学位論文
Doctoral Thesis
Fibroblast growth factor-2による腱板修復促進に関する研究
(The enha nce ment o f r o ta to r cuf f tend o n -to -b o ne hea ling b y tr ea t ment w ith f ib r o b la st gr o w th facto r -2)
徳永 琢也 Takuya Tokunaga
熊本大学大学院医学教育部博士課程医学専攻運動骨格病態学
指導教員 水田 博志 教授
熊本大学大学院医学教育部博士課程医学専攻整形外科学
2015年3月
学 位 論 文
Doctoral Thesis
論文題名:Fibroblast growth factor-2による腱板修復促進に関する研究
(The enhancement of rotator cuff tendon-to-bone healing by treatment with fibroblast growth factor-2)
著 者 名 :徳永 琢也
(単名) Takuya Tokunaga
指導教員名 : 熊本大学大学院医学教育部博士課程医学専攻整形外科学教授 水田 博志 審査委員名 : 機能病理学教授 伊藤 隆明
病態生化学教授 山縣 和也 皮膚病態治療再建学教授 尹 浩信 損傷修復学講師 立石 智
2015年3月
1
目次
1. 要旨 ··· 3
2. 発表論文リスト ··· 4
3. 謝辞 ··· 5
4. 略語一覧 ··· 6
5. 研究の背景と目的 5-1. 腱板の構造と機能··· 8
5-2. 腱板断裂の臨床 ··· 9
5-3. 腱板骨結合部の発生と結合部構成細胞の特異的分子マーカー ··· 12
5-4. 腱組織幹細胞および腱組織の自然修復過程 ··· 13
5-5. 腱板自然修復過程および内因性に発現する成長因子 ··· 14
5-6. Fibroblast growth factor-2(FGF-2)とFGF-2による組織修復促進 ··· 15
5-7. 目的··· 16
6. 研究方法 6-1. 実験動物および実験デザイン ··· 18
6-2. ラット腱板修復モデルの作製 ··· 18
6-3. FGF-2の投与 ··· 19
6-4. 組織学的評価 ··· 19
6-4-1. 組織標本作製 ··· 19
6-4-2. 一般組織染色法および組織学的評価方法 ··· 20
6-4-3. 免疫染色 ··· 21
6-4-4. Section in situ hybridization ··· 22
6-4-5. 修復組織における組織学的コラーゲン線維配列とScxおよびTnmdの mRNA発現の局在との関連性評価 ··· 24
6-5. Real-time RT-PCR ··· 24
6-5-1. 修復組織からのRNA抽出とcDNA作製 ··· 24
6-5-2. Real-time RT-PCRによる解析 ··· 25
6-6. 生体力学試験 ··· 25
6-7. 統計学的解析 ··· 26
2
7. 研究結果
7-1. 組織学的評価 ··· 27
7-2. 増殖期細胞の評価 ··· 28
7-3. 間葉系幹細胞マーカーの評価 ··· 28
7-4. 腱骨結合部細胞マーカーの評価 ··· 28
7-5. 生体力学試験 ··· 31
8. 考察 ··· 33
9. 結語 ··· 36
10. 図表 ··· 37
11. 参考文献 ··· 49
3
1. 要旨
[ 目的 ] 中年以降に急増する腱板断裂は肩の疼痛や機能低下の原因となり、保存療法に抵抗する場合は 腱板修復術が行われる。しかし、術後早期の再断裂は未解決の課題であり、修復を生物学的に促進する 新たな治療法への要望は高い。先行研究においてFibroblast growth factor-2(FGF-2)の局所投与が腱板修 復を促進する可能性が示されているが、これまで腱骨結合部を構成する細胞の特異的マーカーが不明で あったため、修復過程における腱骨結合部構成細胞の参画、およびFGF-2のこれらの細胞への影響は十 分解明されていない。本研究の目的は、近年同定された腱板付着部の軟骨細胞のマーカーである転写因 子SRY-box containing gene 9(Sox9)、腱細胞のマーカーである転写因子Scleraxis(Scx)とⅡ型膜貫通型
タンパクTenomodulin(Tnmd)の腱板修復部における発現と組織学的所見および力学強度を評価し、腱
板修復過程におけるSox9、Scx、Tnmd発現細胞の参画、およびFGF-2のこれらの細胞への影響を明らか
にし、FGF-2の修復促進作用を検証することである。
[ 方法 ] 20週齢雄性Sprague-Dawleyラットの左肩棘上筋腱修復モデルにFGF-2(5μg)を含んだゼラチ ンハイドロゲル担体を腱骨間に留置したFGF-2群と、PBS含浸担体を留置した対照群を作製し比較した。
術後2、4、6、8、12週に組織を採取し、修復部の組織学的所見はスコアを用いて、力学強度は張力測定 器を用いて評価した。また、修復組織における軟骨細胞と腱細胞のマーカーとしてSox9、ScxおよびTnmd の発現をreal-time-RT-PCR、in situ hybridizationおよび免疫染色で評価した。さらに、修復部における増殖 期細胞をproliferating cell nuclear antigen(PCNA)の免疫染色、また、間葉系幹細胞をマーカー(CD73、 CD90、CD105)の免疫染色により評価した。
[ 結果 ] 術後4週以降のFGF-2群において、腱骨間の血管豊富な疎性線維性組織が減少し、密な腱様組
織による腱骨間結合を認め、6週以降の修復部の力学的強度が有意に上昇した。さらに、FGF-2群の修復 部における術後4週のSox9、4-8週のScx、および4-12週のTnmdの発現が有意に上昇した。また、FGF 群において修復組織のより広範囲に分布するScxとTnmdの発現が認められたが、両群共に結合部骨表 面におけるTnmdの発現は認められなかった。さらに、FGF-2群の術後2、4週の修復部における増殖期 細胞割合が増加し、対照群より多くの間葉系幹細胞マーカー陽性細胞が認められた。
[ 考察 ] FGF-2投与群においてScxとTnmdの発現が上昇する時期と一致して、組織学的所見の改善や修
復部の力学的強度の上昇が観察されたことから、ScxやTnmdを発現する腱系譜細胞は、腱骨結合部の修 復過程において重要な役割を担っていることが示唆された。しかし、FGF-2の担体からの放出は2週間 以内と想定されるため、術後4週以降のScxとTnmdの上昇は、FGF-2の直接的な腱細胞分化誘導では なく、初期の修復部における未分化前駆細胞集団の増加を介した作用であることが示唆された。
[ 結論 ] ラット腱板修復モデルに対するFGF-2の投与は、修復早期において腱板修復に参画する未分化 前駆細胞の増加を介して組織学的、生体力学的に腱板修復を促進することが示唆された。
4
2. 発表論文リスト 関連論文
Takuya Tokunaga, Chisa Shukunami, Nobukazu Okamoto, Tatsuya Okada, Takuya Taniwaki, Kiyoshi Oka, Hidetoshi Sakamoto, Junji Ide, Hiroshi Mizuta, and Yuji Hiraki.
FGF-2 Stimulates the Growth of Tenogenic Progenitor Cells to Facilitate the Generation of Tenomodulin-Positive Tenocytes in a Rat Rotator Cuff Healing Model.
Am J Sports Med. 43: 2411-2422, 2015.
その他の論文
Tokunaga, T., Ide, J., Arimura, H., Nakamura, T., Uehara, Y., Sakamoto, H., and Mizuta, H.
Local Application of Gelatin Hydrogel Sheets Impregnated With Platelet-Derived Growth Factor BB Promotes Tendon-to-Bone Healing After Rotator Cuff Repair in Rats.
Arthroscopy. 31: 1482-1491, 2015.
Uezono, K., Ide, J., Tokunaga, T., Arimura, H., Sakamoto, H., Nakanishi, Y., and Mizuta, H.
Effect of Postoperative Passive Motion on Rotator Cuff Reconstruction With Acellular Dermal Matrix Grafts in a Rat Model.
Am J Sports Med. 42: 1930-1938, 2014.
Uezono, K., Ide, J., Tokunaga, T., Sakamoto, H., Okamoto, N., and Mizuta, H.
Effect of immobilization on rotator cuff reconstruction with acellular dermal matrix grafts in an animal model.
J Shoulder Elbow Surg. 22: 1290-1297, 2013.
5
3. 謝辞
本研究を完遂するにあたり多くの御指導、御鞭撻を賜りました熊本大学大学院生
命科学研究部整形外科学分野の水田博志教授に深く感謝申し上げます。また、研究
全般に渡り多くの御指導、御鞭撻を賜りました京都大学再生医科学研究所生体分子
設計学分野の開祐司教授、広島大学大学院医歯薬保健学研究院基礎生命科学部門生
体分子機能学の宿南知佐教授に深く感謝申し上げます。また、本研究の遂行にあた
り多くの御助言、御指導、御支援を賜りました熊本大学医学部附属病院関節再建先
端治療学寄付講座の井手淳二特任教授、熊本大学工学部機械システム工学科の坂本
英俊教授、熊本大学大学院生命科学研究部整形外科学分野の教室の皆様に心より感
謝申し上げます。
6
4. 略語一覧
ADL : activities of daily living BMP :bone morphogenetic protein
cDNA : complementary deoxyribonucleic acid COMP : cartilage oligomeric matrix protein CTGF : connective tissue growth factor DAB : 3,3-diaminobenzide tetrahydrochloride DEPC : diethylpyrocarbonate
DIG :
digoxigenin
ECM : extracellular matrix
EDTA : ethylenediaminetetraacetic acid FGF-2 : fibroblast growth factor-2 HCl : hydrochloric acid
HE :hematoxylin and eosin
IGF :Insulin-like growth factors MSC : mesenchymal stem cell NaCl : sodium chloride PBS : phosphate buffered saline PCNA : proliferating cell nuclear antigen PDGF : platelet derived growth factor PFA : paraformaldehyde
rhFGF-2 : recombinant human FGF-2 RNA : ribonucleic acid
RNase :ribonuclease ROI : region of interest
RT-PCR : reverse transcription polymerase chain reaction
Scx : scleraxis
SD rat : Sprague Dawley rat Sox9 :SRY-box containing gene 9 SSC : standard saline citrate TBS : Tris-buffered saline
7
TE : Tris-EDTA
TEA : triethanolamine
TGF-β : transforming growth factor, beta
Tnmd : tenomodulin
tRNA : transfer ribonucleic acid TSPCs : tendon stem/progenitor cells
8
5. 研究の背景と目的
5-1. 腱板の構造と機能
腱板(rotator cuff)とは肩甲上腕関節を取り囲む肩甲下筋(subscapularis muscle)、棘
上筋(supraspinatus muscle)、棘下筋(infraspinatus muscle)、および小円筋(teres minor
muscle)の上腕骨停止部の腱性部分を指す(図1)[1, 2]。腱板の上腕骨停止部は腱組織
が線維軟骨層を介して上腕骨に付着する構造(fibrocartilagenous enthesis)を呈しており、
腱骨間の線維軟骨層はtidemarkによって非石灰化線維軟骨層と石灰化線維軟骨層にわけ
られ、軟組織と硬組織の境界を形成している(図2)[3, 4]。
腱骨付着部を形成する各層の組織学的特徴として、腱組織層ではコラーゲン線維が張
力方向に規則正しく配列し、各線維束はendotenonという膜につつまれ、腱全体はepitenon
という膜に包まれており[5, 6]、細胞成分としては紡錘形の核をもち細胞質の目立たない
腱細胞(腱線維芽細胞)が線維束に沿って配列している。また、非石灰化線維軟骨層で
は腱組織から連続するコラーゲン線維が張力方向へ規則正しく配列し、細胞成分として
は円形・卵円形の核をもつ線維軟骨細胞が線維方向に連なって配列している。さらに深
層の石灰化線維軟骨層では、石灰化した細胞外基質の間に円形核をもつ細胞質の豊富な
線維軟骨細胞が存在するが、この層では非石灰化線維軟骨層にみられるような細胞の柱
状配列はあまりみられない。また、上腕骨との境界は複雑に入り組んだパズル状の構造
を呈し、力学的に強靭な付着部を形成している[3, 5]。
腱板の機能には主動作筋としての作用と関節の安定化作用がある。主動作筋としての
9
作用は、棘上筋による上腕の外転、棘下筋や小円筋による外旋、肩甲下筋による内旋が
挙げられる。一方、関節の安定化作用としては、肩関節の動作時に各腱板構成筋が互い
に均衡して上腕骨頭を肩甲骨の関節窩に引きつけることで、動作中に骨頭中心を関節窩
に対して一定の位置に保持し、関節に動的な安定性をもたらしている[7]。
5-2. 腱板断裂の臨床
腱板断裂(rotator cuff tear)は腱板が断裂し線維の連続性が失われた状態を指す。棘
上筋腱の断裂が最も多く、そのほとんどが血流に乏しい上腕骨付着部の近傍1.5cm以内、
つまり組織学的には腱組織から非石灰化線維軟骨層の領域で断裂する[8]。一般に腱板断
裂は転倒や転落などによる肩関節下方からの外力、重量物を持ち上げる際の牽引外力、
および投球動作などの繰り返しの外力などによって生じるが[9]、中年以降では日常生活
動作の中の軽微な外力で断裂が生じることもあり、その発生の基盤に加齢に伴う腱板の
変性、すなわちコラーゲン線維の細小化および配列不整、粘液変性、硝子化変性、軟骨
化生、石灰化、血管増生、脂肪浸潤などによる生体力学的強度の低下が関係していると
考えられている[10-12]。
腱板断裂はその到達度により完全断裂(全層断裂)と不全断裂(部分断裂)に分類さ
れ、さらに、完全断裂はその大きさにより小断裂(<1cm)、中断裂(1~3cm)、大断
裂(3~5cm)、広範囲断裂(>5cm)と分類される[13]。
症状としては運動時痛に加えて安静時痛、特に夜間痛による睡眠障害をしばしば認め
10
る。また、断裂によって動作筋としての機能が低下するため、上腕の外転筋力(棘上筋
腱断裂)、外旋筋力(棘下筋腱断裂)、内旋筋力(肩甲下筋腱断裂)が低下し、上肢の
拳上や結帯動作などが困難となり、日常生活動作(ADL)に支障をきたす[14]。
腱板断裂に対する治療としては、活動性がそれほど高くない中高年者においては、腱
板の断裂により生じた二次的な変化を解消し、残存する腱板機能の維持を目的とした対
症的な保存療法が行われる[9, 14]。具体的には局所の安静、消炎鎮痛剤の内服や外用、
ヒアルロン酸製剤や副腎皮質ステロイドの関節内注射などの薬物療法によって症状の緩
和をはかり、関節機能の維持を目的としたストレッチ、可動域訓練、筋力強化などの理
学療法が行われる。これらの保存療法によって症状が軽快し、日常生活の支障が軽度と
なる症例もあるが、完全断裂例では自然治癒は起こらないとされ、腱板筋の萎縮や筋力
低下は徐々に進行することから[15, 16]、活動性の高い若年者や3から6ヶ月間の保存療
法に抵抗する症例に対しては、断裂腱板の解剖学的な修復を目的とした腱板修復術が選
択 さ れ る[14, 17]。 術 式 は 上 腕 骨 の 元 の 付 着 部 に 作 製 し た 骨 溝 に 腱 断 端 を 縫 着 す る
McLaghlin 法や、元の付着部の骨を浅く切除して縫合糸アンカーを用いて腱断端を縫着
する方法が基本となる。従来は直視下手術、あるいは関節鏡補助下ミニ・オープン法が
行われていたが、1990年代中頃より術後の肩関節機能に重要である三角筋に対してより
低侵襲である鏡視下腱板修復術が一部で行われはじめ、最近では、手術手技や手術器械
の改良に伴って、鏡視下腱板修復術が急速に普及しつつある[17]。
現 在 の 腱 板 修 復 術 に お け る 大 き な 課 題 の 一 つ と し て 術 後 の 再 断 裂 が 挙 げ ら れ る
11
[18-20]。その頻度は報告によって幅があるが、最近のエビデンスレベルⅠからⅡの文献
を集積したシステマティックレビューによると21.7%(185/861例)と比較的高頻度であ
ることが知られている[18]。また、再断裂のほとんどは術後 6 ヵ月以内の比較的早期に
起こることから[21]、その発生には不十分な修復反応による力学的な強度回復の遅延が
関係していると考えられている。そのため臨床の現場では、術後数週間にわたる装具固
定期間を設け慎重なリハビリテーションが行われ、社会復帰までに長い治療期間を要し
ている。
これらの課題を克服するために、腱板修復能を生物学的に促進し、修復部の強度の早
期回復を達成する新規の治療法が求められている。これまで、基礎研究レベルでは成長
因子や幹細胞の投与、足場材料などの一部で組織学的所見の改善や力学強度の上昇など
の有効性が示唆されている報告も散見される一方で[22-24]、目的とする効果を認めなか
ったとする報告もあり[25]、その結果は一致していない。その一因として、長年にわた
り腱板を構成する主要な細胞である腱細胞系譜の特異的な分子マーカーが判明しておら
ず、腱板修復過程に寄与する細胞成分やその制御因子に関する評価が困難であったこと
が挙げられる。そのため現状では、腱板修復後に自律的に起こる修復機転に関する分子・
細胞レベルでの理解は十分とは言い難く、従来の研究における組織学的所見の改善や力
学強度の上昇といった修復促進のメカニズムはほとんど解明されていない。より効果的
な修復促進治療の確立のためには、腱板修復過程の分子・細胞レベルでの理解を深め、
促進治療の標的因子を明確にすることが重要であると考えられる。
12
5-3. 腱板骨結合部の発生と結合部構成細胞の特異的分子マーカー
腱板と骨の結合部はマウス胚発生過程では、筋と軟骨原基の間を腱原基が連結する形
態を呈する。軟骨原基は側板中胚葉の間充織組織を由来とする軟骨前駆細胞によって胚
性12.5日(E12.5)までに形成され[26]、生後に内軟骨性骨化によって骨組織に置換され
上腕骨となる。この軟骨原基中に存在する軟骨前駆細胞、および軟骨細胞の分化マーカー遺伝子
としてHMG型転写因子であるSRY box-containing gene 9(Sox9)が知られている[26, 27]。一方、
発生過程の初期において筋と軟骨原基とを連結する腱原基もまた、側板中胚葉の間充織
組織を由来とする未分化細胞集団によって形成される。近年、この腱原基中に存在する
前駆細胞を含む腱細胞系譜の分化マーカー遺伝子として塩基性helix-loop-helix(bHLH)
型の転写因子である Scleraxis(Scx)が報告され[28, 29]、マウス四肢腱組織発生過程の
早期(E10.5)よりその発現がみられることから、腱細胞の初期分化マーカーと考えられ
ている[5, 26, 30]。また、Scxの遺伝子欠損マウスは腱組織の細胞外基質の減少や線維構
造の粗雑化といった腱組織低形成の表現型を示し、さらに I 型コラーゲン遺伝子など腱
組織形成に関連する遺伝子の発現が著明に低下、あるいは欠落することから[29]、腱発
生の初期過程において Scx は重要な役割を担っていることが示唆されている[5, 28, 29,
31]。
近年、その他の主要な腱組織特異的な分子マーカー遺伝子としてⅡ型膜貫通型糖タン
パクであるTenomodulin(Tnmd)が報告された[32-34]。マウス胚の四肢腱組織の発生過程に
おいて、Scxの発現がE10.5より上昇しE12.5をピークとしてE13.5には低下するのに対して、Tnmd
13
の発現はより後期のE13.5より急激に上昇し、出生後の成熟腱細胞においても、その発現は高いレ
ベルで持続することから[30]、Tnmdは腱組織の後期分化マーカーと考えられている[33]。
最近、これらの細胞特異的マーカーの解析により、腱付着部構造(enthesis 構造)の形成に
は腱原基や軟骨原基を形成する各々の前駆細胞集団とは区別される前駆細胞集団、すなわちScxと
Sox9 を共発現し、腱細胞や軟骨細胞への多分化能を有する前駆細胞集団が重要な役割を担って いることが明らかとなった[26, 27]。これらの結合部を構成する細胞の特異的分子マーカ
ーを用いた解析手法は、修復に寄与する細胞成分の評価、あるいは修復組織の質的評価
にも有用である可能性が示唆されている[33]。
5-4. 腱組織幹細胞および腱組織修復過程
近年、Biらは、biglycanおよびfibromodulinの遺伝子欠損マウスを用いた研究により、
成体の腱組織中に自己複製能と腱細胞への分化能を有する腱組織幹・前駆細胞(TSPCs)
が存在することを示し、さらにこのTSPCsをMatrigelに播種してマウス皮下に移植する
ことで腱様組織が誘導されることを報告した[35]。また、Dymentらは、マウス膝蓋腱欠損修
復モデルの修復過程において、損傷後1週までの初期に腱傍組織より未分化間葉系前駆細胞が修復
部に動員され、損傷後2週よりScxを発現する腱前駆細胞に分化することを報告した[36, 37]。さら
に、ラット腱板修復モデル、あるいはラット膝蓋腱欠損修復モデルの修復部にウイルス
ベクターを用いて Scx 遺伝子を導入した骨髄由来間葉系幹細胞や腱由来幹細胞を投与する
ことで、組織学的・生体力学的に組織修復を促進することが示されている[38, 39]。これ
14
らの一連の報告は、成体の腱組織の修復過程において、内在性の腱組織幹・前駆細胞に
よる修復能が限定的ながら備わっている可能性、さらに、これらの細胞集団の増殖刺激
あるいは修復部への動員が修復促進につながる可能性を示唆させるものである。一方で、
これらの報告の大部分が腱実質の修復モデル(腱-腱修復)を用いた研究であるため、腱
板修復術後(腱-骨修復)の修復過程におけるScxを発現する腱組織幹・前駆細胞の参画に
ついては未解明である。
5-5. 腱板自然修復過程および内因性に発現する成長因子
腱板自然修復過程において正常組織と同等の力学的強度の回復や組織学的に正常な構
造の再生は困難とされている[40-42]。ラット腱板自然修復過程においては、術後10日ま
でに多形核白血球、リンパ球、形質細胞、マクロファージなどの炎症細胞や、未分化細
胞および線維芽細胞の浸潤と新生血管の増生がおこり、腱骨間は疎な肉芽組織によって
充填される。術後4週で修復組織内の炎症細胞はほぼ消失し、不規則に配列したコラー
ゲン線維と骨との部分的な結合が観察される[42]。術後 8 週以降には修復組織内の細胞
密度の減少や毛細血管の減少、およびコラーゲン線維の張力方向への配列などの修復組
織の成熟の兆候が認められるが、術後16週が経過しても腱骨間には血管豊富な疎性線維
性瘢痕が介在し[42]、修復部の力学的強度は正常組織と比較して大幅に劣ることが示さ
れている[40]。
これまで、ラット腱板修復モデルの術後1週から2週にかけてFGF-2、BMP-12、BMP-13、BMP-14、
15
COMP、CTGF、PDGF-B、TGF-β1の発現が上昇すること[43]、また、ウサギ腱板(棘上筋腱)欠損
修復モデルの修復組織内の線維芽細胞にFGF-2、血管内皮細胞にPDGF-B、血球細胞に TGF-β や
IGF-1が発現することが報告されており[44]、腱板修復部において内因性に発現している様々な成長
因子が、修復の過程においてそれぞれ重要な役割を果たしていると考えられており、これらの因子
の遺伝子組み換えタンパクの投与や、遺伝子導入細胞の投与による修復促進治療への応用が試みら
れている[23, 24]。
5-6. Fibroblast growth factor-2(FGF-2)とFGF-2による組織修復促進
塩基性線維芽細胞増殖因子(basic Fibroblast growth factor: bFGF, Fibroblast growth
factor-2: FGF-2)は間葉系細胞を含む様々な細胞に対して増殖・遊走活性を有する FGF
ファミリーに属する生体内因子であり[45]、前述のように腱板修復過程の早期の修復部
で内因性に発現し、修復過程において重要な役割を果たしていると考えられている[43,
44]。FGF-2はヘパリン結合性タンパク質であり、生体内ではヘパラン硫酸を含む細胞外
基質に結合して存在し、組織損傷時に修復部へ放出され作用する[45]。
FGF-2はin vitroの実験系において、間葉系幹細胞を含む複数の幹細胞に対して分化能を維持した
まま増殖を強力に促進すること[46-48]、また、ヒト腱板由来細胞を含む様々な動物種由来の腱細胞
の増殖を強力に促進することが示されている[49]。さらに、in vivoの実験系においても、様々な組
織における再生・修復促進効果が報告されている。修復部へ局所注射した FGF-2 は 48
時間以内に消失することから[50]、修復部に活性を保持したまま持続的に投与する方法
16
として、これまでに浸透圧ポンプを用いて持続投与する方法[51, 52]や徐放担体としてヘ
パリン[53]、fibrin sealant[54]、またはゼラチンハイドロゲル[50]を用いた投与、あるいは
ウイルスベクターを用いた遺伝子導入細胞の投与[55]などが行われ、腱・靭帯組織[53, 54,
56]、骨組織[57]、軟骨組織[52, 58, 59]、歯周組織[60, 61]などに対する再生・修復促進効果が報告
されている。中でも軟骨組織[58, 59]や歯周組織の修復過程において[60]、FGF-2は修復
初期での未分化な前駆細胞の修復部への動員、あるいは増殖促進を介して修復を促進する
ことがこれまでに示されている。
腱板修復におけるFGF-2の効果に関しては、ラット腱板修復モデルに対するFGF-2含
有fibrin sealantの局所投与によって早期の組織学的所見および生体力学的強度が改善す
ることがこれまでに示されており[54]、腱板修復促進治療に応用できる可能性が示唆さ
れている。しかしながら、FGF-2 がどのようにして腱板修復過程を促進するか、特に修
復への寄与 が 想定される細胞 に及ぼす 影響に関 してはほとんど評 価されておらず 、
FGF-2の腱板修復促進治療への応用のためにはさらなる研究が必要である。
5-7. 目的
これまでの背景をまとめると、腱板修復術後の再断裂率の低下および治療期間の短縮
のために、腱板の修復を生物学的に促進する治療法が求められている。これまで、成長
因子や幹細胞の投与、足場材料などの基礎研究が行われているが、修復促進治療の標的
となりうる細胞成分が未だに不明確であるため、各々の修復促進メカニズムに関しても
17
不明な部分が多く残されている。
これまでの報告を踏まえ、FGF-2の局所投与により腱骨結合部を形成する組織幹・前
駆細胞集団が刺激されることで、腱板修復過程を促進するのではないかと考えた。そこ
で本研究では、腱板修復過程における腱組織幹・前駆細胞の修復への参画、およびFGF-2
によるこれらの細胞への影響を評価するために、近年同定された腱骨結合部を構成する細
胞特異的マーカー遺伝子を用いた解析手法に着目した。
本研究の目的は、腱板修復過程における腱骨結合部を構成する細胞特異的マーカーの
発現の評価、組織学的評価および生体力学評価によって、腱板修復過程におけるFGF-2
の作用を明らかにすることである。
18
6. 研究の方法
6-1. 実験動物および実験デザイン
実験には日本SLC(浜松)から購入した19から21週齢のSDラット(平均475
± 37
g)を合計156 匹使用した。なお、本実験は熊本大学動物実験委員会からの承認を得て行った (承認番号
B26-057)。ラット左肩腱板(棘上筋腱)急性損傷修復モデルを作製し、1肩あたり5 μgのFGF-2
(rhFGF-2, 科研製薬株式会社, 東京)を含浸させたゼラチンハイドロゲル担体(MEDGEL PI5, 株
式会社メドジェル, 京都)を腱骨間に留置したFGF-2群とPBS 含浸担体を留置した対照群(各群
78匹)を比較した。術後2、4、6、8、12週に各群、各タイムポイントにつき6肩ずつ組織学的評 価(一般染色, 免疫染色, in situ hybridization)を行い、また、各群、各タイムポイントにつき6肩ず
つreal-time reverse transcription polymerase chain reaction(RT-PCR)を行い、腱骨結合部細胞関連マー
カー遺伝子の発現を評価した。また、術後6、12週に各群、各タイムポイントにつき9肩ずつ生体
力学試験を行った。
6-2. ラット腱板修復モデルの作製
ラット腱板(棘上筋腱)急性損傷修復モデルはThomopoulosらの報告[42]に従って作製した。19
から20週齢雄のSDラットにペントバルビタール(25-30 mg/kg)の腹腔内投与による全身麻酔導
入後、左肩直上に2 cmの皮膚切開を加え、三角筋を線維方向に割いて棘上筋腱を露出し、メスを
用いて上腕骨付着部で鋭的に切断した。その後、付着部に残存した軟部組織および線維軟骨層を高
速回転ドリル(ProxxonミニルーターセットMM100, キソパワーツール, 大阪)を用いて、骨髄か
19
らの出血が観察されるまで完全に除去した後、腱断端にMason-Allen変法で5-0プロリン糸をかけ
て元の位置に修復した。その際、FGF-2 もしくは PBS を含浸させたゼラチンハイドロゲル担体
(MEDGEL PI5)を腱骨間に留置し、三角筋、皮下、皮膚を4-0ナイロン糸で縫合した。術後は外
固定を行わずcage activityを許可した。組織は術後2、4、6、8、および12週に肩甲骨と上腕骨を一
塊として摘出した。
6-3. FGF-2の投与
実験には科研製薬株式会社より提供されたrhFGF-2(recombinant human FGF-2)を使
用した。また、FGF-2 の投与担体としてメドジェル株式会社(京都)より購入した生体
吸収性ゼラチンハイドロゲルシート(
MedGel PI5
)を使用した[50]。投与前に、各肩あたり1 mgのシートに、500 μg/mLに調整したFGF-2溶液を 10 μl(1肩あたり5 μg)含浸
させ、室温で60分培養した後、修復部に留置した[50]。対照群では、PBSのみを含浸、
培養後に修復部に留置した。
6-4. 組織学的評価 6-4-1. 標本作製
ペントバルビタール(25-30 mg/kg)の腹腔内投与により深麻酔を導入後に、灌流ポンプ
(FPC-100, アズワン, 大阪)を用いて4%PFAで灌流固定後に組織を摘出し、4%PFAを用いて4°C
で16 時間の後固定を行った。次に、組織をエタノール系列で脱脂した後に、モース氏液(10%ク
エン酸, 22.5%ギ酸)に浸して4°Cで約7日間脱灰した。脱灰後、組織はエタノール系列による脱水、
20
およびキシレンによる透徹後にパラフィン包埋した。肩関節棘上筋腱から上腕骨修復部を中心に4
µmの冠状断の組織切片を作製し各染色に使用した。組織標本作製に使用した溶液の希釈には、
RNase free水もしくはDEPC処理水を使用した。
6-4-2. 一般組織染色法および組織学的評価方法
修復部の組織学的所見はヘマトキシリン・エオジン(HE)染色およびpicrosirius-red染色し
た組織切片を、光学顕微鏡(Olympus BX-51, オリンパス, 東京)で観察し、 画像はデジタルカ
メラ(DP70, オリンパス)で撮影し、画像解析ソフト(Image J, National Institutes of Health,
Bethesda, USA)を用いて評価した。
各組織切片における修復部の組織学的成熟度は、これまで報告されている修復部の細
胞密度、血管数、コラーゲン線維配列の3項目による評価法[62-65]を一部改編したスコ
アリングシステム(合計12点満点)(表1)を用いて半定量的に評価した[66]。すべて
の項目で連続3切片を評価し、観察者バイアスを避けるため各切片の実験群およびタイ
ムポイントを盲目化して評価した。
細胞密度は倍率100倍で観察したHE染色切片上の腱骨移行部領域(骨表面から100-400 μm
近位)から無作為に3箇所の関心領域(200 × 200 μm)を選択し、領域内のすべての有核細胞をカ
ウントし、平均細胞密度(cells/mm2)を算出した。また、各切片の値は正常肩組織(n=6)切片上
の腱骨移行部領域で同様にカウントして算出した値を100%として標準化して表示した。
血管数は倍率100倍で観察したHE染色切片上の腱骨移行部領域内(骨表面から100-400 μm
近位)のすべての血管をカウントし、平均血管数を算出して記録した。
21
修復部のコラーゲン線維配列は、倍率12.5倍で偏光顕微鏡下に撮影したpicrosirius-red染色画
像を用いて評価した[38, 64, 66, 67]。この画像では、コラーゲン線維が一定方向に配列した領域で
は回折光により明領域として観察され、この画像をImage Jで8-bit Gray Scale画像(黒色領域, 0;
白色領域, 255)に変換すると、明領域は白色領域(Gray Scale高値)として観察されるため、こ
のGray scale値を指標として切片上の修復組織のコラーゲン線維配列を評価した。腱骨移行部領域
(骨表面から100-400 μm)から無作為に10箇所の関心領域(100 × 100 m)を選択し、Image
Jを用いて各領域のGray scale値を測定し、10箇所の平均値を算出して記録した。測定誤差を減
少させるために、すべての切片を回転台上で360度回転し、最も回折光が観察される位置で画像を
撮影した。また、各切片の値は同様の条件で撮影した正常肩組織(n = 6)切片の値を100%として
標準化して表示した。
6-4-3. 免疫染色
腱骨修復部における増殖期細胞をPCNAの免疫染色、またSox9発現細胞を免疫染色により評価
した。また、間葉系幹細胞は陽性マーカーとしてCD73、CD90、CD105の発現を、陰性マーカー
(血球系細胞マーカー)として CD34、CD45 の発現を免疫染色により評価した[68]。マウス抗
PCNA モノクローナル抗体(clone PC10)は Dako(Copenhagen, Denmark)から、ウサギ抗
Sox9 ポリクローナル抗体(AB5535)とマウス抗 CD105 モノクローナル抗体(#05-1424)は
Millipore(Billerica, USA)から、ウサギ抗CD73ポリクローナル抗体(AB175396)とマウス抗
CD90モノクローナル抗体(AB225)とウサギ抗CD34ポリクローナル抗体(AB 64480)とウサ
ギ抗CD45ポリクローナル抗体(AB10558)抗体はAbcam(Cambridge, UK)からそれぞれ入手
22
した。二次抗体としてラット組織用ヒストファインMAX-POをニチレイバイオサイエンス(東京)
から入手した。
組織切片(4 μm)をキシレンで脱パラフィンし、希釈エタノール系列で親水化した後、抗原賦活
化処理のため切片を緩衝液(Target Retrieval Solution, pH9.0, DAKO)に浸し60°Cで16時間加温した。
切片をメタノールで希釈した0.3%過酸化水素に室温で10分間浸して内因性ペルオキシダーゼ活性
阻害処理を行った。PBSによる洗浄後、10%正常ヤギ血清(ニチレイバイオサイエンス)を用いて
室温で30分間、非特異的結合阻害処理を行った。PBS洗浄後、1次抗体(PCNA; 1:200, Sox9; 1:200,
CD73; 1:200, CD90; 1:200, CD105; 1:50, CD34; 1:100, CD45; 1:1000)を4°Cで16時間反応させた。PBS
による洗浄後、2次抗体は1次抗体の動物種に応じてヒストファインMAX-PO(M)もしくはヒス
トファインMAX-PO(R)を用いて、室温で30分間反応させた。次にDAB溶液(同仁化学, 熊本)
に浸し室温で3分間の発色反応を行い、0.5%マイヤーヘマトキシリンで対比染色を行い、脱水、透
徹、封入処理の後、光学顕微鏡で観察した。陰性コントロールは、1次抗体を使用せずに同様の処
理を行った切片を評価した。
PCNAおよびSox9の発現は倍率200倍で観察した連続3切片上の腱骨移行部領域(骨表面から 100-400 μm近位)から無作為に3箇所の関心領域(200 × 200 μm)を選択し、領域内の全ての
有核細胞および陽性細胞をカウントし陽性細胞率を算出して半定量的に評価した[58]。
また、術後2週、4週の修復早期の間葉系幹細胞の評価として陽性マーカー(CD73、CD90、
CD105)の発現と、陰性マーカー(CD34、CD45)の発現を連続切片の免疫染色により評価した。
23
6-4-4. Section in situ hybridization
組織切片におけるScxとTnmdのmRNAの局在はDIG標識したRNAプローブを用いたin situ
hybridizationの手法で評価した[69]。組織切片(5 μm)をキシレンで脱パラフィンし、希釈エタノー
ル系列で親水化した後、TE緩衝液(10mM Tris-HCl pH 8.0, 1mM EDTA)で20 μg/mlに調整した
proteinase K(Gibco BRL, Rockville, USA)を用いて室温で10分間処理し、4%PFAに浸し室温で10 分間再固定した。PBSで洗浄後に0.2 M HClで10分間処理し、再度PBSで洗浄後にアセチル化処
理として0.25%無水酢酸含有0.1 M TEAに室温で10分間浸した。その後PBSによる洗浄、希釈エ
タノール系列による脱水、風乾を行った。
ScxとTnmdに対するDIG標識RNA probe(sense probe, antisense probe)はDIG RNAラベリング キット(Roche, Mannheim, Germany)を用いてin vitro転写反応により作製した。京都大学再生医科
学研究所、生体分子設計学分野で作製されたcDNA配列(rat Scx, 726 bp; rat Tnmd, 607bp)を含むプ
ラスミド(pBSII SK+, Stratagene, La Jolla, USA, pCRII-TOPO, Invitrogen, Carlsbad, USA)を制限酵素で
直線化して鋳型として用いた。作製したprobeを溶解したhybridization buffer(50%ホルムアミド, 10
mM Tris-HCl, 1×Denhardt溶液, 10%Dextran sulfate, 600 mM NaCl, 0.25%SDS, 1 mM EDTA [pH 8.0], and 200 μg/ml tRNA)をサンプルに滴下後、HybriSlipTM (Sigma-Aldrich, St Louis, USA)で被覆し、50°C
で16時間hybridizationを行った。その後、切片を50%ホルムアミド含有2倍SSCに浸して62°Cで
40分間洗浄し、TNE緩衝液中で20 μg /mlに調整したRNase A溶液(Sigma-Aldrich)に浸し、37°C で30分間培養した。その後、2倍SSCおよび0.2倍SSCで62°Cで40分ずつ洗浄した。probeの発
色は抗DIG-AP Fab fragment抗体(Roche)とBM Purple(Roche)を用いて行った。陽性コントロー
24
ルとしてマウス胚上肢(E16.5)切片を用い、また、陰性コントロールとして、各sense probeを使
用した切片を用いた。
6-4-5. 修復組織における組織学的コラーゲン線維配列とScxおよびTnmdのmRNA発現の局在との
関連性評価
修復組織のコラーゲン線維配列とScxおよびTnmdの発現の関連性を評価するため、術後8週の両群
のpicrosirius-red染色切片とin situ hybridizationの切片上の修復部を9箇所の領域(関節側、中間層、滑
液包側/腱実質、縫合部周囲、腱骨移行部)に分割し(図9C)、各領域におけるコラーゲン線維配
列とScxおよびTnmdの発現の相関を評価した(n = 12)。各領域における組織学的コラーゲン線維配
列は6-4-2.に記載した方法で評価した。また、ScxおよびTnmdの発現はhybridization signalの染色性に
より、(1)陰性、(2)弱陽性、(3)中等度陽性、(4)強陽性の4段階のグレードを用いて半定量的
に評価し[40, 42]、各領域で最も高い段階を記録した。
6-5. Real-time RT-PCR
6-5-1. 修復組織からのRNA抽出とcDNA作製
すべてのタイムポイントにおいて腱骨間の修復組織を採取し、液体窒素で凍結した後、マルチビ
ーズショッカー(安井機器, 大阪)を用いて破砕した。総RNAはTRIzol Plus RNA Purification Kit
(Invitrogen)を用いて抽出、精製した。精製したRNA(1 μg)を用いてThermoscript RT kit(Invitrogen)
により逆転写反応を行い、cDNAを作製した。
25
6-5-2. Real-time RT-PCRによる解析
Real-time PCRによる解析はABI7500 system(Applied Biosystems, Foster, CA, USA)を用いて行っ た。TaqMan Universal PCR Master Mixおよび各遺伝子に対するプライマー(TaqManTM Gene Expression
Assays, Scx [Assay ID: Rn01504576_m1], Tnmd [Rn00574164_m1], Sox9 [Rn01751069_mH], 18S ribosomal RNA [Rn03928990_g1])はApplied Biosystemsより購入した。50°Cで2分間、95°Cで10
分間の反応の後、95°Cで15秒、60°Cで1分間反応させるプロトコールで40サイクルのPCR反応
を行った。各遺伝子の発現量は18S ribosomal RNAを内因性コントロールとしてΔΔCt法を用いて定
量し、正常棘上筋腱における発現を1として表示した[70]。
6-6. 生体力学試験
生体力学試験は、熊本大学工学部研究棟Ⅰ実験室の小型張力測定器(STA1225, オリエンテック
社, 東京)を用いて行った。上腕骨から肩甲骨を一塊として採取した検体は、試験まで-80°Cで保存
し、試験前に37°Cで解凍した後、肩甲骨、棘上筋、縫合糸表層の修復組織および縫合糸を除去し、
修復部幅が2 mmとなるように検体を標準化した[54, 64-66]。次に、張力測定器の万力に紙やすりを
貼付し上腕骨付着部から3mm近位で腱を固定した後、張力負荷の方向に腱が平行になるように上
腕骨をもう一方の万力に固定した。全ての検体は0.1 Nの張力で前負荷をかけ、10 mm/minの速度
で破断するまで負荷をかけた。また、破断部位を記録し、検体の幅および厚みをデジタルマイクロ
キャリパー(DT-100, 新潟精器, 新潟)を用いて測定し、断面積(mm2)を長方形に近似して算出
した。コンピューターに自動記録された荷重変位曲線より最大破断強度(N)、剛性(N/mm)、最
26
大破断応力(N/mm2)を算出した。正常肩対照群として、右肩正常肩(n = 9)について同様に評価
した。
6-7. 統計学的解析
全てのデータは平均値 ± 標準偏差として表示した。各データのFGF-2群と対照群間の比較には
Mann-Whitney U test.を用いた。また、各データの各群内のタイムポイント間の比較には
Kruskal-Wallis testを行い、post hoc testとしてScheffe testにより多群間を比較した。また、修復組織 の各領域におけるコラーゲン線維配列とScxまたはTnmd発現の相関の検定にはスピアマンの順位
相関係数を用いた。全ての統計学的解析には、Stat View version 5.0(SAS Institute Inc., Cary, NC, USA)
を使用し、有意水準5%未満を統計学的に有意とした。
27
7. 研究結果
7-1. 組織学的評価
術後2週では両群において血管と細胞が豊富で疎な線維性肉芽組織が腱骨間に認められ、偏光顕
微鏡下に観察した picrosirius-red 染色切片の修復組織内に回折光領域はほとんど観察されなかった
(図3F, P)。術後の経過に伴って、修復部の細胞密度の減少や、回折光領域の増加など修復組織成
熟の兆候が認められたが、対照群では12 週経過後も腱骨間には血管が豊富で配列が不規則な線維
性組織が介在していた(図3e)。一方、術後4週以降のFGF-2群では、対照群と比較してこれらの
疎性線維性組織は明らかに減少し、腱骨移行部の修復組織内により多くの回折光領域が認められた
(図3)。観察期間内において、両群共に、線維軟骨層からなる正常の付着部構造は再生されなかっ
た。
スコアを用いた組織学的成熟度の項目別評価では、術後8週、12週の血管数がFGF-2
群において対照と比較して有意に少なく(8週, P = .020; 12週, P = .013)(図4B)、
また、術後4週以降のコラーゲン線維配列がFGF-2群において有意に高値であった(4
週, P = .007; 6週, P = .025; 8週, P = .025; 12週P = .025)(図4C)。術後2、4、6、8、
および12週の組織学的スコアは FGF-2 群で、3.2±0.4、4.8±0.4、6.3±1.0、8.0±1.7、
9.3±1.2、対照群で、3.0±0.0、4.0±0.6、5.2±0.4、5.8±0.4、7.2±1.0 であり、両群
共に経時的に増加し、術後4週以降のFGF-2群において対照群と比較して有意に高値で
あった(4週, P = .045; 6週, P = .037; 8週, P = .013; 12週P = .013)(図4D)。
28
7-2. 増殖期細胞の評価
修復組織における増殖期細胞はPCNAの免疫染色により評価した。修復後早期(2、4週)のFGF-2
群において対照群より多数のPCNA陽性細胞が認められた(図5A, B, F, G)。術後6週以降では、
両群ともに PCNA 陽性細胞は明らかに減少し、主に修復部の上腕骨表面の領域に観察された(図
5C, D, E, H, I, J)。PCNA陽性細胞率は術後2、4週においてFGF-2群で対照群と比較して有意に高
値であった(2週, FGF-2群, 45.4 ± 9.3%vs. 対照群, 30.8 ± 5.7%; P = .016; 4週, FGF-2群, 41.9 ± 8.0%vs.
対照群, 29.0 ± 7.4%; P = .025)(図5K)。術後6、8、12週では両群間に有意な差はみられなかった(6
週, FGF-2群, 23.8 ± 5.3%vs. 対照群, 19.7 ± 2.7%; P = . 423; 8週, FGF-2群, 22.3 ± 5.7%vs. 対照群, 19.7
± 2.7%; P = . 263; 12週, FGF-2群, 19.6 ± 5.3%vs. 対照群, 19.0 ± 3.8%; P = .749)(図5K)。
7-3. 間葉系幹細胞マーカーの評価
修復部における間葉系幹細胞は連続切片を用いた免疫染色で評価した。陽性マーカーとして
CD73、CD90、CD105、また、陰性マーカーとしてCD34およびCD45を評価した。術後2、4週の
FGF-2群の修復組織において、対照群と比較してより多くのCD73、CD90、CD105陽性細胞を認め
た(図6K-M)。両群共に観察期間内の修復部において、CD45陽性細胞はほとんど認められず(図
6E, J, O, T)、CD34陽性細胞は血管周囲以外にほとんど認められなかった(図6D, I, N, S)。
7-4. 腱骨結合部細胞マーカーの評価
修復部における腱細胞の評価としてScx、Tnmdの発現を、修復組織を用いたReal-time RT-PCRお
よびSection in situ hybridizationにより解析した。Real-time RT-PCRによるScxの発現解析では、術後
29
4週(FGF-2群, 1.8 ± 0.8 vs. 対照群, 0.8 ± 0.3; P = .010)、6週(FGF-2群, 3.3 ± 1.5 vs. 対照群, 1.2 ± 0.4;
P = .010)、8週(FGF-2群, 1.6 ± 0.3 vs. 対照群, 1.0 ± 0.1; P = .004)のFGF-2群において対照群と比較
して有意に高値であった。2週(FGF-2群, 1.1 ± 0.5 vs. 対照群, 1.1 ± 0.5; P = .631)と12週(FGF-2
群, 1.2 ± 0.4 vs. 対照群, 0.8 ± 0.2; P = .150)では両群間に有意な差はなかった(図7A)。
In situ hybridizationではScxに対するhybridization signalは両群のすべての観察時期の腱実質の紡錘 形細胞に認められた(図8A, E, I, M, Q, U)。腱骨移行部の修復組織においては4週以降の縫合糸周
囲の紡錘形・卵円形細胞にわずかに認められた。対照群の腱骨間の血管豊富な線維性修復組織内の
細胞ではScxに対するhybridization signalはほとんど認められなかったが(図8C, G, K)、4週以降の
FGF-2群の上腕骨表面の紡錘形細胞でScxのhybridization signalが検出された(図8S, W)。
Tnmdの発現はReal-time RT-PCRでは、術後4週(FGF-2群, 2.1 ± 0.7 vs. 対照群, 0.9 ± 0.4; P = .004)、
6週(FGF-2群, 4.7 ± 1.6 vs. 対照群, 1.6 ± 1.4; P = .010)、8週(FGF-2群, 3.7 ± 0.8 vs. 対照群, 1.8 ± 0.2;
P = .004)、および12週(FGF-2群, 2.1 ± 0.7 vs. 対照群, 1.2 ± 0.2; P = .016)のFGF-2群において対照
群と比較して有意に高値であった。2週(FGF-2群, 0.9 ± 0.8 vs. 対照群, 0.7 ± 0.6; P = .631)では両
群間に有意な差はなかった(図7B)。
Tnmdに対するhybridization signalは、Scxと同様に両群のすべての観察時期の腱実質の紡錘形細
胞に認められた(図8B, F, J, N, R, V)。対照群では、縫合糸周囲から骨移行部の疎な線維性組織にか
けてそのsignalはほとんど検出できなかったのに対して、FGF-2群ではTnmdのsignalの分布が縫
合糸の周囲まで認められた。しかし腱骨移行部の骨表面の領域では、両群共に観察期間内において
Tnmd のsignalは認められなかった(図8D, H, L, P, T, X)。
30
次に、修復組織における組織学的なコラーゲン線維配列とScxおよびTnmdのmRNA発現の局在
との関連性を評価するため、術後8週の両群のピクロシリウスレッド染色切片とin situ hybridization
切片上の修復部を9箇所の領域(関節側、中間層、滑液包側/ 腱実質、縫合部周囲、腱骨移行部)
に分割し(図9C)、各領域(合計108領域)におけるコラーゲン線維配列とScxおよびTnmdの発
現の相関を評価した(図9D, E)。各領域におけるScxの発現とコラーゲン線維配列との間には、弱
い正の相関が認められた(スピアマンの順位相関係数 [ρ] = .27, P < .001)(図9D)。一方、Tnmdの
発現とコラーゲン線維配列との間には、強い正の相関が認められた(ρ = .88, P < .001)(図9E)。
修復部におけるSox9の発現をReal-time RT-PCRと免疫染色により評価した。Real-time RT-PCRで
は、術後4週(FGF-2群, 1.8 ± 0.4 vs. 対照群, 0.5 ± 0.4; P = .004)のFGF-2群において対照群と比較
して有意に高値であった。2週(FGF-2群, 0.9 ± 0.4 vs. 対照群, 0.6 ± 0.4; P = .337)、6週(FGF-2群, 1.8
± 0.3 vs. 対照群, 1.7 ± 0.6; P = .749)、8週(FGF-2群, 2.4 ± 0.8 vs. 対照群, 3.1 ± 1.3; P = .200)、および
12週(FGF-2群, 2.1 ± 0.6 vs. 対照群, 2.4 ± 0.5; P = .337)では両群間に有意な差はなかった(図10A)。 免疫染色では、Sox9の発現は主に腱骨間の修復組織の卵円形核を有する細胞に認められたが、腱
実質部から縫合糸周囲にかけて存在する卵円形、紡錘形核を有する細胞にもわずかに認められた
(図10B-G)。修復部におけるSox9陽性細胞率は、術後4週(FGF-2群, 23.7 ± 4.9% vs. 対照群, 14.0
± 3.6% P = .010)のFGF-2群において対照群と比較して有意に高値であった。2週(FGF-2群, 13.4 ±
6.6% vs. 対照群, 10.1 ± 4.8%; P = .337)、6週(FGF-2群, 22.3 ± 5.7% vs. 対照群, 19.7 ± 2.7%; P = .263)、
8週(FGF-2群, 11.2 ± 1.5% vs. 対照群, 10.3 ± 1.1%; P = .423)、および12週(FGF-2群, 11.3 ± 3.4% vs.
対照群, 10.1 ± 2.6%; P = .729)では両群間に有意な差はなかった(図10H)。
31
7-5. 生体力学試験
正常肩9検体および術後6、12週において各群9検体ずつ評価した。正常肩9検体中、3検体が
腱骨付着部で剥離し、残りの6検体は腱実質部で破断した。FGF-2群、対照群の検体は全て腱骨間
の修復部で破断した。正常の腱骨付着部の最大破断強度は42.0 ± 5.56 N、剛性は27.0 ± 5.56 N/mm、
断面積は2.09 ± 0.13 mm2、最大破断応力は19.7± 2.44 N/mm2であった。
術後6週の修復部の最大破断強度(FGF-2群, 15.05 ± 3.32 N vs. 対照群, 7.92 ± 2.54 N, P =.009)、剛
性(FGF-2群, 7.16 ± 3.28 N/mm vs. 対照群, 3.43 ± 1.24 N/mm, P =.005)、最大破断応力(FGF-2群, 4.24
± 0.96 N/mm2 vs. 対照群, 2.34 ± 0.93 N/mm2, P =.001)はFGF-2群で対照群と比較して有意に高値であ った。断面積(FGF-2群, 3.56 ± 0.35 mm2 vs. 対照群, 3.53 ± 0.51 mm2, P =.627)は両群間に有意な差
は認めなかった(図11)。
術後12週の最大破断強度(FGF-2群, 23.8 ± 4.61 N vs. 対照群, 13.8 ± 4.69 N, P =.003)、最大破断応
力(FGF-2群, 6.92 ± 1.10 N/mm2 vs. 対照群, 3.88 ± 1.12 N/mm2, P =.003)はFGF-2群で対照群と比較
して有意に高値であった。剛性(FGF-2群, 8.98 ± 3.58 N/mm vs. 対照群, 6.83 ± 1.25 N/mm, P =.310)、
断面積(FGF-2群, 3.42 ± 0.28 mm2 vs. 対照群, 3.53 ± 0.26 mm2, P =.354)は両群間に有意な差は認め
なかった(図11)。
32
8. 考察
本研究ではラット肩腱板(棘上筋腱)急性損傷修復モデルを用いて、FGF-2の局所投与の修復部
における組織学的所見、増殖期細胞、間葉系幹細胞、腱骨結合部関連細胞マーカー、力学強度への
影響について組織学的評価、免疫組織学的手法、real-time RT-PCR、in situ hybridization、生体力学試
験の手法を用いて評価した。
ラット腱板急性損傷修復モデルの自然修復過程では、術後 10 日目までに炎症細胞と未分化間葉
系細胞の浸潤と新生血管の増生がおこり、腱骨間は修復性肉芽組織によって充填される。経過に伴
い修復組織内の細胞密度と毛細血管は減少し、部分的にコラーゲン線維と骨との結合や力学方向へ
の配向性がみられるが、16週が経過しても血管網に富む疎性線維性組織が腱骨間に介在し、正常組
織は再生しない[42]。本研究のPBS含浸担体を留置した対照群では、これまでの報告と同様に、術
後12週が経過しても腱骨間には疎な線維性組織が介在していた(図3e)。一方、FGF-2群では、術
後4週より腱骨間の疎な線維性組織の減少とコラーゲン線維配列の改善を認め、4週から12週の組
織学的成熟度スコアは対照群と比較して有意に高値であった(図 4D)。また、生体力学試験では、
術後12週におけるFGF-2群の最大破断強度、剛性、最大破断応力はそれぞれ、健常組織の56.7%、
33.3%、35.1%と観察期間において正常水準の強度回復には至らなかったが、術後6、12 週ともに
修復部の力学的強度はFGF-2群で有意に高値であった(図11)。腱骨間に形成される血管が豊富で
不規則に配列した線維から構成される組織は、力学負荷方向に配列したコラーゲン線維から構成さ
れる組織より脆弱であることが過去の大型動物モデル(ヒツジ棘下筋腱修復モデル)を用いた研究
で示されており[71]、これは本研究におけるFGF-2投与による修復部の力学強度の上昇と、同部位
33
の組織学的な線維性瘢痕の減少、およびコラーゲン線維配列の改善との関連性を示唆させる結果と
考えられる。
これまでScx発現細胞の増加が、修復過程において配列したコラーゲン線維の形成を促進し、力
学的強度に優れた腱様組織の形成を促す可能性が示されている。GulottaらはScxをウイルスベクタ
ーにより導入した骨髄由来間葉系幹細胞をラット腱板修復モデルに投与することで、術後4週にお
いて組織学的、生体力学的に修復を促進することを報告し[38]、また、TanらはScx導入腱由来幹細
胞をラット膝蓋腱損傷モデルに投与することで腱の修復を促進することを示した[39]。マウス胚の
四肢の腱発生における腱前駆細胞から成熟腱細胞への分化の過程では、Scxの発現はE10.5より上
昇し、E12.5を発現のピークとしE13.5には低下する。一方、Tnmdの発現はE13.5より急激に上昇
し、出生後も腱細胞における発現は持続する[30]。さらに、このScxとTnmdの時間的な発現パター
ンはマウス膝蓋腱の修復過程においても同様に確認されている[70]。本研究では、FGF-2 の投与に
より、術後4週以降の修復組織内により多くのScx発現細胞がみられ、術後4週から8週の修復部
におけるScx発現が上昇することを示した(図7A)。これらの結果は、本モデルの修復過程におい
て、より多くのScx発現腱前駆細胞が修復部に出現したことを示唆する。さらに、FGF-2群の術後
4週から12週にわたる持続的なTnmd発現上昇(図7B)もまた、より多くのTnmd発現細胞が修復 へ寄与したことを示唆する結果と考えられる。Tnmd 陽性の紡錘形細胞は、コラーゲン線維が高度
に配列した領域に一致して観察され、特に修復中期以降のFGF-2群において、Tnmd陽性細胞の分
布は縫合糸近傍の修復組織内へと拡大し、その発現は組織学的にコラーゲン線維が配列している領
域と一致していた(図9E)。これらの修復過程における空間的なTnmdの発現パターンは、腱発生
34
過程の後期でみられる発現パターンと類似していることから[33, 72]、Tnmdは腱修復過程において
も腱細胞分化および修復組織の質的評価に有用なマーカーであることが示唆される。
本研究で用いたゼラチンハイドロゲル担体からのFGF-2の放出は約2週間であることが示されて
いることから[50]、FGF-2群での修復中期(4週)以降のScxおよびTnmdの発現の上昇は投与した
FGF-2による直接的な腱細胞系譜への分化促進作用ではないと考えられる。本実験では、FGF-2群
の修復初期において修復組織内のPCNA陽性細胞率が増加していること(図5)から、FGF-2によ
る腱様組織形成は、修復早期の組織幹細胞、未分化前駆細胞の増殖促進を介した作用であることが
示唆される。最近、Dymentらはマウス膝蓋腱修復モデルの修復過程において、まず損傷後1週ま
でに腱傍組織より未分化間葉系前駆細胞が修復部に動員され、損傷後2週頃よりScx発現腱前駆細
胞が修復部に出現することを示した[36, 37]。この事実は、初期の未分化間葉系前駆細胞の修復部へ
の出現が、後の修復組織における腱細胞への分化と密接に関係していることを示唆するものである。
また、これまでの細胞培養系による研究において、FGF-2は間葉系幹細胞および前駆細胞の分化能
を維持したまま増殖を強力に促進することが示されている[46-48]。さらに、FGF-2の局所投与は動
物モデルを用いた骨、軟骨、および歯周組織の修復の初期過程において組織幹細胞や前駆細胞の増
殖や遊走を刺激することで再生の誘導、あるいは組織修復を促進することが報告されている[58, 60]。
これらの一連の報告と、本実験でのFGF-2の投与による早期の修復組織内のMSCマーカー発現細
胞の増加(図6)という結果から、FGF-2による腱板修復の促進は、修復初期における組織幹・前
駆細胞の増殖刺激を介した作用であることが示唆された。
本研究にはいくつかの考慮すべき限界が存在する。まず、本研究でのラット棘上筋腱の急性断裂
35
修復モデルは、臨床における断裂後の経過による棘上筋の萎縮や腱の変性などの病態を反映してい
ない。また、健常な小型動物と臨床上の患者との治癒能や修復機転は同様であるとは言い難く、今
後、陳旧性断裂モデルなどの病態モデルやより大型の動物モデルによる検討の余地が残されている。
しかし、本研究に用いたラットモデルは、ヒトと解剖学的に類似していることから修復機序の解明
や修復の促進に関する研究に広く用いられている有用なモデルである[40, 42, 54, 62, 63]。次に、本
研究の組織学的評価では、サンプルの一部の切片から領域を抽出するため、サンプリングバイアス
の存在は否定できない。しかしながら、生体力学評価や分子生物学的評価を含めた複数の評価法を
用いて得られた結果は、概ね整合性が取れており、本実験においてFGF-2の一定の効果は示されて
いるものと考えられる。
本研究ではFGF-2が特定の前駆細胞の増殖を選択的に促進するかは明らかとなっていないが、本
モデルのFGF-2群において中期以降にみられたScxとTnmdの発現の上昇は、FGF-2が修復早期の
修復部における未分化前駆細胞の増殖を刺激し、中期以降の腱系譜細胞の増加に有利な微小
環境をもたらしたものと考えられた。さらに、注目すべき所見として、FGF-2群の術後4週におい
て一過性にみられたSox9の発現上昇がある。これは腱付着部構造の形成に重要な役割を持つSox9
とScxを共発現する前駆細胞集団が[27]、修復過程においても限定的に参画している可能性を示唆
する結果である。本研究により、修復早期に出現する前駆細胞(Scx 発現前駆細胞)の刺激が修復
促進につながる可能性、すなわちScx発現前駆細胞が修復促進治療の標的となる可能性が示唆され
た。腱板修復に寄与する前駆細胞集団の特徴の理解やそれらの標的細胞の動員、増殖、分化の制御
機構のさらなる研究は、より効果的な生物学的腱板修復促進治療の確立に有益であると考えられる。
36
9. 結語
ラット腱板修復モデルに対するFGF-2の投与は、修復早期において腱骨部の修復に参 画する未分化前駆細胞の増殖刺激を介して組織学的、生体力学的に腱板修復を促進する ことが示唆された。
37
10. 図表
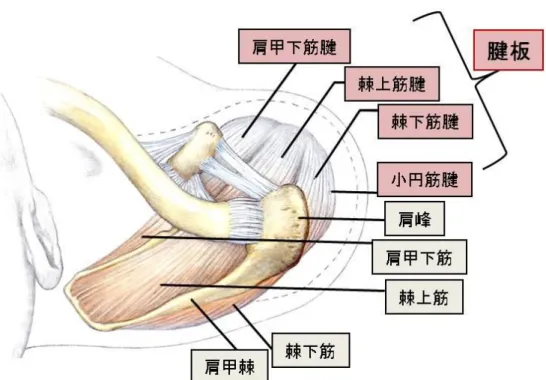
図1. 肩甲上腕関節と腱板の構造
腱板(rotator cuff)とは、肩甲上腕関節の周囲を取り囲む肩甲下筋(subscapularis muscle)、
棘上筋(supraspinatus muscle)、棘下筋(infraspinatus muscle)、および小円筋(teres minor
muscle)の上腕骨付着部近傍の腱性部分を指す(文献2より改変引用)。
38
図2. 腱板骨付着部の構造
腱板の上腕骨停止部は腱組織が線維軟骨層を介して上腕骨に付着する構造(fibrocartilagenous
enthesis)を呈している(A)。腱組織ではコラーゲン線維が張力方向に規則正しく配列し、
各線維束はendotenonという膜につつまれ、腱全体はepitenonという膜に包まれており、
細胞成分としては紡錘形の核をもち細胞質の目立たない腱細胞(線維芽細胞)が線維束 に沿って配列している(B)。また、非石灰化線維軟骨層では腱組織から連続するコラ ーゲン線維が張力方向へ規則正しく配列し、細胞成分としては円形・卵円形の核をもつ 線維軟骨細胞が線維方向に連なって配列している。さらに深層の石灰化線維軟骨層では、
石灰化した細胞外基質の間に円形核をもち細胞質の豊富な線維軟骨細胞が存在するが、
この層では非石灰化線維軟骨層にみられるような細胞の柱状配列は目立たない。また、
上腕骨との境界は複雑に入り組んだパズル状の構造を呈し、力学的に強靭な付着部を形 成している(C, D)。
39
表1. 組織学的スコアリング
40
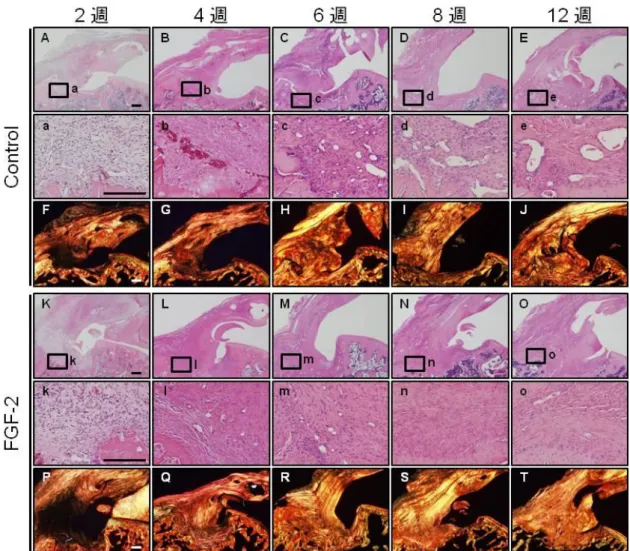
図3. 組織学的評価
腱骨修復部の組織学的な所見をHE染色(A-E, K-O)と偏光顕微下picrosirius-red(F-J, P-T)染色に より評価した(各群、各タイムポイントn = 6)。a-e, k-oはそれぞれA-E, K-O内の拡大像である。
術後2週(A,K)では両群共に腱骨間に血管と細胞が豊富で、不規則配列したコラーゲン線維から なる肉芽組織が形成され(F, P)、経過に伴い細胞密度や血管数の減少が観察されたが、術後12週 の対照群の腱骨間には血管に富む不規則に配列したコラーゲン線維からなる線維性組織が介在し ていた(E, J)。一方、FGF-2群では対照群と比較してこれらの修復組織は明らかに減少し、一定方 向に配列したコラーゲン線維からなる腱様の修復組織により骨と連続していた(O, T)(Scale bars = 100 µm)。
41
図4. 組織学的評価結果
修復組織の組織学的成熟度は細胞密度、血管数、コラーゲン線維配列によるスコアを用 いて評価した(各群、各タイムポイントn = 6)。項目別にみると、術後6週の細胞密度お よび 8 週、12 週の血管数が FGF-2 群において対照群と比較して有意に少なく、術後 4 週以降のコラーゲン線維配列がFGF-2群において対照群と比較して有意に高値であった。
3項目による組織学的スコアは術後4週以降のFGF-2群において対照群と比較して有意 に高値であった(*P < 0.05, Mann-Whitney U test)。
42
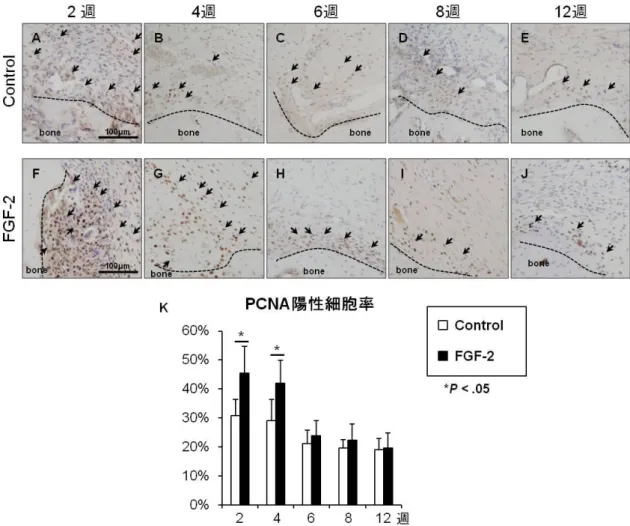
図5. 修復部における増殖期細胞の評価
修復部における増殖期細胞をproliferating cell nuclear antigen(PCNA)の免疫染色により評価した
(各群、各タイムポイントn = 6)。修復後早期においてはFGF-2群(F, G)において対照群(A, B)
より多くのPCNA陽性細胞が認められた。術後6週以降では、両群ともにPCNA陽性細胞は明ら かに減少し、主に上腕骨と修復部の境界の領域に観察された(C-E, H-J)、(矢印、PCNA陽性細胞)、
(Scale bars = 100 µm)。修復部におけるPCNA陽性細胞率は術後2週、4週においてFGF-2群で対
照群と比較して有意に高値であった。術後6、8、12週では両群間に有意な差はみられなかった(K)、
(*P < 0.05, Mann-Whitney U test)。