Effect of Acute Paraquat Toxicity on Ascorbic Acid Levels of Liver and Plasma in Mice
10
0
0
全文
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
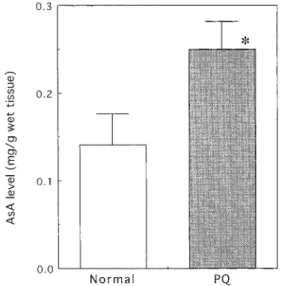
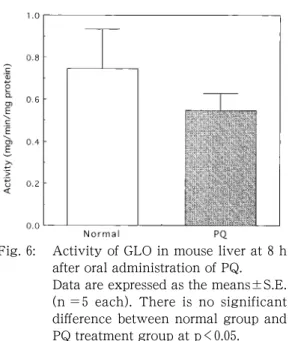
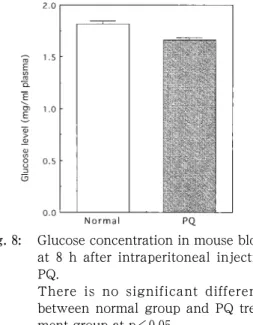
図

関連したドキュメント
熱力学計算によれば、この地下水中において安定なのは FeSe 2 (cr)で、Se 濃度はこの固相の 溶解度である 10 -9 ~10 -8 mol dm
Insulin and glucagon levels in living related liver transplantation : Their interaction with the recovery of graft liver function(.
The mGoI framework provides token machine semantics of effectful computations, namely computations with algebraic effects, in which effectful λ-terms are translated to transducers..
An example of a database state in the lextensive category of finite sets, for the EA sketch of our school data specification is provided by any database which models the
A NOTE ON SUMS OF POWERS WHICH HAVE A FIXED NUMBER OF PRIME FACTORS.. RAFAEL JAKIMCZUK D EPARTMENT OF
A lemma of considerable generality is proved from which one can obtain inequali- ties of Popoviciu’s type involving norms in a Banach space and Gram determinants.. Key words
de la CAL, Using stochastic processes for studying Bernstein-type operators, Proceedings of the Second International Conference in Functional Analysis and Approximation The-
[3] JI-CHANG KUANG, Applied Inequalities, 2nd edition, Hunan Education Press, Changsha, China, 1993J. FINK, Classical and New Inequalities in Analysis, Kluwer Academic
