An improved
assay
method
for fibroblast
gelatinolytic
enzyme
Kichibee Otsuka, Mitsuhiro Ohshima, Masamichi Kaku,
Takaki Kajima, Masato Fukuoka, Yukitoshi Kaiya and Kantaro Suzuki
Department of Biochemistry, Nihon University School of Dentistry
(Received 14 June and accepted 7 November 1997)
Abstract: A useful gelatinolytic enzyme assay for fibroblasts, utilizing a novel sample preparation method for collagenase with dithiothreitol (DTT) treatment to inactivate endogenous collagenase inhibitors, was developed using soluble fluorescein isothiocyanate (FITC)-labeled gelatin. The substrate, gelatin was prepared by heating commercially
available FITC-labeled type I collagen. The
denatured collagen was cleaved with purified trypsin and partially purified fibroblast gelatinase, and the
digested FITC-fragments were measured
fluorometrically. The intensity of the fluorescence was in proportion to the reaction time and enzyme
concentration. Both enzyme activities were
measurable within the nanogram range of enzyme preparations. The enzyme activity was detected after
4-aminophenylmercuric acetate (APMA) treatment
which was completely inhibited by metalloproteinase inhibitors, but not by serine- and cysteine-proteinases' inhibitors. Conditioned media of human periodontal ligament fibroblasts (PLF) and gingival fibroblasts (GF) were separately treated with DTT prior to the enzyme assay, and then the assay was performed in the presence of APMA. The enzyme activities of PLF and GF were 106- and 55-fold higher than those of the conventional gelatinase assay which was carried out without DTT treatment. This assay method allowed the measurement of gelatinolytic enzyme activity when tissue inhibitors of metalloproteinases were present in the fibroblast culture medium.
Key words: fibroblast gelatinolytic enzyme; FITC-labeled gelatin; TIMP inactivation.
Introduction
Matrix metalloproteinases (MMPs) play an important role in tissue remodeling, degradation, and wound healing, since they break down collagens and other proteins in the extracellular matrix of the periodontium
(1, 2). Since many tissues produce both MMPs and tissue inhibitors of metalloproteinases (TIMPs), sensitive enzyme assay methods for MMPs that avoid the effects of TIMPs are necessary for accurate determination of MMP
activities. As substrates for MMPs, fluorescent peptides are available (3) that allow for the examination of MMP activities with high sensitivity. However, protein substrates are preferred since they more closely resemble true physiological substrates. Radiolabeled substrates for MMPs have also been used in some studies (4-6), but these are expensive and hazardous. On the other hand, most fluorescent protein derivatives for conventional
proteinase assays (7, 8) do not have as highly sensitive as radiolabeled substrates (9). As for other assay methods,
gelatin-substrate enzymography has also been used to assay gelatinase and stromelysin (10-13) with relatively high sensitivity, because TIMP was separated by the
electrophoresis. However, this would be laborious and it would be difficult to determine the enzyme activity
precisely.
Recently, as fluorescein isothiocyanate (FITC)-labeled type I collagen has become more widely available, we have used it to develop a new collagenase assay (14, 15)
that is 3-10 times more sensitive than the conventional collagenase assays. Furthermore, it is useful to determine the functional activity in the presence of two different types of collagenase inhibitors which are considered to be
identical with TIMPs, using the culture medium of human
periodontal ligament fibroblasts and gingival fibroblasts as samples. Utilizing the above procedures, we attempted to establish a fibroblast gelatinolytic enzyme assay method employing heat-denatured FITC-labeled type I collagen as a substrate.
Materials and Methods Chemicals
The chemicals used and their sources were as follows: FITC-labeled bovine type I collagen from Elastin Products; 4-aminophenylmercuric acetate (APMA),
gelatin-agarose gel, bovine serum albumin (BSA), penicillin G, gentamicin sulfate, N-ethylmaleimide
(NEM) and phenylmethylsulfonyl fluoride (PMSF) from Sigma; Ultrogel AcA 54 from IBF Biotechnics; dithiothreitol (DTT) from Boehringer Mannheim; a-minimum essential medium (ƒ¿-MEM) and trypsin from Gibco; fetal bovine serum (FBS) from Sebak; amphotericin B from Flow Laboratories; dimethylsulfoxide (DMS0) and ethylenediamine-tetraacetic acid (EDTA) from Dojindo Laboratories;
prestained MW markers from Bio-Rad Laboratories; and sodium dodecyl sulfate (SDS) and o-phenanthroline from Wako. All other reagents were of analytical grade. Correspondence to Dr. Kichibee Otsuka, Department of Biochemistry,
Nihon University School of Dentistry, 1-8-13 Kanda-Surugadai, Chiyoda-ku, Tokyo 101, Japan
Cell Culture
Human gingival fibroblasts (GF) were obtained from
gingival connective tissue explants as described by Ohshima et al. (16, 17). At the 5th passage of GF culture, the cells were seeded at a density of 106 cells per T-75 (75 cm2) flask (Corning), and cultured for two weeks at 37•Ž with a-MEM supplemented with 10 % heat-inactivated FBS and antibiotics (100 ƒÊg/ml penicillin G, 50 ƒÊg/ml
gentamicin sulfate, 0.3 ƒÊg/ml amphotericin B) under a humidified atmosphere with 5 % CO2/95 % air. Then, the culture medium was replaced with serum-free a-MEM containing the above antibiotics, and the cells were further cultured for 5 days; half of the culture medium was collected, and fresh medium was added daily.
Preparation of partially purified gelatinase
Thirty milliliters of the GF-cultured medium was
pooled and concentrated by ammonium sulfate precipitation at 10-90 % saturation. The precipitate was dissolved with a one-tenth volume of gelatinase assay buffer, and dialyzed against the same buffer. The buffer was composed of 0.2 M NaC1, 5 mM CaC12, 0.05 % Brij 35 and 0.02 % NaN3 in 50 mM Tris-HC1 buffer, pH 7.4 (18,19).
The dialyzed solution was then applied to Ultrogel AcA 54 column (1.6 •~ 90 cm) equilibrated with the above buffer, and eluted at a flow rate of 10 ml/h. The eluate was fractionated into 2.0-m1 aliquots, in which
gelatinolytic activity was determined by gelatinase enzymography. The fractions containing the enzyme activity was pooled for further purification.
The pooled eluate was subjected to gelatin-agarose gel affinity column chromatography as described by Hibbs et al. (20) and Hipps et al. (21), and eluted with 5 % DMSO. The enzyme activity in the eluted fractions was examined by enzymography as described above. The fractions containing high gelatinase activity were pooled and used as partially purified gelatinase.
Enzymography
Gelatin-substrate enzymography was performed as described by Overall and Limeback (11), and Gangbar et al. (12). Briefly, a discontinuous 10 % cross-linked
SDS-polyacrylamide mini-slab gel containing 40 ƒÊg/ml gelatin was used. Twelve microliters of sample and 4.0 ƒÊl of electrophoresis sample buffer (2 % SDS, without urea and reducer) were mixed, and electrophoresed at 150 V for 1 h. The slab-gel was rinsed twice with 2.5 % Triton X-100 solution, and then incubated at 37•Ž for 2 h in the enzyme assay buffer. The gel was fixed for 5 min with
10 % acetic acid, and stained with Coomassie brilliant blue (CBB) G 250. Clear bands indicating gelatin digestion by the enzyme were observed on the slab gel against the blue background stained by CBB G 250.
FITC-gelatin preparation
FITC-labeled type I collagen was dissolved in 10 mM acetic acid to give 0.2 % collagen, and treated by heating at 60•Ž for 30 min as described by Sunada and Nagai (4),
Nakano and Scott (5), Hipps et al. (21), and Harris and Krane (22). Prior to use, FITC-labeled gelatin solution was diluted with an equal volume of the enzyme assay buffer (pH adjusted to 8.5), and 0.1 % gelatin solution was used as a substrate for the following gelatinolytic enzyme assay.
Assay conditions for gelatinolytic enzyme activity
The reaction mixture consisted of up to 25 ƒÊl of enzyme sample, 50 ƒÊl of the assay buffer including 0.1 % BSA and 2 mM APMA, and 25 ƒÊl of 0.1 % FITC-labeled
gelatin. The reaction was performed using covered 1.5-ml polypropylene micro-test tubes. For the trypsin assay, the above reaction mixture was utilized without BSA and APMA. After shaking the reaction mixtures in the tubes thoroughly, the mixture was incubated at 37•Ž for 60 min. The reaction was terminated by adding 200 of trichloroacetic acid (TCA) and tannic acid (TA) mixed solution (TCA/TA solution) and then shaking vigorously. The tubes were allowed to stand at 4•Ž for 10 min. The TCA/TA-insoluble proteins were precipitated by centrifugation at 4,000 rpm for 10 min with a centrifuge
(Tomy MC-15A).
A 100111 aliquot of the above supernatant was diluted with 250 ƒÊl•@ of 0.5 M Tris-HC1 buffer to ensure that the entire sample was at the proper pH for fluorometrical assay of FITC-gelatin fragments digested. Three hundred
was taken for measuring the intensity by a microplate reader (Corona MTP-32, Corona Electric, Japan) combined with a fluorescence attachment (Corona MTP-F2) at an excitation wavelength of 490 nm and an emission wavelength of 530 nm.
One-hundred percent relative fluorescence was considered to equal the total fluorescence intensity derived from FITC-gelatin fragments hydrolyzed by an excess amount of trypsin (125 ƒÊg/tube). Plotted points were the averages of duplicate determinations. Individual values were used to calculate the coefficient of correlation.
Furthermore, molecular weights of FITC-gelatin fragments degraded by the enzyme were examined by SDS-polyacrylamide gel electrophoresis (SDS-PAGE). After centrifugation of the enzyme reaction mixture, the supernatant was dialyzed against distilled water, and then lyophilized. The lyophilized supernatant was mixed with electrophoresis sample buffer (2 % SDS, 2 % urea, 2 % mercaptoethanol), and electrophoresed on a 7.5 % SDS-PAGE mini slab gel, which was stained with CBB R 250 according to the method of Hayashi and Nagai (23).
To determine the optimum pH for the assay conditions, enzymography was carried out in assay buffer at various
pH values according to Piening and Rieder-Henderson (24). To clarify enzyme types, metallo-, serine or cysteine proteinase, of this reaction, inhibitory effects of 10 mM EDTA (19, 25-27), 10 mM o-phenanthroline (26, 27), 1 mM PMSF (28), and 5 mM NEM (29) were examined by enzymography, respectively.
Fibroblast gelatinolytic enzyme assay in crude solution
Four pairs (cases) of human specimens of periodontal ligament fibroblasts (PLF) and gingival fibroblasts (GF) were seeded separately into 24-well plates (Corning) at 2
×104 cells/well, and incubated for two weeks in culture medium as described previously (15, 17). The cells were then cultured for an additional 72 h with serum-free ƒ¿-MEM containing 0.1 % BSA and antibiotics, and the fibroblast-conditioned media were harvested. Prior to the enzyme assay, each conditioned medium was treated with a final concentration of 10 mM DTT for 30 min at 35•Ž to inactivate any endogenous metalloproteinase inhibitor, and dialyzed exhaustively against assay buffer at 4•Ž for
18 h to remove DTT (14, 15). Samples for the conventional gelatinolytic enzyme assay (1, 30) were dialyzed directly against the assay buffer without DTT treatment. Two to five IA of the DTT-treated media was used for the enzyme assay, whereas 25 ƒÊl of the untreated media was used for the conventional assay. The experiment was performed in triplicate for each case, and the data were expressed as means and standard deviations for triplicate wells or four cases in terms of mU/ƒÊg DNA
(15, 17).
To examine the effects of this sample preparation method on gelatinolytic enzyme activity, APMA-treated active gelatinase and progelatinase from GF were treated with DTT and dialyzed against assay buffer. Active enzyme was abolished almost completely with DTT treatment, and the activity was not recovered after treatment with APMA. The proenzyme was not affected by DTT treatment, and the activity was detected only after APMA activation.
Results
Partial purification of fibroblast gelatinase
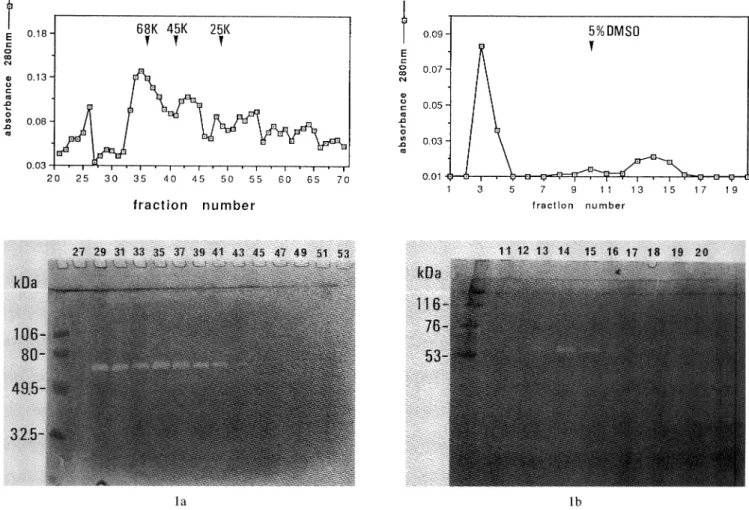
The soluble proteins in GF-cultured medium were concentrated with ammonium sulfate and dialyzed against the gelatinase assay buffer. The concentrated proteins were then applied to Ultrogel AcA 54 column, and gelatinolytic enzyme activity in each eluted fraction was determined by gelatin enzymography. The protein concentration in each fraction was also measured with a
spectrophotometer (Shimadzu CL-1200) at 280 nm. The elution profile of the proteins and the enzymography are shown in Fig. la. Clear bands on the enzymogram showed gelatinolytic enzyme activity in eluate fraction numbers 29 to 39. The molecular weight of the enzyme
1a 1b
Fig. 1 Enzymography on Ultrogel AcA 54 (a) and a gelatin-agarose affinity gel (b)-eluted sample. Concentrated gingival fibroblast
conditioned medium was applied to an Ultrogel AcA 54 gel-filtration column (a) followed by gelatin-agarose affinity gel (b)
and monitored for gelatin-degrading activity by enzymography and absorbance (added graphs), as detailed in Materials and
Methods. Added graphs for Ultrogel AcA 54 (a) - ; absorbance 280 nm, for gelatin-affinity column (b) - ; absorbance
was estimated to be approximately 72 kDa from the enzymogram. The above fractions were collected and pooled, applied to a gelatin-agarose affinity column, and the enzyme activity in each fraction was examined by enzymography. Clear bands were observed at fraction numbers 13 to 15 (Fig. lb), which eluted with 5 % DMSO. These fractions were used as partially purified gelatinase.
Solubility of undigested or digested
FITC-gelatin at various concentrations of TCA/TA
solution
Prior to determining gelatinolytic activity, the solubility of undigested or digested FITC-gelatin fragments were examined with various concentrations of TCA/TA solution. Undigested or trypsin digested FITC-gelatin were precipitated by the combination of 5 % or 10
% TCA mixed with several concentrations of TA. The lowest fluorescence from supernatant fractions of undigested FITC-gelatin (blank) were obtained when the combination of 5 % TCA/0.05 % TA, 10 % TCA/0.05 % TA, and 10 % TCA/0.01 % TA solutions were used. In terms of the fluorometrical intensity of the
TCA/TA-soluble FITC-fragments entirely digested by 125 lag trypsin, the highest fluorescence was obtained by the
combination of 10 % TCA/0.01 % TA among the above TCA/TA solutions (Fig. 2). Therefore, we used 10 % TCA/0.01 % TA solution to terminate the enzyme reaction, and to determine the soluble FITC-fragments.
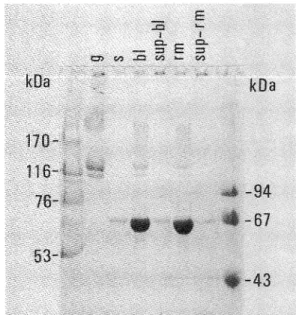
To determine molecular weights of proteins precipitated under this assay condition, proteins in the enzyme assay
mixture was separated by SDS-PAGE. Proteins of
molecular weights below 67,000 including the enzyme digested-gelatin fragments were not precipitated by this condition (Fig. 3).
The optimal pH for measuring intensity of the fluorescence derived from soluble FITC-fragments was examined next, and a pH of 8.5 was found to be the most effective pH between 7.5 and 9.0 (Fig. 4).
Fig. 2 Solubility of undigested or digested FITC-gelatin at various concentrations of TCA/TA mixed solutions. Undigested gelatin (blank) or 125 mg trypsin digested gelatin fragments (total) were precipitated by combinations of TCA/TA solution. -• - ; 10 % TCA for blank, ---•›-- ; 5 % TCA for blank, ---•ž-- ; 10 % TCA for total, ---•¢--; 5 % TCA for total.
Fig. 3 Determination of TCA/TA soluble proteins. Molecular weights of proteins in the enzyme reaction mixture were examined by SDS-PAGE, followed by CBB R 250 staining. g; gelatin substrate, s; partially purified gelatinase, bl; reaction mixture without enzyme before centrifugation, sup-bl; supernatant of bl after centrifugation, rm; reaction mixture before centrifugation, sup-rm; supernatant of rm after centrifugation.
Fig. 4 Effects of pH on fluorescence of FITC-fragments. After centrifugation, fluorescence of supernatant from undigested gelatin (blank) or digested gelatin fragments (total) was examined with 0.5 M Tris-HC1 buffer, pHs between 7.5 and 9.0 with pH intervals at 0.5. •\•\ ; blank,--- ; total.
Gelatinase assay conditions
Since gelatinolytic activity in the culture medium was in an inactive form (latent form or proenzyme), the
enzyme was activated by APMA-treatment. The
relationship between preincubation time and the partially purified gelatinase (65 ng) activity is shown in Fig. 5.
Since the highest enzyme activity was observed at time
zero, substrate and APMA were added at the same time
just before the start of the enzyme reaction. In addition,
since the optimal pH range for this enzyme assay was
between pH 6.9 and 8.4, we decided to use the buffer at
pH 7.4 for the following assay (Fig. 6).
8a
8b Fig. 5 Effect of preincubation time with APMA on partially
purified gelatinase. Sixty-five nanograms of partially purified gelatinase was activated with 2 mM APMA at
37•Ž for various times and then incubated with substrate at 37•Ž for 60 min.
Fig. 6 Optimum pH of partially purified gelatinase. Gelatin substrate gel strips containing enzyme were incubated in the assay buffer at the indicated pH at 37•Ž for 2 h, then stained with CBB G 250.
Fig. 7 Time dependence of the hydrolysis of FITC-gelatin by partially purified gelatinase. Twenty-four nanograms of partially purified gelatinase was incubated at 37•Ž in assay buffer containing 0.1 % BSA and 2 mM APMA. For partially purified gelatinase, r = 0.994.
Fig. 8 Hydrolysis of FITC-gelatin by partially purified gelatinase (a) or trypsin (b). Partially purified gelatinase (a) was incubated in assay buffer containing 0.1 % BSA and 2 mM APMA. Trypsin (b) was incubated in the assay buffer without BSA and APMA. Both enzymes were incubated at 37•Ž for 60 min. For partially purified gelatinase (a), r = 0.999; for trypsin (b), r = 0.998.
The time dependence of the hydrolysis of FITC-gelatin by 24 ng of partially purified gelatinase is shown in Fig. 7. Because the intensity of the fluorescence from the degraded substrate was proportional for up to 90 min of the enzyme reaction, 60 min was employed for the assay.
The cleavage of FITC-gelatin by partially purified
gelatinase and trypsin was proportional to the concentrations of both enzymes (Fig. 8a, b). The most useful concentrations of partially purified gelatinase were 1 to 15 ƒÊl (4.8 to 72 ng protein), and those for trypsin were 2 to 30 ng for the 60 min incubation period. The fluorescence derived from hydrolyzed FITC-gelatin was
proportional to 40 % of the degradation of the total amount of FITC-gelatin used (Fig. 8a).
The inhibitory effects of EDTA, o-phenanthroline, PMSF and NEM are shown in Fig. 9. The enzyme activity was entirely blocked by both EDTA and
o-phenanthroline, which are known as pharmacological metalloproteinase inhibitors. The enzyme activity was not inhibited by PMSF or NEM.
Fibroblast gelatinolytic enzyme assay in crude solution
The gelatin-degrading activities of the sample
preparations of both conditioned media from PLF and GF are shown in Table 1. The cleavage of FITC-gelatin according to these crude samples were proportional to the enzyme concentration and reaction time up to 90 min. According to the conventional sample preparation, the
gelatinolytic activity of PLF was 16 mU, and that of GF was 33 mU per lig DNA on average. By employing our new sample preparation, on the other hand, gelatinolytic
activity was markedly increased. Using this procedure, the gelatinolytic enzyme activity of PLF was 1707 mU
and that of GF was 1919 mU per lig DNA. These data revealed that our method was 106-fold higher for PLF and 55-fold higher for GF than the conventional assay methods.
Discussion
FITC-gelatin is suitable for determination of
gelatinolytic enzyme activity because it is similar to physiological substrates. FITC-gelatin is easy to prepare by heating FITC-labeled type I collagen at 60•Ž, it is stable during storage, and inexpensive. We used it for carrying out the gelatinolytic enzyme assay, and were able to obtain sufficient sensitivity equivalent to that of
previous radioactive assays (5-100 ng trypsin) (31) without any problems related to radioactive waste or other health hazards.
Fig. 9 Inhibitory effects of some reagents. Gelatin substrate gel strips containing enzyme were incubated in the assay buffer containing indicated inhibitors at 37•Ž for 2 h, then stained with CBB G 250. C; control, E; EDTA, O; o-phenanthroline, P; PMSF, N; NEM.
Table 1 Gelatinolytic enzyme activity of PLF and GF conditioned media
Four pairs (cases) of PLF and GF confluent cultures were further incubated with 0.1 % BSA containing ƒ¿-MEM for 72 h.Harvested conditioned media were directly dialyzed against assay buffer (conventional preparation) or treated with 10 mM DTT at 35•Ž for 30 min and dialyzed against assay buffer (new preparation), and tested for gelatinolytic enzyme activity in the presence of 1 mM APMA at 37•Ž for 60 min. The results represent the means and standard deviations of triplicate wells (for each case) or four cases (mean).
To handle gelatin with the TCA and TA mixture, the solution was shaken thoroughly to precipitate the undigested gelatin for this assay. Although Twining (9) reported that 5 % TCA was useful for precipitating undigested casein, we found that the solution should be a mixture of 10 % TCA and 0.01 % TA to precipitate undigested gelatin, and to obtain a low blank (Fig. 2).
In order to determine the fluorescence derived from digested soluble FITC-gelatin fragments, the above solution should be neutralized with 0.5 M Tris-HC1 solution, pH 8.5. Sufficient FITC fluorescence will not be produced under lower pH conditions (9). We found that the fluorescence was also decreased at higher pH values (Fig. 3).
Gelatin digestion in this assay was inhibited by EDTA (19, 25-27) and o-phenanthroline (26, 27), but not by PMSF (28) and NEM (29), indicating this enzyme activity was of metalloproteinases.
Fibroblast gelatinase is well known as metallopro-teinase 2 (MMP-2), and organomercurials such as APMA are needed in order to activate the enzyme (6, 21). APMA was necessary for activating the enzyme, but the enzyme activity was markedly decreased by incubation with APMA without FITC-gelatin (Fig. 5). Although BSA was added to the above preincubation mixture, it was not effective in protecting the active enzyme against APMA treatment. The above observations indicated that the substrate would be necessary in order to maintain the enzyme activity after APMA activation. A similar observation was reported by Fridman et al. (6), although the reason why APMA both activated and inhibited the enzyme was unknown. They also found that the optimal activation time with APMA treatment was 15 min, but that further incubation markedly reduced the gelatinase activity. Our results, however, indicated that the optimal activation time was zero. We presume this discrepancy was due to the enzyme existing as a complex with TIMP-2, as described by Goldberg et al. (32) and Stetler-Stevenson et al. (33), whereas Fridman et al. (6) employed human recombinant 72-kDa gelatinase for determining the proper APMA treatment time.
Meikle et al. (2) have stated that fibroblasts synthesize and secrete MMPs in latent form (proenzymes) and TIMP. Once MMPs were activated, they would be trapped if sufficient amounts of TIMP were secreted by the same fibroblasts, and thus inactivated. For instance, Heath et al. (1) detected MMPs in tissue culture medium of gingiva, but failed to detect MMPs including gelatinase in the culture medium of gingival fibroblasts, although they did detect a potent amount of inhibitor. Pettigrew et al. (30), and Richards and Rutherford (31) failed to detect
collagenase activity in PLF culture media while they
detected the immunoreactive procollagenase and
procollagenase mRNA (35). To solve this problem, we developed a new sample preparation procedure (14, 15) for detecting collagenase activity in human PLF, and found that the activity was 3- to 10-fold higher compared with the conventional methods (1, 30, 35). This was because collagenase inhibitors coexisting in the medium were inactivated by DTT treatment without any effect on
procollagenase (15). If the active form of fibroblast gelatinase was also inhibited by TIMP as described by Hipps et al. (21), we hypothesized that DTT treatment would be similarly effective for detecting gelatinolytic enzyme activity in the culture medium of fibroblasts. The present study revealed the clear effectiveness of DTT
treatment followed by APMA in detecting the
gelatinolytic activity of PLF and GF culture media (Table I), which would contain two different types of metalloproteinase inhibitors (TIMPs) as described by Ohshima et al. (14). This study also revealed that secretion of gelatinolytic enzyme by PLF and GF occurred without any stimulation in vitro, whereas Nakano and Scott (5) used concanavalin A (ConA), and Hipps et al. (21) used phorbol myristate acetate (PMA) for stimulation of GF to produce gelatinase.
As DTT treatment of the medium caused inactivation of the active gelatinolytic enzyme, the present sample preparation procedure would not be suitable when active enzymes exist in the sample. Since little active form of this enzyme is present in culture media, the DTT treatment is unlikely to affect the analysis. Therefore, the present sample preparation procedure would be useful for detecting fibroblast gelatinolytic activity without any purification steps when the enzyme is present in proenzyme form.
Acknowledgment
This study was supported in part by Research Grant for Assistants of Nihon University Research Grant for 1996.
References
1. Heath, J. K., Gowen, M, Meikle, M. C. and Reynolds, J. J. (1982) Human gingival tissues in culture
synthesize three metalloproteinases and a
metalloproteinase inhibitor. J. Periodontal Res., 17, 183-190
2. Meikle, M. C., Heath, J. K. and Reynolds, J. J. (1986) Advances in understanding cell interactions in tissue resorption. Relevance to the pathogenesis of periodontal disease and a new hypothesis. J. Oral Pathol., 15, 239-250
3. Netzel-Arnett, S., Mallya, S. K., Nagase, H., Birkedal-Hansen, H. and Van Wart, H. E. (1991) Continuously recording fluorescent optimized for five human matrix metalloproteinases. Anal. Biochem.,
195, 86-92
4. Sunada, H. and Nagai, Y. (1980) A rapid micro-assay method for gelatinolytic activity using tritium-labeled heat-denatured polymeric collagen as a substrate and its application to the detection of enzymes involeved in collagen metabolism. J. Biochem., 87, 1765-1771 5. Nakano, T. and Scott, P. G. (1986) Purification and
characterization of a gelatinase produced by fibroblasts from human gingiva. Biochem. Cell Biol. 64, 387-393
6. Fridman, R., Bird, R. E., Hoyhtya, M., Oelkuct, M., Komarek, D., Liang, C. -M., Berman, M. L., Liotta, L. A., Stetler-Stevenson, W. G. and Fuerst, T. R. (1993) Expression of human recombinant 72 kDa gelatinase
and tissue inhibitor of metalloproteinase-2 (TIMP-2): Characterization of complex and free enzyme. Biochem. J., 289, 411-416
7. Wunsch, E. and Heidrich, H. G. (1963) Zur
quantitativen Bestimmung der Kollagenase. Hoppe-Seyler' s Z. Physiol. Chem., 333, 149-151
8. Spencer, P. W., Titus, J. S. and Spencer, R. D. (1975) Direct fluorimetric assay for proteolytic activity against intact proteins. Anal. Biochem., 64, 556-566 9. Twining, S. S. (1984) Fluorescein
isothiocyanate-labeled casein assay for proteolytic enzymes. Anal. Biochem., 143, 30-34
10. Heussen, C. and Dowdle, E. B. (1980) Electrophoretic analysis of plasminogen activators in polyacrylamide
gels containing sodium dodecyl sulfate and copolymerized substrates. Anal. Biochem., 102, 196-202
11. Overall, C. M. and Limeback, H. (1988) Identification and characterization of enamel proteinases isolated from developing enamel. Amelogeninolytic serine
proteinases are associated with enamel maturation in pig. Biochem. J., 256, 965-972
12. Gangbar, S., Oveall, C. M., McCulloch, C. A. G. and Sodek, J. (1990) Identification of polymorphonuclear leukocyte collagenase and gelatinase activities in mouthrinse samples: correlation with periodontal disease activity in adult and juvenile periodontitis. J. Periodontal Res., 25, 257-267
13. Sodek, J. and Overall, C. M. (1992) Matrix
metallo-proteinase in periodontal tissue remodeling. Matrix Suppl., 1, 352-362
14. Ohshima, M., Kuwata, F., Sasai, Y., Otsuka, K. and Suzuki, K. (1993) Collagenase and collagenase inhibitors secreted by cultured human periodontal ligament cells. J. Nihon Univ. Sch. Dent., 35, 109-117 15. Ohshima, M., Otsuka, K. and Suzuki, K. (1994)
Interleukin-1p stimulates collagenase production by cultured human periodontal ligament fibroblasts. J. Periodontal Res., 29, 421-429
16. Ohshima, M., Kuwata, F., Otsuka, K., Saito, R., Sato, K., Shioji, S. and Suzuki, K. (1988) Alkaline
phosphatase activities of human periodontal ligament cells. J. Nihon Univ. Sch. Dent., 31, 208-217
17. Ohshima, M., Otsuka, K., Kuwata, F., Fujikawa, K., Ito, K. and Suzuki, K. (1993) Gingival epithelial cells secrete a substance which increases the collagenolytic enzyme activity of periodontal ligament cells. J. Nihon Univ. Sch. Dent., 35, 57-64
18. Overall, C. M., Wiebkin, O. W. and Thonard, J. C.
(1987) Demonstration of tissue collagenase activity in vivo and its relationship to inflammation severity in human gingiva. J. Periodontal Res., 22, 81-88
19. Overall, C. M., Wrana, J. L. and Sodek, J. (1989) Transforming growth factor-ƒÀ regulation of collagenase, 72 kDa-progelatinase, TIMP and PAI-1 expression in rat bone cell populations and human fibroblasts. Connective Tissue Res., 20, 289-294 20. Hibbs, M. S., Hasty, K. A., Seyler, J. M., Kang, A. H.
and Mainardi, C. L. (1985) Biochemical and
immunological characterization of the secreted forms of human neutrophil gelatinase. J. Biol. Chem., 260, 2493-2500
21. Hipps, D. S., Hembry, R. M., Docherty, A. J. P., Reynolds, J. J. and Murphy, G. (1991) Purification and characterization of human 72-kDa gelatinase (Type IV collagenase); Use of immunolocalisation to demonstrate the non-coordinate regulation of the 72- kDa and 95-kDa gelatinase by human fibroblasts. Biol. Chem. Hoppe Seyler, 372, 287-296
22. Harris, E. D. and Krane, S. M. (1972) An endopeptidase from rheumatoid synovial tissue culture. Biochim. Biophys. Acta, 258, 556-576 23. Hayashi, T. and Nagai, Y. (1979) Separation of a
chains of type I and III collagens by SDS-polyacrylamide gel electrophoresis. J. Biochem., 86, 453-459
24. Piening, C. and Riederer-Henderson, M. A. (1989) Neutral metalloproteinase from tendons. J. Orthop. Res., 7, 228-234
25. Sellers, A. and Reynolds, J. J. (1977) Identification and partial characterization of an inhibitor of collagenase from rabbit bone. Biochem. J., 167, 353-360
26. Pettigrew, D. W., Ho, G. H., Sodek, J., Brunette, D. M. and Wang, H. M. (1978) Effect of oxygen tension and indomethacin on production of collagenase and neutral proteinase enzymes and their latent forms by porcine gingival explants in culture. Arch. Oral Biol., 23, 767-777
27. Vaes, G., Eeckhout, Y., Lenaers-Claeys, G., Francois-Gillet, C. and Dreutz, J. E. (1978) The simultaneous release by bone explants in culture and parallel activation of procollagenase and of a latent neutral proteinase that degrades cartilage proteoglycans and
denatured collagen. Biochem. J., 172, 261-274 28. Moss, D. E. and Fahrney, D. (1978) Kinetic analysis
of differences in brain acetylcholineesterase from fish or mammalian sources. Biochem. Pharmac., 27, 2693-2698
29. Brubacher, L. J. and Glick, B. R. (1974) Inhibition of papain by N-ethylmaleimide. Biochemistry, 13, 915-920
30. Pettigrew, D. W., Sodek, J., Wang, H. -M. and Brunette, D. M. (1980) Inhibitors of collagenolytic enzymes synthesized by fibroblasts and epithelial cells from porcine and macaque periodontal tissues. Arch. Oral Biol., 25, 269-274
31. Sevier, E. D. (1976) Sensitive, solid-phase assay of proteolytic activity. Anal. Biochem., 74, 592-596 32. Goldberg, G. I., Marmer, B. L., Grant, G. A., Eisen,
A. Z., Wilhelm, S. and He, C. (1989) Human 72- kilodalton type IV collagenase forms a complex with a tissue inhibitor of metalloproteinase designated TIMP-2. Proc. Natl. Acad. Sci. USA, 86, 8207-8211 33. Stetler-Stevenson, W. G., Krutzsch, H. C. and Liotta,
L. A. (1989) Tissue inhibitor of metalloproteinase (TIMP-2). A new member of the metalloproteinase inhibitor family. J. Biol. Chem., 264, 17374-17378
34. Richards, D. and Rutherford, R. B. (1988) The effects of Interleukin 1 on collagenolytic activity and prostaglandin-E secretion by human periodontal-ligament and gingival fibroblast. Arch. Oral Biol., 33, 237-243
35. Richards, D. and Rutherford, R. B. (1990) Interleukin-1 regulation of procollagenase mRNA and protein in periodontal fibroblasts in vitro. J. Periodontal Res., 25, 222-229