その役割に関する研究
大森, 俊郎
https://doi.org/10.11501/3099962
出版情報:Kyushu University, 1994, 博士(農学), 論文博士 バージョン:
権利関係:
第3章 全麹大麦焼酎もろみの特徴
3 - 1 緒言
前章で, もろみ中のグリセリンの生成源について 検討し, 大麦焼酎もろみ中 のグリセリンが酵母および大麦麹に由来すること, 主原料にも麹を使用した全 麹のもろみに おいて酵母が生成するグリセリンは一般的な仕込み配合である麹 歩合33%よりも重量比で約250/0増加することを述べた。
焼酎製造における麹の役割は主に酵素およびクエン酸の供給源として考えら れている7 3 )。 焼酎麹にはα-アミラーゼ グルコアミラーゼおよび酸性プロ
テアーゼなどの醸造に 必須の酵素が含まれる。 米麹は原料の溶解に必要十分量 の酵素を含み28), 米焼酎においては麹使用量を減らす方法や麹を使用せず,
酵素剤のみで糖化し, 発酵する製造方法も検討されている74)。 一方, 大麦焼 酎もろみは大麦の溶解性が悪いこともあり 大 麦からのグルコースの生成が発 酵を律速している28 )と報告されている。 そのため 使用原料に占める麹の割 合は大麦焼酎の方が高く, 一般に大麦焼酎は麹歩合30--- 330/0, 米焼酎は麹歩合 25"-'30%で製造されている75 )。 全麹仕込み では 麹中の不飽和脂肪酸に よ り酵母のalcohol acetyl transferase活性が低下し, 酵母によるエステル生成が抑 制され9), 麹の風味が強い焼酎となる75 ) 。 また, 原料中のでんぷんが麹菌 によって消費されるため原料当たりのアルコール収量が低下すること, あるい は製麹する手聞が加わることなどによるコスト高で, 全麹仕込みは一般にはほ
る必要がある。 そこで, 本章では大麦麹中のグリセリン動向, 全麹もろみの特 徴について検討 した結果を述べる。
3-2 実験方法
原料大麦
第1章1-2に示 した。
大麦麹
第2章2 -2に示 した。
酵素の抽出および活性測定
酵素の抽出およびトアミラーゼ活性の測定は第2章2-2で示 した方法に従っ て行った。
グルコアミラーゼ(GAase)活性の測定7 6 ) は次のように行った。すなわち,
2%可溶性でんぷん(Merck)溶液5mlにO.2M酢酸緩衝液(pH5.0) 1mlを加え,
5分予熱後, 粗酵素液O.5mlを添加し, 400Cで20分間インキユベートした。 こ の後, 2Mの水酸化ナトリウムO.5mlを加えて, 反応、を停止させ, 室温で約30分 間放置後, 2Mの塩酸0.5mlで中和した。 反応液中のグルコース濃度はグルコー
スBーテストワコー(和光純薬工業)を用いて, 測定した。 また, ブランクは
2Mの水酸化ナトリウムを添加した後に粗酵素液を加え, 以下同様に操作した。
可溶性デンプンから400Cで60分間に1mgのグルコース を生成する 活性を1unitと
し, 大麦麹19当たりのグルコアミラーゼ活性は(3-1)式に従って求めた。
60 . _ 1
.. 100
GPase ( unit/g-koji) =生成 グルコース量(mg)Xτz×τγ×τz×T555 (3-1) 酸性プロテアーゼ(APase)活性の測定7 7 ) は次のように行った。 すなわち,
2%カゼイン (関東化学)溶液1.5mlにMcIlvaine緩衝液(pH3.0 )1ml を加え,
380Cで5分間予熱後, 5倍に希釈した粗酵素液0.5mlを加え, 同温度で60分間反応
させたo 0.4Mトリクロロ酢酸(TCA)溶液を3m l添加して反応を停止させ,
JIS5種Aろ紙で沈殿をろ別した。 さら に, 0.4M炭酸ナトリウム5mlとタンパク 質測定用フェノール試薬(関東化学)1mlにろ液0.5ml, 脱イオン水0.5mlを添加 し, 380Cで3 0分間反応させ 660nmの吸光度 を求めた。 ブランクはTCA 溶液を 添加した後に粗酵素液を加え, 以下同様に操作した。 プロテアーゼの作用に よ
り遊離したアミノ酸量をチロシンとして定量するために, サンプルとブランク との吸光度差を求め, 予め求めたチロシン量と660nmにおける吸光度との関係
から, 生成チロシン量(μg)を求めた。酸性プロテアーゼ 活性は4 00Cで60分間 にlμgのチロシン相当 量の呈色を示す活性を1unitとし, 大麦麹19当たりの活性
は(3-2)式に従って求めた。
APase (unit/g北oji) ==生成チロシン量(μg)Xとx.2一x !旦X1 0.5 20 5 ( 3-2) また, カゼイン溶液は次のように調製した。 すなわち, カゼイン2gを50mlの
た方法で行った。
大麦麹の脂肪酸
大麦麹 50gをエーテルで24時間抽出し, 抽出した粗脂肪に内標準としてヘプ タデカン酸10mgを加え, 減圧濃縮し, 濃縮液中の遊離脂肪酸をGCに供した。
GC分析はキャピラリーガスクロマトグラフ(Perkin Elmer, 8500型)で行った。
カラムはNukol(ゆ0.53mmX15m, 膜厚0.5μm, Supelco)を使用し, カラム温度 は1100C(1分保持)から80C/minで2100Cまで昇温させ, 17分保持するプログラ ムで行った。 注入口 検出器温度はそれぞれ2500Cとし 検出器はFIDを使用し た。キャリアーガスはヘリウムガスを使用し 流速1.6ml/min, スプリット比 10:1, サンプル注入量0.2μlとした。
大麦麹の麹菌体量
大麦麹中の麹菌体量は大内ら7 8 ) の方法を一部改変して行ったO すなわち,
大麦麹 19に脱イオン水1mlを加え, 沸騰水中 で30分保持し, 冷却後, 30秒間 ホモジナイズした。 さらに, アマノ2(天野製薬, 焼酎醸造用市販酵素剤)を 20mg添加し, 550Cで3時間振とう処理した後, 3,000rpmで10分間遠心分離し?
沈殿物をさらに 脱イオン 水で3回洗浄し た 。 沈殿物にO.lM酢酸 緩 衝 液
(pH5.0) 10mlとウスキザイム(和光純薬, 細胞壁溶解酵素)30mgを加え,
370Cで3時間振とう処理し, 3,000rpmで"10分間遠心分離後, 上澄液を0.8μmフィ
ルターでろ過し, N-acetylglucosamine比色定量試料とした。
N -acety 19l uc os ami ne比色定量法はReissigら79) の方法に準じて行ったO す なわち, 試料0.5mlに0.8Mホウ酸カリウム溶液O.lmlを加え, 激しく沸騰し ている水浴 中で正確に 3分間加熱し た。 ただちに, 冷却後, 予め用意した p-dimethylaminobenzaldehyde (DMAB)試薬を3ml加え, 十分にかくはん後 38�
39tの水浴中に浸し 正確に20分放置後, 585nmの吸光度を測定した。 DMAB 試薬は次のように調製し た。 すなわち, 10Mの塩酸を12.50/0 (v/v)含む氷 酢酸100mlにDMABIOgを溶解し, DMAB試薬とした。 検量線作成のため使用し た菌体は, Czapek培地で300C, 20日間液体培養したAspergillus kawachiiの菌体 をアセトン乾燥させたものを使用した。
酵母
第2章2-2に示した大麦焼酎用酵母BAW-6を使用した。
小仕込試験
麹歩合330/0および全麹の 仕込み配合(第2章Table2-1)で大麦焼酎もろみを 製造した。
もろみ中の脂肪酸
65%精麦を用い, 総原料60g (麹原料20g, 主原料40g), 汲水90mlで仕込ん
した。GC分析は大麦麹中の脂肪酸の分析と同一条件で行った。
酵母の菌体内脂肪酸
発酵終了後 の大麦焼酎もろみを遠心分離(1,500rpm, 10分間)し, 固形物を 除去した後, さらに顕微鏡観察でデンプン粒が観察されなくなるまで脱イオン 水での洗浄, 遠心分離(3,000rpm, 10分間)を繰り返し, 酵母を集菌した。 集 菌酵母を脱イオン水で3回洗浄し, 酵母に 付着した脂肪酸を除去するためにア セトンで3回洗浄後, さらに石油エーテル:ジエチルエーテル(2:1)で3回洗浄 し, 凍結乾燥した。 凍結乾燥酵母5mgをアンプル管に取り, 50/0塩化水素/メタ ノーjレ溶液0.5mlを用い 減圧下で1100Cで3時間メチル化した。 得られた脂肪酸 メチルにヘプタデカン酸メチル1ml(lg/ l)と水1mlを加え, さらに, 石油エー テル3mlを加えて 脂肪酸メチルを抽出した。 抽出液は減圧下で乾固させた後,
ヘキサン50μlに溶解し, GCに供した。
GC分析はFID検出器付きのガスクロマトグラフ(目立, 263-80型)を用いて 行った。 カラムはG-525(ゆ1.2mmX40m, 膜厚2μm, 化学品検査協会)を使用
した。 カラム温度は1500C(5分間保持)から50C/minで2000Cまで昇温させ,
15分保持するプログラムで, 注入口, 検出器温度はそれぞれ2500Cで行った。
キャリアーガスはヘリウムを用い, 流速4Oml/min, サンプル注入量は1μlとした。
盆笠
大麦麹中のグリセリン, グルコースおよびクエン酸の定量は次のように行っ たOすなわち, 大麦麹50gに脱イオン水100mlを加えて, ホモジナイズ後,
0.8ドmのフィルターでろ過し, HPLC分析に供した。 グリセリン, グルコースの
測定は第1章1 - 2と同様に行った。 また, クエン酸の定量は グリセリン, グ ルコースと同じ分離条件で, 検出器は紫外分光検出器(Waters, UV48 4)を用
い, 210nmで 測定した。
エタノールはアルコール分析計(BTIC, AB-1B) , 密度は密度計(京都電子 工業, DA300)でそれぞれ分析を行った。
もろみ1ml当たりの酵母の生菌数はサンプルを適宜希釈し, 0.15%酵母エキス,
0.2%ポリペプトン 10/0グルコース 0.1%リン酸2 水素lカリウム, 0.040/0硫酸マ グネシウム7水和物および20/0寒天のプレート培地に埋包し 300Cで48時間培養 後,得られたコロニー数から求めた。 もろみ中の酵母の全生菌数は, 遠心分離 により求めたもろみの液相量にもろみ1ml当たりの生菌数 を乗じて求めた。 出 麹酸度は国税庁所定分析法注解8 0 ) に従って測定した。 麹の水分は麹10gを ンヤーレに正確に秤量し, 1100Cで24時間乾燥させ 乾燥前後の値から求めたO
のα-アミラーゼ, グルコアミラーゼ\酸性フ。ロテアーゼ、およびリバーゼの活性
はそれぞれ94, 132, 1 9,500および28unit/g-dried kojiであった。 また, 大麦麹中 のクエン酸, グルコースおよびグリセリンの濃度はそれぞれ1.78, 4.61および O.95g/100g-dried kojiであった。
清酒米麹8 1 )あるいは焼酎破砕米麹1 0 )では製麹中に麹中の脂肪酸が顕著 に増加し, 脂肪酸が酵母の生理, 代謝に影響する 8 2 -8 4 )ことが報告されてい る。しかしながら 大麦麹の脂肪酸に関する 報告はないこと から, 大 麦麹の 脂肪酸を測定した。 その結果をTable3-2に示す。 リノール酸 は大麦麹中の遊離 高級脂肪酸の約60%を占め, 乾燥大麦麹100g当たり 229mgで、あった。 次いで,
オレイン酸, パルミチン酸の順番であった。 一般的な麹歩合330/0仕込みでは主 原料に大麦を使用し 全麹仕込みでは主原料に大麦麹を使用する 。 このことか
ら, 麹歩合の違いはもろみ中のリノール酸やオレイン酸などの不飽和脂肪酸濃 度に大きな違いをもたらすことが示唆された。
3-3-2 製麹中の成分変化
a. 大麦麹中のグリセリンとクエン酸
Table 3-1. General analysis of barley-koji.
α-Amylase 94 unitJg-dried koji Glucoamylase 132 unitJg-dlied koji Acid protease 19,500 unit/g-dried koji
Lipase 28 unitJg-dlied koji
Citric acid 1. 78 g/100g-dried koji Glucose 4.61 g/lOOg-dried koji Glycerol 0.95 g/lOOg-dlied koji
Moisture 27.5 %
These data were the mean value of ten samples
determined. Barley koji was prepared according to the ordinary method.
Table 3-2. Amounts (mg) and composition (%) of fatty acids of barley and barley北oji.
Materials
会æ��_lg_(!f_ f�t_ty _号�_i_q�_(,!叫 99_�-?!!�_s�p!�)_ _a
Composition of fatty acids (%)
C12:0 C14:0 C16:0 C16:1 C18:0 C18:1 C18:2 C18:3 Total U/S ratioc
Barley NDb 0.15 15.16 ND 1.62 3.38 25.94 Trace 46.24
ーーー苧ーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーーー喧骨骨ーー_ -
0.3 32.8 3.5 7.3 56.1 100 1.73
ND 0.59 59.95 ND 5.34 72.59 228.68 20.97 388.12
Barley-k�ii __.l_一一一一 一一一一一一一一一一一一一一一一一一一一一一一一一一一一一 一 4.88
0.2 15.4 - 1.4 18.7 58.9 5.4 100
a Abbreviations: CI2:0, lauric acid; C14:0, myristic acid; C16:0, palmitic acid; CI6:1, palmitoleic acid; C18:0, stearic acid; CI8:1, oleic acid; C18:2, linoleic acid; C18:3, linolenic acid.
b N ot detected.
c U/S ratio indicates: (CI6:1+_C18:1+ C18:2+ C18:3)/(C12:0+ C14:0+ C16:0+ C18:0)
製麹中の麹中のグリセリン, クエン酸, グルコース, 麹 菌体量および水分の 経時変化を Fig.3-1に示す。 グリセリンとクエン酸は18時間目に検出された後,
徐々に増加した。 焼酎麹の製造方法の特徴は製麹後半に温度を低下させ,
Aspergillus kawachiiのクエン酸生成を促すことにある85) 。 グリセリンとクエ ン酸は低温期でも増加し, 両者の生成パターンは同じ傾向を示した。 グルコー スは12時間自に検出された後, 徐々に増加し, 30時間目に最大値4.2gjl00g-koji を示した。 しかしながら, グルコース濃度は低温期では減少し, 44時間目で 3.4g/100g-koji まで低下した。 麹菌体量は30時間目まで増加したが, 低温期では ほぼ一定となり, 増殖は停止したものと考えられた。
焼酎麹のクエン酸生成の目安として製造管理で使用される出麹酸度と麹中の グリセリン濃度との関係を検討した。 その結果をFig.3-2に示す。 出麹酸度と麹 中のグリセリン濃度とは(3-3)式で表され (r=O. 74) , 出麹 酸度が麹中のグ リセリン濃度の管理指標になりうることが示唆された。
グリセリン濃度(gjl00g-koji) = 1.4X出麹酸度 (ml-0.1N NaOH)-O.4 (3-3) b. 大麦麹中のグリセリンとグリセリドの経時変化
第2章2-3-1で述べたように, 一般に使用される40---45時間製麹した大
麦麹は遊離グリセリンを含み, さらに, 前項でこれが製麹後半の低温期に増加 することを述べた。 ここで は, 大麦麹に由来するグリセリンを詳細に検討する ために, 大麦麹中の遊離グリセリン(F-GL)とグリセリド(G-GL)の製麹中
(ポ) ω」コ窃一02
(月、。ミ'OOOF\窃)ωω00コ一。
40 30
20 5 4
3
2 40
30
2 (00)
225caεωト
�崎、.ヌ?、 ι一、一 -一 之 O'
0> I
o 0>
ò 0 F 。
、- --
0> 去、
、- 0)
、-〆ε 主。一 ω〉〉の=ωQ弘三
20co一」ZO--O」ωoh一。
50 0
。
。 40
Koji
fermentation time(h)
20 30 10
Fig. 3-1. Glycerol, citric acid, glucose, mycelia weight and moisture content in barley
shochu koji
during
koji
fermentation.Symbols: 0 , glycerol�ム,citric acid�口,glucose�・,mycelia weight� • ,moisture. Line indicates the temperature.
。
。
08
。 7
5
2 6
4 3
(守主'OOOF\O)一O』ωO〉一。
5 4
Acidity (m/-O.1 N NaOH)
2 3
Fig.3-2. Relationship between glycerol and the acidity of barley
koji.
が, 大麦麹 で は時間経過に従って 増加し, 特に30時間以降の低温期に顕著に増 加した。 麹中のリバーゼ活性は30時間まで徐々に増加し, 36時間以降の低温期 には急激に増加したO これは, グリセリドの 減少と良く一致し ており, 製麹中
にAspergillus kawachiiが生産し たリバーゼの作用により グリセリドからグリセ
リンが生成されていることが示唆された。
3-3-3 発酵中の成分変化 a. 発酵 経過と酵母数
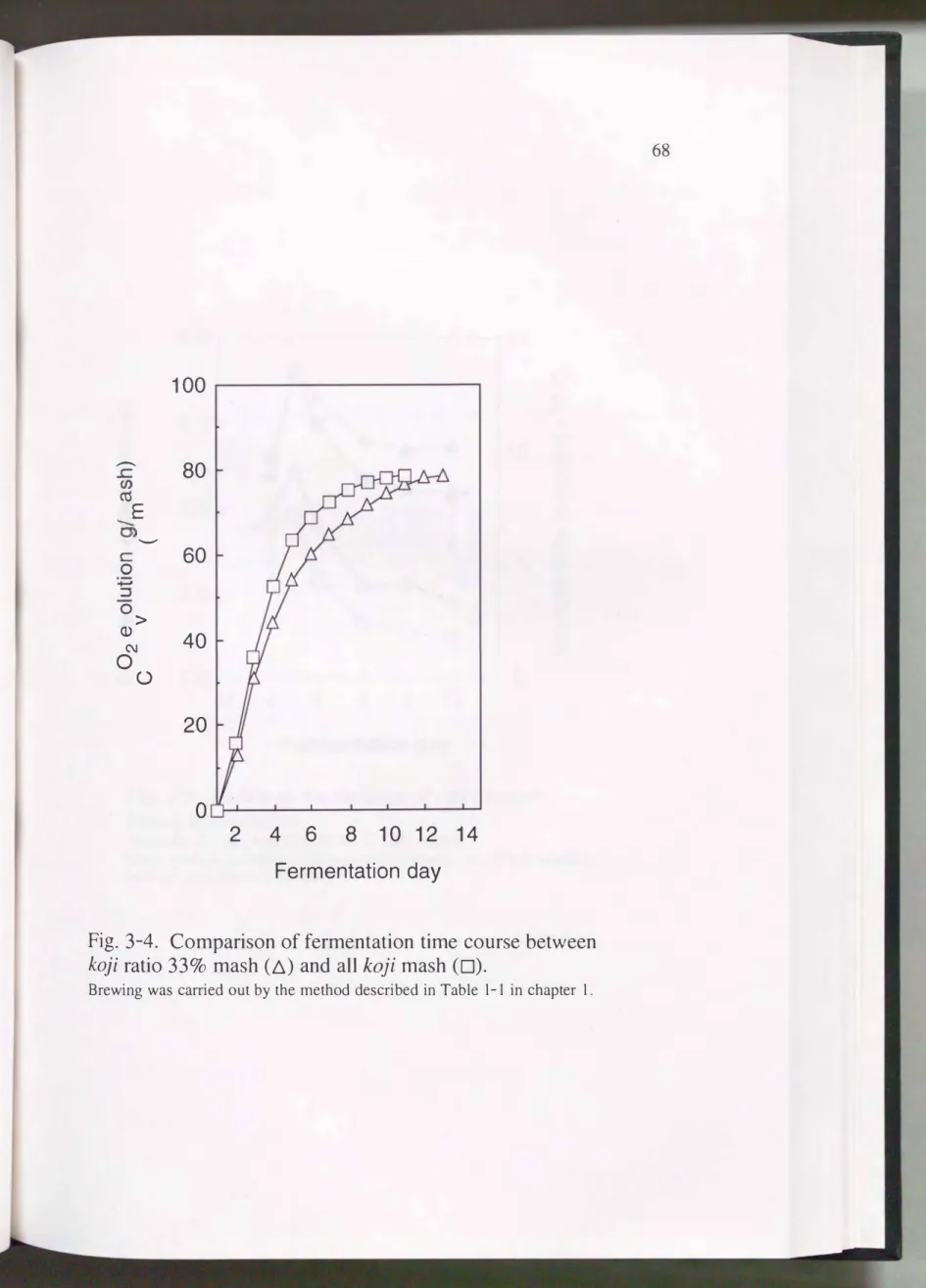
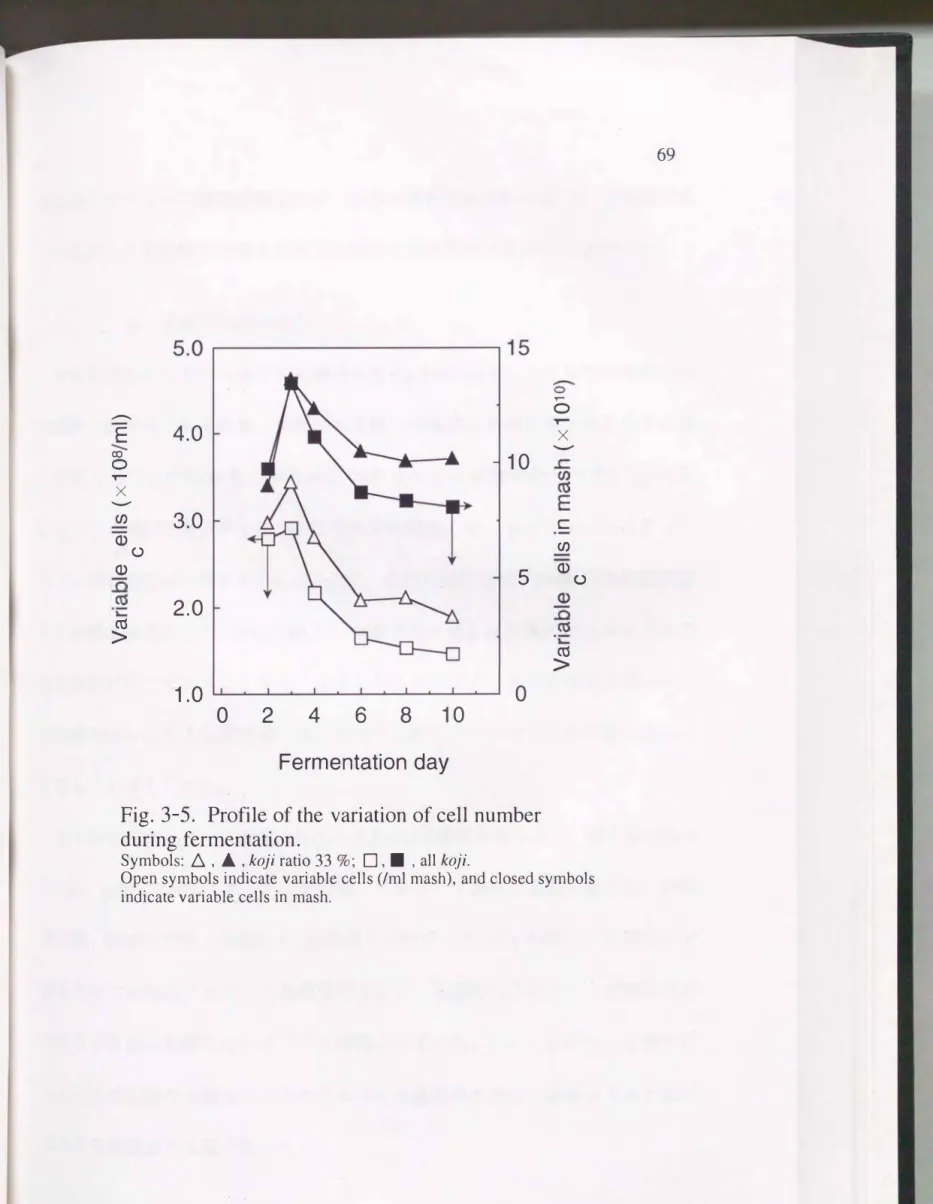
Fig.3-4および、Fig.3-5に総原料300g (麹原料100g)の仕込み を行ったときの発 酵経過および酵母数の変化 を示す。 発酵終了時の炭酸ガス発生量は全麹, 麹歩 合33%もろみともおよそ80gで、差違は見られなかったが, 全麹は麹歩合33%よ
り仕込み当初の発酵の進行が速か った。 もろみ1ml当たりの酵母の生菌数 は,
2.9�3.5 X 108cellsで3日目に最大となり その後発酵の進行に伴って1.4---1.9 X l08cellsまで減少した。 また 1ml当たりの生菌数は発酵の全期間を通じて全麹 の方が低くかっ た。 これに対し, もろみ中の酵母の全生菌数は13X 1010cellsで3 日目に最大となり その後徐々に減少した。 米焼酎の全麹もろみにおいて はも
(qov『刀mw一」℃'oh一Cコ)(0)kC一〉一ちのωωca一J 30 25
15 10 20
5
。 36 42
30 24 18
。 200
150 100 250
50
。
(JOEOR悶凶一JO,LR悶凶)一O」ωO〉一。』00一日の江
Time
(h)
ofkoji
fermentationFig.
3-3.Ratio of glycerol and lipase activity of koji
during koji fermentation.
( ) 正
、
何ωE
、 0
司 二、�
4C 0 コ�
O ω 〉
o tコC\I
100
80
60
40
20
。
2 4 6 8 10 12 14 Fermentation day
Fig. 3-4. Comparison of fermentation time course between
koji ratio 339もmash (ム) and all koji mash (口).
Brewing was carried out by the method described in Tab1e 1-1 in chapter 1.
...
-‘、
ちE
4.0
10
守ーー
×
、同国".
ω 3.0
ω 。
。 5
工cs3
� 2.0
〉cu
1.0 。
。 2 4 6 8 10
Fermentation day
Fig. 3-5. Profile of the variation of cell number during ferrnentation.
Symbols:ム, , koji ratio 33 %;口,・, all koji.
p。-句、
マ-。
守ーー
×
、ーー"
4ω
EE = 百
c
cn
ω ()
4ω ω コ
�
〉の
Open symbols indicate variable cells (lml mash), and closed symbols indicate variable cells in mash.
b. 各成分の経時変化
大麦焼酎主もろみ中の成分の経時変化をFig.3-6に示す。 もろみ中の各成分は
主原料(麹歩合330/0は大麦, 全麹は大麦麹) の成分に基因すると考えられる違 いが見られた。 すなわち, もろみ中の グリセリンは麹歩合 330/0で、2.7g/Zから 6.1g/Hこ, 全麹で5.0g/1から10.2g/1にそれぞれ増加した。 主もろみにおけるグリ
セリ ンの増加量はそれぞれ3.4, 5.2g/1で, その大部分は12� 144時間の発酵が盛 んな時間に生成された(Fig.3-6A)。 麹歩合330/0および全麹の仕込みは酒母の 製造条件が同じであることから, 主もろみにおけるグリセリン増加の違いは,
主原料の違いによる発酵環境の差に起因する酵母のグリセリン生成量の違いに よるものと考えられた。
もろみ中のグルコース濃度は仕込み直後の3時間自が最も高く, 麹歩合33%で 0.55%, 全麹で0.900/0であった。 その後, グルコース濃度は急激に低下し, 24時 間以降, 麹歩合33%, 全麹とも0.250/0以下で推移した(Fig.3-6B)。 米焼酎の全 麹もろみで初期のグルコース濃度は約100/0で, 高濃度のグルコースが酵母のグ リセリン生成に影響している4 0 )と報告されている。 しかしながら, 上述した ように大麦焼酎の全麹もろみ中のグルコース濃度は米焼酎の全麹もろみと比べ ると十分無視できる量であった。
15
A 10
5
(一\O)一O」ωo去の
。
0.5
(ポ)
ωω00コ一。
0.5
4.0
3.5 (JF)万一oco一主O
工己
96 120 144 168 192 216 240 264
Fermentation
(h)
72 48
。 24
Fig.
3-6.Glycerol (A), glucose (B), citric acid (C) concentration and pH (D) of barley
shochumash during fermentation.
Symbols: Â ,koji ratio 33 %;口 ,Allkoji.
に高くなった。 このことから, もろみのpHはクエン酸濃度に依存していると考 えられた(Fig.3-6D)。
3-3-4 もろみ中の脂肪酸と酵母の菌体内脂肪酸
a. もろみ中の脂肪酸
Table3-2に示すように, 大麦と大麦麹とでは脂肪酸に顕著な違いがあり, 麹 歩合がもろみ中の脂肪酸に影響することは明らかなことである。 ここでは, 仕 込み当初のもろみ中の遊離脂肪酸を調べた。 もろみ中のパルミチン酸, オレイ ン酸およびリノール酸の経時変化をFig.3-7に示す。 脂肪酸濃度の差は, 仕込み 直後においてのみ見られ パルミチン酸, オレイン酸およびリノール酸とも全 麹で多かった。 麹中に最も多く含まれるリノール酸のもろみ中の濃度は顕著に 異なり, 全麹もろみは麹歩合330/0もろみのおよそ1.7倍であった。 また, リノー ル酸, オレイン酸は1日目から2日目にかけて濃度の低下が見られたO これは酵 母が脂肪酸を取り込んだためではないかと推定された。
b. もろみから分離した酵母の菌体内脂肪酸
これまで麹歩合の違いが酵母の菌体内脂肪酸にどのような影響を与えるか明
1.5
0.5 (2ε) co--c」HCωocoo万一oco一ω
一oc
一」
0 1.5
0.5 (2ε) cozcおCφOC00万一06220
0 1.5
0.5
(2ε)
cozc」HCωQC00 万一ocoEE志向比
。 8 10
Fermentation day 6
4 2
Fig. 3-7. Change of free higher fatty acids in barley shochu mash.
Symbols: 0 , koji ratio 33%�・, al1-koji.
す。YEPD培地で培 養した酵母の菌体内脂肪酸は, パルミトオレイン酸が最も 多く, 次いでオレイン酸, パルミチン酸であったのに対し, もろみから分離し た酵母の菌体内脂肪酸ではリノール酸が主要な脂肪酸になっていたO また, も ろみ中の酵母の菌体内脂肪酸量に対するパルミチン酸の割合はYEPD培地と同 程度であったのに比べ, パルミトオレイン酸やオレイン酸の割合は顕著に減少 した。また, 主原料に大麦を使用した麹歩合330/0もろみから分離した酵母 に比 べ, 麹を用いた全麹もろみから分離した酵母ではリノール酸の菌体内脂肪酸に 占める比率がさらに増大していた。
乾燥菌体内の脂肪酸総量はYEPD培地で3.44mg /l00mg, 麹歩合330/0もろみ で 12.36mg/l OOmg , 全麹もろみで、15.93mg/l00mgとなり, もろみから分離した酵母 の方が顕著に多かった。 また, 麹歩合による脂肪酸総量の違いは3 .57mg/1 OOmg であったが, この内の7割がリノール酸の違いによるものであった。
以上のことから, 酵母はもろみからリノール酸を取り込んで利用していると 考えられ, 特にリノール酸は麹歩合の影響を大きく受けていた。
Table 3-3. Composition of free fatty acids extracted from barley shochu main mash at the initial stage of fermentation.
Amount of fatty acids (mg/l-mash) b
Koji ratio a Composition of fatty acids (%)
Cl的Cl的 ClωC16:1 C18:0 C18:1 C18:2 C18:3 Total U/S rati
g
33% (Standard) NDC 6.9 112.8 ND 19.9 42.3 225.8 ND 407.7
1.7 27.7 - 4.9 10.4 55.4 - 100 1.92
ND 13.7 184.6 ND 28.5 121.5 468.4 ND 816.7
100% (All koji) 一一一一一一一一一一一一一一一一一一一一 一一一一一一一一一一一一一一一一一一一 2.6 1 1.7 22.6 - 3.5 14.9 57.4 - 100
a Koji ratio (%) = (weight of koji materials) / (weight of total materials) x 100.
b Abbreviations were shown in Table 3-2 in chapter 3.
c Not detected.
d U/S ratio was shown in Table 3-2 in chapter 3.
ンはリバーゼ活性の増加パターンと類似しており, このことから大麦麹中のグ リセリドがリバーゼにより分解され減少したものと考えられた。 大麦麹に蓄積 するグリセリン(a)は, Aspergillus kawachiiが生成したグリセリン(b)と製 麹中にAspergillus kawachiiが生産したリバーゼ、の作用でグリセリドから生成し
たグリセリン(c)とに分けられた。 この両者の割合は, Fig.3-1およびFig.3-2の 結果からおよそ4:1と推定され, Aspergillus kawachiiが生成したグリセリンの方 が多いと考えられた。 一方, 大麦はグリセリンを含まず, リバーゼの作用でグ リセリンとなるグリセリド(e)のみであり, 42時間製麹の大麦麹のグリセリ
ド(ct)は大麦のおよそ半分の量であると推察された。 第2章で述べたように,
もろみ中のリバーゼ活性が低いことから, リバーゼの作用でグリセリンとなる 大麦麹のグリセリド(d)および大麦中のグリセリド(e)は発酵中もそのまま の状態にあると考えられた。
酵母のグリセリン生成は発酵初期のグルコース濃度が影響する3 7 , 4 0 )と報 告されている。 清酒の米麹の約300/0が可溶性糖であり, 麹原料は発酵初期の糖 の供給源としても重要である8 6 )との報告があるように, 一般に製麹すること で麹中にグルコースが蓄積され, 麹歩合が高いほど仕込み直後のもろみ中のグ ルコース濃度は高くなると推察される。 三枝ら1 0 )は製麹時間が異なる破砕米
Barley
, , , , , , , , , , , , ,
Barley koji
Glycerol
Fig. 3-8. How glycerol of barley koji comes from. Free glycerol (F-GL) in barley koji (a) is consisting of glycerol produced by Aspergillus kωvαchii (b) and glycerol derived from glyceride of koji digested by lipase (c) during koji fermentation. Whole glycerol containing in barley (e) and glycerol in koji derived from glyceride of barley (d) after koji fermentation.
仕込みでは, もろみ中のグルコース濃度は仕込み直後の最も高いときでおよそ 1%にすぎず\米焼酎の全麹仕込みとは明らかに異なっていた。
一般的な仕込み配合で ある麹歩合330/0の大麦焼酎もろみでの糖化, 発酵につ いては岩野ら2 6・2 8 )によって詳細な検討がなされている。 それによれば, 大 麦の溶解速度が米と比べて遅いために, 大麦焼酎もろみにおいては酵素による 大麦からのグルコース生成が発酵を律速していると報告されている。 大麦麹は グルコースを含むため, 全麹もろみ中のグルコース濃度は高く 推移すると考え られた。 しかし, 実際は大麦麹中のグルコース濃度が約30/0と低く, Fig.3-6Bに 示すようにもろみ中のグルコー ス濃度は仕込み直後に全麹の方が高い傾向が見 られたものの, 24時間以降0.250/0 以下で推移し, 麹歩合の影響をほとんど受け ていなかった。 麹中のグルコースは短時間で酵 母に消費され, その後は原料か らのグルコース生成が発酵を律速したと推定される。 グルコース濃度が酵母の グリセリン生成に及ぼす影響は次章で述べる。
培地中に存在する脂肪酸は速やかに酵母菌体内に取り込まれ, 増殖や香気成
分の生成に影響し, 特にリノール酸などの不飽和脂肪酸はエステル生成を阻害 する8 3 , 8 4 )と報告されている。 Table3-3 に示すように, YEPD培地で培養した 酵母からリノール酸は検出されず\焼酎もろみから分離した酵母にのみ含まれ,
しかも全麹もろみから分離した酵母の方が多かった。 また, この結果は, 麹歩 合が高い仕込みでは麹中の不飽和脂肪酸のために酵母のエステル生成が抑制さ れたという報告と符合する。 以上のように, 大麦麹中の脂肪酸がもろみ中の脂 肪酸に反映し, これが酵母に影響しているのは明らかである。 麹中の脂肪酸が 酵母のグリセリン生成に及ぼす影響については第4章および第5章で述べる。
3-5 小括
大麦麹および大麦焼酎の全麹もろみの特徴を明らかにすることを目的に実験 を行った。 大麦麹は大麦と比べ脂肪酸含量が多く, 特に不飽和脂肪酸組成の増 加が顕著であった。 大麦麹中のグリセリンは低温期の30時間以降に増加し , こ れはクエン酸生成パターンと同じ傾向を示した。 大麦中のグリセリドは製麹36 時間以降減少し始め, 42時間では大麦の約550/0まで減少した。 遊離グリセリン は製麹時間の経過に従って増加し , 特に36時間以降顕著に増加した。 一方, 麹 中のリバーゼ活性は30時間まで徐々に増加し 36時間以降は急激に増加した。
麹中に蓄積されるグリセリンは , Aspergillus kawachiiの代謝によって生成され るものとAspergillus kωvachiiが生成したリバーゼ、の作用でグリセリドから生成 されるものがあると推察された。
発酵は麹歩合33%よりも全麹の方が速く進行したが, 発酵終了時の炭酸ガス 発生量は同じであった。 もろみ1ml当たりの酵母の 生菌数は発酵の全期 間を通
生成された。 もろみ中のグルコース濃度は24時間以降0.5%以下で推移し, 麹歩 合の影響を受けなかった。 脂肪酸は全麹もろみの方が多く, 特に大麦麹に多く 含まれたリノール酸の違いが大きかった。 麹歩 合は発酵終了後のもろみから分 離した酵母の菌体内脂肪酸にも影響した。
第4章 全麹もろみにおける酵母のグリセリン高生成要因
4 - 1 緒言
もろみ中のグリセリンは, 焼酎製品の香気に間接的に影響する成分であり,
麹歩合33%もろみにグリセリンを添加した実験の結果, もろみ中のグリセリン
濃度が8---18g/1の範囲にあるとき, 焼酎製品の品質が最も好ま しいことを第l 章で述べた。 また, 第2章で一般的に行われる麹歩合33%仕込みのもろみ中の グリセリ ン濃度はおよそ6g/1であることを述べた が, 焼酎製品の品質を向上す るには, さらにもろみ中のグリセリン濃度を高める必要がある。もろみ中のグ
リセリン濃度を高めるた めには, 仕込み条件を変える方法および酵母のグリセ リン生成能を高める方法とがあるが, 本章では仕込み条件でグリセリン濃度を 高めるために, 全麹もろみで酵母のグリセリン生成が向上した要因を検討した。
酵母のグリセリン生成は培地の初期条件に影響される。 Pnachal and Stewart8 7 ) は,酵母の グリセリン生成が浸透圧の増加に伴って増加すること, Hemming
and Adler8 8 ) は グリセリン生成が塩化ナトリウムによって増加し, 浸透圧の作 用というよりむしろ"specific stress solutes"によるものではないかと報告してい
る。また, ワイン醸造においてはグルコースの初期濃度が高いほど増加し3 7 )
pHなどにも影響される8 9 ) ことが報告され 米焼酎の全麹もろみにお いては 酵母のグリセリン生成が高濃度のグルコースによって影響されると推察されて
いる40 ) 。
らに米と大麦とは溶解性に違い がある。 このこ とから, ワインや米焼酎もろみ において報告されている酵母 のグリセリン生成条件が大麦焼酎もろみにおける グリセリン生成条件とは必ずしも一致しないのではないかと考えられる。
前章で述べた大麦麹の成分および全麹もろみの特徴を踏まえて, 本章では全 麹もろみで酵母のグリセリン生成が増加した要因を検討した。
4-2 実験方法
酵母
第2章2-2に示した大麦焼酎用酵母BAW-6を使用した。
グルコース濃度およびクエン酸濃度を考慮した仕込み試験
グルコースの影響を調べた実験では, 麹歩合330/0仕込みの主もろみにグルコー
ス3, 6, 12および50gをそれぞれ加え, 仕込みを行った。 基本的な仕込み配合 は第2章Table2-1に示した配合比で行ったが , 糖源の量を同じにするため大麦 量をそれぞ、れ4, 8, 15および、62g減らして仕込みを行った。
クエン酸の影響を調べた実験では, 麹歩合330/0仕込みの主もろみにクエン酸 4および、9gをそれぞれ加え, 第2章Table2-1に示した配合で仕込みを行ったO
グルコース高濃度下でのクエン酸の影響を調べた実験では, グルコース50g とクエン酸4, 9および15gをそれぞれ加え, 第2章Table2-1に示した配合で主原 料の大麦量 を62g減らして仕込みを行った。 発酵 は前培養したBAW-6を2X 108cells/mlに調整した懸濁液10mlを接種後, 250Cで行った。
脂肪酸を添加した仕込み試験
第2章Table2-1 に示した仕込み配合で仕込みを行い, パルミチン酸100mg,
オレイン酸150mgおよびリノール酸500mgを主もろみに添加した。 発酵は前培 養したBAW-6を2X 108cells/m lに調整した懸濁液10mlを接種後, 250Cで行い, 炭 酸ガス減少量で発酵を管理した。
分析
グリセリン, グルコースの定量は第l章1 -2に, クエン酸の定量は第3章 3-2に, エタノールの定量は第3章3-2に示した方法で行った。
4-3 実験結果
4-3-1 グルコースの影響
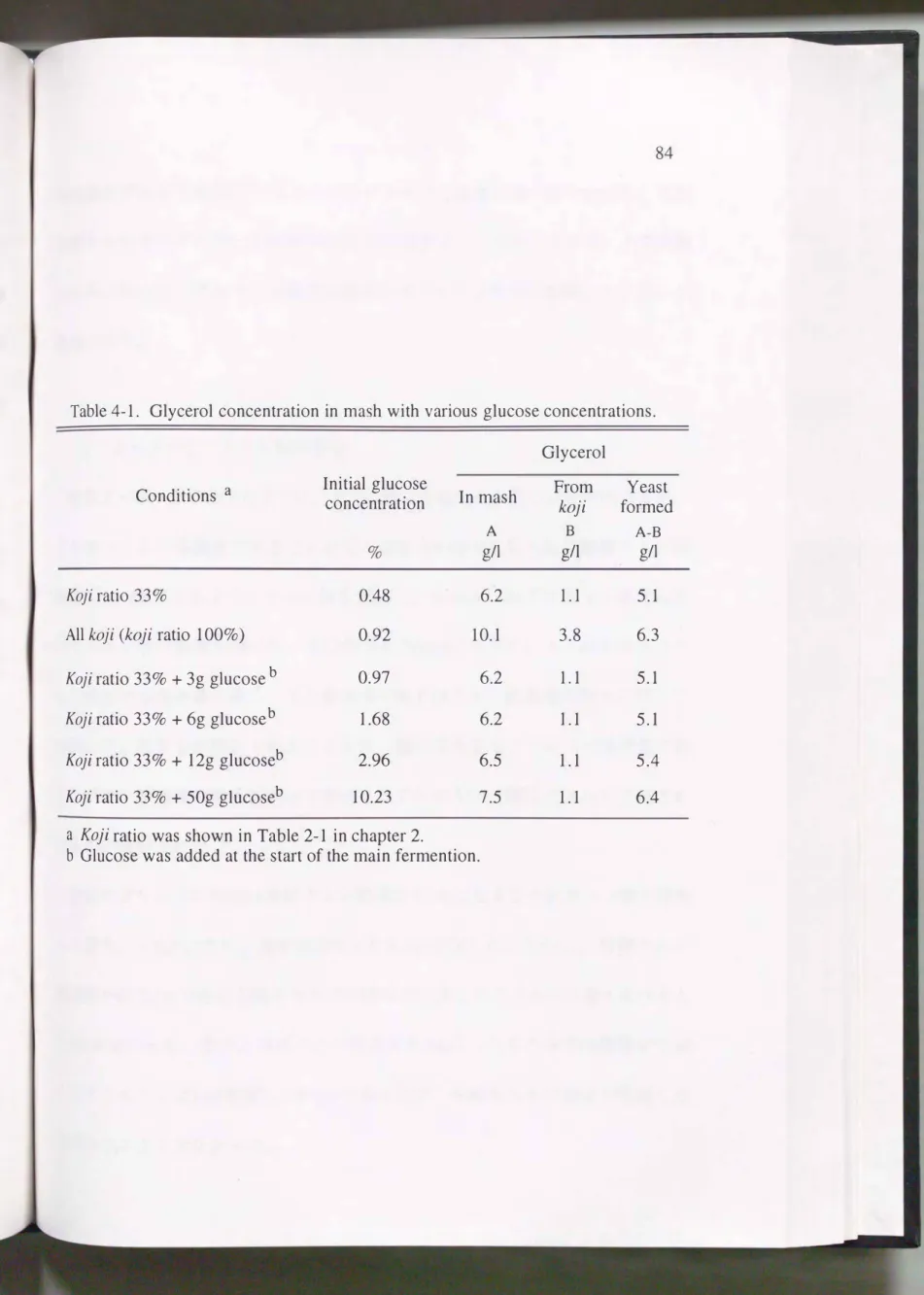
大麦焼酎の主もろみにグルコースを添加して生成するグリセリン濃度を比較 した。その結果をTable4-1に示す。 もろみ中のグリセリン濃度は麹歩合33%よ り全麹の方が3.9g/l高く, また, 初期グルコース濃度で30/0になるようにグルコー スを添加したもろみで、O.3g/1多かった。 しかしながら, グルコースを添加しでも
Table 4-1. Glycerol concentration in mash with various glucose concentrations.
Conditions a 1nitial glucose concentratlon
%
Koji ratio 33% 0.48
All koji (koji ratio 100%) 0.92
Koji ratio 33% + 3g glucose b 0.97 Koji ratio 33% + 6g glucoseb 1.68
Koji ratio 33% + 12g glucoseb 2.96 Koji ratio 33% + 50g glucoseb 10.23
a Koji ratio was shown in Table 2-1 in chapter 2.
1n mash A g/l
6.2 10.1 6.2 6.2
6.5 7.5
b Glucose was added at the st紅t of the main fermention.
Glycerol
From Yeast koji formed
B A-B
g/l g/l
1.1 5.1
3.8 6.3
1.1 5.1
1.1 5.1
1.1 5.4
1.1 6.4
初期濃度がおよそ20/0以下のもろみではグリセリン濃度に違いはなかった。 大麦 焼酎もろみ中のグルコース濃度は1%以下で推移する。 このことから, 大麦焼酎 もろみにおいて, グルコース濃度は酵母のグリセリン生成に影響していないと 推察された。
4-3-2 クエン酸の影響
前章3-3-2で述べたように, クエン酸は全麹もろみ中におよそ1%含まれ,
グルコースより高濃度であることから, 麹歩合33%の主もろみに初期クエン酸 濃度で1, 2%になるようにクエン酸を添加し, もろみ中のグリセリン濃度に及 ぼすクエン酸の影響を調べた。 その結果をTable4-2に示す。 もろみ中のグリセ リン濃度は全麹が最も高く また麹歩合330/0ではクエン酸濃度の増大に伴って 増加した。 麹歩合が同じであることから, 麹に由来するグリセリンは同量であ り, クエン酸濃度の増大に伴って増加したグリセリンは酵母に よって生成され たものと推定された。
酵母のグリセリン生成は初期クエン酸濃度が10/0になるようにクエン酸を添加 した場合, 5.8g/1 となり, 麹歩合330/0よりO.7g/1増加した。 しかし, 初期クエン 酸濃度が同じ1%である全麹もろみでの酵母が生成したグリセリン量と比べると
O.5g/ 1少なかった。 また, 初期クエン酸濃度を2%にしたもろみでの酵母が生成
したグリセリンは19/1増加して6.1 g/lで、あったが, 全麹もろみで酵母が生成した グリセリンより少なかった。
Table 4-2. Glycerol concentration in mash with various citric acid concentration.
Conditions
Koji ratio 33%
All koji (koji ratio 100%)
Koji ratio 33% + 4g citric acid a
Koji ratio 339も+ 9g citric acid a
lnitial citric acid concentratìon
%
0.31 1.01 1.04 1.98
ln mash
A g/l 6.2 10.1 6.9 7.2 a Citric acid was added at the start of the main fermentation.
Glycerol
From Yeast
koji formed
B A-B
g/l gll
1.1 5.1
3.8 6.3
1.1 5.8
1.1 6.1
4-3-3 高濃度グルコース下でのクエン酸の影響
高濃度グルコース下でのクエン酸の影響をグルコース初期濃度とクエン酸初 期濃度を調整した麹歩合33%もろみで調べた。 その結果をTable4-3に示す。 グ ルコース濃度を100/0になるように調整したもろみ中のグリセリン濃度は7.5g/1で グルコース濃度未調整の麹歩合33%よりも多かった。 もろみ中のグリセリン濃 度はさらにクエン酸濃度の増大に伴って増加し, クエン酸濃度10/0で、0.4g/l, 濃 度2%で1.4g/1それぞれ増加した。 以上のように, クエン酸濃度はグルコース高 濃度下でも酵母のグリセリン生成に影響することが明らかになった。
4-3-4 高級脂肪酸の影響
麹歩合330/0 の大麦焼酎もろみに高級脂肪酸を添加し 発酵後のもろみ中のグ リセリン濃度を比較した。 添加した脂肪酸は, もろみに比較的多く含まれたパ ルミチン酸, オレイン酸およびリノール酸とし, それぞれ全麹仕込みに相当す る量を計算で求めて 添加した。 その結果をTable4-4に示す。 パルミチン酸だけ を添加したもろみ中のグリセリン濃度 はコントロール(脂肪酸無添加の麹歩合 33%もろみ)よりも減少した が, それ以外はいずれも高かった。 全麹仕込み相
当量のパルミチン酸 オレイン酸およびリノール酸を添加することで酵母のグ リセリン生成は麹歩合33%のもろみよりもO.4g/l増加し, 脂肪酸濃度の違いが全 麹もろみで の酵母のグリセリン高生成の要因となっていると考えられた。 最も
Table 4-3. Effect of citric acid on glycerol concentration under high glucose concentration mash.
Glycerol
Conditionsa 1nitial citric acid From Yeast
concentratlon 1n mash koji formed
A B A-B
% g/l g/l g/l
Control 0.29 7.5 1.1 6.4
Control + 4g citric acid 1.02 7.9 1.1 6.8
Control + 9g citric acid 2.03 8.9 1.1 7.8
Control + 15g citric acid 3.07 9.4 1.1 8.3
a Control was a mash of koji ratio 330/0 to which 50g of glucose was supplemented.
Glucose and citric acid were added to the second mash. lnitial concentration of glucose in the mash was approximately 100/0.
Table 4-4. Effects of fatty acid supplement to barley shochu mash on glycerol formation.
Koji ratio Fatty acid (mg/300g barley)a
o rA ρU FLV vd G
g/1 gll
Yeast formed
A-B gll C16:0 C18:1 C18:2
1n mash From koji
A B
33% 0 0 0 6.2 1.1 5.1
All koji 0 0 0 10.1 3.8 6.3
100 0 0 5.9 1.1 4.8
o 150 0 6.9 1.1 5.8
o 0 500 6.7 1.1 5.6
33% 100
100 0 100
150 0 150 150
0 500 500 500
6.7 6.7 6.9 6.6
1.1 1.1 1.1 1.1
5.6 5.6 5.8 5.5
a C16:0, Palmitic acid, 100mg/300g-barley; C18:1, oleic acid, 150mg/300g-barley;
C18:2, linoleic acid, 500mg/300g-barley.
Brewing was done by 300g of barley with BA W -6.ー Fatty acids were added to the main mash. Temperature was maintained at 25 oC. Koji ratio was shown in Table 2-1 in chapter 2.
ミチン酸は酵母のグリセリン生成を抑制した。 リノール酸だけを添加したもろ み中のグリセリン濃度は6.5g/1で、コントロールよりも増加し, リノール酸は酵母 のグリセリン生成を促進した。 また オレイン酸とリノール酸を添加したもろ み中のグリセリン濃度は6.8g/1で, これはリノール酸のみを添加したもろみより
も多かったO しかし, オレイン酸のみを添加したもろみと同量であり, オレイ ン酸とリノール酸を同時に添加しでもグリセリン生成に対する相乗効果は得ら れなかったO
以上のように 脂肪酸が酵母のグリセリン生成に影響し 全麹もろみで酵母 のグリセリン生成が高まる要因の一つは脂肪酸であることが明らかになった。
しかしながら, オレイン酸およびリノール酸は酵母のグリセリン生成を増加さ せたのに対し, パルミチン酸は減少させるなど脂肪酸によってグリセリン生成
に対する影響は異なっていた。
4-3-5 クエン酸および脂肪酸の濃度を調整した
もろみ中のグリセリン濃度
全麹もろみと同じクエン酸および脂肪酸の濃度になるように 麹歩合33%の 王もろみにクエン酸および脂肪酸を添加し, 発酵後のもろみ中のグリセリン濃
度を比較した。 その結果をTable4-5に示す。 全麹もろみにおける酵母のグリセ リン生成は麹歩合33%もろみと比べ1.2g/1多く, 6.3g/1 であった。 これに対し,
もろみ中のクエン酸および脂肪酸を全麹もろみと同濃度になる ように添加する ことで, 酵母のグリセリン生成は麹歩合330/0もろみ よりも1.0 g/ 1増加して,
6.1g/1となり, これは全麹での酵母のグリセリン生成量と同程度であった。 以上 の結果, 全麹もろみで酵母がグリセリンを高生成したのはもろみ中のクエン酸 および脂肪酸の濃度の違いに基因するものであることが明らかになった。
4-4 考察
米焼酎の全麹もろみでは高濃度グルコースが酵母のグリセリン生成を向上さ せている4 0 )と報告されている。 しかし, 大麦焼酎の全麹もろみ中のグルコー ス濃度は, 第3章3-3-3で述べたように米焼酎の全麹もろみと比べると明 らかに小さく, 最高でおよそ1%にすぎなかった。 麹歩合330/0もろみの初期グル コースを調整したもろみでグリセリン生成を比較した結果, 初期のグルコース 濃度がおよそ30/0以上で向上したO このことから, 大麦焼酎の全麹もろみで酵母 のグリセリン生成が高まることにグルコース濃度はほとんど関与していないと 考えられた。 また, この結果はもろみに酵素剤を添加すること などにより, も ろみ中のグルコース初期濃度を高めることでグリセリン濃度向上が可能である ことを意味しており, これについては第8章で述べる。
Table 4-5. Glycerol formation by shochu yeast BA W -6 in barley shochu mash supplemented citric acid and fatty acid.
Glycerol
From Yeast
Exp. Conditions 1n nlash koji formed Note
A
g/l
Barley koji + Barley 6.2
2 1 + citric acid a 7.2
+ C16:0, C18:1, C18:2
3 Barley koji 10.1
B
g/1
l.1 l.1 3.8
A-B
g/l
5.1 6.1 6.3
Koji ratio 330/0 (standard type)
(All koji)
a Citric acid, 4g/300g-barley; C16:0, Palmitic acid, 100mg/300g-barley; C18:1, oleic acid, 150mg/300g-barley; C18:2, linoleic acid, 500mg/300g-barley.
クエン酸は, 製麹中にAspergillus kawαchiiが生成し, 麹中に蓄積されてもろ
みに持ち込まれる。 第3章3-3-3で述べたように, 全麹もろみ中のクエン 酸濃度は発酵の進行に伴って徐々に低下するもの のグルコース濃度よりも常に 高く推移することから, 酵母のグリセリン生成 に及ぼすクエン酸の影響を検討 した。 全麹もろみで酵母が生成したグリセリンは, 麹歩合330/0もろみより1.2g/l 多かったのに対し, 全麹もろみと同じなるようにクエン酸の初期濃度を調整し た麹歩合33%もろみで酵母が生成したグリセリンは, 麹歩合33%もろみより O.7g/1多かった。 このように, クエン酸濃度を同じ条件にした麹歩合33%もろみ でのグリセリンの増加量(ü.7g/l)は, 麹歩合の違いに 伴う酵母のグリセリン生 成量の差(1.2g/l)のおよそ6割に達しており, 全麹もろみで酵母のグリセリン 生成を増加させ ている最も大きな要因 はクエン酸濃度であると考えられた。 次 章では合成培地を用いて グリセリン生成に及ぼすクエン酸の影響を詳細に検 討し, その結果について述べる。
麹を主原料とした場合 もろみ中の脂肪酸初期濃度が高いことから, 主もろ みに脂肪酸を添加して影響を調べた。 その結果, 脂肪酸添加による酵母のグリ セリン生成 の増加量は麹歩合によるグリセリンの差のおよそ3割に相当した 。
もろみ中の脂肪酸濃度の違いが全麹もろみで酵母のグリセリン生成に影響し,
これがクエン酸に次いで2番目に大きな要因であることが明らかになった。 ま た? オレイン酸およびリノール酸は酵母のグリセリン生成を増加させた のに対
し, パルミチン酸は滅少させるなど脂肪酸の種類によってグリセリン生成に対
て代謝産物への影響は異なるという点で著者の結果と一致する。
Table4-5の結果から, 全麹もろみにおける酵母のグリ セリン高生成は, クエ
ン酸および脂肪酸の濃度の違いに基因すると結論された。
4-5 小括
大麦焼酎全麹もろみでの酵母のグリセ リン高生成要因を検討した。 麹歩合 33%の主もろみにグルコースを添加した結果 グルコースの初期濃度が2%以下 ではもろみ中のグリセリン濃度に影響せず 大麦焼酎全麹もろみでの酵母のグ リセリン生成にグルコースはほとんど影響しないと推定された。
麹歩合33%の主もろみに全麹もろみと同濃度になるようにクエン酸を添加し
た結果, 酵母のグリセリン生成は増加した。 グリセリンの増加量は全麹と麹歩
合330/0とのもろみ中のグリセリン量の差のおよそ6割に達した。
パルミチン酸 オレイン酸およびリノール酸を全麹もろみと同濃度になるょ
っに麹歩合33%の主もろみに 添加した結果, 酵母のグリセリン生成量は増加し
た。また, 全麹もろみと同濃度になるように, クエン酸と脂肪酸を添加したと ころ, 酵母は全麹もろみと同程度のグリセリンを生成した。 以上の結果から,
大麦焼酎全麹もろみで酵母がグリセリンを高生成したのは, クエン酸および脂 肪酸の濃度の違いに基因するものと結論された。