. The seasonal maxima in mortality were observed in September of 1956, and in August of 1957. Generally, the beetles cease from oviposidon and feeding acfcivity approximately three or four days before their death. Dry condition is harmful for animals. When the soil in the container dried up, the beetles stopped their oviposition and feeding, and became inactive. In general, oviposition is observed during the period from. late June to the last part of August. During the oviposition period the beetles feed on leaves of the food plants. In the early part of September, small numbers of beetles were caught on leaves of Prunus sargentii Rehder in the grounds of Hokkaido Gakugei University. Insectary observations indicated a brief period of activity and feeding after oviposition was completed. 2. Parthenogenetic reproduction So far as the author is aware, no report has so far been made by any worker concerning the details of reproduction of this beetle {Pse'^docn 7orhinus bifascici.tus).. Dissections of 139 adults obtained in Sapporo and vicinity have revealed that all of the individuals under observation were females. As noted above, 79 individuals were collected just after their emergence and caged separately in small petri dishes with soil and leaves of food plants. All of these beetles deposited eggs under captivity ; most of the eggs that were deposited developed normally. On the basis of these experimental results there is no doubt — 316 -.
(22) Some Oecological Observations of Three Species of Curculionid Weevils but that this species reproduces by parthenogenesis. 3. The egg. The egg (Fig. 36) measures approximately 0.87 mm (from 0.85 to 0.92 mm) in length by 0.43 mm (from 0.41 to 0.44 mm) in width. It is elliptical in form, and weighs 0.17 mg on the average. Except for the fact that the egg of this species is slightly larger than that of Catapiomis gracilicornis, it is difficult to distinguish the eggs of the two species. The original color of the freshly deposited egg is shiny milky white; the color changes to dull one to three days later, and then to light brown as further development proceeds. Shortly before hatching, the tips of the mandibles of the larva are visible through the egg covering. 4. Oviposition records In order to obtain some information on oviposition, a preliminary examination was made. with thirty beetles in 1956. Further investigation was carried out with 65 individuals in 1957. The beetles were placed separately in small petri dishes with about 5 mm of sifted moist soil. Throughout the entire work, fresh leaves of Rumex obtusifolius were kept in each container. The eggs are usually deposited on the leaves of host plants through ovipositor in a manner similar to that described above in Catapionus gracilicornis (Figs. 3036). A female laid 4 to 10 eggs within 10-20 minutes, The freshly emerged beetles need about 30 days before the commencement of oviposition (Table 17). Table 17. Length of the period (days) from emergence to the beginning of ovi position in Pseudocneorhinzis bifasoiatus, from the data based on 12 individuals collected on June 3, 1957. Beetle no.. The data on which oviposition. begins. No. 1. No. 2. No. 3. No. 4 No. 5. No. 6. No. 7. No. 8. No. 9 No. 10 No. 11 No, 12 Total Mean. June. June. June. June. July 1. July 2. July. July 5. July 6. 28. 29. 30. 30. 3. July 6. July 6. July 8. Preparative. period. F<??°?v4)aS25 26 27 27 28 29 30 32 33 33 33 35 258 d2a9y^ for tion. It seems apparent that a comparatively long time ranging from 25 to 35 days with an average of 29,8 is required before the egg-laying begins (Table 17), For purpose of obtaining oviposition records, the leaves in the rearing container were examined daily for the presence of eggs. The data thus obtained during the entire egg-. laying period in 1957 are summarized in Table 18. Beetles Nos. 5 and 23 escaped before oviposition began. No eggs were deposited by beetles Nos. 6, 9, 26, 28, 31, 34, 35, 36, 37, 39, 40, 42, 43, 44, 46, 47, 48, 49, 50, 51, 52 and 53 during the entire experiment. Examinations by dissection made on October 16 indicated that all of them were females, and further a comparatively large parasitic worm which is regarded as a larva of a certain arthropod was found in every non-depositing female. There is no doubt but that the degeneration of gonads due to the parasitic worms caused the beetles to fail in their egg formation. Daily records during the whole egg-laying period, the number of beetles alive, the number ovipositing, and the average number of eggs per day per living beetle, are shown in Chart 9. The greatest number of eggs were deposited during the first part of July though egglaying under insectary conditions showed a comparatively long duration from June 8 to — 317 —.
(23) Yasushi Takenouchi Table 18. Oviposition records for Pseitdocneorhimcs bifasciatzis at Sapporo, during May, June, July, August and September 1957. The data are based upon. daily records from 55 beetles and are summarized for the approximately 10-day period. June June June July July July Aug. Aug. Aug. Sept, Sept. Sept. Oct.. Beetle No.. Date 1 11 21 1 11 21 1 11 21 1 11 21 1. Col- to to to to to to to to to to to to to lected June June June July July July Aug. Aug. Aug. Sept. Sept. Sept. Oct.. 10 20 30 10 20 31 10 20 31 10 20 30 10 Number of eggs deposited. 1. May 26 0. 4. 21. 3 4. 0 May 30 0 1. 8. 0 0 0 0 0. 0 11 2. 0 0. 0 0. 5 0. 35. 35. 2. 5 6 7. Escaped. 9 10 11 12. 13. 14. 15 16 17 18. June. 0 0 0 0 0 0. 0. 6 14. 0 0 0. 0 7 23. 39. 30. 4 14 16. 32 45. 0 0. 10. 8. 7. 2 4. 6 6. 10. 34 19 22 0. 0. 7. 0. 54 5. 49. 60 53 23 48. 0. 19. 0. 0 0. 13 17 0. 21 22. June 16 0. 0 0. 23. 0 37. 0 0 0 0 0 0 0 0 0 0 0 0 0. 15 16. 40. 20 23. 24 25 26 27. 28. 29. 30. 31 32 33 34 35. 36. 0. Escaped 0-. 0. 0 0 0 0 0. 0 0 0 0 0 0 0 0. 37 38 39 40. 0 June 26 0. 41. 0. 42 43. 44. 45 46 47 48 49. 50 51 52 53 54 55. 0 0 0 0 0. 0 0 0 0 0. 0 0. 0 0. 0. 0 0 0 0 0 0 0. 0 0 0. 0. 0. 0. 0. 0 0 0. 0. 0. 0 12 0 0. 35 4. 48 0. 17 0. 36. 0 4 0 0 17. 0 20 0 0 4. 36 0 0 52. 44. 36. 21. 23 0. 16. 0. 51 31 51 24 32. 0. 33 0 14. 34 58 21. 0 17 0. 15. 0. 0. 0. 5. 35. 0 0 0 0 0. 12. 0 0 0 0. 0 0. 0. 0 0 0 0 0 0. 0. 0. 0. 0 0 0 0 0 0 0. 0 0 5. 0 0 0 0 0 0 0 0 0 0. 19 60. 0 0 0 0 17 0 0 13 0. 0 0. 16. 0. 0 27 23 68. 20 25. 0. 38. 11 0 30 55. 0 0. 14. 0 37 5. 0 11. 66 0 0. 0. 0. 0. 0. 25 26. 57 24. 24 49. 16. 0 17 41. 21 29. 18 31. 52. 17 16 24. 0. 8 0 28 18 0 0 0 9 0. 38. 0 0 0 0 0. 29. 0 0 2. 47. 0 0 0 0. 14 0 0. 15 0 0. 34. 20. 0 0 0 20. 0 0 0. 22. 0. 0. 0 0 0 0. 0 0 0 0 0. 0. 0. 0. 0 0. 0. 0 0 0. 0. 0 0 0 0. 0. 0. 1 0. 2 26. 8. 0. 0 0 8 0 0.
(24) Some Oecological Observations of Three Species of Curculionid Weevils. '%^^^^. ,-'>'<»»^^,,,<^,,,^'!-*-» '-"^. '~^"^~^i^'\^^ •"-.",:, Mii.r ?K> rr:np. l->k', « .'Oli'r BI-i-Tt-iu -f(P l.t'ifi) E<<ii;i • W.. HO (1%;; pt'S ('•A*' PES Bi.S.U.E. I1!1'. IN) ili/Tf FTTN ;. •^ ^-.^. iwy 'I. U^J^-^r'^--^^^ i. I. '. '.. ^^. ^'. I?. '. '. -*<'•. JUNE. '. '. !. •*'. hi. M'W. .. ~'. ^Y*^. .?<!. "W " HiH/h. -V.VM. (*^i*«*<-+^.*.''». .>). WWW. 10. '. 30. <1. .*. .••»..i^. !(•. ;0. ••SSV\m?».. <. ». 10 10. W. .*1. Chart 9. Graphs showing a close correlation between temperature and egg-laying activities in 30 specimens of Pseudocneorhmus bifasciatzis during their oviposition period. The weevils ramained alive for about one month after egg-laying.. 1-IC ij-^9. AUW. Juiy. SEPTEMBER. Chart 10. A summarized record on oviposition data from Pseudocneorhim<,s bifasciatus given in Table 18. The curve shows the percentage of total number of eggs deposited in ten-day intervals, in an entire. period of oviposition.. -319—. f. •X-SV6F-K ^ . ».'.
(25) Yasushi Takenouchi. September 29 (Charts 9, 10). The data presented in Chart 9 show that air temperatur& affected the egg-laying activity of the beetles. During the season in 1957, a total of 3,975 eggs were deposited by 31 females, ranging. from 1 to 478 eggs per beetle with an average of 128.2 (Table 19). The smallest number laid by any one beetle in one day was 1, whilst the greatest number was 20, with a daily average of 5.6 eggs per living beetle. During a period of egg-laying, the adults live actively. and feed up to the time of their death (Chart 9 and Table 19). Table 19. Summary of the activities of 31 egg-laying Psezidocneorhimis bifasciatus at Sapporo, during May, June, July, August, and. September, 1957 Minimum. AIaximum. Average. Number of eggs/day/living beetle. 20. 0. 5.6. Total number of eggs per beetle. 478. 1. 128.2. Length of ovi position period (days). 107. 1. 43.9. Days from the onset of egg-laying. 146. 8. 57.1. to its death. Individual females deposited their eggs quite irregularly during their oviposition period. In most cases (167 out of 310 cases), adults deposited some eggs for a day, and then rested for several days before resuming their egg-laying activities. The data are presented in Table 20. Table 20. Correlation between number of days in which egg'-laying was. continued and their frequencies in the oviposition period (1957) Days in which e;gg-laying was continued No. of cases. 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 15. Total. 167. 73. 29. 13. 10. 8. 3. 1. 1. 1. 1. 1. 310. In 30 beetles under captivity from July 20 to October 27, 1956, a total of 5,453 eggs were laid by 28 females. The number of eggs laid per individual ranged from 8 to 474 eggs with an average of 194.75 (Table 21). Two non-depositing animals were proved as females in dissection. The largest number deposited by any one beetle in one day was 30, with a daily Table 21. Summary of the activities of 28 egg-laying Psezt.docneorhimis bifasciatz(,s at Sapporo, during July, August, September, and November, 1956. Maximum. Minimum. Average. 30. 0. Total number of eggs laid per beetle. 473. 8. 194.75. No. of days of deposition per beetle. 158. 10. 51.3. No. of eggs/day/living beetle. — 320 —. 3.8.
図

+7



関連したドキュメント
To measure vulnerability we have some parameters that are toughness, binding number, vertex integrity, and scattering number [5].. The problem “given a graph G , decide whether
Typically, the value of the Coarse Shutter Width Total registers is limited to the number of rows per frame (which includes vertical blanking rows), such that the frame rate is
As long as pest infestation continues, make repeat applications a minimum of 7 days apart but do not apply more than 16.0 ounces (0.3 pounds active ingredient) per acre per
In recent years, its abuse has been spreading, especially among the youth in their 20's, and the number of arrests for cannabis abuse in 2004 reached a record high of 2,209 (108.7%
Amount of Remuneration, etc. The Company does not pay to Directors who concurrently serve as Executive Officer the remuneration paid to Directors. Therefore, “Number of Persons”
Apply in a minimum of 2 gallons of water (or a total carrier volume) per acre by air or a minimum of 20 gallons of water per acre by ground. For improved efficacy, light
Pre-Harvest Interval (PHI) from planting application: 3 days (leaves); 125 days (corms) Maximum amount allowed per crop season: 24.0 fluid ounces/Acre (0.38 lb AI/Acre) Maximum
frame_length_lines determines the number of row periods (T ROW ) per frame. This includes both the active and blanking rows. The minimum vertical blanking value is defined by the
