博 士 学 位 論 文
タンパク質の変性中間体および凝集体形成メカニズムの
熱力学的研究
Thermodynamic study of transition mechanism of
intermediate and aggregate state of protein
近畿大学大学院
総合理工学研究科理学専攻
博 士 学 位 論 文
タンパク質の変性中間体および凝集体形成メカニズムの
熱力学的研究
Thermodynamic study of transition mechanism of
intermediate and aggregate state of protein
平成 30 年 1 月 9 日
近畿大学大学院
総合理工学研究科理学専攻
目次 頁 序論 1 1 章 ミオグロビンのソルビトール誘起モルテングロビュール状態における 熱力学的及び速度論的解析 1.1 序論 5 1.2 実験 1.2.1 試料 7 1.3 結果および考察 1.3.1 ソルビトール存在下における構造解析 8 1.3.2 MG 状態の熱力学的解析 9 1.3.3 MG 状態の熱安定性(CD 測定) 12 1.3.4 MG 状態の熱安定性(DSC 測定) 14 1.3.5 ソルビトールの添加による MG 状態への転移機構 16 1.3.6 MG 状態の速度論的特性 18 1.4 1 章まとめ 21 2 章 1,4-ジオキサン添加によるシトクロム c の線維状凝集体の熱力学的研究 2.1 序論 22 2.2 実験 2.2.1 試料 23 2.3 結果および考察 2.3.1 シトクロム c の DSC 測定 24 2.3.2 シトクロム c の CD 測定 MG 状態の熱力学的解析 26 2.3.3 シトクロム c の状態図 28
2.4 2 章まとめ 29 3 章 1,4-ジオキサン添加によるミオグロビンの凝集体形成の熱力学的および 速度論的研究 3.1 序論 30 3.2 実験 3.2.1 試料 30 3.3 結果および考察 3.3.1 ミオグロビンのヘムの脱離と CD 測定 31 3.3.2 ミオグロビンの DSC 測定 33 3.3.3 静置時間依存性 37 3.3.4 凝集における活性化体積 40 3.4 3 章まとめ 42 総論 43 補足:実験装置・測定 (A) 円二色性測定 45 (B) 蛍光測定 45 (C) 走査型電子顕微鏡測定 46 (D) 動的光散乱光測定 46 (E) 示差走査熱量測定 46 (F) 粘度測定 47 (G) 密度測定 47 (H) 高圧密度測定 47
参考文献
序論 タンパク質は疎水性相互作用、水素結合、静電的相互作用、S-S 結合などのエンタ ルピー的要素と構造や溶媒和などのエントロピー的要素により、熱力学的にわずかな 安定性によって二次構造や三次構造を有した規則正しい立体構造を形成し、機能を発 現している。このような状態は“天然状態(Native:N 状態)”と呼ばれ、構造的に折 りたたまれていることから“折りたたまれた状態(Folded state)”と呼ばれることも ある。N 状態のわずかな安定性は生体内で効率的に構造変化や機能発現を実現する上 で重要な要素であるが、一方で、その小さな安定性のために温度や塩濃度、有機溶媒 や pH などの外的要因により容易に変性し、変性状態(Denatured:D 状態)へと転移 する。D 状態は変性の要因によって様々であり、尿素などの変性剤による変性状態(変 性剤変性)、pH による変性状態(酸変性状態・アルカリ変性状態)、温度による変性 状態(熱変性状態・低温変性状態)、圧力による変性状態(圧変性状態)、有機溶媒に よる変性状態(有機溶媒変性)など、名称だけでなく構造の崩壊の程度も一様ではな い[1-5]。これは構造崩壊のメカニズムが外的要因に応じて異なることを表している。 一般に D 状態は N 状態と比べて折りたたまれた構造が崩れており、構造的には“折 りたたまれていない状態(Unfolded state)”と呼ばれることもある。強アルカリや高 温などの極端な条件以外の変性においては、非共有結合の切断によるアンフォールデ ィングであるため、外的要因を取り除く(例えば酸性条件を中性に戻す)ことにより、 タンパク質は再び N 状態に転移(フォールディング)する。このような可逆的な構造 転移をもとに、タンパク質の安定性は N 状態と D 状態のギブズエネルギー差(ΔGo) として熱力学的(平衡論的)に表現される。 ΔGo=−RT ln K (1) K=[N]/[D] (2) ここで K は平衡定数、[N]と[D]は各状態の濃度、R は気体定数(8.314 J K-1 mol-1)、T は絶対温度[K]である。 タンパク質にはこのような N 状態と D 状態とは別に、速度論・平衡論的に中間的
な状態である変性中間体が存在する[6-9]。タンパク質の変性中間体は、アンフォール ドした構造から N 状態にフォールディングする過程に平衡論・速度論的に存在する準 安定な構造であると考えられており、立体構造の形成メカニズムを理解する上でこの ような中間体の構造や安定性を明らかにすることが重要である。変性中間体の中で、 特に二次構造が N 状態に近く三次構造が崩壊した状態をモルテングロビュール状態 (Molten Globule:MG 状態)と呼び、多くの研究者によってその構造的・熱力学的な 特性が研究されている[10-12]。 また、タンパク質の基本形態の一つとして、近年、N 状態、D 状態、MG 状態の他 に凝集体構造が考えられている[13-15]。タンパク質の凝集構造には、タンパク質が無 秩序に集まったアモルファス凝集体(Amorphous Aggregation:AA)や規則正しい凝 集体(Ordered Aggregation:OA)がある。OA 状態にはタンパク病の原因となるアミ ロイド(Amyloid)[16]と呼ばれる線維状凝集体(Fibrous Aggregation: FA)があり、発 病のメカニズム解明や予防の観点からも、その形成メカニズムを解明することが望ま れている。 タンパク質は分子量が大きいため非常に複雑な相互作用のバランスでその構造が 形成されている。このような“規則正しく”かつ“複雑”という二面性が、タンパク 質の構造形成メカニズムを解明する上で重要なポイントとなる。N 状態の構造は結晶 構造解析や NMR などを用いて明らかにすることができるが、非天然状態については その不均一性のため、構造決定をすることが難しい。このようなタンパク質の非天然 状態への構造転移を観測する上で、巨視的な情報を与える熱測定を含む熱力学的なア プローチは有効な手段となる。複雑な相互作用で構成されるタンパク質も熱力学的な 法則に従っており、タンパク質の状態転移に伴う熱力学的測定から得られる熱力学量 の基礎的データの蓄積は、タンパク質の構造や安定性、ひいては機能発現のコントロ ールを実現する上で、重要な研究課題である。 以上の観点から、本研究では非天然状態である変性中間体、変性状態、凝集体の形 成機構に関し、以下の 2 点についてその解明を目指した。詳細な背景や目的は各章に
おいて記述する。
<目的1> 糖によって誘起される変性中間体の構造と形成メカニズムの解明(1 章) 主論文1 J. Therm. Anal. Calorim. (2016) 123, 1861-1869.
<目的2> 有機溶媒により誘導される凝集体形成メカニズムの解明(2, 3 章) 主論文 2 Thermochimica Acta. (2018) 659, 8-12.
主論文 3 Netsu Sokutei. (2018), in press.
糖アルコールや有機溶媒のような共溶媒の添加は、温度や圧力と比べると実験的に 容易であり、水溶液中のタンパク質に摂動を与える上で有効な手段である。本論文で は、共溶媒添加によるシンプルな外的摂動によって非天然状態を誘起し、下記の手法 を用いてその構造や熱力学的特性を明らかにした。 二次構造含有量の決定・・・円二色性分散計 J-820 W(JASCO, Japan) フォールディングの推定・・・蛍光測定装置 F-4500(HITACHI, Japan) 熱変性・凝集化熱力学量の決定・・・断熱型示差走査熱量計 VP-DSC(Malvern) ・・・DSC VII evo(SETARAM, France)
粒子径の決定・・・自動マイクロ粘度計 AMVn(Anton Paar, Austria) ・・・動的光散乱光度計 Zetasizer Nano ZSP(Malvern) 凝集構造の決定・・・走査型電子顕微鏡 S-4800(HITACHI, Japan) 部分比容の決定・・・振動型精密密度計 DMA4500(Anton Paar, Austria)
・・・高圧対応精密密度計 DMA512(Anton Paar, Austria)
本研究で用いたタンパク試料は、ウマ心筋由来のミオグロビン(17.8 kDa)とウシ 心臓由来のシトクロム c(12.3 kDa)を用いた。それぞれ、ヘム基を有する典型的な 単量体のタンパク質であり、ミオグロビンは疎水性相互作用などの非共有結合により
ヘム基と結合し、シトクロム c はシステインを介したジスルフィド結合による共有結 合でヘム基が結合している。そのため、ミオグロビンは変性にともないヘム基の脱離 が生じるが、シトクロム c では脱離しないなど、構造のコアとしてのヘム基の役割を 比較することができる。また、ともにα ヘリックスを多くもつタンパク質であり、二 次構造の変化を測定するのにおいても適している。さらに、シトクロム c はある条件 下で MG 状態や線維状凝集体に転移する報告[17,18]があり、ミオグロビンについても 凝集体形成に関する報告[19,20]なども多数あるため、それぞれ比較するための適切な モデルタンパク質である。
1 章 ミオグロビンのソルビトール誘起モルテングロビュール状態における 熱力学的及び速度論的解析 1.1 序論 タンパク質の立体構造は、熱力学的には水素結合、疎水性相互作用、静電的相互作 用とファンデルワールス力などのエンタルピー的要因と、構造や溶媒和などに起因す るエントロピー的要因により形成されている。球状タンパク質が水中においてフォー ルディング構造(天然状態)を形成する際には、一般に多くの疎水性アミノ酸は水を 避けてタンパク質の内側に存在し、逆に多くの親水性アミノ酸は水に露出している。 このような疎水性物質が水との接触を避けて互いに集まる作用は“疎水性相互作用” と呼ばれ、タンパク質が立体構造を形成する上で重要な役割を果たしている。温度や 圧力、pH、変性剤、塩、有機溶媒などの外的要因によってタンパク質が変性すると、 フォールディング構造が崩壊し、多くの疎水性アミノ酸は溶媒中に露出する[1-5]。タ ンパク質が変性する原因は外的要因によって異なるが、いずれにおいても言えること は、天然状態よりも変性状態のほうが自由エネルギー的に安定であるということであ る。天然状態と変性状態の自由エネルギー変化は両状態のエンタルピー変化とエント ロピー変化の大小関係によって決定される。そのため、変性の原因を一つに決定する ことは困難であり、エンタルピーやエントロピーの役割や影響の大きさを個別に明ら かにするためには、構造変化のための適切な外的要因を選ぶ必要がある。塩の添加は 主に静電的相互作用に影響を与え、有機溶媒の添加は水素結合、静電的相互作用、疎 水性相互作用に影響を与えることが知られている。また、一般には糖の添加はタンパ ク質を安定下させることが知られているが、我々は過去の研究において、シクロデキ ストリンのような環状オリゴ糖がタンパク質の安定性を低下させることを見出して いる[21,22]。これは、変性によって露出した疎水性側鎖にシクロデキストリンが選択 的に包接することにより変性状態が安定化し、相対的に天然状態が不安定化したもの である。前述したようにタンパク質の安定性は天然状態と変性状態の差であるため、
添加物のような外的要因がどちらの状態に大きく作用するのかを明らかにすること が重要となる。特に、疎水性側鎖は天然状態においてはタンパク質内部に存在する傾 向があるため添加物との接触機会が少なく、タンパク質の変性によって溶媒に露出す ることで初めて添加物と接触(相互作用)することから、添加物の影響を調べる上で 良いターゲットとなる。また、疎水性側鎖への添加物の相互作用はタンパク質の疎水 性相互作用に直接的に影響を与えることから、タンパク質の立体構造形成に及ぼす疎 水性相互作用の役割を明らかにすることにつながる。疎水性側鎖の溶媒への露出を添 加物によって制御することができれば、タンパク質の安定性や機能のコントロールに 繋がると考えられる[21,22]。
タンパク質のモルテングロビュール(Molten Globule : MG)状態は天然(Native : N) 状態と変性(Unfolded : U)状態の間の中間的な状態である[6-11]。タンパク質の立体 構造形成機構を明らかにするためには、天然状態や変性状態だけでなく MG 状態のよ うな変性中間体の構造や安定性、形成機構を明らかにすることが重要である。そのた め、MG 状態の熱力学的、分光学的、速度論的な研究が 1990 年代より多く行われて きたが、その多くは塩によって誘起された静電的相互作用に起因する MG 状態につい てであった[12,23-27]。一般に、酸性条件下のタンパク質は表面電荷が正に帯電するた め、分子内静電反発によって二次構造や三次構造が崩壊し、変性(酸変性)する。こ の酸変性状態(Acid Unfolded:AU 状態)に塩を添加することによって、静電反発が 緩和され巻き戻り、二次構造は存在するが三次構造が崩壊した中間的な MG 状態にな る。塩添加による MG 形成は実験条件的にも容易であるため、多くの塩誘起 MG 状態 に関する研究が行われ、静電的相互作用のフォールディングへの影響について多くの 議論がなされてきたが、疎水性相互作用の効果に関する議論は十分にはなされてこな かった。我々は過去の研究において、酸変性状態のタンパク質にポリオールを添加す ることによって、静電反発の緩和ではなく、疎水性相互作用を強化することによって MG 状態を誘導する新しい方法とその MG 状態の熱力学的特性について報告した[17]。 ポリオール誘起 MG 状態の構造や安定性は塩誘起 MG 状態と異なっており、新しい変
性中間体の存在を示唆するとともに、適当な添加物を加えることにより疎水性相互作 用をコントロールし、タンパク質の構造や安定性を制御できる可能性を示している。 しかし、疎水性相互作用による MG 状態に関する報告例は少なく、疎水性相互作用の 役割や MG 状態の熱力学的特性を議論するには至っていない。 本章では、ポリオール誘起 MG 状態への転移機構を明らかにするために、6 つのヒ ドロキシ基を有し、過去の我々の研究において最もタンパク質の巻き戻り効果が大き かったソルビトールを用い、ウマ心筋由来のミオグロビンの変性中間体の構造および 安定性について調べた。ミオグロビンは過去に測定したシトクロム c のような典型的 な単量体のヘムタンパク質であり、その塩誘起の MG 状態は多く研究がなされている [28-32]。DSC、CD、粘度計を用いた平衡論的解析や選択的溶媒和解析、速度論的解 析により、ソルビトール誘起 MG 状態の熱力学的特性を明らかにした。 1.2 実験 1.2.1 試料 モデルタンパク質として用いたウマ心筋由来ミオグロビンは Sigma-Aldrich(95 %) から購入し、milli-Q 水を用いて試料調製した。あらかじめ濃度の濃いミオグロビン溶 液を調製し分光光度計を用いて、モル吸光係数555 = 12.92×103 M-1 cm-1[33]での吸光度 測定により濃度を決定した。ソルビトール、水酸化カリウム、クロロ酢酸、塩酸はキ シダ化学の特級試薬を精製することなく使用した。水酸化カリウムとクロロ酢酸を用 いた緩衝溶液(10 mM)は pH 2.1 であった。 分光測定のため、ミオグロビンの濃い溶液を一定量用いてソルビトール溶液と混合 し、タンパク質濃度(0.3 mg mL-1)を一定に調製した様々なソルビトール濃度(0-5 M) の試料溶液を上記の緩衝溶液中で調製した。各溶液の質量比を決定するために、タン パク質およびソルビトールの正確な濃度を、溶液の密度を用いて決定した。DSC と粘 度測定のために、濃いミオグロビン溶液を一定量用いてソルビトール溶液と milli-Q 水と混合することにより、タンパク質(3-4 mg mL-1)およびソルビトール(0, 3.0, 3.5,
4.0 M)を含む試料溶液を調製し、沈殿を防ぐために最後に塩酸を 10 mM になるまで 加えた。また、各濃度のソルビトール溶液をリファレンスとして調製した。 1.3 結果および考察 1.3.1 ソルビトール存在下における構造解析 タンパク質の円二色性は、溶液中のタンパク質 の二次構造変化を測定する上で有効的な手法で ある。222 nm の波長におけるモル楕円率はタンパ ク質内の α へリックスの含有量を反映する[34]。 Fig. 1 に 20oC におけるミオグロビンの水中(pH 6.4)および各ソルビトール濃度(0, 1.0, 2.0, 3.0, 4.0, 5.0 M、pH2.1)水溶液中の遠紫外 CD スペクトル を示した。ソルビトール 0 M(pH 2.1)の 222 nm におけるモル楕円率を水溶液中の値と比較する と、強度が大きく減少しているため、ミオグロビ ンの二次構造は失われ、酸変性状態に転移したと 考えられる。酸変性状態のミオグロビンにソルビ トールを添加すると、濃度の増加とともに CD 強 度は負に増加した。ソルビトール濃度 5.0 M では、 222 nm の楕円率は天然状態と同程度であるが、 208 nm の楕円率が天然状態よりも強度が負に大 きいため、天然状態とは異なる構造に転移してい ると考えられる。また、トリプトファンなどの芳 香族アミノ酸周りの環境(つまり、フォールディ ングの有無や三次構造変化)を反映する近紫外 CD スペクトル(Fig. 2)を見ると、pH 2.1 およびソルビトールを 4 M まで添加したスペクトルはほとんど楕円率が変化せず、 200 210 220 230 240 250 -20 -15 -10 -5 0 [ ] / 1 0 3 d eg c m 2 d m o l -1 / nm
Fig. 1 Far-UV CD spectra of myoglobin in sorbitol solutions (0, 1.0, 2.0, 3.0, 4.0, and 5.0 M) at pH 2.1 and 20 oC. Native state (in water):open circle, acid-unfolded state (in pH2.1):filled triangle, MG state (in pH 2.1 and 5.0 M sorbitol):filled square. 250 275 300 325 350 -0.1 0.0 0.1 0.2 [ ] / 1 0 3 d e g c m 2 d m o l -1 / nm
Fig. 2 Near-UV CD spectra of myoglobin in sorbitol solutions (0, 1.0, 2.0, 3.0, 4.0, and 5.0 M) at pH 2.1 and 20 oC. Native state (in water):open circle, acid-unfolded state (in pH2.1):filled triangle, MG state (in pH 2.1 and 5.0 M sorbitol):filled square.
天然状態とは大きく異なっており、三次構造が崩 壊していると考えられる。またソルビトール 5 M を添加した結果も、255 nm 付近にピークが見られ るが天然状態とは異なる構造になったと考えられ る。Fig. 3 に励起波長 290 nm の蛍光スペクトルを 示した。蛍光スペクトルではトリプトファン周り の溶媒の極性を反映するため、近紫外 CD と同様 に三次構造変化を評価できる。天然状態では 323 nm に極大のピークが観測され、酸変性すると、335 nm にピークがレッドシフトした。これは、タンパ ク質が酸変性することにより、内側に存在したトリプトファンが水溶液中に露出した ためだと考えられる。ソルビトールを添加すると、濃度が増加するにつれてブルーシ フトし、トリプトファン周りが疎水性環境に戻ったと考えられる。CD と蛍光の結果 から、二次構造はソルビトールの添加により、酸変性状態から天然状態に近い構造に 巻き戻ったが、三次構造はソルビトールを加えてもまだ壊れていると考えられる。つ まり、ミオグロビンはシトクロム c[17]と同様に、天然状態とは異なる中間状態(MG 状態)に巻き戻ったと考えられる。 1.3.2 MG 状態の熱力学的解析 ミオグロビンの二次構造のソルビトール濃度依存性は、ソルビトール誘起 MG 状態 に対する熱力学的情報を示す。Fig. 4 に様々な温度における 222 nm のモル楕円率のソ ルビトール濃度依存性を示した。酸変性状態(AU)から MG への転移は全ての系で 可逆的であった。AU と MG の二状態転移(AU⇔MG)におけるギブズエネルギー変 化(G = GMG-GAU)は式(3)を用いて算出した。
[ ] [ ]
] [ ] [ ln ln MG AU G RT K RT (3) 300 325 350 375 400 425 450 0 2 4 6 8 10 A rb it ra ry u n it / nmFig. 3 Fluorescence spectra of myoglobin in sorbitol solutions (0, 1.0, 2.0, 3.0, 4.0, and 5.0 M) at pH 2.1 and 20 oC. Native state (in water):open circle, acid-unfolded state (in pH2.1):filled triangle, MG state (in pH 2.1 and 5.0 M sorbitol):filled square.
ここで、R は気体定数、T は各測定における温度[K]、K は平衡定数、[]AUと[]MGは それぞれ AU 状態と MG 状態のモル楕円率の 値、また、[]は AU⇔MG 転移における任意の 濃度でのモル楕円率である。算出したギブス エネルギー変化をG = 0 kJ mol-1付近でプロ ットしたものが Fig. 5 である。いずれの温度 においても 2 つのパラメーターの間に直線関 係が見られたため、ソルビトール非存在下(0 M)における AU 状態から MG 状態へのギブ ズエネルギー変化(Go)は式(4)より算出 された[35]。 ] [ o sorbitol m G G (4) ここで、m は AU⇔MG 転移の協同性を示す値である。また、m とソルビトール濃度 から転移の中点濃度である Cmを算出できる。酸変性状態からソルビトール 0 M にお けるミオグロビンのソルビトール誘起 MG 状態への転移に対する熱力学量を Table 1 にまとめた。ソルビトール 0 M における MG 状態のGoの値は 6.72-9.13 kJ mol-1(pH 2.1)とミオグロビンの尿素変性における値(28.0 kJ mol-1)と比較すると非常に小さ い値を示した[36]。ソルビトール非存在下における AU⇔MG 転移に対するエンタルピ ー変化(Ho)、エントロピー変化(So)および熱容量変化(CPo)はGo の温度依 存性と以下の熱力学的関係式(5)-(7)より算出した。
T T G H / 1 d / d o o (5) T G H S o o o (6) T H C d d o o P (7) 0 1 2 3 4 5 -20 -15 -10 -5 0 [ ]222 / 1 0 3 d e g c m 2 d m o l -1 Sorbitol/MFig. 4 Plots of []222 as a function of
sorbitol concentration at 10oC (filled circle), 20oC (filled triangle), 25oC (filled square), 30oC (filled inverted triangle), and 35 oC (filled diamond). Solid lines represent the theoretical fits to a two-state transition model using the parameter values shown in Table 1.
得られた 25o C における各熱力学量を Table 1 に示した。 ミオグロビンでのHoの-27.8 kJ mol-1を過 去のシトクロム c の MG 状態の値(-78 kJ mol-1)と比較すると非常に小さかった[17]。 また、エントロピー項(TSo)が負の値を示 していることから、MG 転移によって構造化 したことがわかる。熱容量変化( o P C )はソ ルビトール系では無視できるほど小さい。通 常、天然状態の熱変性におけるCPoは、およ そ 75 J K-1 mol-1(アミノ酸 1 mol あたり)[37,38] となる。これは、変性によってタンパク質の
ASA(Accessible Surface Area, 溶媒露出表面積)が増加するため、水和量が増加し、 変性状態の熱容量が増加するためだと考えられる。つまり、MG 転移における小さな
o P
C
は MG 状態の水和量が酸変性状態とあまり変わらないことを示している。
Table 1 Thermodynamic parameters for the sorbitol-induced MG state of myoglobin T o C ΔGo kJ mol-1 M kJ mol-1 M-1 Cm M ΔHo kJ mol-1 TΔSo kJ mol-1 ΔCp kJ mol-1 K-1 10 6.72 ± 0.45 -3.37 ± 0.19 2.00 -27.8 -36.2 0 20 7.67 ± 0.18 -3.60 ± 0.08 2.13 25 8.37 ± 0.39 -3.82 ± 0.15 2.19 30 8.82 ± 0.31 -3.97 ± 0.13 2.22 35 9.13 ± 0.22 -3.92 ± 0.09 2.33 溶液中でのタンパク質の構造の広がりは固有粘度のような流体力学的特性に反映 0 1 2 3 4 5 -5 0 5 10 G / k J m o l -1 Sorbitol/M
Fig. 5 Plots of G as a function of sorbitol concentration at 10oC (filled circle), 20oC (filled triangle), 25oC (filled square), 30oC (filled inverted triangle), and 35 oC (filled diamond). Solid lines represent the theoretical fits to a two-state transition model using the parameter values shown in Table 1.
される[39]。25o C 水中での天然状態、10 mM 塩酸中での AU 状態と 10 mM 塩酸およ びソルビトール 3.0 M 中での MG 状態のミオグロビンの固有粘度はそれぞれ 4.2、32、 10.8 cm3 g-1となった。これらの流体力学的値は、MG 状態の三次元構造が緩やかに詰 まっており、N 状態と AU 状態の中間の大きさであること強く示唆している。以上の 結果より、ソルビトール誘起 MG 状態は二次構造があり、三次構造は N 状態ほど巻 き戻っておらず、水和の多い非常に膨潤した構造であることがわかった。 1.3.3 MG 状態の熱安定性(CD 測定) タンパク質の温度に対する熱的挙動 は様々な熱力学量を反映する。様々な ソルビトール濃度におけるミオグロビ ンのモル楕円率(222 nm)の温度依存 性を Fig. 6 に示した。温度増加にとも なって楕円率強度の減少、つまりミオ グロビンのへリックスの含量が減少 しており、熱変性状態に転移したと考 えられる。また、ソルビトール濃度が 増加するとともに転移曲線は高温側に シフトしており、MG 状態はソルビトールの添加により熱に対して安定化していると 考えられる。ソルビトール濃度が低い場合、低温においてもモル楕円率の強度が小さ いことから、Fig. 1 で示したようにまだ MG 状態に巻き戻っていないと考えられ、3.0 M 以上では低温で MG 状態に転移していると考えられる。222 nm における MG 状態 の楕円率強度は N 状態に近く、二次構造量が同等であることを示唆しているが、温度 に対する耐性は大きく異なっている。MG 状態(3~5 M)の楕円率強度は温度増加に ともなって減少しており、二次構造の減少、つまり熱変性状態へ転移していることが わかるが、非常に協同性が低い転移を示している。一方、N 状態(in water)は 80℃ 20 40 60 80 -15 -10 -5 N (in water) 5.0 M 4.0 M 3.0 M 2.0 M 1.0 M [ ] 222 /1 0 3 d eg cm 2 d mo l -1 T/oC
Fig. 6 Temperature dependence of []222 of
myoglobin in buffer solution (pH 2.1) at various concentration of sorbitol and in water (pH 6.4).
付近から協同性の高い(転移が始まると協同的に崩壊が進行する)熱変性を示した。 熱力学的には熱変性における協同性の違いは熱変性における熱容量変化の違いを反 映している。MG⇔AU の二状態転移と転移に対する熱容量変化を考察するために、 熱転移における熱力学量を式(3)と(8)から最小二乗法により求めた。
m m P m m 1 ln T T T T T C T T H G (8) それぞれ熱変性に対する、Tm は変性中点温度であり、ΔHm は変性中点におけるファ ントホッフのエンタルピー変化、ΔCpは熱容量変化になる。得られた熱力学量を Table 2 に示した。これらは様々なソルビトール濃度における変性中点温度での MG 状態か ら熱変性状態への転移に対する熱力学量を示しており、Table 1 に示したソルビトール 非存在下における 25o C での AU 状態から MG 状態への転移に対する熱力学量と単純 に比較することは難しい。ΔHmの値より、MG 状態から熱変性状態への転移は吸熱反 応になり、ソルビトール濃度が増加するにつれて値は大きくなった。しかし、天然状 態から熱変性状態への転移に比べると ΔHm は半分以下になった。高温でのモル楕円 率を比較すると、熱変性状態はソルビトール濃度に大きく依存せず、ΔHmの差は MG 状態と天然状態の差を反映したと考えられる。つまり、小さい ΔHmの値は MG 状態 が天然状態よりも相互作用数が少ないことを強く示唆している。また、熱容量変化ΔCp の値を比較すると、MG から熱変性状態への変化量は天然から熱変性状態への変化量 より小さかった。このような小さなΔCpが、温度に対する転移の協同性の低さを表し ている。(熱力学的にΔCpはΔG の温度に対する曲率を反映するからである。)さらに、 ΔCpの値はタンパク質の水和量に大きく依存するため、値が小さいということは MG 状態の水和量が非常に少なく、熱変性状態と同程度であると考えられる[37,38]。これ らの熱力学量はソルビトール誘起の MG 状態が天然状態と同程度の二次構造をもっ ているが、立体構造を形成する相互作用の量は少なく、水和量の多い膨潤した構造で あることを示している。Table 2 Thermodynamic parameters for the thermal unfolding of the N and MG states of myoglobin obtained by CD State pH Sorbitol M ΔHm kJ mol-1 Tm o C ΔCp kJ mol-1 K-1 N 6.4 0 579 ± 6 83.4 ± 0.1 11.5 ± 1.8 MG 2.1 3.0 121 ± 1 42.6 ± 0.1 0.3 ± 0.2 2.1 4.0 140 ± 2 59.2 ± 0.1 1.3 ± 0.3 2.1 5.0 206 ± 3 72.8 ± 0.1 2.4 ± 0.8 1.3.4 MG 状態の熱安定性(DSC 測定) ソルビトール誘起 MG 状態の DSC 結 果を Fig. 7 に示した。比較のために天 然状態と酸変性状態の温度依存性を合 わせて示した。pH 2 の AU 状態におけ る DSC 測定では熱転移に伴うピークが 観測されなかったため、AU 状態と熱変 性状態は熱力学的に近い状態であるこ とを示している。しかし、ソルビトー ルを加えると幅広く小さい吸熱ピーク があり、濃度が増加するとピークが高 温側にシフトした。天然状態と MG 状態の熱変性における変性中点温度 Tmと DSC よ り求まったエンタルピー変化Hcal、および半値幅 T1/2を Table 3 に示した。
Table 3 Thermodynamic parameters for the thermal unfolding of the N and MG states of myoglobin obtained by DSC State pH Sorbitol M Hcal kJ mol-1 Tm o C T1/2 o C N 6.4 0 327.5 ± 3.5 78.9 ± 0.1 3.9 ± 0.1 MG 2.0 3.5 39.3 ± 5.1 51.6 ± 0.8 21.1 ± 0.9 2.0 4.0 55.5 ± 6.1 53.2 ± 0.9 19.1 ± 0.6 20 40 60 80 100 20 MG (4.0 M) N AU Cp / k J m o l -1 K -1 T / oC MG (3.5 M)
Fig. 7 DSC curves of myoglobin in each state. N in water, AU in 10 mM HCl, MG (3.5 M) in 3.5 M sorbitol with 10 mM HCl, MG (4.0 M) in 4.0 M sorbitol with 10 mM HCl.
MG 状態の変性に対するHcalの値は非常に小さく、天然状態からの値に比べおよそ 10-20 %程度の値を示した。半値幅は天然状態からの値に比べ MG 状態は非常に大き く、CD 測定の結果でも示されたように協同性が低いことを表している。これらの熱 力学量の違いはソルビトールにより誘起された状態が天然状態とは異なり MG 状態 であることを示し、ソルビトール誘起 MG 状態の三次構造は天然状態よりも相互作用 数が少ないことを示唆している。熱測定(DSC)から直接的に求まったHcalと CD 測 定の値をもとに熱力学の関係式より求まった(一般にファントホッフエンタルピーと 呼ばれる)Hm(Table 2)は異なっていた。Hmは二状態転移を仮定することにより 熱力学的に得られるため、Hcal とは必ずしも一致するわけではない。通常、Hm と Hcalの比が 1 よりも大きい場合、熱転移に会合や解離の過程を伴うことを示している [37,38]。しかし、タンパク質の全体的な構造変化を反映する DSC によって得られる 熱力学量は、二次構造の変化のみを観測する CD 測定から得られる熱力学量と同じで ない可能性があるため、単純な比較は難しい。また、CD で得られた Tmは DSC より も高く、ソルビトール濃度の増加にともない同様の傾向を示した。これらの結果は、 三次構造が崩壊した後に二次構造が変性したことを示している。 後藤ら [31]によると、アポミオグロビンの塩誘起 MG 状態は、本測定により得られ たミオグロビンのソルビトール誘起 MG 状態と同様に、低い協同性と小さいHcal (80-90 kJ mol-1 , 80oC)を示している。糖アルコールはたとえ高濃度であっても、イ オン対を容易に形成しないため、水の誘電率を大きく低下させることがなく、静電的 相互作用にあまり影響しないと考えられる[40]。つまり、それぞれの MG 状態の熱力 学的性質は似ているが、ソルビトールは静電的相互作用に影響を与えることなく、疎 水性相互作用の影響によりリフォールディングを誘発したと考えられる。 過去の研究でのシトクロム c のソルビトール誘起 MG 状態は、本測定により得られ たミオグロビンのソルビトール誘起 MG 状態の熱力学的性質と異なり、高い協同性と Hcalを示していた[17]。このように、ソルビトールの添加により疎水性相互作用が強 化され酸変性状態がリフォールディングしたものの構造や熱力学的特性が大きく異
なっていることは、タンパク質の立体構造形成機構が個々のタンパク質によって異な っていることを示唆している。ミオグロビンとシトクロム c はともに α へリックスが 多いタンパク質であるが、分子量や柔軟性(圧縮率)が大きく異なっている。また、 大きな違いは、ともにヘムタンパク質であるがその結合様式である。シトクロム c の ヘム基は共有結合により結合しているため、変性してもヘム基が外れることはない。 しかし、ミオグロビンのヘム基は非共有結合により結合しているため、変性するとヘ ム基が外れて、より協同性と熱安定性の低いアポミオグロビンへ転移する[30, 31]。ヘ ム基の存在がヘムタンパクにとっては、天然状態だけでなく MG 状態の厳密な三次構 造を保つためには重要な因子であると考えられる。 1.3.5 ソルビトールの添加による MG 状態への転移機構 熱力学的観点から、ソルビトールを 加えたときに AU 状態から MG 状態へ 転移する機構は、MG 状態が AU 状態 よりも安定であることを示している。 MG 状態へのソルビトールの影響をギ ブズエネルギー変化で示した(Scheme 1)。ここで、G0とG はそれぞれ pH 2.1 の水中における AU 状態から MG 状態への自由エネルギー変化と pH 2.1 のソルビトー ル溶液中における AU 状態から MG 状態への自由エネルギー変化を示している。また、 AU tr G とGtrMGはそれぞれ AU 状態と MG 状態の pH 2.1 における水中からソルビトー ル溶液中への変化を示している。Table 1 のG0は MG 状態が 25oC においてソルビト ール非存在下での AU 状態よりも 8.37 kJ mol-1ほど不安定であると示した。25o C にお けるソルビトール 4 M でのG の値は-6.92 kJ mol-1と見積もられ、G はG0より小さ い値となった。水中での AU 状態がソルビトール中へ移行し、その後 MG 状態へと転 Scheme 1 Diagram of free energy changes in the processes.
移するルートのGtrAU+ G と、水中での AU 状態が MG 状態に転移しその後ソルビ トール溶液中に移行するルートのG0+GtrMGは等しくなる。言い換えれば、G-G0 とGtrMG-GtrAUは等しくなる。つまり、pH 2.1 において、各溶液中での AU 状態か ら MG 状態への自由エネルギー変化の差は、各状態を水中からソルビトール溶液中へ 移行させる自由エネルギー変化の差と等しくなる。G0はG よりも大きいことから、 AU tr G はGtrMGよりも大きくなるはずである。これは、多成分溶液の熱力学を用いて 説明することができる。三成分溶液系で水を第 1 成分、タンパク質を第 2 成分、ソル ビトールを第 3 成分とすると、ソルビトールの選択的溶媒和3は式(9)になる[41]。
3 2
T,μ1,μ3 3 m / m (9) ここで、m は各成分のモル濃度を示し下付き数字は 1 が水、2 はタンパク質、3 はソ ルビトールを示す。3はバルク溶媒と比較してどの成分がタンパク質周りに選択的に 存在しているのかを示す。3 と実際の水とソルビトールの結合数(B1と B3)との間 の関係は3 = B3-(m3/m1)B1で示される。つまり、AU と MG の選択的溶媒和3の差 である33MG 3AUは以下の式(10)-(12)で表される。
AU μ3 μ1, T, 2 3 MG μ3 μ1, T, 2 3 3 m /m m /m (10)
AU
T,μ1,μ3 3 MG 3 1 3 AU 3 MG 3 3 B B m /m B B (11)
3
3 dlnK/dlna (12) ここで、K は AU と MG 間の平衡定数であり、a3はソルビトールの活量を表す[42-44]。 3の値は AU 状態と MG 状態のどちらに選択的にソルビトールが結合しているのか を表す。式(12)を用いて3 の値を求めるため、ソルビトール濃度と平衡定数の対 数の相関を Fig. 8 に示した。m3は pH 2.1 でソルビトールの活量は報告されていないた め、活量係数が一定であると仮定し、a3の代わりに用いた。Fig. 8 の傾きが3の値と なるため、全ての温度で3はおよそ 3 となる。また、すべて3が正の値であるため、(∂m3/∂m2)MGは(∂m3/∂m2)AU より大きくなる。これら の結果は、ソルビトール分子が AU 状態よりも MG 状態に選択的に結合、または、水分子が AU 状態よ りも MG 状態から選択的に排斥されていることを 意味している。これは、ソルビトール分子が 6 個の ヒドロキシ基を有しており、AU 状態で表面に露出 した疎水性側鎖から選択的に排斥されているもの と思われる。つまり、疎水性側鎖が露出していない MG 状態は AU 状態と比べてソルビトールが排斥さ れておらず、このソルビトールの各状態への親和性の違いによって、ソルビトール存 在下では AU 状態よりも MG 状態のほうが相対的に安定(平衡が移動する)になると 考えられる。Table 1 に示したように、MG 状態で観測されたCP0は無視できるほど小 さいため、MG 状態の水和量(B1MG)と AU 状態の水和量(B1AU)は大きく異なるわけでは なく、3 の正の値は主にソルビトールの結合数が異なることを示すと考えられる (B3MG > B3AU)。3はおよそ 3 であるので、3 つのソルビトール分子が AU 状態に比べ て MG 状態に多く結合しているか、MG 状態に比べて AU 状態から排斥されていると 考えられる。以上のように、ソルビトールの添加による MG 状態への巻き戻りは、ソ ルビトールの MG 状態への選択的溶媒和(=AU 状態からの選択的排斥)を介して、 生じていることが明らかとなった。 1.3.6 MG 状態の速度論的特性 フォールディングおよびリフォールディング過程をストップトフローCD により測 定した。Fig. 9 はソルビトール 0 M と 3.3 M における中性から酸性への pH ジャンプ における[]222のストップトフロー測定の結果である。ソルビトール非存在下(A)では、 およそ 200 ms 程度で[]222強度が減少していることから、ミオグロビンのα へリック 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 -2 -1 0 1 2 ln K ln m 3
Fig. 8 Wyman plot for the sotbitol effect of myoglobin at 10oC (filled circle), 20oC (filled triangle), 25oC (filled square), 30oC (filled inverted triangle), and 35 oC (filled diamond).
スが減少していることがわかり、天然 状態から酸変性状態に転移しているこ とを示している。Konermann は pH 3.4 条件で 441 nm のミオグロビンのヘム 基の吸光度を観察すると、ミオグロビ ンは 2 段階の変性を示し、これは 0.7 s でヘムが解離し溶媒へ溶け出し、0.16 s で中間体が形成することに対応したと 報告している[45, 46]。一方、ソルビトール 3.3 M(B)では、初め強度が減少しているが、 その後一定の値になり、700 ms 以降において強度が負に増加した。この結果は、酸性 条件になると一度α へリックスの量は減少するが、時間が経つにつれて巻き戻り α へ リックスの量が増加することを示している。つまり、速度論的には、N 状態から AU 状態を経由して MG 状態に転移すると考えられる。N 状態から AU 状態への転移と AU 状態から MG 状態への転移を単純な一次反応で仮定すると、見かけの速度定数 kapp は以下の式(13)を用いて決定できる。 f app i 222 exp( ) k t (13) ここで t は時間であり、222は各時間に対する 222 nm のモル楕円率、iは初め のモル楕円率、fは最後のモル楕円率を示す。それぞれ、N⇒AU 転移については、 初めが N 状態で最後が AU 状態であり、N⇒MG 転移については、初めが N 状態で最 後が MG 状態のモル楕円率を表す。見かけの速度定数 kappの値を算出し、混合後の塩 酸濃度([HCl]fin)に対してプロットしたものを Fig. 10 に示した。 ソルビトール存在下において、混合後の塩酸濃度が 6 mM の場合は N⇒AU の kapp の値は 7.79 ms-1となり、ソルビトール非存在下における値(13.5 ms-1)より 40 %程度 小さい値を示した。また、すべての[HCl]finにおいて、ソルビトール存在下の kappの値 は小さくなった。流体力学的観点から 3.3 M のソルビトール溶液で得られた粘度の値 0 500 1000 1500 -40 -30 -20 -10 0 10 [ ] 222 / 1 0 3 d eg c m 2 d m o l -1 t / ms
Fig. 9 Time dependence of []222 of myoglobin at
pH jumping from neutral to acidic in the presence of 0 M (A) and 3.3 M (B) sorbitol.
(A)
(9.6 mPa s, 25o C)は、水の粘度の値(0.982 mPa s, 25oC)よりも非常に高いため、溶 媒の影響により N⇒AU の kappの値が減少したと 考えられる。Hagen[47]や Tang ら[48]は様々なタ ンパク質のフォールディングに対する時間は溶 媒の粘度の増加とともに長くなることを示した。 また別の観点から kappの値が減少した理由として、 3.3 M の溶媒中ではソルビトールが体積分率で 60 %程度に相当するため、ソルビトール分子がタ ンパク質の変性を立体障害的に阻害したことが 考えられる。 一方、ソルビトール存在下において、[HCl]fin
が 6 mM の場合は AU⇒MG の kappの値は 3.21 ms-1となり、N⇒AU の値(7.79 ms-1)
と比較すると半分程度でゆっくり巻き戻っていることがわかる。ソルビトール存在下 では[HCl]finに対して kapp,N⇒AUは正の相関を示し、kapp,AU⇒MGは負の相関を示した。つ
まり、N⇒AU の転移は pH が低下することによってアスパラギン酸やグルタミン酸が 中性化し、リジンやアルギニンなどの正に帯電したアミノ酸間で静電反発が生じてす ぐに進行したと考えられる。また、AU⇒MG のリフォールディング過程では、1.3.4 で示したように、AU から 3 つのソルビトール分子が選択的に排斥されるなどの複雑 なメカニズムにより徐々に進行すると考えられる。 AU から MG 状態への転移は AU と MG との間の平衡反応の緩和過程とみなすこと ができる。したがって、見かけの速度定数 kapp,AU⇒MGは kAU⇒MGと kMG⇒AUの和(kapp,AU ⇒MG = kAU⇒MG + kMG⇒AU)、また、平衡定数 K=[AU]/[MG]は kAU⇒MG/kMG⇒AUとなる。kAU ⇒MGは AU から MG への速度定数であり、kMG⇒AUは MG から AU への速度定数である。 1.3.2 の Fig. 4 で示した平衡系の結果より、10.9oC でソルビトール 3.3 M の平衡定数 K は 10.7 と見積もられていることから、kMG⇒AUは 1.84×10-4 s-1で kAU⇒MGは 1.98×10-3 s-1 となり、この環境条件(10.9o C、3.3 M)において MG 状態から AU 状態へのアンフォ 5 6 7 8 9 10 0 5 10 15 20 25 30 AU ⇒ MG N ⇒ AU N ⇒ AU 10 3 k a p p / s -1 [HCl]fin / mM
Fig. 10 Plots of kinetic constants of folding and refolding of myoglobin against the final concentration of HCl. From N to AU in the absence of sorbitol (open circle) and from N to AU (filled triangle) and from AU to MG (filled square) in the presence fo 3.3 M sorbitol.
ールディングは AU 状態から MG 状態へのリフォールディングはよりも 10 倍遅いこ とが明らかとなった。 1.4 1 章まとめ 酸変性したミオグロビンはソルビトールの添加によって巻き戻り、その構造は CD スペクトルや蛍光スペクトルの違い、熱変性における小さなエンタルピー変化、低い 協同性、小さい熱容量変化から判断されるように、天然状態のものと同様の二次構造 ではあるが膨潤した中間的な状態(MG 状態)であった。この巻き戻りは MG 状態と 比較して AU 状態からソルビトールが 3 分子選択的に排斥したことにより誘導された ものであり、疎水性相互作用の相対的な強化によるものである。天然状態から MG 状 態への転移は、AU 状態を介して進行しており、直接的な N→MG 転移は生じていな い。今回ミオグロビンにおいて得られたソルビトール誘起 MG 状態は過去のシトクロ ム c とは大きく異なっており、ヘム基の存在がヘムタンパクにとっては、天然状態だ けでなく変性中間体の構造形成においても重要な因子であると考えられる。
2 章 1,4-ジオキサン添加によるシトクロム c の線維状凝集体の熱力学的研究 2.1 序論 タンパク質の立体配座はフォールディングされた天然状態、天然状態で存在する三 次構造や二次構造を失いアンフォールディングした変性状態、モルテングロビュール のような中間状態、アモルファス凝集のような会合状態、およびアミロイド病に起因 するような規則正しい凝集体などがある。規則正しい凝集体は多くのタンパク質で観 測されており、タンパク質の基本形態の一つである可能性がある[13-15]。このような 凝集形成機構の解明は、タンパク質病の予防と治療の双方の面で重要である。これま で凝集体の構造については、核磁気共鳴(NMR)や X 線構造解析、円偏光二色性(CD) [49-55]で明らかにされているが、凝集プロセスに関する熱力学的データはほとんど存 在せず、J.Kardos ら[56]と Sasahara ら[57]は等温滴定型微少熱量計(ITC)や示差走査 熱量計(DSC)による2-microglobulin の凝集過程の速度論的および熱力学的特性を報 告している程度である。熱力学的データが少ない理由の一つとして、凝集体形成によ りタンパク質の溶解度が低下するため、分光強度の変化を定量的に解釈することが難 しいことが挙げられる。一方、熱分析は凝集や沈殿を含む全反応を巨視的に観測する ため、分光法では得られないデータを補正する上で効果的な手法である。 また、凝集プロセスを観測するためには、タンパク質を凝集させるために適切な条 件を選択することも重要である。凝集させる手法の一つとして、タンパク質の等電点 付近でタンパク質間の静電反発を減少させる方法がある。しかし、この方法では多く の場合、規則正しい構造の凝集体ではなくアモルファス状の沈殿物を生じる。また、 アミノ酸置換によって規則的なアミロイド凝集体を形成することが報告されている が[58,59]、全てのタンパク質に適用できるわけではなく、汎用性が低い。さらに、凝 集を引き起こす手法の一つに超音波法があるが[60,61]、超音波は非常に大きな熱を発 生するため凝集プロセスに関する熱力学的情報を検出することは容易ではない。また、 緩衝溶液の変化なども熱測定や熱分析の測定値に影響を及ぼすため、凝集に関する熱
力学的情報を正確に決定するためにはできるだけ凝集化の条件を単純にすることが 望ましい。 我々はこれらの問題点を解消するため、1,4-ジオキサンを共溶媒としてタンパク質 に添加し、加熱することによって凝集化に伴う熱力学量を観測する手法を確立した。 この凝集化方法は単純であるため、病原性のタンパク質だけでなく、一般的な球状タ ンパク質の凝集機構を明らかにするモデルとして用いることもでき、凝集を起こすモ デルタンパク質の数が増えることはその機構を明らかにするために有効である。本研 究で用いたシトクロム c は典型的な球状および単量体のタンパク質であり、特殊な条 件下ではあるが非天然状態に関する報告もある[17,62]。また、線維状凝集体に関する 報告[18,63]などもあるため、適切なモデルタンパク質である。この章ではシトクロム c の線維状凝集体形成における構造と転移機構ならびに熱力学量について考察を行う。 2.2 実験 2.2.1 試料 ウシ心臓由来のシトクロム c は Sigma-Aldrich(95%)から購入し、milli-Q 水を用い て試料調製した。1,4-ジオキサン(関東化学、純度 99.9%)は、モレキュラーシーブ ス 4A (キシダ化学, 1/16 ペレット) を、 真空系を用いて 10-3 ~ 10-2 Pa の高真空で まず室温で 30 分程度排気した後、約 453 K で 8 時間以上排気し活性化したものを用 いて脱水した。あらかじめ濃度の高いシトクロム c 溶液を調製し 280 nm での吸光度 測定により濃度を決定した。高い濃度のシトクロム c 溶液を所定の重量だけとり、ジ オキサンおよび milli-Q 水を混合することにより試料調製を行った。シトクロム c の 最終的な濃度およびジオキサンのモル分率は溶媒と溶液の重量と密度データから得 られた希釈率を用いて決定した。溶液のタンパク質濃度は DSC 測定ではおよそ 0.50 w/w%であり、CD 測定では 0.050 w/w%であった。シトクロム c の量は 1,4-ジオキサン および水の量と比較すると無視できるくらい小さいので、溶液のジオキサンモル分率 は 1,4-ジオキサンおよび水の二成分系として計算した。
2.3 結果および考察 2.3.1 シトクロム c の DSC 測定 シトクロム c の各モル分率におけるジ オキサン溶液の DSC 曲線を Fig. 11 に示 した。シトクロム c が熱変性するときに は吸熱ピークが見られ、ジオキサンモル 分率が増加するにつれて吸熱ピークは 低温側にシフトした。吸熱ピークはジオ キサンのモル分率が 0-0.20 の間で観測さ れた。吸熱ピークは変性することにより シトクロム c の二次構造や三次構造が失 われることを反映する。また連続測定を 行ったところ同様のピークが観測され たため、熱変性は可逆的であることが分 かる。 モル分率が 0.20 以上では発熱ピーク が観測された。二回目の連続測定ではピ ークが観測されず、赤色であったシトク ロム c 溶液は加熱後に赤色の沈殿物が発 生し溶液は無色になった。つまり、発熱 反応は不可逆でありシトクロム c の大部 分は最初の加熱において凝集したものと 考えられる。また、モル分率が 0.4 以上では加熱による発熱ピークは観測されなかっ た。加熱後の試料を SEM で観測したところ、Fig. 12 で示したとおり、モル分率 0.3-0.4 の範囲で試料溶液中に構造の異なる沈殿物が観測された。これらの結果は、発熱ピー クが変性状態から線維状沈殿への不可逆的転移を示すことを示唆している。一般に、 20 30 40 50 60 70 80 90 -0.4 -0.2 0.0 0.2 0.4 2 3 4 Exo. 0.30 0.25 0.20 0.15 0.10 0 0.05 H ea t fl o w / m W T / oC
Fig. 11 DSC results of cytochrome c in (1-x) water + x dioxane. The numbers in the figure are the mole fractions of dioxane, x. the dotted line are integration baselines.
Fig. 12 SEM images of thermal-induced aggregated cytochrome c. (a)dioxane mole fraction, x = 0.30; (b) x = 0.40.
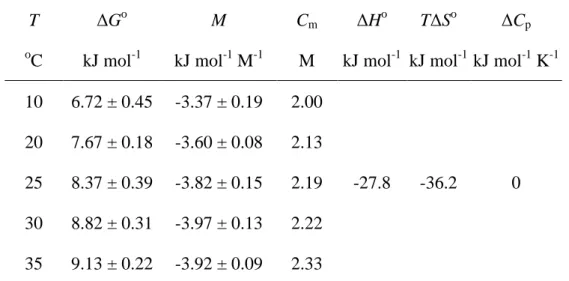
タンパク質の凝集現象は変性中もしくは変性後に起こることが多い[64,65]。これは変 性によって露出した疎水性残基が疎水性相互作用による分子間会合に寄与するため である[66,67]。DSC 測定に用いる濃度では熱変性と凝集を別々に観測できる例はほと んどない。したがって、多くの場合、各転移の熱力学量を定量的に評価することは困 難である。しかし、Fig.11 に見られるようにモル分率 0.20 においては変性および凝集 のピークが両方観測されており、各転移に関するエンタルピー変化を決定することが 可能になった。 熱変性および線維状凝集体への転移に対する、温度の半値幅はそれぞれおよそ 10o C と 4o C であり、線維状凝集体への転移は非常に協同的であることがわかる。 サンプル調製後、試料を 20o C で 24 時間静置したときの DSC 結果には有意な変化 はなく、加熱前のシトクロム c の状態は時間に対する依存性は見られなかった。 DSC 測定により得られた変性中点温度 Td、凝集中点温度 Ta、エンタルピー変化(測 定により得られた熱をセル内のタンパク質の物質量で割って決定)の平均値と標準偏 差を Table 4 に示した。
Table 4 Thermodynamic parameters for thermal denaturation and aggregation of cytochrome c in (1-x) water + x 1, 4-dioxane.(*Comparison with literature value under similar condition (pH 4.6, 40 mM glycine buffer); Td=78.0oC, ΔHd=440 kJ mol-1 [68])
mole fraction of dioxane Td Hd Ta Ha o C kJ mol-1 oC kJ mol-1 0.00* 83.4 ± 0.1 298 ± 8 - - 0.05 62.9 ± 0.3 278 ± 7 - - 0.10 52.1 ± 0.3 214 ± 8 - - 0.15 43.8 ± 0.5 153 ± 10 - - 0.20 37.5 ± 1.0 98 ± 18 83.1 ± 0.2 -347 ± 8 0.25 - - 64.9 ± 1.5 -424 ± 11 0.30 - - 53.6 ± 1.2 -456 ± 9 0.40 - - - - ジオキサンのモル分率が増加すると、Taは減少し、凝集にともなうエンタルピー変
化Haは数値的に増加した。線維状凝集体への転移エンタルピー変化Haは-347 から -456 kJ mol-1であるのに対して熱変性に伴うエンタルピー変化Hdは 298 から 98 kJ mol-1であった。熱変性の吸熱量と比較して、線維状凝集体への転移における発熱のエ ンタルピー変化の方が絶対値的に大きいことから、線維状構造は単量体の天然状態と 比較すると分子間相互作用の形成により、相互作用の多い構造を有することを強く示 唆している。Barone ら[64]は pH 5.0 で熱変性後のウシ血清アルブミン(BSA)の凝集 について、タンパク質濃度が低い場合、凝集における大きい負のエンタルピー変化 (-685 kJ mol-1)と変性における大きい正のエンタルピー変化(524 kJ mol-1)と報告
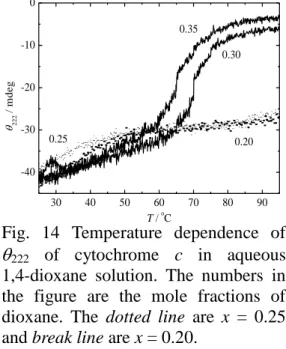
した。また、Morel ら[69]はニワトリ由来の α-spectrin の SH3-domain の線維状凝集体 における熱分解と熱変性にともなう部分エンタルピー(100 kJ mol-1)が同程度である ことを示した。これは球状タンパク質の熱変性における部分エンタルピーより有意に 小さく、α-spectrin の SH3-domain の線維状凝集体の内部の相互作用の密度が比較的低 いことを示している。しかし、シトクロム c のジオキサンを起因とした線維状凝集体 は BSA の凝集体と同様、熱変性のエンタルピー変化より凝集体形成におけるエンタ ルピー変化が数値的に大きかった。つまり、シトクロム c の線維状凝集体は天然状態 と比較すると内部の相互作用の密度が比較的多いと考えられる。 2.3.2 シトクロム c の CD 測定 タンパク質の構造転移におけるエンタルピー変化は転移前後の構造の相違を反映 する。そのため転移前と転移後の構造を決定する必要がある。Fig. 13(a)は、各モル分 率および 25o C での 1,4-ジオキサン水溶液中のシトクロム c の CD スペクトルである。 また、楕円率 222 nm の値はタンパク質の二次構造(主に α へリックス)の含有量を 反映している[34]。Fig. 13(b)に各ジオキサンモル分率に対する 222 nm の楕円率をプロ ットした。水中のタンパク質のスペクトル(x = 0.0)の形と強度は、シトクロム c が 水中では天然状態でα へリックスを多く有することを示している。モル分率が 0 から 0.15 の間では、楕円率強度が変化しなかった。0.2 から 0.3 の間ではスペクトルの形状
は天然状態と同様であるが、強度が有意に増加し た。この強度の増加はα へリックスの含有量が増 加していることを示している。モル分率 0.25 以上 では熱変性をあらわす DSC の吸熱ピークが観測 されていなかった(Fig. 11)。一般に、二次構造が 存在し、三次構造が崩壊したモルグロビュール状 態などは熱変性にともなう吸熱ピークを示す [17,62]。しかし、モル分率 0.25 以上において吸熱 ピークが観測されなかったということは、ジオキ サンにより誘起された状態が熱力学的に熱変性 状態に近い中間体であることを示唆している。ま た、モル分率 0.35 以上では、楕円率強度が急激に 減少し、モル分率 0.45 で 0 に減少した。これはシ トクロム c が凝集したことにより、不溶化したた めだと考えられる。ジオキサンはタンパク質の二 次および三次構造に影響を及ぼすことから、タンパク質の構造は、ジオキサンモル分 率に強い依存性があると考えられる。つまり、タンパク質の二次構造はジオキサンの 低い比誘電率(D = 2.1)によって、水中に比べペプチド間の水素結合が誘起されるた めに増加する[70,71]。しかし、この低い比誘電率はタンパク質の疎水性相互作用を弱 めるため、タンパク質の三次構造が崩壊すると予想される。実際に、シトクロム c の 凝集温度 Taは、高い比誘電率を示すジメチルスルホキシド(DMSO, D = 47)および 1-プロパノール(D = 20)水溶液中と比較すると 1,4 ジオキサン水溶液中の Taは低く、 凝集後の構造も異なっている。また、ジオキサンは水の性質にも影響を及ぼすと考え られる。ジオキサンは低い比誘電率を有するが水素結合部位を有するため全モル分率 で水に可溶である。ジオキサン水溶液のモル分率 0.15 における負の過剰エンタルピー [72]やモル分率 0.3 における負の過剰体積[73]に反映されるように、ジオキサンと水と 210 220 230 240 250 260 -50 -40 -30 -20 -10 0 0.15 0.50 0.40 0.30 x = 0 / m d eg / nm 0.0 0.1 0.2 0.3 0.4 0.5 -50 -40 -30 -20 -10 0 222 / m de g xDioxane
Fig. 13 (a) CD spectra of cytochrome c in (1-x) water + x 1,4-dioxane. The number beside the thick line is the mole fraction of dioxane, x. (b) 222
of cytochrome c in (1-x) water + x 1,4-dioxane.
(a)
の間には強い相互作用が存在している。また、X 線回折や NMR においてもクラスタ ーが存在していることが示されている[74]。1,4 ジオキサンと水との相互作用はタンパ ク質が天然状態を形成するのに必要な水とタンパク質との間の相互作用に影響を及 ぼすため、常温においても変性状態へ転移するものと考えられる。 Fig. 14 に x=0.2, 0.25, 0.3 および 0.35 におけるシトクロム c の θ222の温度依存性を示 した。モル分率 0.30 以上では、加熱により、強度が急激かつ不可逆的に減少した。加 熱後、赤色のタンパク質溶液が無色になり、試 料溶液中に赤色の沈殿が観測されており、強度 の低下は凝集によるタンパク質濃度の低下を示 している。0.25 未満のモル分率では、強度の急 激な低下は測定範囲内では観測されなかった。 得られた凝集体の中点温度はモル分率 0.30 で約 70oC であり、これは 2.3.1 で示した DSC 結果(Fig. 11, Table 4)の 53.6oC より高いことから、凝集は タンパク質濃度に依存することを示している。 2.3.3 シトクロム c の状態図 Fig. 15 にシトクロム c の転移温度 Tdおよび Taに基づく状態図を示した。0 から 0.15 のモル分率では天然状態(N)は加熱時に熱変性状態(DT)に変換されるが測定温度 範囲内では線維状凝集体(FA)への転移は観測されなかった。25o C では、ジオキサ ンモル分率が増加するにつれてヘリックスリッチな中間状態(Ddio)に変換した N 状 態が熱変性のエンタルピー変化が徐々に 0 に減少し、Ddio状態が熱力学的に DT状態に 近いことが示された。25o C でモル分率 0.4 以上では、CD 強度の低下(Fig. 13)より 明らかなように、加熱前にシトクロム c は凝集し不溶性になった。また、加熱前の凝 集体を SEM により観測したところ、線維状ではなくアモルファスな凝集体(AA)で あった。モル分率が 0.2 から 0.3 のとき、Ddio状態は加熱時に大きな発熱ピークをとも 30 40 50 60 70 80 90 -40 -30 -20 -10 0 0.20 0.25 0.30 0.35 222 / m d e g T / oC
Fig. 14 Temperature dependence of
222 of cytochrome c in aqueous
1,4-dioxane solution. The numbers in the figure are the mole fractions of dioxane. The dotted line are x = 0.25 and break line are x = 0.20.
ない FA 状態に変化した。しかし、モル分率 0.40 以上ではシトクロム c は加熱前に AA 状態に変換されていたため、発熱ピーク が観測されなかった。Lin ら[18]は添加された アルコール(TFH および HFIP)濃度とタン パク質濃度に対するホロシトクロム c、アポ シトクロム c およびアポシトクロム c の凝集 状態の相図を示し、本研究と同様に特定の狭 い領域でアモルファスおよび線維状凝集体を 生成することを示している。これらの結果は 凝集前の状態、特に可溶性のへリックスリッ チ状態への転移が線維状凝集体形成に重要で あることを示唆している。 2.4 2 章まとめ 本章では共溶媒として 1,4 ジオキサンを添加し DSC によるシトクロム c の線維状凝 集体形成における熱力学量を決定した。線維状凝集体への転移は熱変性にともなう吸 熱のエンタルピー変化より数値的に大きい発熱のエンタルピー変化を示し、熱力学的 に線維状凝集体が単量体の天然状態と比較して多くの分子間相互作用を有する構造 であることを示唆した。CD 測定の結果は加熱前の構造が線維状凝集体への転移に重 要であることを示した。CD 測定により得られた Taはタンパク質濃度が薄いため DSC 測定により得られたものより高かった。本研究では走査速度やタンパク質濃度の影響 については言及していないが、より詳細な凝集形成機構を明らかにするためには速度 論的な検証が必要であると考えられる。 0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40 20 40 60 80 100 DT AA Ddio FA N T ra n sition te m p er atur e / oC x
Fig. 15 State diagram of cytochrome c in (1-x) water + x 1,4-dioxane solution. The black circles represent Td and Ta. the
triangles and open circles represent 25% and 75% of the transition, respectively. N = native state, DT = thermally denatured
state, FA = fibrous aggregation, Ddio =
dioxane denatured state, and AA = amorphous aggregation.
3 章 1,4-ジオキサン添加によるミオグロビンの凝集体形成の熱力学的および 速度論的研究 3.1 序論 2 章で示したように、基本構造の一つである規則正しい凝集体構造の形成メカニズ ムを明らかにすることは、ミスフォールディング過程の解明につながるとともに、タ ンパク質病の予防[75,76]やタンパク質溶液の長期保存においても重要である。本章で は、1,4-ジオキサンの存在下における示差走査熱量計(DSC)によるミオグロビンの 熱変性および凝集体形成にともなう熱力学量を決定した。溶液中の構造および沈殿し たミオグロビンは CD や動的光散乱(DLS)、走査型電子顕微鏡(SEM)により測定 した。ジオキサンのモル分率、静置時間および圧力が凝集化に及ぼす影響を熱力学的 および速度論的に分析した。ミオグロビンは典型的な球状および単量体であり、規則 正しい凝集体を含む非天然状態に関する多くの報告[19,20]があるため良好なモデル タンパク質である。また、ヘム基(ポルフィリン環)を非共有結合により結合するな ど、シトクロム c と異なる結合様式であり、凝集の構造や形成過程に及ぼすヘム基の 影響を議論する上でもよいモデル系である。また、構造の柔軟性を反映する圧縮率[77] は、ミオグロビンが 8.98×10-5 MPa-1であり、シトクロム c の 0.066×10-5 MPa-1の値よ りも非常に大きく、凝集に対する柔軟性の影響を議論する上でもよいモデル系である。 3.2 実験 3.2.1 試料 ミオグロビンは Sigma-Aldrich から購入したウマ心筋由来のものを用いた。ミオグ ロビンは milli-Q 水で透析した。あらかじめ濃度の濃いミオグロビン溶液を調製し、 吸光度測定により濃度を決定した。1,4-ジオキサン(関東化学、純度 99.9 %)は、モ レキュラーシーブス 4A (キシダ化学, 1/16 ペレット) を、 真空系を用いて 10-3~ 10-2 Pa の高真空でまず室温で 30 分程度排気した後、約 453 K で 8 時間以上排気し

![Fig. 4 Plots of [] 222 as a function of sorbitol concentration at 10 o C (filled circle), 20 o C (filled triangle), 25 o C (filled square), 30 o C (filled inverted triangle), and 35 o C (filled diamond)](https://thumb-ap.123doks.com/thumbv2/123deta/9938324.1390442/15.892.119.796.106.594/plots-function-sorbitol-concentration-triangle-inverted-triangle-diamond.webp)


![Fig. 9 Time dependence of [] 222 of myoglobin at pH jumping from neutral to acidic in the presence of 0 M (A) and 3.3 M (B) sorbitol](https://thumb-ap.123doks.com/thumbv2/123deta/9938324.1390442/24.892.349.770.112.361/time-dependence-myoglobin-jumping-neutral-acidic-presence-sorbitol.webp)

![Table 4 Thermodynamic parameters for thermal denaturation and aggregation of cytochrome c in (1-x) water + x 1, 4-dioxane.(*Comparison with literature value under similar condition (pH 4.6, 40 mM glycine buffer); T d =78.0 o C, ΔH d =440 kJ mol -1 [68]](https://thumb-ap.123doks.com/thumbv2/123deta/9938324.1390442/30.892.182.704.792.1078/thermodynamic-parameters-denaturation-aggregation-cytochrome-comparison-literature-condition.webp)