カチオン性側鎖を持つポルフィリンによるグアニン 四重鎖
DNAの安定化機構のӕ明
2005 山下 健
Stabilization of Guanine Quadruplex DNA by the Binding of Porphyrins with Cationic Side Arms
2005
Takeshi Yamashita
Stabilization of Guanine Quadruplex DNA by the Binding of Porphyrins with Cationic Side Arms
Takeshi Yamashita
The telomerase is a target for the design of new anti-cancer drugs because telomerase is active in 85-90 % of human tumor cells and this enzyme combines and elongates telomeres by catalyzing the addition of telomere sequence so that cancer cells are immortalized. Telomere is the nucleoprotein structure at the end of chromosomes consisting of guanine-rich tandem repeats of the sequence d(TTAGGG) in human and forms G-quadruplex structure under physiological conditions. One of the approaches to telomerase inhibition is stabilization of G-quadruplex DNA by small molecules and suppression of the interaction between the telomerase and telomere. One of small molecules stabilizing G-quadruplex DNA is tetramethylpyridiniumyl porphyrin (TMPyP4) that binds externally at the end of quadruplex with its porphyrin ring stacked on the G-tetrad, and the four peripheral pyridinium groups are located in the groove of the quadruplex. I synthesized novel porphyrins with cationic pyridinium and trimethylammonium arms at para (pPy, pTm) or meta (mPy, mTm) position of all phenyl groups of tetratolyl porphyrin expecting their stronger interaction with G- quadruplex by both porphyrin ring and cationic side arms than well-characterized TMPyP4. I also synthesized ZnmPy in order to clarify the binding mode of cationic porphyrins with G-quadruplex. The results are summarized below.
Interaction of double-stranded DNAs and cationic porphyrins
Based on the melting temperature (Tm) of double-stranded DNAs, TMPyP4 and novel catonic porphyrins (pPy, pTm, mPy, and mTm) were found to stabilize the DNA almost equally. Absorption spectra and induced CD (iCD) spectra, however, show that TMPyP4 and novel porphyrins bind differently; TMPyP4 binds by self-stacking while pPy, pTm, mPy, and mTm bind mainly groove of double-stranded DNA, although all porphyrins bind to the surface of the DNA.
Interaction of G-quadruplex and cationic porphyrins
The quadruplex structure was found to be stabilized more by the meta-isomers than by the para-isomers and TMPyP4, and one mole equivalent of all porphyrins was sufficient to the stabilization of the quadruplex, as revealed by the increase of Tm of the quadruplex. Absorption spectra, iCD spectra, and fluorescence resonance energy transfer (FRET) spectra show that all porphyrins bind G-quadruplex at the same position, suggesting that porphyrins bind by external stacking with G-tetrad at the end of G-quadruplex or bind groove of the G-quadruplex.
Interaction of G-quadruplex and Zinc porphyrin
Because zinc porphyrin has five-coordinate structure, it does not favor intercalation and external stacking. Thus I synthesized ZnmPy and studied spectroscopically in order to clarify the interaction between cationic porphyrins and G-quadruplex. ZnmPy stabilized G-quadruplex almost equally to mPy, although 2-3 mole equivalents were required to sufficiently stabilize the quadruplex. ZnmPy binds groove of G-quadruplex as revealed by absorption spectra, iCD spectra and FRET spectra.
Modeling of cationic porphyrin / G-quadruplex complexes
I calculated minimum energy of cationic porphyrin / G-quadruplex complexes in various binding modes. The smallest energy was obtained for the groove binding, indicating that cationic porphyrins energetically favor groove binding to the G- quadruplex in a face-on manner and stabilize the G-quadruplex structure.
Conclusion
I synthesized novel cationic porphyrins and revealed that meta-isomers (mPy and mTm) stabilized G-quadruplex structure more strongly than para-isomers (pPy and pTm), and that cationic porphyrins bind groove of G-quadruplex face-on. All porphyrins bind the same position of G-quadruplex, thus the position of the cationic side arms is very important to stabilize the G-quadruplex structure. Since telomerase inhibition activity of a drug is strongly related to the stabilization of quadruplex DNA structure,
目次
第一章 緒ۗ 1
第二章 新֩カチオン性ポルフィリンの合成
第一節 序論 10
第二節 5,10,15,20-tetra (a-bromo-p-tolyl)porphyrin (pBr)の合成 11 第三節 5,10,15,20-tetra (a-bromo-m-tolyl)porphyrin (mBr)の合成 12
第四節 pPyの合成 13
第五節 pTmの合成 13
第六節 mPyの合成 14
第七節 mTmの合成 14
第三章 二重鎖DNAとカチオン性ポルフィリンとの相互作用
第一節 序論 15
第二節 ポルフィリン添加に伴う二重鎖DNAのTmの測定 15
第三節 CTDNA添加に伴うポルフィリンの吸収スペクトル変化 18
第四節 CTDNA添加に伴うポルフィリンの誘֬CD 20
第五節 小括 24
第四章 四重鎖とカチオン性ポルフィリンとの相互作用
第一節 序論 26
第二節 ポルフィリン添加に伴う四重鎖のTmの測定 28 第三節 四重鎖添加に伴うポルフィリンの吸収スペクトル変化 30 第四節 四重鎖添加に伴うポルフィリンの誘֬CDスペクトル 32 第五節 四重鎖添加に伴うポルフィリンのFRETスペクトル 35
第五章 四重鎖とポルフィリン亜鉛錯体との相互作用
第一節 序論 44
第二節 ZnmPyの合成 44
第三節 ZnmPy添加に伴う四重鎖のTmの測定 45
第四節 四重鎖添加に伴うZnmPyの吸収スペクトル変化 46 第五節 四重鎖添加に伴うZnmPyの誘֬CDスペクトル 46 第六節 四重鎖添加に伴うZnmPyのFRETスペクトル 47
第七節 小括 49
第六章 モデリングڐ算を用いた結合様式の検討
第一節 序論 50
第二節 ポルフィリンのスタッキングによるG-tetradの安定化 51 第三節 四重鎖のݱ格にあるリン酸基の荷の分布 53 第四節 ポルフィリンの四重鎖に対する結合様式の検討 55
第五節 小括 60
第七章 総括 61
実験の 64
引用文献 76
ࡤ辞 86
ફГच ࣜۗ
¸ॶ་Ʒ¹ƥƶૣিƓǑ۔ƳࠛǓdžƯ¹ບÐƲഗ֡ưٓƟ¹ƥǔƳઊƩ࣬
ƫƮƕƧº৶ۼƶࠑළ໓ƶਐǚۂƮLJǓư¹৶ۼdžNJƲƗƷ¹ڤԌ¹ѯư Օ৾औƔࠑළۍПƶरϳǚ࠷ƟƮƒǒ¹1950 ైનƳహǓưڥըࡏՔƔ 1ϳƳ ƲƫƮƌǓºdžƧ¹1981 ైϱݔƷ¹ϔॠি൝Ɣ 1ϳưƲƫƮƒǒ¹ƥƶঝƷ
ైÐरअƟ۔ƯƷࠑළ໓ƶ30%ǚৱljǓdžƯƳƲƫƮƌǓ1)ºƛƶۍПƷ¹ Օ৾औƳƷܨি൝ࡐƔ¹ڥըࡏՔƳƷݔϜޱƲƱƔӬದƝǔ¹ƥǔǑƶഗ֡
ƶຊේnj࠲ƳۉଙƲܑүǚ࠷ƟƮƌǓƶƳઓƟƮ¹ƔǛƲƱƶϔॠি൝Ƴ Ʒ¹ГǚࣩƕܑƲƔณƨƳӬದƝǔƮƌƲƌƛưƳƊǓº়ऺެභƔƔ ǛҠơǓۍПƷ¹ǟȆDzǣÓǺÓ(ԩ߱ਬ¹࠘ӱਅ¹දਅ¹Ҡԙ൝ࡐƲƱ) Ƴǐǒॶઑƶ০ڐইƯƊǓ DNAƔЂ(ƔǛЋஞࠃƶԩҠ)Ɵ¹ൖৄƝǔǓƛ ưƯ়ऺެභưƷЂƲǓެභƔিƝǔƮƗǓƛưƔƕƫƓƙưƲǓº়ऺެ
භƳƷ¹ЂƟƧ DNA ǚƟƧǒ¹ƥƶЂƔ࠲ƣƲƌषݜƷ¹ƥƶެභ
࠺ॱƔ࠺ߗơǓƛưƯƔǛҠƟƧެභǚࣩơǓिҠౖ֞ƔƟƮƌǓºdž Ƨ¹ઑఝƶืћެභƳǐƫƮNJƔǛҠƟƧެභƷܫڛƝǔࠑึƟƮƌǓºƟƓ ƟƲƔǑ¹ЂƳǐǒƥƶिҠౖ֞ƔƲǗǔƮƒǒ(DNA ЋஞࠃnjƔǛ ຫরЋஞࠃƶഩԩҠ)¹džƧ¹ȒȨȝÓǺÓ(ǡǟȦǴ¹ࠚු¹ѻൟƲƱ)ƶы
ƳǐǒืћެභƔƎdžƗ߁ພƟƲƌգ؈ƳƲƫƮƌǓषݜ¹ƔǛެභƔौ
Ɵ¹ƔǛਭॎƳƲǓºƔǛҠƶӬࠀƷ¹ǟȆDzǣÓǺÓƳǐǓ DNA ƶЂƯ Ɗǒ¹ƥǔǑƶЂƳǐǒ¹ƔǛЋஞࠃƶԩҠ¹DNAЋஞࠃnjƔǛຫর ЋஞࠃƶഩԩҠƔࢲƲƫƮƔǛެභưƲǓƛưƓǑ¹ƔǛƷЋஞࠃƶഗ֡ư
ۗƎƛưƔƯƕǓºdžƧ¹ǟȆDzǣÓǺÓưȒȨȝÓǺÓƶંƗƷ¹॒ࠧnjׇ
ѱƲƱശ૫ƶিԩư॥ƗշٱƟƮƌǓƛưƓǑ¹িԩ࢙ՖഗưNJۗƎƛưƔƯ ƕǓº
¸ƔǛƶ࠲Ʒ¹ؤࣗưਙॱƳൟƙǑǔǓºؤࣗưƷ¹ӱҭ
njදਅƲƱƶƛưƯƔǛƔdžƨЃƟƮषݜƳƷܑƨƔ¹ਙॱƳЃ
ƟƮƌǓषݜƳƷܕƓƲƌºГධ¹ਙॱƷ¹ܨƔǛޱnjȖȦȝȮޱƲƱƶ
ޱǚພƌƮ࠲ơǓධƯƊǒ¹ਙॱƳЃƟƧषݜnjƔǛǚՏਙƳ࠲ơƛ ưƔƯƕƲƌषݜƯNJ¹ƔǛƶપƕƝǚࣹƝƗơǓƛưƯѩฮܑүnjLJƲƱ ƶऔीǚཋǑƚǓƛưƔ֛ગƯƕǓºܨƔǛޱƷ¹ެභमӳܨƔǛޱưൟࠃ
இ࠲ƶ2ࡾ་Ƴൟ་ơǓƛưƔƯƕǓºެභमӳܨƔǛޱƷ¹ƔǛެ
භƶದА½ौƳഁঅƯƊǓ൝ࡐƶݜǚਰӳơǓનࡤዓܨޱ¹ƔǛެභƳઓ ƟƮਏવஇƳ௪ƗܨƔǛܨি൝ࡐ¹ެභƶൟ༡ƳࢲƲࣹըƶ௪ƕǚࠐlj ǓƛưƳǐǒƔǛެභǚࠑึƝƣǓࣹը߁ພ¹ƥǔƳҦƐƮ¹DNAǚǺÓ ǭǾȃưƟƧǝȦǨȦҠޱnjDzǴȒȤǼȮƳનഒƝǔǓಋؽߋઑƲƱƔƊǓ2)º ƟƓƟƲƔǑ¹ƌƢǔƶޱNJƔǛެභƳઓơǓਏવƔහƟƗࡶƳޙি½
ौƶଙƟƌެභƳ߁ພƟ¹ڥעۑࣺ¹ેโ¹ऎҠ֎औीƲƱƶ؊ƌ߁ພƔ֬
ǓưƌƎڠƔƊǓºГධ¹ൟࠃஇ࠲Ʒ¹ƔǛެභ۞ƶଦǚൟࠃȧ ȔȦƯஇưƟƧޱƶƛưƯ¹߁ພƶړۑƔ֛ગƝǔǓƔ¹ࡑພҠƝǔƮ ƌǓƔdžƨࣺƲƗ¹džƧ¹ࡑްƳࠑළࠧۥƔ֬ǓƲƱࠈǗƴ߁ພƔඦݥƝ ǔƮƌǓºdžƧ¹ެභƷ֕ƬNJƶЂƔLJࢲƲƫƮƔǛҠơǓƛưƓǑ¹Ɣ ǛެභƷ়ƳંࡾƳǗƧƫƮƌǓºƥƶڤү¹ƥƶিҠԙஇഗ໊ԙஇࡐƷં
ࡾંບƯƊǒ¹ƔǛެභƶࡾ་ƳǐƫƮܨƔǛޱƳઓơǓՕࢄƔЂƲƫƮƌ ǓƛưƓǑ¹ƌƢǔƶ൝ƳƒƌƮNJ¹உ҃ƯƕǓƔǛƶࡾ་ƳઓƟƮۙӧƔ ƊǓºƥƶƧljƌƗƬƓƶޱǚ௬Ƴ¹džƧƷݜ໊இƲ್ࣔƳ߹ພơǓഁ
ƔƊǒ¹ંޱ൮ພƔ݉ǗǔƮƌǓºƥƛƯ¹ఃƯંƗƶࡾ་ƶƔǛƳ߹
ພƯƕ¹ƔǛެභਏવƔݗƌ¹Ƭdžǒ߁ພƶࣺƲƌ൝ǚӬದơǓƛưƔ כยưƲƫƮƌǓº
¸ȁȨȜǝ(telomere)Ʒ¹२Ԍি൝ƶ৾ॐઑƶงൟƶƛưƯ¹ǩȥDzȞ܃Ư íงîǚϼฒơǓtelosưíൟîǚϼฒơǓmerosƓǑ߁ǑǔƧ܃ƯƊǓºȁ ȨȜǝƶѻ֏༞Ʒ¹1978ైƳȁȃȤȍȜȅǚພƌƧڽףƓǑฯǑƓƳƝǔ3)¹
۔džƯƳંƗƶࡾƶ༞ƔൟƓƫƮƌǓ4-7)ºƥƶ༞Ʒ¹ǫǝȆȮƳമǛƨ
༞ǚƟ¹ȍȃƳƒƌƮ ƶ ѻ֏ƶ٤ǒඉƟ༞ƔঝǨȨѻ֏Ƴ
ƫƮ੶ƌƮƒǒ¹ƥƶ3’งൟƷ¹100-200ѻ֏¹Г෭ޒൟƔఈࢽƟƮƌ
Ǔ8-10)ºdžƧ¹ƛƶǫǝȆȮƳമǛƨѻ֏༞ƷਙƮƶႄస་௫൝4)ƨƙƯƲƗ
ȁȃȤȍȜȅ3)njݏඟ5)¹ǦǟǮ6)¹ो൝ƶDzȨǟȇȅǵȅ7)ƲƱdžƯඐƝǔƮ
ƒǒ(Table 1)¹ƥƶܲ੧NJࠨƮƌǓƛưƓǑનǗǒƶܑƓƲƌౖ֞ǚƟƮƌ
ǓƛưƔੀƝǔǓº
Table 1. Sequence of telomere.
ࡑްƳȁȨȜǝƷ¹৾ॐઑƶक़॒nj¹ਭƐ¹งຈݜƲƱƓǑ৾ॐઑǚࡸǓ ưۗƎࢲƲԥǚƫƮƌǓƛưƔƫƮƌǓ10, 11)ºƝǑƳ¹৾ॐઑง
ൟƷRNAȒȤǟșÓൟǚൖৄƯƕƲƌƧlj¹ൖৄƶƧƻƳ50-200ѻ֏ࢹƟ (งൖৄ๓ભ¹ Figure 1)12, 13)¹ƊǓ୷ஶƗƲǓưǝȘȃÓDzǴǚСƕ֬ƛơ ƛưƓǑ¹ȁȨȜǝƷ¹ެභࢆฮƶࠉưNJ݂ƐǑǔƮƌǓºƥǔƳҦƐƮЋ ஞஇཀऔƶƌƗƬƓƳƷ¹ઑެභƶൟ༡ࢆฮƔƌƶư௬Ƴ¹ȁȨȜǝન ࡤƳЂऺƔƊǓषݜƔඦݥƝǔƮƌǓƛưƓǑ14-17)¹ཀҠ۔नưNJธৣƲշٱ ƔƊǓƛưƔ࠷މƝǔƮƌǓºГධ¹ȁȨȜȤÓǷƷ¹1985ైƳȁȃȤȍȜȅ ǚພƌƧڽףƓǑದۂƝǔƧ18)º1989ై¹ȁȃȤȍȜȅƶȁȨȜȤÓǷଓٸRNA ǚǪȨÓȆȮǫƟƧưƕƓǑג࡛ݏਬƯƊǓưຊੀƝǔƮƌƧ19)ºƥƶۼ¹
1997-1998ైƳȍȃǚսljƧݏਬȁȨȜȤÓǷƶǰȑȡȆǾȃƔ࠰ƌƯǪȨÓ
ɒɈ
ɆɭɩɒɡɊ ȫȤȳ
ȷɭȤɌɊȺɊ
ॆC. elegans ڐೠ
(TTAGGG)n (TTGGGG)n (TTAGG)n (TTTAGGG)n
(TG1-3)n (TTAGGC)n
lack
lack start replication
end replication
: template DNA : RNA primer : product DNA : DNA polymerase
Figure 1. Model of end replication problem. RNA primer binds template DNA and starts replication by polymerase. After replication, product DNA is shorter than template DNA because RNA primer portion is not replication.
ȆȮǫƝǔ¹ƥƶDNA༞ƔڸƝǔƧڤү¹ȁȨȜȤÓǷƷ¹DNA༞ƶ
ƓǑNJ¹ƥƶܲ੧ƓǑNJג࡛ݏਬƯƊǓƛưƔฯƟƧ20-22)ºȁȨȜȤÓǷ Ʒ¹ƔǛެභƶ85-90%Ưದ۔ƟƮƒǒ23, 24)¹৾ॐઑƶงൟƯƊǓȁȨȜǝ ƳڤݜƟȁȨȜǝ༞ƶॖସǚ॑౷Ɵ¹ƥƶସƝǚГ୩ƳඐƬƛưƯƔǛެභ ǚഩࠑҠƟƮƌǓ(Figure 2)25-28)ºdžƧ¹ȁȨȜȤÓǷƷ¹ȍȃƯƷ1132ǝȚȉ
߱ƓǑƲǓג࡛ݏਬൟư5’-CUAACCCUAA-3’ưƌƎ11ѻ֏ƶȁȨȜǝ༞
ưஇƲ༞ǚսLj451ȇǪȧǥǼȄƶଓٸRNAƶࡶƳమƬƯܲƝǔƮƒ ǒ¹ƥƶג࡛ݏਬȝǼÓȐƷࡾǚଵƐƮǐƗඐƝǔƮƌǓºȁȨȜȤÓǷ Ʒ¹ࡿᗡದিưธৣƲշٱƔƊǓƛư29)¹חƻ¹ȁȨȜȤÓǷƷ়ऺƲઑެභ
Figure 2. Model of elongation of telomere by telomerase.
C A A U C C C A A U C
3' 5'
T T A G G G T T A G G G T T A G G G
C C A
C A A U C C C A A U C
3' 5'
T T A G G G T T A G G G T T A G G G
C C A
C A A U C C C A A U C
3' 5'
T T A G G G T T A G G G T T A G G G T T A G G G
C C A
C A A U C C C A A U C
3' 5'
T T A G G G T T A G G G T T A G G G T T A G G G
C C A
T T A G G G T T A G G G G G
C C A
T T A G G G T T A G G G T T A G G G T T A G G G
C C A A A T C C C A A T C C C A A T C
telomerase fixation
telomere elongation by telomerase
telomerase movement
polymerase extension
telomere elongation by telomerase
ƯƷǃưǛƱದ۔ƟƮƌƲƌƛưƓǑ¹ƔǛਏવƔݗƗ¹߁ພƶࣺƲƌॠ ƟƌܨƔǛޱƶǺÓǭǾȃưƟƮ֛ગƝǔƮƒǒ30)¹ࡑްƳȁȨȜȤÓǷƶଓ ٸRNAǚǺÓǭǾȃưƟƧǝȮǼǶȮǴnj31, 32)¹ȁȨȜȤÓǷƶԩǚຫর ơǓǺȮȌǪࡐƶದ۔௮33, 34)¹džƧƷࣹൟࠃǚພƌƮȁȨȜȤÓǷƔȁȨȜ ǝƳڤݜơǓƶǚਰӳơǓධƲƱƔڽףƝǔƮƌǓ35, 36)º
¸DNAƷ¹ǝȂȆȮ¹ǫǝȆȮ¹DzȃDzȮ¹ǼȚȮƶ߾Ƭƶѻ֏ư¹ȥȮ߱¹ ȂǥǨDzȥȗÓǴƓǑܲƝǔƮƌǓºǝȂȆȮưǼȚȮ¹ǫǝȆȮưDzȃDz ȮƔƥǔƦǔWatson-CrickٸƶਬڤݜǚٺƟ¹ୋऺమ෭ƶޒƔג݉ƟƮƌ ǓమࢲǑƣǛܲ੧ǚưƫƮƒǒ37)¹ȍȃƯƷެභƳƒƌƮ30Ғѻ֏ઓƊǓº ƟƓƟƲƔǑ¹ȁȨȜǝ٤ǒඉƟ༞ǚƬГ෭ޒൟƔ¹িઑఝऽڨҟƯ߾
ࢲޒܲ੧ǚưǓƛưƔޚؼඦݥƝǔƧ38, 39)º߾ࢲޒƷ¹ǫǝȆȮƳമǛƨГ෭ ޒƔǒƧƧdžǔƧܲ੧ǚưƫƮƒǒ¹džƨিઑఝƯƷƥƶܲ੧ƔԏీƝǔƮ ƌƲƌƔ¹ȁȨȜǝ40, 41)njc-mycƶȒȨȝÓǺÓЋஞࠃ42, 43)¹ืћǫȨȑȥȮƶ ǴǟǾǼЏ44)ƲƱƔ߾ࢲޒܲ੧ǚࡷǒƎǓ༞ǚƟƮƌǓƛư¹džƧ¹߾
ࢲޒƳڤݜơǓǺȮȌǪࡐƔƊǓƛư45-48)¹߾ࢲޒܲ੧ǚӕƕГ෭ޒƳơǓݏ ਬƔƊǓƛư49-53)ƲƱƓǑ¹߾ࢲޒƷিઑఝƯࢲƲԥǚƫƮƌǓƛưƔ
࠷މƝǔƮƌǓº߾ࢲޒƷ¹మƬƶൟƓǑܲƝǔƮƒǒ¹ГƬƷ¹TTAȦ ÓȒൟƯ¹NJƎГƬƷ߾ƬƶǫǝȆȮƔHoogsteenٸƶਬڤݜƳǐƫƮٺ
ƝǔƧǫǝȆȮ߾ࢲޒ൲ൟáG-tetradâƯƊǓ54, 55)ºԌ࠶֡ำǚພƌƧӕ
ǐǒǑǔƧຜњଇƯƶܲ੧Ʒ¹TTAȦÓȒƔൟࠃյڤݜƟƧG-tetradƶઓ Ԓਅחƻ੩ƳƊǒ¹߾෭ƶDNAޒƔג൲݉ƳܕƟƮƌǓƛưƔඦݥƝǔƮ ƌǓ54)ºdžƧ¹Xਅڤईܲ੧ӕǐǒǑǔƧڤईଇƯƶܲ੧Ʒ¹ൟࠃյڤݜ
ƟƧG-tetradƶ੩ǚTTAȦÓȒƔઓԒਅरƳਜ਼ƫƮƒǒ¹߾෭ƶDNAޒƷ¹൲
݉ƳܕƟƮƌǓƛưƔඦݥƝǔƮƌǓ55)ºƝǑƳ¹ຜњଇưڤईଇධƳƒ ƌƮ¹ߡƬƶG-tetradƔπ-π۶߁ພƳǐƫƮଽਅஇƳ൵ǛƯƒǒ¹ƥƶ൲յ ƶِյƳƷؽੳǟǥȮáNa+, K+ƲƱâƔƟƥƶܲ੧ǚϩ୩ҠƟƮƌǓºƛ ƶ߾ࢲޒܲ੧ǚϩ୩ҠơǓƛưƳǐƫƮ¹ȁȨȜȤÓǷƶ ȁȮȒȧÓȃƔ
ȁȨȜǝDNAƳڤݜơǓƛưǚਰӳƝǔ¹ƥƶڤү¹ȁȨȜȤÓǷƶॖସಶ҃
ǚਰӳƯƕǓƛưƔൟƓƫƮƌǓ56)ºޚؼ¹߾ࢲޒƶงƳƊǓG-tetradƳઓƟ
Ʈπ-πǴǺǾǨȮǫơǓƛưƯ¹߾ࢲޒܲ੧ǚϩ୩ҠơǓමݖգǚơǓҠݜ൝
Parallel G-quadruplex DNA
G G G
G
G G
G
G
G G G G
Antiparallel G-quadruplex DNA
G G G
G
G G
G
G
G G G G
N H H
N N
N N N
H H R
O H
N N N N
N H H
R O
H
N N N N
H H
R O H N
N N N H
R
O
N
G-tetrad
A B C
M+
Figure 3. (A) Chemical structure of G-tetrad formed by Hoogsteen-type hydrogen bonding network (dashed lines). A metal (M) is accommodated at the center of the G-tetrad. (B) Schematic representation of the antiparallel quadruplex DNA. The arrows represent the direction (5' to 3') of the sugar phosphate backbone. (C) Schematic representation of the parallel quadruplex DNA. The arrows represent the direction (5' to 3') of the sugar phosphate backbone.
C A A U C C C A AU C
3'
5' T T A G G G
G G C C A
G G G
G G G
G G G
G G G
T T A G G G G G
C C A
G G G
G G G
G G G
G G G
Inhibition of telomerase fixation Formation of G-quadruplex Structure
ƔඦݥƝǔƮƒǒ57-67)¹ƥƶƛưƓǑ߾ࢲޒܲ੧ƶϩ୩ҠƳƷ¹Ҡݜ൝ƶದॐ
ƶƔࢲƯƊǓư݂ƐǑǔƮƌǓº
¸߾ ࢲޒܲ੧ǚϩ ୩ҠơǓҠ ݜ൝ƶГƬƯ ƊǓ5,10,15,20-tetra-(N-methyl-4- pyridyl)porphyrin (TMPyP4, Figure 5A)Ʒ¹߾ࢲޒƳઓƟƮݗƌ९ཋƔƊǒ¹ԯ Ƭ؊ƌȁȨȜȤÓǷਰӳԩǚƬƛưƔඦݥƝǔƮƌǓ68-71)ºdžƧ¹ƥƶڤ ݜƷ¹߾ࢲޒƶงƳƊǓG-tetradưȘȦȐǞȥȮգƔǴǺǾǨȮǫƟƮƒǒ¹ գƶࢍඈƳƊǓ߾ƬƶȏȥdzȆǡț֏Ʒ¹ƥǔƦǔ߾ƬƊǓ߾ࢲޒƶܷƳ
ƟƮƌǓºȘȦȐǞȥȮգƶưG-tetradƶƷǃDŽ௬ƠƯƊǓƛưƓǑ¹
TMPyP4ƶࡶƲϩ୩Ҡƶ֞ܲƷ¹ȘȦȐǞȥȮգưǫǝȆȮѻ֏յƯƶπ-π۶
߁ພƳǐǓӱǴǺǾǨȮǫƯƊǓưƝǔƮƌǓºГධ¹TMPyP4ƶϳЂ
ઑƯƊǓ5,10,15,20-tetra-(N-methyl-2-pyridyl)porphyrin (TMPyP2, Figure 5B)Ʒ¹
TMPyP4ƳೝǁƮȁȨȜȤÓǷਰӳԩƔୢƌ69, 70)ºƛƶǐƎƳ¹ȁȨȜȤÓǷ
ਰӳԩƷ¹ȘȦȐǞȥȮգƳڤݜƟƮƌǓ֏ƶϳƔธৣƳշٱƟƮƌ
ǓºTMPyP4ǚսljƧǦǼǥȮȘȦȐǞȥȮƷ¹మࢲޒDNAnjߡࢲޒDNAư
ƶ۶߁ພƳƬƌƮƷંƗƶ૯ۂƔඦݥƝǔƮƌǓƛư72-93)¹džƧ¹ȘȦȐǞ ȥȮ࠺ॱࡿᗡࢥƔƊǓƛưƓǑ94-96)¹ȘȦȐǞȥȮƷ¹߾ࢲޒưҠݜ൝ư ƶ۶߁ພǚଳǁǓरƯພƲڽףਬưƌƎƛưƔƯƕǓº
Figure 5. Chemical structure of (A)TMPyP4 and (B) TMPyP2.
NH N N
HN
N N N
N
A B
N H N N
HN
N N
N
N
¸ƥƛƯଙࡣƷ¹߾ࢲޒǚTMPyP4ǐǒ؊Ɨϩ୩ҠơǓƛưƯ¹ǐǒ؊ƌȁȨ ȜȤÓǷਰӳԩǚ࠷ơƛưǚ֛ગƟƮ¹߾Ƭƶ੩ޒƳǦǼǥȮƶ֏ǚ
ƬȘȦȐǞȥȮǚ০ڐ½ݜƟƧºpTmưpPyƷ¹ȜǸϳƶȐǢȆȦ֏ƶȌ ȤϳƳǦǼǥȮƶȃȥȜǼȦǝȮȝȆǡț֏ưȏȥdzȆǡț֏ǚƥǔƦǔ੩ ޒƳƫƮƒǒ¹TMPyP4ưೝǁƮ੩ޒƔସƌºdžƧ¹mTmưmPyƷ¹ƥǔƦ ǔȜǸϳƶȐǢȆȦ֏ƶȜǺϳƳ֏ƔƊǒ¹ƥǔƦǔpTmưpPyƶϳЂ
ઑƯƊǓºdžƢ¹మࢲޒDNAưƶ۶߁ພƳƬƌƮંƗƶ૯ۂƔƊǓƛưƓ Ǒ¹మࢲޒDNAưȘȦȐǞȥȮưƶ۶߁ພǚຈӕҜஶ(Tm)¹טࢌǴȕǪȃȦ¹
Ѥమॐ(circular dichroism, CD)ǚ߹ພƟƮଳǁƧº࠰ƳƛǔǑƶȘȦȐǞȥȮ
ƶ߾ࢲޒƶϩ୩Ҡƶ֒ຌǚଳǁǓƧljƳTmǚ੯୩ƟƧºƥƟƮƛǔǑƶȘȦȐ ǞȥȮƶϩ୩Ҡƶ֞ܲƶЊƌǚଳǁǓƧljƳטࢌǴȕǪȃȦ¹CDǴȕǪȃȦ¹ fluorescence resonance energy transfer (FRET) ǴȕǪȃȦƶ੯୩¹חƻȝȂȥȮǫ ڐ߬ǚ݉ƫƧº
ફమच¸ॠ֩ǦǼǥȮȘȦȐǞȥȮƶݜ
ફГ২¸ࣥཉ
¸TMPyP4 Ʒ¹Wheelhouse ǑƳǐƫƮ߾ࢲޒƳઓƟƮڤݜƟ¹ƥƶܲ੧ǚϩ୩
ҠơǓƛư¹חƻݗƌȁȨȜȤÓǷਰӳԩǚơǓƛưƔඦݥƝǔƮƌǓ68, 69)º ƥƶࡶƲϩ୩Ҡƶ֞ܲƷȘȦȐǞȥȮգư G-tetrad յƯƶπ-πǴǺǾǨȮǫƳǐ ǓºdžƧ¹Han ǑƳǐƫƮ TMPyP4 ƶϳЂઑƯƊǓ TMPyP2 Ʒ¹TMPyP4 ǐǒNJȁȨȜȤÓǷਰӳԩNJୢƌƛưNJඦݥƝǔƮƌǓºƛƶƛưƷȁȨȜ ȤÓǷਰӳԩƳƷ¹ȘȦȐǞȥȮգƳǐǓ۶߁ພƶLJƲǑƢ¹ȘȦȐǞȥ ȮգƶȜǸϳƓǑࢽƮƌǓ੩ޒƶыƔપƕƌƛưǚϼฒƟƮƌǓºƥƛƯଙ ࡣƷ¹ȘȦȐǞȥȮգƳҦƐƮ੩ޒƳǐǓ۶߁ພƳǐƫƮ TMPyP4 ǐǒNJ९ ཋ½ڤݜƔरअơǓƛưǚ֛ગƟ¹ॠ֩ǦǼǥȮȘȦȐǞȥȮǚ০ڐ½ ݜƟƧºdžƢ¹߾ࢲޒƶȥȮ߱֏ưƶ۶߁ພƔ؊ƗƲǓƛưǚ֛ગƟƮ¹
TMPyP4 ǐǒNJସƌǦǼǥȮȏȥdzȆǡț੩ޒǚƬॠ֩ȘȦȐǞȥȮ pPy
ưƥƶϳЂઑƯƊǓ mPyǚݜƟƧºdžƧ¹֏ƶЊƌƳǐǓыǚଳ ǁǓƧljƳ¹ȃȥȜǼȦǝȮȝȆǡț֏ǚƬpTmưmTmǚݜƟƧº
Figure 6. Chemical structure of synthetic meso-tetraphenylporphyrins, pPy, pTm, mPy, and mTm.
CH2 N(CH3)3
N CH2
CH2 N(CH3)3 N CH2 NH
N N HN R2
R2
R2
R2
R1
R1 R1
R1
pTm
R1 =H
R2 = R2 = R1 = mPy
H
mTm
H
R2 = R1 = R2 =H pPy
R1 =
ફమ২¸5,10,15,20-tetra (α-bromo-p-tolyl)porphyrin (pBr)ƶݜ
¥Scheme 1 ƳǦǼǥȮȘȦȐǞȥȮ pPy חƻ pTmƶۍưƲǓ pBr ƶݜ
ډ༲ǚ࠷ơºp-tolunitrile ǚࢽದ൝ࡐưƟƮພƌ¹benzene ଇƯ p-tolunitrile ƶȜ ǼȦ֏ǚ N-bromosuccinimide ǚພƌƮȑȨȝҠƟα-bromo-p-tolunitrile ǚƧº
࠰Ƴ¹α-bromo-p-tolunitrile ƶȆȃȥȦ֏ǚ benzene ଇƯ diisobutylalminium hydride(DIBAL-H)ǚພƌƮճیƟα-bromo-p-tolualdehyde ǚƧº࠰Ƴ¹CHCl3 ଇ Ưα-bromo-p-tolualdehyde ư pyrrole ǚ ށ ݜ Ɵ Lindsay ǚ ພ ƌ Ʈ 5,10,15,20-tetrakis(α-bromo-p-tolyl)porphyrin (pBr)ǚƧ97)º
CN
CN CHO
CHO H
N
CH3 CH2Br CH2Br
CH2Br
NH N N
HN Br
Br
Br
Br N-Bromosuccinimide
BPO
benzene
triethylamine DDQ
4 eq. 4 eq.
dry benzene reflux
BF3-Et2O
DIBAL-H
reflux
p-tolunitrile α−bromo-p-tolunitrile α−bromo-p-tolualdehyde
pyrrole
pBr
CHCl3
ફߡ২¸5,10,15,20-tetra (α-bromo-m-tolyl)porphyrin (mBr)ƶݜ
¸Scheme 2 ƳǦǼǥȮȘȦȐǞȥȮ mPy חƻ mTm ƶۍưƲǓ mBr ƶݜ
ډ༲ǚ࠷ơºm-tolunitrileǚࢽದ൝ࡐưƟƮພƌ¹benzene ଇƯm-tolunitrileƶ ȜǼȦ֏ǚN-bromosuccinimideǚພƌƮȑȨȝҠƟα-bromo-p-tolunitrileǚƧº
࠰Ƴ¹α-bromo-m-tolunitrileƶȆȃȥȦ֏ǚbenzeneଇƯdiisobutylalminium hydride (DIBAL-H)ǚພƌƮճیƟα-bromo-m-tolualdehyde ǚƧº࠰Ƴ¹CHCl3 ଇƯα- bromo-m-tolualdehyde ư pyrrole ǚށݜƟ Lindsay ǚພƌƮ 5,10,15,20- tetrakis(α-bromo-m-tolyl)porphyrin (mBr)ǚƧ98)º
Scheme 2. Synthetic pathway of mBr CN
CH2Br
CN CHO
CH2Br
CHO
CH2Br
HN
NH N N
HN Br
Br
Br Br
CH3
triethylamine DDQ benzene
reflux
benzene
BF3-Et2O
N-Bromosuccinimide BPO
4 eq. 4 eq.
DIBAL-H
mBr
m-tolunitrile α−bromo-m-tolunitrile α−bromo- m-tolualdehyde
pyrrole
reflux CHCl3
ફ߾২¸pPyƶݜ
pBrƳӄवƶpyridineǚҦƐ¹1.5յճ໙ƟƮpPyǚƧ(Scheme 3)º
Scheme 3. Synthetic pathway of pPy
ફ۵২¸pTmƶݜ
pBrƳӄवƶȃȥȜǼȦǝȚȮǣǺȉÓȦຜњǚҦƐ 80˚CƯ14յҦేƟ pTmǚƧ(Scheme 4)º
NH N N
HN N
N
N
N
trimethylamine pBr
pTm
80 °C, stir
NH
N N HN N
N
N
N
pyridine pBr
pPy
reflux
ફང২¸mPyƶݜ
mBrƳӄवƶpyridineǚҦƐ¹1.5յճ໙ƟƮmPyǚƧ(Scheme 5)º
Scheme 5. Synthetic pathway of mPy
ફࡆ২¸mTmƶݜ
mBr ƳӄवƶȃȥȜǼȦǝȚȮǣǺȉÓȦຜњǚҦƐ 80˚C Ư 14 յҦే
ƟmTmǚƧ(Scheme 6)º
NH
N N HN N
N
N N
pyridine mBr
mPy
reflux
NH
N N HN N
N
N N
mBr
trimethylamine
mTm
80 °C, stir
ફߡच¸మࢲޒDNAưǦǼǥȮȘȦȐǞȥȮưƶ۶߁ພ
ફГ২¸ࣥཉ
¸߾ࢲޒưҠݜ൝ưƶ۶߁ພƳƬƌƮƷ¹ؼై¹ীǛƳڽףƝǔƮƒǒ¹ƥ ƶڤݜບƳƬƌƮNJ¹Ԍ࠶֡ำ(nuclear magnetic resonance, NMR)54)nj Xਅ ڤईܲ੧ӕ55)ǚພƌƮӕฯƝǔƮƕƮƌǓºƟƓƟƲƔǑ¹Xਅڤईܲ੧Ʒ¹ ຜњଇƯƶܲ੧ǚಶьƟƲƌषݜƔƊǓƛưnjԌ߱ưҠݜ൝ưƶൖݜઑƶڤई ҠƷຓϾƯƷƲƌƛư¹džƧ¹NMR ư X ਅڤईӕưNJƳƥƶӕƳƷࡻ
ƲָౖƔഁƲƛưƲƱඋƳڠƙǓºГධ¹טࢌǴȕǪȃȦnj CD¹ڏ
ǴȕǪȃȦƲƱƷૐƯNJຓϾƳ੯୩ơǓƛưƔƯƕ¹ޙ۔NJݗƌƛưƓǑ¹ ƛǔǑƶࡹǚພƌƮҠݜ൝ƶ߾ࢲޒƳઓơǓ۶߁ພƳƬƌƮຓϾƳ૯Ǔƛ ưƔƯƕǓǐƎƳƲǔƸ¹߾ࢲޒƳڤݜơǓҠݜ൝ƶӬದǚॳljƮƌƗरƯ
ќƯƊǓºȘȦȐǞȥȮƷ¹26πࠃڈǚƩ¹400 nm പؼƳπ"πਐЃƳ
֬ПơǓȘȦȐǞȥȮƶ Soret ખưǐƸǔǓ؊ƌטࢌƔƊǓºdžƧ¹ȘȦ ȐǞȥȮƷ¹؊ƌڏNJದơǓºȘȦȐǞȥȮƶ Soret ખƷ¹ȘȦȐǞȥȮƶ գ؈ीઙƳљധƯƊǒ¹ຓϾƳƥƶҠnj¹ڏ؊ஶƶҠǚୈƎƛưƔƯƕ ǓƛưƓǑ¹DNAưƶ۶߁ພǚൟԙஇࡹƓǑۂƮƌƗरƯພƲڽףਬ
ƯƊǓº۔ƶưƛǕ¹ƛǔǑƶࡹǚພƌƧ߾ࢲޒưȘȦȐǞȥȮưƶ
۶߁ພƳƬƌƮƶඦݥƷࣺƲƌºƥƛƯ¹ଙࡣƷ¹మࢲޒ DNA ưȘȦȐǞȥ Ȯưƶ۶߁ພƳƬƌƮƷơƯƳંƗƶ૯ۂƔඦݥƝǔƮƌǓƛưƓǑ 72-96)¹ džƢॠ֩ƳݜƟƧȘȦȐǞȥȮưమࢲޒ DNA ưƶ۶߁ພƳƬƌƮ¹మࢲ ޒ DNAƶ Tm¹טࢌǴȕǪȃȦ¹֬ CDǴȕǪȃȦǚພƌƮڸƟ¹ƥƛƓ ǑǑǔƧऻඦǚ֏Ƴ߾ࢲޒưƶ۶߁ພƳƬƌƮڸơǓƛưƳƟƧº
ફమ২¸ȘȦȐǞȥȮҦƳಳƎమࢲޒDNAƶTmƶ੯୩
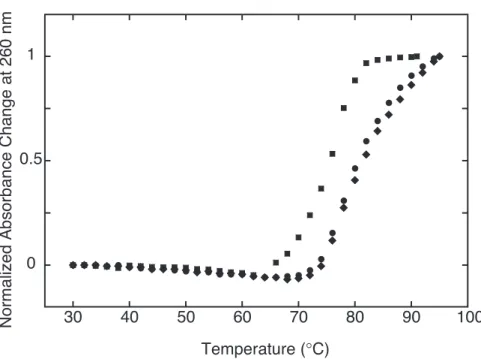
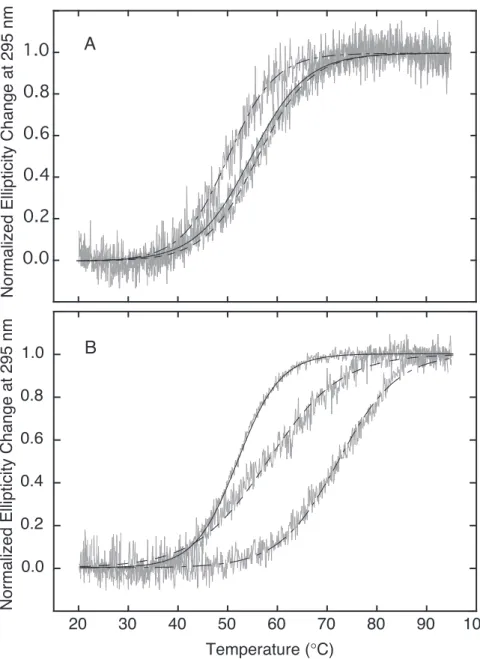
¸ஔਘƶమࢲޒ DNA ƯƊǓ߷ǡDzؖਈ DNA(Calf thymus DNA, CTDNA)ư חƻݶӖݜƟƧ ¹ ¹ ¹ ưƶൖݜઑƶϩ୩ǚଳǁ
ǓƧljƳTmǚ20 mM sodium phosphate buffer, 20 mM NaCl pH 7.0ଇƯ੯୩ƟƧº మࢲޒ DNA ƷǑƣǛܲ੧ǚٺơǓưѻ֏ઓյƶǴǺǾǨȮǫƳǐǒטஶ ƔۑࣺơǓºҜஶरअƳưNJƲƌГ෭ޒ DNA ƔƟƮГ෭ޒƳƲƫƮƌƗ ƳƟƧƔƫƮטஶƶरअƔհ੯ƝǔǓºTmƷ¹మࢲޒ DNAƔՏਙƳГ෭ޒ ƳǃƱƙƧưƕƶଇƶҜஶǚ࠷Ɵ89, 99)¹TMPyP4njݶӖݜƟƧpPy¹pTm¹ mPy¹mTmƶమࢲޒDNAƳઓơǓϩ୩Ҡǀƶ֒ຌǚ૯ǓƛưƔƯƕǓº
¸džƢ¹CTDNAƨƙƶưƕƷ¹Tm = 75.0 ˚CưƲƫƧ(Figure 7)º࠰ƳƥǔƦǔ ƶȘȦȐǞȥȮưCTDNAƔR = 0.1 (R = [porphyrin] / [base pair])ƶưƕƶൖݜઑ ƶ Tm ǚ੯୩ƟƧºdžƧ¹ȘȦȐǞȥȮҟƯ CTDNA ƨƙƶƳೝǁƮƱ ǔƨƙTmƔरअƟƧƓǚ∆TmƯഒƟƧºTMPyP4ƷTm = 81.0 ˚CưƲǒ∆Tm Ɣ6.0˚CƯƊƫƧ(Table 2)ºГධ¹ݶӖݜƟƧ4ࡾ་ƶҠݜ൝pPy¹pTm¹mPy¹ mTmƷ¹ƥǔƦǔ∆TmƔ6.0¹5.5¹4.5¹3.5˚CưƲƫƧ(Table 2)ºpPyưpTm
ƯƷ∆Tmƶ૮ƔTMPyP4ưǃDŽ௬Ơ૮ưƲǒ¹௬୷ஶϩ୩ҠƟƮƌǓư݂ƐǑ
ǔ¹mPy ư mTm Ʒ∆Tm ƶ૮ƔࡲՒୢƗƲƫƮƒǒϩ୩Ҡǀƶ֒ຌƷઓஇƳ
ୢƗƲƫƮƌǓºdžƧ¹֏ƳǐǓϩ୩Ҡǀƶ֒ຌƷ¹ȏȥdzȆǡț֏੩ޒ ǚơǓpPy ưmPyƷ¹ȃȥȜǼȦǝȮȝȆǡț֏੩ޒǚơǓpTmưmTm ǐǒƥǔƦǔ 0.5¹1.0˚C ưǃǛƶǗƢƓƨƙݗƗƲƫƧºƛƶƛưǐǒ
֏ƳǐǓыƷǃưǛƱƲƌƛưƔǗƓƫƧº
¸ϱरƶƛưƓǑమࢲޒ DNA ƶϩ୩ҠƳƷ੩ޒƶସƝnjƥƶϳƳշٱƲƗ ǃDŽ௬ƠƯƊǓƛưƔൟƓƫƧºƛƶƛưƓǑ¹ƌƢǔƶȘȦȐǞȥȮNJమࢲ ޒDNAƳઓƟƮ௬ƠǐƎƲڤݜǚƟƮƌǓƛưƔ࠷މƝǔƧº
Figure 7. Normalized absorbance changes of 38.6 µM base pair CTDNA at 260 nm in the absence (closed squares) or presence of 3.9 µM TMPyP4 (R = 0.1, closed diamonds) and mPy (closed circles) against temperature in 20 mM sodium phosphate buffer, 20 mM NaCl (pH 7.0).
¸¸¸Table 2#Tm and ∆Tm of CTDNA in the absence or presence of cationic porphyrins 0
0.5 1
30 40 50 60 70 80 90 100
Temperature (°C)
Normalized Absorbance Change at 260 nm
TMPyP4 81.0 6.0
pPy 81.0 6.0
mPy 79.5 4.5
CTDNA only 75.0
pTm 80.5 5.5
mTm 78.5 3.5
Tm (°C) ∆Tm (°C) porphyrin
ફߡ২¸CTDNAҦƳಳƎȘȦȐǞȥȮƶטࢌǴȕǪȃȦҠ
¸DNA ҦƳಳƎȘȦȐǞȥȮƶ Soret ખƶҠƓǑȘȦȐǞȥȮƶڤݜບ
ǚ୩ơǓƛưƔƯƕǓºDNA ѻ֏ઓյహƝǔǓषݜ(intercalation)Ʒ¹Soret ખƶئપטࢌౡସƔપƕƗȧǾȄDzȐȃƟ(≥ 15 nm)¹džƧ؊ƌॐܑү(≥ 35%) ǚ࠷Ɵ¹DNA ƶǫȦÓȑƳڤݜƟƮƌǓषݜ(groove binding)Ʒ¹ࣹƝƗȧǾȄ
DzȐȃƟ(≤ 8 nm)¹džƧࡴƌॐܑү(≥ 10%)NJƟƗƷॐܑүǚ࠷ơƛưƔ૯
ǑǔƮƌǓ 72-74, 85, 100-103)ºdžƧ¹Soret ખƶҠƶӄ୷ƓǑ DNAưƶڤݜӄ୷ǚ
੯ơǓƛưƔƯƕǓº
¸džƢޚࣖƳTMPyP4ưమࢲޒDNAưƶڤݜƳƬƌƮଳǁƧº20 mM sodium phosphate buffer, 20 mM NaCl pH 7.0ଇƯTMPyP4ƳCTDNAǚҦƟƮƌƗư
420 nmƶטஶƔࣦÐƳۑࣺƟ¹ئપౡସƷȧǾȄDzȐȃƟƧºdžƧ¹טࢌ
ƔۂǑǔƧƧljƛƶ൲݊ಶ҃ƷГ૫ӭƯƊǓƛưƔ࠷މƝǔƧ(Figure 8A)º
¸࠰ƳݶӖݜƟƧŏࡾ་ƶҠݜ൝ pPy¹pTm¹mPy¹mTm ưమࢲޒ DNA ư ƶڤݜƳƬƌƮଳǁƧ(Figure 8B)ºԇȘȦȐǞȥȮƳ CTDNA ǚҦƟƮƌ Ɨư¹ƥǔƦǔ 410 nm പؼƶטஶƔۑࣺƟئપౡସƔȧǾȄDzȐȃƟƧº ƟƓƟƲƔǑ¹ƥƶҠƷரଇdžƯƷטࢌǚ࠷ƟƧƔ¹TMPyP4 ưƷЂƲ
ǒR = 0.20പؼƓǑטࢌƷӱǔƮƌƫƧƛưƓǑ¹൲݊ಶ҃ƷࣺƲƗưNJ
మ૫ӭƊǓƛưƔǗƓƫƧº
¸džƧ¹ƥǔƦǔƶȘȦȐǞȥȮƶȧǾȄDzȐȃ(∆λ)ưॐܑү(H)ǚ Table 3 ƳdžưljƧºƌƢǔƶȘȦȐǞȥȮNJȧǾȄDzȐȃƷǟȮǺÓǦȧÓDzȢȮǚ
࠷ơ15 nmǐǒNJࣹƝƓƫƧ (8.0-13.0 nm)ºdžƧ¹ॐܑүƷ¹TMPyP4ưpPy¹ mTm ƯƷƥǔƦǔ 50.1¹44.0¹43.0%ưƲǒǟȮǺÓǦȧÓDzȢȮǚ࠷ơ 35%
ǐǒNJપƕƗƲƫƧƔ¹pTmưmPyƷ¹ƥǔƦǔ32.5¹32.3%ưࣹƝƗƲƫƧº ƛǔǑƶڤүƓǑ¹TMPyP4 ư pPy¹mTm Ʒ¹ǟȮǺÓǦȧÓDzȢȮưమࢲޒ DNA ƶӱƳڤݜƟƮƌǓධƶҧౖƔ¹pTm¹mPy Ʒ¹మࢲޒ DNA ƶǫ ȦÓȑƳڤݜƟƮƌǓҧౖƔ࠷މƝǔƧº
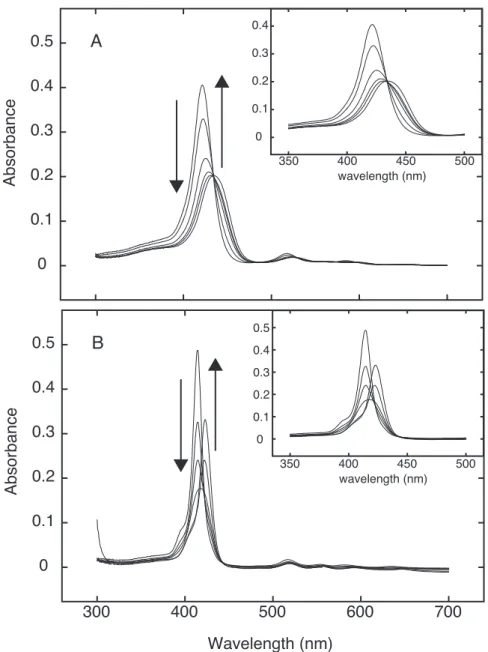
Figure 8. Absorption spectral change of TMPyP4 (A) and mPy (B) on the addition of CTDNA.
The porphyrin concentration was 1.5 µM. The spectra were recorded in 20 mM sodium phosphate buffer, 20 mM NaCl (pH 7.0). A, R values were as follows ∞, 1.08, 0.52, 0.39, 0.31, 0.10. B, R values were as follows ∞, 0.90, 0.60 ,0.40, 0.05, 0.01. Insets show enlarged spectra in the Soret region.
0 0.1 0.2 0.3 0.4 0.5
0 0.1 0.2 0.3 0.4 0.5
300 400 500 600 700
Wavelength (nm)
0 0.1 0.2 0.3 0.4
350 400 450 500
wavelength (nm)
0 0.1 0.2 0.3 0.4 0.5
350 400 450 500
wavelength (nm)
A
B AbsorbanceAbsorbance
Table 3. Spectroscopic data for the porphyrins bound to CTDNA. ∆λ, bathochromic shift.
Hypochromicity (H) was determined by the equation H = (εf — εb) / εf × 100, where εf and εb
represent the molar absorptivities of free and bound porphyrins, respectively.
ફ߾২¸CTDNAҦƳಳƎȘȦȐǞȥȮƶ֬CD
¸CTDNA ưƶڤݜƳಳƎȘȦȐǞȥȮƶ֬ CD ǴȕǪȃȦǚ੯୩ƟƧºȘ
ȦȐǞȥȮ࠺ઑƷഩ্ਬǚƫƮƌƲƌƧljƳ CD Ʒհ੯ƝǔƲƌƔ¹ǨȤ ȦƲൟࠃƯƊǓ DNAưڤݜơǓƛưƯ SoretખപؼƳCDƔ֬ƝǔǓºƛƶ ƛưǚພƟƮȘȦȐǞȥȮƔమࢲޒ DNA ƳઓƟƮƱƶǐƎƳڤݜƟƮƌǓ Ɠ୩ơǓƛưƔƯƕǓºГೃƳ¹়ƶȏÓǪǚ࠷ƟƧषݜƷǫȦÓȑڤݜ¹
ാƶȏÓǪǚ࠷ƟƧषݜƷѻ֏ઓյǀƶǟȮǺÓǦȧÓDzȢȮ¹়ưാƶȏÓ Ǫǚ࠷ƟƧषݜƳƷ DNA ഒरƯƶ࠺ۡǴǺǾǨȮǫ(self-stacking)ư֟ੳƝ ǔǓ72, 73, 75, 83, 85, 100)º
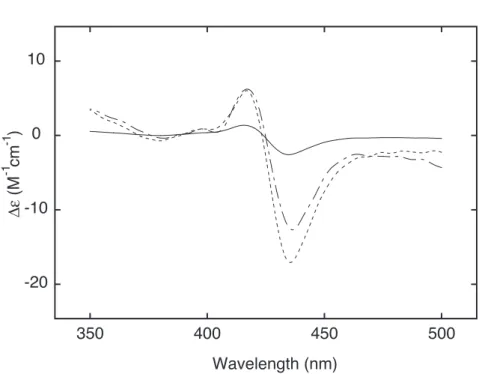
¸džƢ¹TMPyP4ƶǴȕǪȃȦǚۂƮLJǓư¹R = 0.1ưR = 0.05¹R = 0.01ƶư ƕƌƢǔNJ়ưാƶȏÓǪƔ֬ƝǔƧƛưƓǑ¹TMPyP4 Ʒ DNA ഒरƯ
࠺ۡǴǺǾǨȮǫƟƮƌǓƛưƔ࠷މƝǔƧ(Figure 9)º࠰ƳݶӖݜƟƧpPy¹ pTm¹mPy¹mTmƶǴȕǪȃȦǚۂƮLJǓư¹pPyƯƷ¹R = 0.1¹0.05ƶưƕ
়ƶȏÓǪƨƙƲƶƳઓƟƮ¹R = 0.01 ƶưƕ়ƶȏÓǪưƥƶସౡସ੩Ƴࡴ
TMPyP4 pPy pTm mPy mTm
13.0 8.0 8.5 9.0 8.0
50.1 44.0 32.5 32.3 43.0
∆λ (nm) H (%) porphyrn
ƌാƶȏÓǪƔհ੯ƝǔƧºpTm ƯƷ R = 0.1ư 0.05¹0.01ƶƌƢǔNJ؊ƌ
়ƶȏÓǪưƥƶସౡସ੩ƳࡴƌാƶȏÓǪƔհ੯ƝǔƧ(Figure 10)ºdžƧ¹mPy
ưmTmƷ¹R = 0.1ư0.05¹0.01ƶƌƢǔNJ؊ƌ়ƶȏÓǪưƥƶସౡସ੩
ưౡସ੩ƳࡴƌാƶȏÓǪƔհ੯ƝǔƧƔ¹R = 0.1ƶưƕƷସౡସ੩Ƴೝǁ Ʈౡସ੩ƶാƶȏÓǪƔ¹R = 0.01 ƶưƕƷౡସ੩ƳೝǁƮସౡସ੩ƶാ
ƶȏÓǪƔƥǔƦǔ؊ƗƲƫƧ(Figure 11)ºƌƢǔƶȘȦȐǞȥȮNJࡶƳ়ƶ ȏÓǪǚ࠷ƟƮƌǓƛưƓǑ¹ƥǔƦǔࡶƳǫȦÓȑڤݜǚƟƮƌǓư݂ƐǑ ǔǓºƟƓƟƲƔǑ¹࣐Ƴ়ƶȏÓǪƶLJƯƷƲƗƥǔƦǔࡴƌാƶȏÓǪ Ɣସౡସ੩Ɠౡସ੩ƶƱƩǑƓdžƧƷධƳۂǑǔƧƛưƓǑ¹ƌƗƬƓƶ ڤݜບƔށƞƫƮƌǓƛưƔ࠷މƝǔƧº
Figure 9. Induced CD spectra of 5 µM TMPyP4 in the presence of CTDNA at R = 0.1 (solid line), 0.05 (dashed line), and 0.01 (chained line). The spectra were recorded in 20 mM sodium phosphate buffer, 20 mM NaCl (pH 7.0).
-20 -10 0 10
∆ε (M cm )-1-1
350 400 450 500
Wavelength (nm)
Figure 10. Induced CD spectra of 5 µM porphyrins in the presence of CTDNA at R = 0.1 (solid lines), 0.05 (dashed lines), and 0.01 (chained lines). A, pPy; B, pTm. The spectra were recorded in 20 mM sodium phosphate buffer, 20 mM NaCl (pH 7.0).
-5 0 5 10 15 20 -10
0 10 20 30
350 400 450 500
Wavelength (nm)
∆ε (M cm )-1-1 ∆ε (M cm )-1-1
A
B
Figure 11. Induced CD spectra of 5 µM porphyrins in the presence of CTDNA at R = 0.1 (solid lines), 0.05 (dashed lines), and 0.01 (chained lines). A, mPy; B, mTm. The spectra were recorded in 20 mM sodium phosphate buffer, 20 mM NaCl (pH 7.0).
350 400 450 500
-10 -5
0 5 10 15 20 25
Wavelength (nm) -10
-5 0 5 10 15 20
A
B
∆ε (M cm )-1-1 ∆ε (M cm )-1-1
ફ۵২¸ࣹԨ
¥TMPyP4 Ʒమࢲޒ DNA ҦƳಳƎטࢌǴȕǪȃȦҠƳƒƌƮ¹טࢌ
ƔۂǑǔƧƛưƓǑ¹ڤݜӄ୷ƔГ૫ӭƯƊǓƛưƔǗƓƫƧºdžƧ¹ݶӖݜ
ƟƧ pPy¹pTm¹mPy¹mTm ƶטࢌǴȕǪȃȦƶҠƷ¹ƌƢǔNJரଇdžƯ Ʒטࢌǚ࠷ƟƧƔ¹మࢲޒ DNA ǚҦƐƮƌƗư࠰ફƳטࢌƓǑӱǔ ƮƌƫƧºƛƶƛưƓǑ¹ݶӖݜƟƧҠݜ൝ƯƷڤݜӄ୷Ɣమ૫ӭϱरƯƊ ǓƛưƔǗƓƫƧºdžƧ¹טࢌǴȕǪȃȦƶڤүưమࢲޒ DNA ҦƳಳƎ
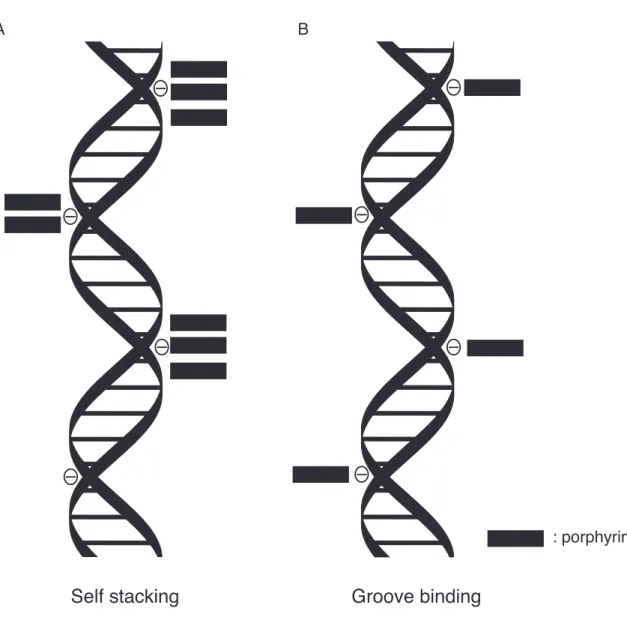
֬CDǴȕǪȃȦƶڤүǚݜஇƳ૨ƟƮ¹TMPyP4ƷDNAഒरƯ࠺ۡǴ ǺǾǨȮǫƟƮ DNA ƳڤݜƟƮƌǓưڤཉƭƙƧ(Figure 12A)ºГධ¹ݶӖݜ
ƟƧȘȦȐǞȥȮƷƌƢǔNJڤݜບƷࡶƳǫȦÓȑڤݜƯƊǓưڤཉƭƙ
Ƨ(Figure 12B)ºƟƓƟƲƔǑ¹ǴȕǪȃȦƔ়ƶȏÓǪƨƙƯƷƲƌƛư¹R
ƶ૮ƳǐƫƮȏÓǪƶౡٺƔพƳҠơǓƛưƓǑ¹ƌƗƬƓƶڤݜƔށ
ƟƮƌǓƛưƔ࠷މƝǔƧºƛƶƛưƷ¹טࢌǴȕǪȃȦƔమ૫ӭϱरƳƲƫ ƧƛưưNJઓ҃ƟƮƌǓº
¸Tm੯୩ƶڤү¹TMPyP4ƯƷ∆Tm = 6.0 ˚CưƲƫƧºdžƧ¹ݶӖݜƟƧҠ ݜ൝ƯƷ¹∆Tm = 3.5-6.0 ˚C ưƲǒTMPyP4 ưપƕƲތƷƲƓƫƧºƛǔǑƶ ڤүǐǒȘȦȐǞȥȮƶڤݜບưȘȦȐǞȥȮƶమࢲޒ DNA ƶϩ୩Ҡǀƶ
֒ຌƷշƔƲƌƛưƔ࠷މƝǔƧº
¸ϱरƶƛưƓǑ TMPyP4 ưݶӖݜƟƧȘȦȐǞȥȮƷƥǔƦǔ࠺ۡǴǺǾ ǨȮǫ¹ǫȦÓȑڤݜưڤݜບƷЂƲǓNJƶƶ¹ƌƢǔNJ DNA ƶഒरƯ ڤݜƟƮƌǓƛưƔǗƓƫƧºDNAƶȥȮ߱֏Ʒຜ౷੩ǚܕƌƮƒǒȘȦȐǞ ȥȮƶ়ƶҼ੩ޒưؼƌϳƳơǓºƛǔǑƶƛưƓǑ¹TMPyP4 ưݶ ӖݜƟƧ 4 ࡾ་ƶȘȦȐǞȥȮƷ¹Ƴ੩ޒƶସƝnjϳ¹֏ƶࡾ་ƶ ыƷƲƗ¹ࡶƳమࢲޒ DNA ݱԋƶȥȮ߱֏ƶാƶҼǚȘȦȐǞȥȮƶ੩ ޒƶ়ƶҼƯଇཋơǓƛưƯ¹ȥȮ߱֏௬߿ƶಶದǚۑࣺƝƣమࢲޒ DNA ǚϩ୩ҠƟƮƌǓư݂ƐǑǔƧº
Figure 12. Model of binding mode of TMPyP4 (A) and pPy, pTm, mPy and mTm (B) against CTDNA. Closed boxes denote the porphyrins.
Groove binding Self stacking
: porphyrin
A B
ફ߾च¸߾ࢲޒưǦǼǥȮȘȦȐǞȥȮưƶ۶߁ພ
ફГ২¸ࣥཉ
¸ફГचƯ֪ࢿƟƧǐƎƳ¹߾ࢲޒǚ؊Ɨϩ୩ҠơǓƛưƔƯƕǓҠݜ൝Ʒ¹
ࡿᗡਏવƶݗƌܨƔǛޱǀƶ҃ພƔ֛ગƯƕǓºdžƧ¹ƛǔǑƶҠݜ൝ƶ߾
ࢲޒǀƶ۶߁ພǚधެƳ૯ǓƛưƔƯƕǔƸ¹ǐǒພƲܨƔǛޱƶ০ڐ½ ӬದƳ໔ƮƮƌƗƛưƔƯƕǓº߾ࢲޒƳઓơǓ൝ƶڤݜưƟƮ¹ƥƶܲ
੧ƓǑǟȮǺÓǦȧÓDzȢȮ¹ӱǴǺǾǨȮǫ(external stacking)¹ǫȦÓȑ ڤݜ¹ȦÓȒǀƶڤݜƶ߾ƬƶڤݜບƔ݂ƐǑǔǓ 104)ºමݖգǚơǓҠݜ
൝ƯƷ¹߾ࢲޒƳઓơǓڤݜບƷࡶƳӱǴǺǾǨȮǫƯƊǒ¹ƥƶමݖգ
ư G-tetrad յƯƶπ-π۶߁ພƔࢲƯƊǓưƝǔƮƌǓ 57-60)ºƟƓƟƲƔǑ¹
ƥƶ੩ޒƳǐǓ߾ࢲޒǀƶ۶߁ພƳƬƌƮƶधެƲඦݥƷࣺƲƌºƥƛƯ¹ ݱԋƷ௬ƠƯ੩ޒƶЂƲǓҠݜ൝ǚພƌƮ߾ࢲޒǀƶ۶߁ພƶЊƌǚଳǁǓ ƛưƯ¹੩ޒƳǐǓ߾ࢲޒǀƶ۶߁ພƳƬƌƮڸƟƧº
¸෭चƯƷ¹G4:5'-CATGGTGGTTTGGGTTAGGGTTAGGGTTAGGGTTACCAC-3' ưƌƎ߾ӖƶȁȨȜǝ٤ǒඉƟ༞ǚƬ39 ઑƶГ෭ޒǥȥǯșÓ(G4)ǚ¹ 4 mM Tris HCl, 1 mM EDTA, 100 mM KCl pH 7.5 ƶऽڨƯǝȆÓȥȮǫƟଳৄƟ ƧNJƶǚ߹ພƟƧº߾ࢲޒƶٺƷ¹࠘ӱ CD ǚ੯୩ơǓƛưƯԏీơǓƛ ưƔƯƕǓ105-109)ºG4ƔГ෭ޒƯơǓժढњऽڨ(4 mM Tris HCl, 1 mM EDTA
pH 7.5)ƳƒƙǓǴȕǪȃȦưೝԔƟƮ¹100 mM KClǚսLjժढњଇƯƷג൲
݉߾ࢲޒƶ295 nmƳ়ƶȏÓǪƔ240 nmപؼƳാƶȏÓǪƔԏీƯƕƧ ƛưƳǐǒƥƶ߾ࢲޒٺǚԏీƟƧ(Figure 13)ºϱҟ¹4 mM Tris HCl, 1 mM
EDTA, 100 mM KCl pH 7.5ƶժढњǚພƌƮǝȆÓȥȮǫƟƧG4ǚ߹ພƟƮ¹
džƢ߾ࢲޒư TMPyP4 חƻݶӖݜƟƧ pPy¹pTm¹mPy¹mTm ưƶൖݜઑƶ ϩ୩ҠƶஶݜƌǚଳǁǓƧljƳ Tm ǚ੯୩ƟƧº࠰Ƴ¹߾ࢲޒǀƶڤݜບǚ
୩ơǓƧljƳ¹߾ࢲޒưԇҠݜ൝ưƶטࢌǴȕǪȃȦư֬CDǴȕǪȃȦ¹ ǴȕǪȃȦǚ੯୩ƟƧº
Figure 13. CD spectra of G4 in 4 mM Tris HCl, 1 mM EDTA (pH 7.5) (A) and 4 mM Tris HCl, 1 mM EDTA, 100 mM KCl (pH 7.5) (B).
-10 0 10 20
-10 0 10 20 30
200 250 300 350
Wavelength (nm)
Ellipticity (mdeg)Ellipticity (mdeg)
A
B