Application of layer-specific markers in the evaluation of
abnormal cytoarchitecture in the olfactory bulb of pro-
kineticin receptor 2 deficient mice
Chihiro Yamazaki, Mamoru Nagano, Mitsugu Sujino, Toshio Terashima', Hitoshi Ozawa2 and Yasufumi
Atsuko Kubo, Shigeyoshi
Department of Anatomy and Neurobiology, Kinki University Faculty of Medicine, 377-2 Ohno-Higashi, Osakasayama, Osaka 589-8511, Japan
'Division of Anatomy and Developmental Neurobiology, Department of Cell Biology and Physiology, Kobe University Graduate School of Medicine,
7-5-1 Kusunoki-cho, Chuo-ku, Kobe 650-0017, Japan
2Department of Anatomy and Neurobiology, Nippon Medical School, 1-1-5 Sendagi, Bunkyo-ku, Tokyo 113-8602, Japan
Abstract
Prokineticin receptor 2 gene mutant mice (Pkr2-1-) show deformity in the olfactory bulb (OB), they lose axonal invasion from the olfac- tory sensory neurons and olfactory interneurons fail to migrate in the OB. In the present study, we examined whether the mutant has an abnor- mality in the laminar structure in the OB by HE staining and immunohistochemistry using anti- tyrosine hydroxylase (TH), parvalbumin (PV), and reelin (RE) with special reference to the layered structure. In Pkr2-1- mice, TH-im- munoreactive neurons were localized in the marginal area of the OB and the number and size of TH-expressing cells were markedly de- creased. The distribution of PV-immunor-
eactive cells showed no apparent difference but the cells were significantly smaller in the mutant is wild-type mice (WT). RE-immunor- eactive mitral cells showed abnormal alignment, the direction of neurite outgrowth was not well coordinated and they were much smaller than those in WT mice. The present findings suggest that not only the impairment of neuronal migra- tion but also the loss of afferent axonal projec- tion from the olfactory epithelium to the OB arrested the maturation and induced apoptosis of PV-, TH-, and RE-immunoreactive neurons in Pkr2-1-.
Key words : olfactory bulb, prokineticin, tyrosine hydroxylase, parvalbumin, reelin
Introduction
The olfactory bulb (OB) is located in the most rostral region of the brain and is involved in olfaction. The OB consists of two apparently distinct parts. The main olfactory bulb (MOB) occupies a large part of the OB and receives projections from olfactory neurons, receiving most of the odor signals while the accessory olfactory bulb (AOB) senses pheromones, receiv- ing projections from the vomeronasal organ.
The MOB shows apparent layers : olfactory nerve layer (ONL), glomerular layer (GL), exter- nal plexiform layer (EPL), mitral cell layer (MCL), internal plexiform layer (IPL), and gran- ular cell layer (GrL), from the surface to the
center.'
Prokineticin is composed of Pkrl (also named EG-VEGF) and Pk2 (also named Bv8). These are multifunctional secreted proteins with 10 cysteine and a molecular weights of 8kDa.2-4
The receptors for these PKs are Pkrl and Pkr2,
Received August 29, 2011 ; Accepted November 16, 2011
35
both of which are G-protein coupled receptors, mediate signal transduction.2,4 The influences of PKs through the activation of PK receptors (PKRs) on several physiological events in the central and peripheral nervous systems, includ- ing intestinal contraction,5 hyperalgesia,6 sper- matogenesis,7-9 neuronal survival," circadian rhythm,11,12 angiogenesis,1344 ingestive behav- ior," and hematopoiesisl" have been reported.
Recently, the importance of Pk2 and its receptor Pkr2 in the development of the olfactory bulb has been demonstrated using Pk2- and Pkr2- deficient mutant mice. The Pk2-'- mice showed asymmetric deformity in the 0B.'7 Pk2 and Pkr2 mRNA are strongly expressed in the OB."
Pkr2-i- mice showed symmetric OB reduction not only in the embryo but also in adult mice.9 Pkr2-i- mice lack axonal projection from the olfactory neurons to their targets in the MOB and, therefore, lack glomeruli in which axonal terminals of olfactory neurons make synaptic contacts on dendrites of the mitral cells.9 The above results suggest that the Pk2-Pkr2 system plays a crucial role in OB development.
In our present study, we examined whether the layered structure of the MOB is preserved in Pkr2 mutant mice. The MOB is known to have various chemical substances. Many chemical substances mark various types of neurons and layers in the OB. We examined the localization of reelin (RE)," parvalbumin (PV)19'2° and tyrosine hydroxylase (TH)21'22-expressing neur- ons that are confined to one or a few layers in the MOB of adult mice.
Materials and Methods
Pkr2 Gene-disrupted mice
Pkr2 disrupted mice were originally described by Matsumoto et al. in 2006.9 The original genetic background was changed from C57BL/6 strain to ICR strain (CLEA Japan, Tokyo) by 10 times backcross mating with the ICR strain to increase the ratio of the survival of neonates since the ratio of survival with the original C57BL/6 was very low. All experiments were performed in compliance with the regulations of the Animal Ethics Committee of Kinki Univer- sity School of Medicine.
Immunohistochemistry
As the cerebral cortex in Pkr2-'- mice does not have any apparent deformities, we deter- mined one plane by using the position of the
rostral tip of the frontal lobe. The cutting plane is symmetrical in the right-to-left axis, vertical to the basal plane of the mouse brain, and is in tangential contact with the rostral tip of the frontal lobe (Fig. 1). The specificity of the anti- TH, anti-PV, and anti-RE antibodies used in the experiments has been described previously.23,24 Eight-week-old male wild-type mice (WT) and mutant were used for this study. Animals were anesthetized with the ether prior to transcardinal perfusion with 4% paraformaldehyde (PFA).
The brain was removed from the skull, fixed with 4% PFA and then embedded in paraffin.
The paraffin-embedded sections (40 pm) were deparaffinized, and immunostained with mouse anti-TH antibody (1 : 2000 ; Millipore, Gemany), mouse anti-PV antibody (1 : 4000 ; Millipore), and mouse anti-RE (1 : 2500 ; Cal- biochem, Germany) as described below. The sections were activated by citric buffer (10 mM citric acid, pH 6.0) for 20 min at 95°C using a microwave, and then methylated by 3% H202 in 50% methanol for 30 min at 4°C. The sections were immersed in the primary antibodies (TH, PV and RE) overnight at 4°C. Subseqently, the sections were processed according to the avidin- biotin-peroxidase (ABC) method using Vector- stain ABC Kits (Vector Laboratories, Burlin- game, CA, USA) for lh at room temperature.
After rinsing in PBS and 0.05 M Tris-HCI buffer (pH 7.4), antigens were visualized using 0.035%
diaminobenzidine (DAB ; Sigma, St. Louis, MO, USA) in 0.05 M Tris-HC1 buffer, contain- ing 0.01% H2O2 for 5-10 min at room tempera- ture. To identify the cytoarchitecture of the OB, the sections were counterstained with hematox- ylin and eosin (HE).
Quantification of cell size
Images were observed under a microscope (BX43 ; Olympus, Tokyo, Japan) and soze were captured by a CCD camera (DP25 ; Olympus).
Twenty cells were chosen randomly per animal (n=3 in each phenotypes) and the cell body was measured using CellSens software (Olympus).
Results are expressed as the mean ± SEM.
Differences between groups were examined for statistical significance using Student's t-test.
Results
Macroscopic observation The OBs in the coronal rostral end of the cerebral
section cortex,
through as defined
the in
36
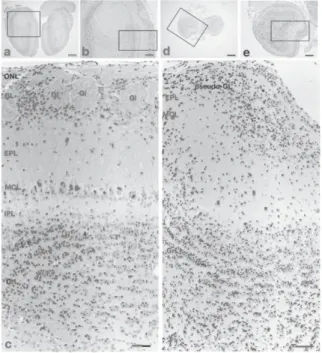
Fig. 1 Schematic view of the olfactory bulb. To compare the cytoarchitectural structure of the olfactory bulb (OB) between wild-type (WT) and Pkr2-'- mice, the reference level is shown by a vertical dashed line.
The reference level is the coronal section through the rostra' end of the cerebral cortex and is vertical to the longitudinal antero-caudal axis of the brain. Other abbreviations : ONL, olfactory nerve layer ; GL, glomerular layer ; EPL, external plexiform layer ; MCL, mitral cell layer ; GrL, granule cell layer ; GI, glomerulus
Fig. 1, were compared between WT and Pkr2-i- mice (see Materials and Methods). All of the OB of WT were oval (Fig. 2a), whereas the OB of Pkr2-i- were rather round (Fig. 2d).
HE staining
In WT mice, the olfactory glomerulus was easily recognized by the oval neutrophil contain- ing a few cell bodies inside and densely packed cell bodies surrounding the vacant structure' (Fig. 2c). The EPL was a cell sparse layer below the glomerular layer and the width of EPL was generally constant in the medial region (Fig. 2a, b, c) of the OB. The MCL was linearly arranged at the boundary between the EPL and the IPL (Fig. 2a, b, c). IPL is a cell-sparse narrow layer localized between the MCL and GrL (Fig. 2c).
In the GrL, small cells were densely distributed, making tightly packed small clusters (Fig. 2c).
In Pkr2-i- mice, the outermost layer of the OB was recognized by the condensation of cell bodies, but the typical vacant oval structure of the olfactory glomerulus was not observed, so it was named the pseudo-glomerular layer (Fig. 2f).
The EPL was also recognized even in the Pkr2-i- mice by its scarcity of cell bodies (Fig.
2f). Beneath the EPL, large clustered cells were present. The triangular shape of the somata of these cells suggests that they were mitral or tufted cells. Assuming that these cells are mitral/tufted
../'•-.:'...'... ..., ...
_... .
..
1(k."
.f;,.
. '
.. .
. . , . ...
. .. ,
a ,-&... - - • _ •Iii: ...-..., .: ,,,,' .. ,:.,* d
-4 •.4_ 1-e-'7.-^F-':.7;.1:...-.rA?:,;:,,(S.1.45.!,
^
f..,,,%1,:tiv‘, , .7S,...-:tr.:;.:4.:e.k. '---.0',..- '4..:,'''''''••• ..,.G. '...,1:-.,-• r.t: '`;'7:.t:i:-...7' ,...:, • .;-°1; 01 .,. A"g^k!"..":;'... Fre` 4.P. - .. 1.: . • • • • .1171%11;. :.:•':•1:7;:i..:7';;;...;.-t.:* ..47...'7.7.:ei:.4,'s.:1::.::i', ....:.1•'' • 'e.; •
,,,.-4';: I- •;';'. .... Y." ..t „•:''''e. .- • .!:': ".,',..' ... '. . lr.v;#1.4.T.7...;.;.::.'. ‘.7*.‘
.i..2; g... ' , f ''' , ' 1 .. ....% :/;fr ' -• • . :, ; ' :; '1 ..:sif:.t.t-i'riti...-•:
;Ept. . - • - : " : In !Z ./ f ' .P 1. ••• 7".... : - : : : l''.' . :...` • ^. :...."•`.:::: r:...., ';':• -,.... ,. *•::;:ii :1'..
, '. 'I . - . ZW...11. ;'..., :!*. .• ''..v.'• ..' A qi: • tl . 7$i . P'...;,.:.'.!..t!' 114 i‘ ',` ^ ' . • • ^ ...!; - - ri i.,..‘ '''...71.'" • • ..-.* 1* f: ; .•10 .: ' .. : .. ..• ' - .. . - ''. ' • '• I•-.44‘ -..' ..- .Fx.,:,•:,,•• ,e-* .. ..., ..:. :"..^.' ; ..! *', ; • 7 , , ' ;" .; ',..., „..-',.: 4.1;,, . : '‘e , , , ,:^• a .''..:, ! , ''. . , .S., ee.;14;.4 . T. 41,;:•••,..:;.'',' ta7.'.)::7!;•, „: 1::', J. , . ••• 4 .•.-r- . ". ,. . ' .I ...• • ,,,- ,I. . ,. .. „ • .. .. - 4 • ... ...
3i....,,,e; 44114,iii;I:"?'t,...'.4e:;t 4 :;...7. y.p,'.k.;;1..r.%,.--,.- ,...v.,.:,.-,;:..?;:,tx:i. ?:
0.,,...,r .„.A,,..:., ,,....k.g....:..::::....f:: ..: :-. ...%7,;...
" . :i 4t:'‘.P,..441'..*--ct:::!..:. 41, f.).11:<*.-.,‘2i.,..:;t4ifi..'!:;.4.4-,,.;:,..,!!,...41:,:::,:. r ,t,...- P...lt . .. '--.•.v4 'f..:...*'...,: :::.,..1
41:44ot; ,.$::,k.,: ...'N.-.'. , 7 • • ..,14. :-.1.4tuilfker...74,.:1.:-L7''''....",!,.,.t.'.`
4.- ,4 ... .,At*!..„•3
:-,,..';.•- „ , - -, .. -z. ,..,...'51",
.!',:".r.,;!"' ge. .P.'1.-,-..-- . 51' ''..is•*-1,;•!::,...e 11;."...%:•...1%,i, '''.'''.7. . 4:4:-= :;... 4;4'4'. ''.**';';..%
• .«,..r.' •,:,.•'• 4'4'
,,..; ... ; ...v.; ei,..1 .„,...i...•• e Z 4' .4,4,,74•.‘t• 4.ii;',:k";•:....?:'sarg-.0M.'":. i: 0.n..:4::,.
-4(e‘''''-.-;-ft,,1-`,..z.se.,...;:t
-....rik.,..•.•:AAV:,,,Se' ...,`":
--, .. -.14,04; "e::'.° ; e••'‘., • ;5*- q.:?..-
• .:,----.;
.1.: ":•-. -.. , --ori - -%....:** ".. • -- : :.„' !.- • L'-• 'v . ; .--,:t`i,r- -,:gy: Z.:- : '., P.--;', l'..,,,i,•
•:',../..---."•tiScv....!-1151...---•:.:;''',...--.'•-'•--.4"....f '
-C••*•:>,,..::.-..:^.•:-:::-.•;..7..,-.,.,...,';'*:•••-•%Vtt:5:;;_..,'..A...`A•"—ri‘f;
Fig. 2 Hematoxylin-eosin (HE)-stained coronal sections of olfactory bulb (OB) of WT (a-c) and Pkr2-i- (d-f) mice.
The rectangles in a and b are enlarged in b and c, respectively, and the rectangles in d and e are enlar-
ged in e and f, respectively. In the OB of the mutant
mouse, neither olfactory nerve layer (ONL) nor
olfactory glomeruli (GL) were identified. Some small cells are scattered in the superficial layer corre-
sponding to ONL, which is named the pseudo-
glomerular layer (Pseudo-GL) in the present study.
Large mitral cells are widely scattered, and therefore
the mitral cell layer (MCL) does not form a unicellar
arrangement as seen in the normal OB. The internal
plexiform layer (IPL) is not identified in the mutant
OB. Other abbreviations : EPL, external plexiform layer ; IPL, internal plexiform layer ; GrL, granule
cell layer ; Gl, glomerulus ; pseudo-GL, pseudo-
glomerular layer. Figure 2c and 2f were rotated 90
degrees to the left from Figure 2b and 2e.
Scale bars, 500,um (a, d) ; 200 Atm (b, e ) ; 50,um (c, f).
cells, cells were smaller than those of WT mice (Fig. 2f). The extension of apical dendrites of mitral/tufted-like cells was not uniform as recog- nized in WT (Fig. 2c, f). Within the clustered mitral cells, there were small neurons packed as in the GrL but the IPL could not be distin- guished (Fig. 20. In the GrL, cells were well packed (Fig. 20 and clusters of cells were also recognized ; however, in Pkr2-1- mice, many neurons were strongly stained with hematoxylin.
Most of the strongly stained neurons were small compared with the granule cells of WT mice.
37
Tyrosine hydroxylase (TH)-immunoreactive neurons
In WT mice, TH-immunoreactive neurons were densely present in the GL, surrounding the glomerulus (Fig. 3a, b) and immunoreactive neural fibers were observed inside the glomer- ulus. The strongly stained neural fibers were thick and well packed (Fig. 3b). In Pkr2-i- mice, a few TH-immunoreactive neurons were observed in pseudo-GL (Fig. 3c, d). The im- munoreactive fibers were not well extended from the cell bodies and were sparse. The number and
.., -
•
, :....
a
ri
.:- ,--:.,..
,.,...,..., . 7 01—, :- ",
PL
MCL
Fig. 3
• •
WI 51.."7, Pt: w. 1 j. rit
^Ite. e", 3
-.5
I it. r
4. 4-4
r t'I
s:.J..0
.
PL
MCL
4
4,1 • ,;*"..4.:
- • ''paecidcizGC.-=--
GrL
•
Tyrosine hydroxylase (TH)-immunoreactive neurons in coronal sections of olfactory bulb
and Pkr2-1- (c, d) mice. Rectangles in a and c are enlarged into b and d, respectively. Black arrow- heads in b and d indicate TH-immunoreactive neur- ons. TH-immunoreactive neurons are distributed in the glomerular layer (GL) of the normal OB (b).
TH-immunopostive neurons located in the shell of the glomerulus elongate their immunopositive den- dritic processes into the core of the glomerulus. In the mutant OB, TH-immunopostive neurons and their neurites are identified in the pseudo-glomerular layer (pseudo-GL) which appears to correspond to the GL of the normal OB. TH-immunopositive neurons are apparently reduced in size in the mutant OB compared with their normal counterparts. Other abbreviations : EPL, external plexiform layer ; GrL, granule cell layer. Scale bars, 100p m (a, c) ; 50 At m (b, d).
:41
•
•
•
^^^•.
active neurons
in a and c are Black arrow-
size of TH-immunoreactive neurons were appar- ently decreased (Fig. 3a, b, c, d).
Parvalbumin (PV)-immunoreactive neurons In the MOB of WT mice, PV-immunoreactive neurons were present in the EPL as reported previously'9•20 (Fig. 4a, b). A small number of weak PV-immunoreactive neurites were observed in the EPL (Fig. 4b). In Pkr2-1- mice, PV-im- munoreactive neurons and neurites were also present in the EPL (Fig. 4c, d).
Reelin (RE)-immunoreactive neurons
In WT mice, RE was expressed by mitral cells in the MCL, tufted cells in the EPL and some
IVY
GL
EPL
MCL
GrL
Fig. 4
6
: 061
ti
M.
04*
EPL iVICL
k.
GrL
• .•
3 ft eL e
MCL • —0.. • • .
4"11
—^•
, r GrL
•
.a1M? 'SULU!: --me Parvalbumin (PV)-immunoreactive neurons in coro- nal sections of olfactory bulb in WT (a, b) and Pkr2 (c, d) mice. The rectangles in a and c are enlarged in b and d, respectively. The Black arrows in b and d indicate PV-immunoreactive neurons. In the normal OB (b), PV-immunoreactive neurons are distributed in the external plexiform layer (EPL). In the mutant OB (d), PV-immunopositive neurons are mainly distributed in the EPL, as in their case of the normal counterparts, but some PV-immunopositive neurons are identified among neurons in the lower third of the pseudo-glomerular layer (pseudo-GL).
PV-immunopositive neurons are apparently reduced in size in the mutant compared with the normal counterpart. Other abbrevisitons : GL, glomerular layer ; MCL, mitral cell layer ; IPL, internal plex- iform layer ; GrL, granule cell layer. Scale bars, 100 pm (a, c) ; 50 p m (b, d).
38
subsets of periglomerular neurons at the bottom of the GL (Fig. 5a, b). Mitral and tufted cells showed moderate to strong staining, but perig- lomerular cells showed moderate staining. In WT, RE-immunoreactive neurons were arranged in a regular row, and the neurite outgrowth has rather consistent direction (Fig. 5a, b, c). In
a t
•- .•
41.
4 , 4 4
,
EPL i
11P , tS1'
;.C.,-*. • ,...•
..C': •
, :.!'..j.:',:;•,
,.., •
..o . *
.
a.
C 01 Fig. 5 -.-
.;
trt 010
• •
'it":4".4
•
Ot‘i
• EPL • •
•
pegaudo-G
INCL • • c - •)
tor • ( • ‘;'/'
. .
••
.
• \ - • .
. 11
• ;7' i
't? •
•
- •
•
\ • 10 -*
c1/4 •
•
a NI
Nre.
• V‘. *U. u "
•
•
Grt
•
— e
***1
ere , aliPs
e-r4 . 7 • I Distribution of reelin (RE)-immunoreactive neurons in coronal sections of olfactory bulb (OB) in WT (a, b, c) and Pkr2-i- (d, e, f) mice. Rectangles in a and d are enlarged in b and e, respectively. And, rectan- gles in b and e are enlarged in c and f, respectively.
The black arrows indicate RE-immunoreactive mitral cells, and black arrowheads indicate RE-immunor- eactive periglomerular neurons. The white arrows and arrowheads indicate the neurites. In the normal OB, RE-immunopositive mitral cells are arranged in a laminar fashion and showed consistent direction of neurite outgrowth, whereas in the mutant OB they do not show any laminar fashion or the consistency in the direction of neurite outgrowth. Other abbrevia- tions : GL, glomerular layer ; EPL, external plex- iform layer MCL, mitral cell layer ; IPL, internal plexiform layer ; GrL, granule cell layer ; pseudo- GL, pseudo-glomerular layer. Figure 5b and 5e were rotated 90 degrees to the left from Figure 5a and 5d.
Figure 5c and 5f were rotated 90 degrees to the right from Figure 5b and 5e. Scale bars, 200 p m (a, d) ; 50 pm (b, c, e, 0.
Pkr2-i- mice, RE-immunoreactive neurons were also localized in both EPL and MCL but the linear arrangement of the RE-immunoreactive neurons and the consistency in the direction of neurite outgrowth was disrupted (Fig. 5c, d, e).
Quantitative analysis of cell sizes
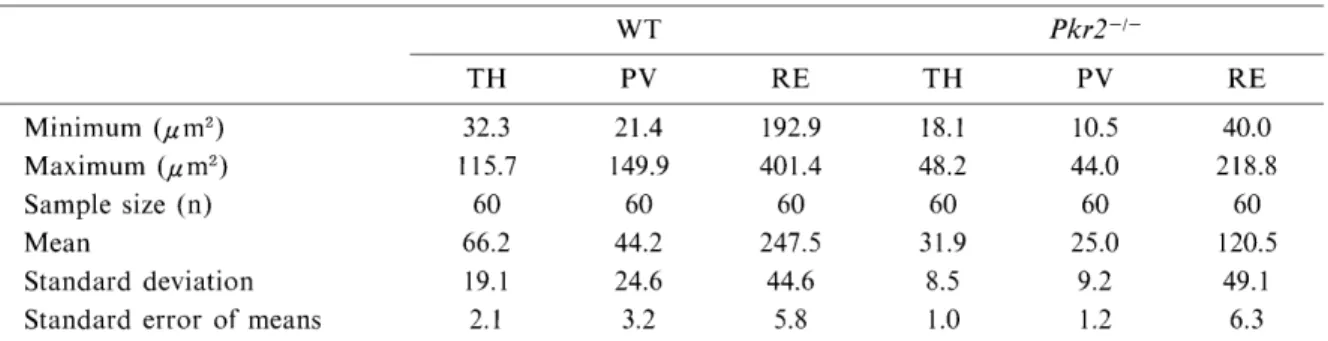
We measured the cell bodies of TH-, PV- and RE-immunopositive neurons in the OB of nor- mal and mutant mice and found that, in all immunoreactive neurons examined, the cell bodies were smaller in Pkr2-1- mice than in their normal counterparts. The mean size of TH-im- munoreactive cell bodies located in the GL was 31.9± 1.0 du m2 (Mean SEM) in Pkr2-1-, signifi- cantly smaller than those (66.2 ± 2.1 ,u m2) of WT
(Student's t test ; p< 0.001). PA-immunor- eactive neurons in the EPL were also significant- ly smaller (p< 0.001) in Pkr2-i- (25± 1.2 ,u m2) than in WT (44.2± 3.2 ,u m2). RE was expressed
in mitral cells, tufted cells and periglomerular cells, as described above, which have apparently different mean sizes of cell bodies. Therefore, we tried to identify mitral cells and to measure the cell bodies. Even in Pkr2-1-, the MCL was distinct from the EPL and pseudo-glomerular layer so that we were able to differentiate mitral cells by their specific location in the MCL and large cell bodies from other RE-immunopostive neurons. We measured the mean cell bodies of RE-immunopositive mitral cells in WT (247.5+
5.8) and in Pkr2-1 (120.5±6.3), suggesting that
Fig. 6
WT PKr2-/-
TH PV RE
The diagrams show the cell body size of tyrosine hydroxide (TH)-, parvalbumin (PV)- and reelin (RE)-immunopositive neurons in WT (gray bar) and Pkr2-i- (open bar) mice.
Cell bodies size of 60 immunopositive neurons (20 neurons per slice ; n=3 for each phenotypes) were measured for each primary antibody. Values are the represent means±SE (n=3). TH-, PV-, and RE- immunopositive neurons were statistically smaller in the mutant OB compared with their normal counter- parts (**p<0.001).
39
Table 1 The size of cell somata of TH-, PV-, and RE-immunopositive neurons in the OB of the normal (WT) and Pkr2-deficinet mutant mice (Pkr2-1-)
WT Pkr2-1-
TH PV RE TH PV RE
Minimum (pm') Maximum (pm') Sample size (n) Mean
Standard deviation Standard error of means
32.3 115.7
60 66.2 19.1 2.1
21.4 149.9
60 44.2 24.6 3.2
192.9 401.4 60 247.5
44.6 5.8
18.1 48.2 60 31.9 8.5 1.0
10.5 44.0 60 25.0 9.2 1.2
40.0 218.8 60 120.5
49.1 6.3
cell bodies of RE-immunopostitive mitral cells were also significantly smaller in Pkr2-1- than in the WT.
Discussion
The present HE-stained sections demonstrated that mutation of the Pkr2 gene affects the laminar structures of the OB. The glomerulus was almost completely lost and mitral cells were not aligned in a row. In the GrL, chromatin condensation in the nuclei of granule cells sug- gests that many granule cells in the OB of Pkr2-1- mice undergo apoptosis. This is also supported by previous studies that demonstrated an increased number of apoptotic neurons by terminal deoxynucleotidyl transferase- mediated- deoxyuridine triphosphate UTP nick-end labe- ling (TUNEL) analysis.'7'25 Granule cells in the GrL of the OB make a dendro-dendritic synaptic connection with dendrites of mitral and tufted cells,26 therefore, it is possible that the impair- ment of granule cells leads to impaired dendro- dendritic synaptic formation between granule cells and mitral cells, and triggers the cell death of granule cells. Therefore, the decreased volume of OB in Pkr2-1- mice could be attributed not only to the erroneous migration of interneur- ons26 but also to the increased number of apoptotic cell deaths of interneurons.
TH, a marker enzyme of dopaminergic neur- ons, is expressed by periglomerular neurons and external tufted cells situated in GL in the OB.27-29 In WT mice, strong TH-immunor- eactive neurons were confined to the GL and TH-immunoreactive nerve fibers were clustered thickly in the glomerulus. The glomerulus was absent in the Pkr2-1- but a few TH-immunor- eactive neurons were present in the pseudo-GL.
Interestingly, TH-immunoreactive nerve fibers were also observed only in the pseudo-GL. This
finding suggests that Pkr2 gene disruption does not affect the migration of TH-immunoreactive periglomerular neurons from their origin in the subventricular zone (SVZ) of the lateral ventricle to their final position, although we could not
exclude the possibility that dopaminergic neur- ons that succeed in migrating to their final position can express TH immunoreactivity. PV, which is one of the neurochemical markers, is calcium-binding protein, and is expressed by neurons in EPL of the mice MOB. The present study has revealed that PV-immunoreactive neur- ons were distributed in the EPL in both WT mice and Pkr2-1- mice. This finding in WT mice is consistent with previous studies.'°"° PV- immunoreactive multipolar neurons, considered to be interneurons, may be generated in the SVZ,
do not have any axons and make synaptic con- tacts with dendrites of mitral cells.29 In the
present study, Pkr2 disruption did not seem to affect the distribution and cell number of PV- immunoreactive neurons, however, in the OB of Pkr2-1- mice, the cell size of PV-immunoreactive neurons was much reduced. These data suggest that Pkr2 gene disruption does not influence the migration of PV-immunoreactive neurons to EPL,29 but affects the maturation process of neurons, including enlargement of the cell size.
PV-immunoreactive neurons have been reported to make synaptic contact with basal dendrites of
mitral cells ;29'35 therefore, the maturation proc- ess of PV neurons may be caused by the absence of normal retrograde factor from mitral cells.
Since mitral cells appear to be abnormal in Pkr2-1-, it is possible that impaired synapse formation of PV neurons with mitral cells may affect the maturation process of PV neurons.
RE, the gene responsible for the reeler muta- tion, is a large extracellular matrix protein, ex-
pressed strongly by external tufted and mitral cells, and weakly in periglomerular cells in
40
PGL.32 In Pkr2-1 mice, we cannot distinguish mitral cells from tufted cells since the alignment of mitral cells is not clear, being scattered in a wider area, and the border between EPL and MCL is ambiguous. The findings suggest that synaptic formation of olfactory nerves with mitral cells is essential for the unicellular align- ment of mitral cells. In addition, the present study clarified that tufted/mitral cells extend their neurites in almost random directions instead of a uniform direction, suggesting that the neurite outgrowth of tufted/mitral cells is regulated by synaptic contact between olfactory nerve endings and mitral cells. Some of the morphological phenotypes in Pkr2-1- mice are very similar to those of other mutant mice that lack axonal projection from the olfactory neur- ons. In fact, Arx,33 D1x533 and fez' (fez)35,38 mutant mice share abnormal features similar to those of Pkr2-1- mice. In these mutants, the ONL is absent due to the lack of projections from the olfactory neurons to the OB. The olfactory bulb is very small and the glomerulus is almost completely lost. The mitral cell layer is widened and mitral/tufted neurons marked by reelin expression are widely dispersed around the border of EPL and GrL. IPL is absent, possibly because of defects of the axonal extension of mitral cells. In contrast, interneurons in the OB are not necessary to arrange mitral cells in the OB. Dxll/D1x2 double knockout mice show a complete absence of GABAergic interneurons, including periglomerular cells and granular cells. The olfactory nerve layer was arranged neatly. Mitral cells did not show abnormal distribution,37 therefore, although impaired migration of olfactory interneurons of the OB also has been reported in Pkr2-i- mice,'2'17 it is probable that the disorder of the layered struc- ture is attributed to the loss of axonal projection from olfactory neurons. As Pk2-deficient mutant mice also show a deformity of the olfactory bulb, similar to those in Pkr2-1- mice," it is highly probable that the PK2 and Pkr2 ligand-receptor system is essential to make a normal layered structure. Thus, in mutants that show similar disorders, as shown in Pk2 and Pkr2 mutant mice, loss of axonal invasion may be the first event which causes the broad spectrum of abnor- mality in the olfactory bulb.
The reduction in the size of the neurons in Pkr2-1- mice observed in the present study could be attributed both to immature development of
neurons and the apoptosis caused by impaired synaptic connection of the neurons. In the developing process of the neuron, the size of cell bodies of the neurons increases associated with synapse formation. And, moreover, the size of the neurons decreases in the process of apoptosis.38 Therefore, it is highly probable that the size reduction of the cell bodies was caused
both by impaired development and apoptosis.
We will further perform studies to investigate the maturation process of PV, RE, and TH-express- ing neurons in future.
Conflict of Interest Statement
None of the authors has a financial relation- ship with a commercial entity that has an interest in the subject of this manuscript
Acknowledgements
We thank Dr. Shun-Ichiro Matsumoto for helpful dis- cussion and Ms. Junko Kawano, Mr. Yutaka Funabashi,
Ms. Toshiko Egami, Ms. Mieko Kashima, and Ms. Yayoi Yamanaka for their skillful technical assistance. This research was supported by a Grant-in-Aid for Scientific Research from the Japanese Ministry of Education, Sci- ence, Sports and Culture of Japan.
References
1. Switzer C, R Olmos Jd, Heimer L, 1.Olfactory sys- tem, in The rat nervous system, G. Paxinos, Editor 1985, Academic press Australia. p. 1-36
2. Li M, Bullock CM, Knauer DJ, Ehlert FJ, Zhou QY (2001) Identification of two prokineticin cDNAs : recombinant proteins potently contract gastrointestinal smooth muscle. Mol Pharmacol 59 : 692-698 3. Negri L, Lattanzi R, Giannini E, Melchiorri P (2007)
Bv8/Prokineticin proteins and their receptors. Life Sci 81 : 1103-1116
4. Soga T, Matsumoto S, Oda T, Saito T, Hiyama H, et al. (2002) Molecular cloning and characterization of prokineticin receptors. Biochim Biophys Acta 1579 : 173-179
5. Wade PR, Palmer JM, Mabus J, Saunders PR, Prouty S, et al. (2010) Prokineticin-1 evokes secretory and contractile activity in rat small intestine. Neur- ogastroenterol Motil 22: e152-161
6. Hu WP, Zhang C, Li JD, Luo ZD, Amadesi S, et al.
(2006) Impaired pain sensation in mice lacking pro- kineticin 2. Mol Pain 2 : 35
7. Maldonado-Perez D, Evans J, Denison F, Millar RP, Jabbour HN (2007) Potential roles of the prokineticins in reproduction. Trends Endocrinol Metab 18 : 66-72 8. Sinisi AA, Asci R, Bellastella G, Maione L, Esposito D, et al. (2008) Homozygous mutation in the pro-
41
kineticin-receptor2 gene (Va1274Asp) presenting as reversible Kallmann syndrome and persistent olig- ozoospermia : case report. Hum Reprod 23 : 2380- 2384
9. Matsumoto S, Yamazaki C, Masumoto KH, Nagano M, Naito M, et al. (2006) Abnormal development of the olfactory bulb and reproductive system in mice lacking prokineticin receptor PKR2. Proc Natl Acad Sci U S A 103: 4140-4145
10. Melchiorri D, Bruno V, Besong G, Ngomba RT, Cuomo L, et al. (2001) The mammalian homologue of the novel peptide Bv8 is expressed in the central ner- vous system and supports neuronal survival by activat- ing the MAP kinase/PI-3-kinase pathways. Eur J Neurosci 13 : 1694-1702
11. Cheng MY, Bullock CM, Li C, Lee AG, Bermak JC, et al. (2002) Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus.
Nature 417: 405-410
12. Masumoto KH, Nagano M, Takashima N, Hayasaka N, Hiyama H, et al. (2006) Distinct localization of prokineticin 2 and prokineticin receptor 2 mRNAs in the rat suprachiasmatic nucleus. Eur J Neurosci 23 : 2959-2970
13. Kisliouk T, Lew N, Hurwitz A, Meidan R (2003) Presence and regulation of endocrine gland vascular endothelial growth factor/prokineticin-1 and its rece- ptors in ovarian cells. J Clin Endocrinol Metab 88 : 3700-3707
14. Monnier J, Samson M (2010) Prokineticins in an- giogenesis and cancer. Cancer Lett 296: 144-149 15. Negri L, Lattanzi R, Giannini E, Felice M De,
Colucci A, et al. (2004) Bv8, the amphibian homologue of the mammalian prokineticins, modulates ingestive behaviour in rats. Br J Pharmacol 142: 181-191 16. Soderhall I, Kim YA, Jiravanichpaisal P, Lee SY,
Soderhall K (2005) An ancient role for a prokineticin domain in invertebrate hematopoiesis. J Immunol 174 : 6153-6160
17. KL Ng , Li JD, Cheng MY, Leslie FM, Lee AG, Zhou QY (2005) Dependence of olfactory bulb neur- ogenesis on prokineticin 2 signaling. Science 308:
1923-1927
18. Pappas GD, Kriho V, Liu WS, Tremolizzo L, Lugli G, et al. (2003) Immunocytochemical localization of reelin in the olfactory bulb of the heterozygous reeler mouse : an animal model for schizophrenia. Neurol Res 25 : 819-830
19. Qin ZP, Ye SM, Du JZ, Shen GY (2005) Postnatal developmental expression of calbindin, calretinin and parvalbumin in mouse main olfactory bulb. Acta Biochim Biophys Sin (Shanghai) 37 : 276-282 20. Kosaka T, Kosaka K (2008) Heterogeneity of parval-
bumin-containing neurons in the mouse main olfactory bulb, with special reference to short-axon cells and betalV-spectrin positive dendritic segments. Neurosci Res 60 : 56-72
21. Baker H, Towle AC, Margolis FL (1988) Differential afferent regulation of dopaminergic and GABAergic
neurons in the mouse main olfactory bulb. Brain Res 450 : 69-80
22. Foster GA, Schultzberg M, Kokfelt T, Goldstein M, Hemmings HC, et al. (1988) Ontogeny of the dopamine
and cyclic adenosine-3' : 5'-monophosphate-regulated phosphoprotein (DARPP-32) in the pre- and postnatal mouse central nervous system. Int J Dev Neurosci 6 : 367-386
23. Okuyama-Yamamoto A, Yamamoto T, Miki A, Terashima T (2005) Changes in reelin expression in the mouse olfactory bulb after chemical lesion to the olfactory epithelium. Eur J Neurosci 21 : 2586-2592 24. Yang Z (2008) Postnatal subventricular zone progen-
itors give rise not only to granular and periglomerular interneurons but also to interneurons in the external plexiform layer of the rat olfactory bulb. J Comp Neurol 506: 347-358
25. Prosser HM, Bradley A, Caldwell MA (2007) Olfac- tory bulb hypoplasia in Prokr2 null mice stems from defective neuronal progenitor migration and differenti- ation. Eur J Neurosci 26 : 3339-3344
26. Hinds JW, Hinds PL (1976) Synapse formation in the mouse olfactory bulb. I. Quantitative studies. J Comp Neurol 169: 15-40
27. Halasz N, Johansson 0, Hokfelt T, Ljungdahl A, Goldstein M (1981) Immunohistochemical identifica- tion of two types of dopamine neuron in the rat olfac- tory bulb as seen by serial sectioning. J Neurocytol 10 : 251-259
28. Mugnaini E, Oertel WH, Wouterlood FF (1984) Immunocytochemical localization of GABA neurons and dopamine neurons in the rat main and accessory olfactory bulbs. Neurosci Lett 47 : 221-226
29. Kosaka T, Kosaka K (2011) "Interneurons" in the olfactory bulb revisited. Neurosci Res 69 : 93-99 30. Ohm TG, Muller H, Ulfig N, Braak E (1990)
Glutamic-acid-decarboxylase-and parvalbumin-like- immunoreactive structures in the olfactory bulb of the human adult. J Comp Neurol 291 : 1-8
31. Toida K, Kosaka K, Heizmann CW, Kosaka T (1994) Synaptic contacts between mitral/tufted cells and GABAergic neurons containing calcium-binding protein parvalbumin in the rat olfactory bulb, with special reference to reciprocal synapses between them.
Brain Res 650: 347-352
32. Ikeda Y, Terashima T (1997) Expression of reelin, the gene responsible for the reeler mutation, in embry- onic development and adulthood in the mouse. Dev Dyn 210: 157-172
33. Yoshihara S, Omichi K, Yanazawa M, Kitamura K, Yoshihara Y (2005) Arx homeobox gene is essential for development of mouse olfactory system. Development 132: 751-762
34. Long JE, Garel S, Depew MJ, Tobet S, Rubenstein JL (2003) DLX5 regulates development of peripheral and central components of the olfactory system. J Neurosci 23 : 568-578
35. Hirata T, Nakazawa M, Muraoka 0, Nakayama R, Suda Y, M Hibi (2006) Zinc-finger genes Fez and Fez-
42
like function in the establishment of diencephalon subdivisions. Development 133: 3993-4004
36. Watanabe Y, Inoue K, Okuyama-Yamamoto A, Nakai N, Nakatani J, et al. (2009) Fezfl is required for penetration of the basal lamina by olfactory axons to promote olfactory development. J Comp Neurol 515 : 565-584
37. Bulfone A, Wang F, Hevner R, Anderson S, Cutforth
T, et al. (1998) An olfactory sensory map develops in the absence of normal projection neurons or GABAer- gic interneurons. Neuron 21 : 1273-1282
38. Kerr JF, Wyllie AH, Currie AR (1972) Apoptosis : a basic biological phenomenon with wide-ranging implication on tissue kinetics. Br J Cancer 26 : 239- 257