題目
高 Ca クリーゼを呈した PTH 産生後腹膜腫瘍 (MFH)の分子学的解析
―異所性にホルモン産生能を獲得する機序―
内 田 香 介
(内分泌・代謝病学専攻)
防衛医科大学校
平成26年度
目 次
第1章 緒言 1頁
第2章 材料と方法 4頁 第1節 症例報告 4頁 第2節 腫瘍と細胞株 5頁 第3節 免疫組織化学 5頁 第4節 mRNA解析 6頁 第5節 PTH遺伝子のcDNA解析とゲノムDNA解析 6頁 第6節 PTH遺伝子プロモーター領域CpG部位のメチル化解析 7頁 第7節 マイクロアレイによる網羅的遺伝子発現解析 8頁
第3章 結果 10頁 第1節 腫瘍細胞でのPTHタンパク質の検出 10頁 第2節 腫瘍細胞でのPTHmRNAの過剰発現 10頁 第3節 PTH遺伝子の構造解析 11頁 第4節 PTH遺伝子プロモーター領域CpG部位のメチル化解析 11頁 第5節 網羅的遺伝子発現解析 12頁
第4章 考察 14頁
第5章 結論 17頁
謝辞 18頁
引用文献 19頁
図表 24頁
第1章 緒言
ホルモンは内分泌器官から産生され、微量にもかかわらず生体の恒常性維持 や成長・生殖にかかわる情報伝達に重要な役割を果たす物質である。ホルモン の産生・分泌は他のホルモン (上位ホルモン・下位ホルモン・他) によって絶妙 に調節されているが、その調節機構は依然として不明な点がある。
内分泌器官から発生する腫瘍の中にはホルモン産生能を持つもの (ホルモン 産生腫瘍) があり、ホルモンの種類によって特徴的な病像を呈するだけでなく、
生命予後も大きく損ねる。ホルモン産生腫瘍の病態形成には、ホルモン制御機 構に関わる情報伝達分子の質的・量的異常が関与していることが推測され、こ れまでに上位ホルモン受容体自体の異常や上位ホルモン受容体共役因子の異常 が報告されている [1-3]。
一方、内分泌器官以外の組織から発生した腫瘍で、本来産生しないホルモン を産生するものを異所性ホルモン産生腫瘍と称する。異所性に産生されるホル モンは、adrenocorticotropic hormone (ACTH) が代表的で、antidiuretic hormone (ADH) や growth hormone releasing hormone (GHRH) なども知られ、すべて単一 遺伝子産物の翻訳・プロセッシングに起因するペプチドホルモンで、複数の律 速酵素による合成経路が必要な甲状腺ホルモンやステロイドホルモンの異所性 産生はみられない。神経内分泌腫瘍 (NET; neuroendocrine cell tumor) によるホル モン産生は、元来組織に点在する活性アミンやペプチドホルモン産生能をもつ 神 経 内 分 泌 細 胞 (neuroendocrine cell) ま た は amine-precursor uptake and decarboxylation (APUD) 細胞に由来する腫瘍細胞からであり [4]、厳密な意味で は「異所性」ではなく「正所性」と称する方が正しい。腫瘍による異所性ホル モン産生の機序は幾つかの仮説が提唱されており、腫瘍細胞が腫瘍化の過程で
1
退行が生じホルモンを産生する脱分化説 [5]、あるいはホルモン産生能を有する 原始細胞 (progenitor cell) が腫瘍化により様々に分化し多彩なホルモンを産生 する異分化説などがあり [6]、これらの過程でジェネティック・エピジェネティ ックな変化を生じ本来発現が抑制されていたホルモンが産生されるようになる と考えられている [7]。たとえば、ジェネティックではエリスロポエチン産生腫 瘍における Von-Hippel-Lindau (VHL) 遺伝子の変異が [8-10]、エピジェネティッ クでは異所性 ACTH 産生腫瘍 (肺小細胞癌由来の培養細胞株 DMS-79、気管支 カルチノイド) における proopiomelanocortin (POMC) 遺伝子プロモーターの脱 メチル化が関与することが示唆されている [11, 12]。しかしながら、ホルモン産 生能を異所性に獲得する機序は未検討・未解明の部分が多い。
副甲状腺ホルモン (PTH; parathyroid hormone) は副甲状腺により産生され、
カルシウム (Ca) 代謝において中心な役割を果たすホルモンで、血中 Ca やビ タミン D がその発現に影響している。副甲状腺細胞由来の腺腫や癌は PTH の制御機構を逸脱して高 Ca 血症を来たすが、副甲状腺に異常が無くても高 Ca 血症をきたす悪性腫瘍体液性高 Ca 血症 (HHM; humoral hypercalcemia of malignancy) が多いことも 1941 年の Albright らの報告以来知られており [13]、異所性 PTH 産生腫瘍と考えられていた。1987 年に副甲状腺ホルモン関 連タンパク質 (PTHrP; PTH-related protein) が発見されて以降、HHM の原 因物質のほとんどが腫瘍により産生される PTHrP であり [14-16]、副甲状腺 に異常の無い真の異所性 PTH は極めて稀 [17-39] と判明している。
PTH 遺伝子は副甲状腺細胞特異的に発現することがよく知られているが、副 甲状腺の継代できる細胞株が樹立できておらず、また異所性 PTH 産生腫瘍が稀 であり、その組織特異性「正所性」および副甲状腺とは無関係な組織による「異 所性」産生の分子機構は不明な点が多い。異所性 PTH 産生腫瘍では、PTH 遺
2
伝子とその上流の再構成・増幅 [17] や PTH 遺伝子プロモーター領域にある CpG 部位の低メチル化 [18] の PTH 産生能獲得への関与が報告されているの みで、解析が少ない。今回、高 Ca クリーゼを呈した異所性 (後腹膜) PTH 産 生悪性線維性組織球腫 (MFH; malignant fibrous histiocytoma) 症例を報告すると ともに、その PTH 産生のメカニズムに迫る目的で、腫瘍の DNA および RNA の解析を行った。PTH 遺伝子の構造異常はなく、またプロモーター領域の CpG ジヌクレオチドの低メチル化も認めなかった。網羅的転写解析の結果から、PTH 遺伝子発現を高める転写因子 Sp3 と NF-Yβ の発現増加、PTH の mRNA を 安定化する因子の発現上昇を認めた。
3
第2章 材料と方法
第1節 症例報告
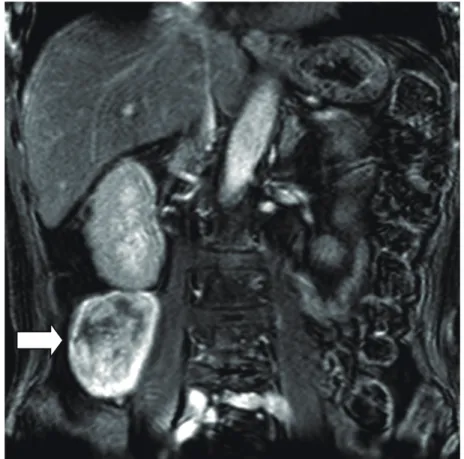
69 歳の男性患者が、多尿、口渇、倦怠感から高 Ca 血症と後腹膜腫瘍が発見 され当院へ入院した。入院時検査所見を表 1 に示す。補正血清 Ca 値は 18.2 mg/dL (基準範囲 8.4-10.4 mg/dL) と高値であった。血漿 PTHrP は 1 pmol/L 未 満 (基準範囲 1.0 pmol/L 未満) で正常であったのに対して、血清 intact PTH (iPTH) は 532 pg/mL (基準範囲 10-65 pg/mL) と高値だった。Magnetic resonance imaging (MRI) で、後腹膜腔に右腎下極に接して腫瘤がみられた (図 1 A)。血清
iPTH 上昇を伴う高 Ca 血症であり、原発性副甲状腺機能亢進症が示唆されたが、
超音波検査や 99mTc-MIBI シンチグラムで副甲状腺の腫大を認めなかった。入院 直後からの補液と zoledronic acid hydrate および elcatonin の投与により、補正
血清 Ca 値は一旦下降した。しかし、入院 16 日後より補正血清 Ca 値が薬物
療法中にもかかわらず再上昇し、後腹膜腫瘍は増大した。Computed tomography
(CT) や骨シンチグラフィーで後腹膜腔以外の腫瘍や転移は見られず、後腹膜腫
瘍が PTH を産生・分泌していることが示唆された。入院後 33 日目に、約 90X
80 X 50 mm 大の腫瘍が右腎および上行結腸とともに外科的に切除された。組織
病理学には、クロマチンの増量した異型核を持つ紡錘形細胞が、大型で強い異 型性を示す腫瘍細胞を散在性に混じながら、びまん性に一部花むしろ様配列を 構築して増殖していた。ビメンチン染色は陽性で、脂肪芽細胞は診断に有意と いえるほど認められず、平滑筋への分化は免疫組織化学で証明されなかったた め、悪性線維性組織球腫 (MFH) と診断された。術後間もなく血清 iPTH は 23 pg/mL へ下降し、補正血清 Ca 値も基準範囲内へ漸減した (図 1 B)。摘出され た腫瘍の一部を DMEM 培地に 2 時間静置し、上清の whole-PTH を測定した
4
ところ、4830 pg/mL (基準範囲 10-39 pg/mL) と高値であった。患者は入院後 55 日目に退院した。術後 6 年経過して、血清 iPTH や Ca は基準範囲内であった が後腹膜に再発を示唆する腫瘤を認めたため、精査加療中である。
第2節 腫瘍と細胞株
事前に患者に対して腫瘍の医学研究のための使用を説明し、文書による同意 を得た上で、本腫瘍は防衛医科大学校病院泌尿器科により切除された後、内分 泌代謝内科で液体窒素中に保存され、また病理部でホルマリン固定後にパラフ ィン包埋され保存された。副甲状腺腺腫は、帝京大学医学部附属病院から提供 された。この論文に記述される腫瘍の取扱い手順は、それぞれ関連する倫理委 員会にて承認された。SS-1 細胞株は、37 歳女性から単離されたヒト肺小細胞 癌由来の PTH だけでなく PTHrP や POMC を産生する新たな細胞株で、石川 真由美先生 (当時の所属は東邦大学糖尿病代謝内分泌センター) からご供与い ただいた [40]。MCF-7 細胞株は、69 歳女性から単離されたヒト乳癌由来の細 胞株で [41]、American Type Culture Collection (ATCC) から購入した。
第3節 免疫組織化学
本腫瘍のホルマリン固定パラフィン包埋切片を、増感を含めた西洋わさびペ ルオキシダーゼ (HRP) による酵素抗体法 (間接法) で免疫染色した。クエン酸 緩衝液 (pH 6.0) 中で熱処理 (95 ℃、40 分) し、抗原賦活した。3 % 過酸化水 素水中で 5 分間インキュベーションし、内因性ペルオキシダーゼを除去した。
一次抗体は、抗 PTH (1-34) マウスモノクローナル抗体 (AbD Serotec, UK) を使 用した。VECTASTAIN Elite ABC Kit (Vector Laboratories, Burlingame, CA) を使用 して標準方法で増感し、発色基質 3, 3’-diaminobenzidine (DAB) で発色した。
5
第4節 mRNA 解析
RNA を、RNA iso (Takara, Shiga, Japan) を使用して標準方法で抽出し、
PrimeScript RT reagent Kit (Takara) を使用 (ランダムヘキサマ―と Oligo dT プ ライマーの両方を利用) して逆転写 (RT; reverse transcription) 反応を 37 ℃ 25 分で行い、85 ℃ 15 秒で逆転写酵素を熱失活させた。標的遺伝子群およびリフ ァ レ ン ス 遺 伝 子 と し て 選 択 し た glyceraldehydes-3-phosphate dehydrogenase (GAPDH) それぞれのプライマーと SYBR Premix Ex Taq (Takara) を使用して相 対定量スタンダードカーブ法で定量的 RT-PCR (qRT-PCR) を施行した。PCR 反 応はシャトル PCR 標準プロトコールに基づき、初期変性 (95 ℃ で 30 秒) を 1 サイクル、続いて 2 ステップ PCR (95 ℃ で 5 秒、60 ℃ で 45 秒) を 40 サイクル行い、最後に融解曲線分析を追加してシングルピークであることを確 認した。プライマーの配列を表 2 に示す。プライマー配列設定には、ゲノム DNA 由来の増幅を避けるために、サイズの大きなイントロンの前後のエキソン に Forward と Reverse のプライマーをそれぞれ Primer3Plus を使用して設計 し、さらに National Center for Biotechnology Information (NCBI) の Basic Local Alignment Search Tool (BLAST) で検索してその特異性を確認した。副甲状腺腺腫 を陽性対照に、MCF-7 を陰性対照にした。検体の測定は最低 3 回行った。
第5節 PTH 遺伝子の cDNA 解析とゲノム DNA 解析
cDNA 解析。腫瘍から RNA を抽出し、PrimeScript RT reagent Kit (Takara) を
用いて PTH の cDNA を合成した。PTH 遺伝子の全コード領域を含む断片を増
幅するように設定したプライマー (表 2) と KOD FX PCR enzyme を使用して PCR 増幅した 697 塩基対の PCR 産物を pBluescript II SK (+) phagemid ベク ター (Stratagene, La Jolla, CA, USA) にクローン化し、Applied Biosystems 3730xl
6
DNA アナライザを使用して塩基配列を決定した。
ゲノム DNA 解析。本腫瘍と患者の末梢血白血球から、Wizard SV Genomic DNA Purification System (Promega, Wisconsin, USA) と QIAmp DNA Mini kit (Qiagen, Hilden, Germany) をそれぞれ使用して、ゲノム DNA を抽出した。PCR 増幅には KOD FX PCR enzyme (Toyobo, Shiga, Japan) を使用した。PTH 遺伝子 上流の再構成の解析は、PTH 遺伝子の転写開始点 (TSS; Transcription start site) から上流約 21 キロ塩基対までの DNA 領域をカバーする、平均 5 キロ塩基対 のそれぞれ両端が重なり合う 4 つの断片に分けて増幅し、副甲状腺腺腫と
MCF-7 細胞からの PCR 産物と比較した。プロモーター領域の塩基配列解析は、
PTH 遺伝子の転写開始点から 5.7 キロ塩基対上流~ 360 塩基対下流の DNA 領域を、約 800 から 1000 塩基対の長さのそれぞれ両端が重なり合う 9 つの DNA 断片に分けて PCR 増幅し、pBluescript II SK (+) phagemid vector にクロー ン化し、2 つから 4 つのクローンの塩基配列を Applied Biosystems 3730xl DNA アナライザを使用して決定した。
第6節 PTH 遺伝子プロモーター領域 CpG 部位のメチル化解析
PTH 遺伝子プロモーター領域の CpG 部位のメチル化状態を、バイサルファ イトシーケンシング法で解析した。本腫瘍と副甲状腺腺腫からゲノム DNA を 抽出し (Wizard SV Genomic DNA Purification System (Promega, Wisconsin, USA))、 MethylEasy Xceed Rapid DNA Bisulphite Modification Kit (Human Genetic Signatures,
Sydney, Australia) を使用してバイサルファイト処理し、バイサルファイト処理後
の配列 にあ わせ て設 計し たプ ライ マー (表 2) と TaKaRa Taq Hot Version (Takara) を使用して PCR 増幅した。PCR 産物を、pGEM-T Easy vector System (Promega, Wisconsin, USA) を使用してクローン化し、本腫瘍からの 9 つのクロ
7
ーンと副甲状腺腺腫からの 3 つのクローンの 2 本鎖の塩基配列を Applied Biosystems 3730xl DNA アナライザを使用して決定した。
第7節 マイクロアレイによる網羅的遺伝子発現解析
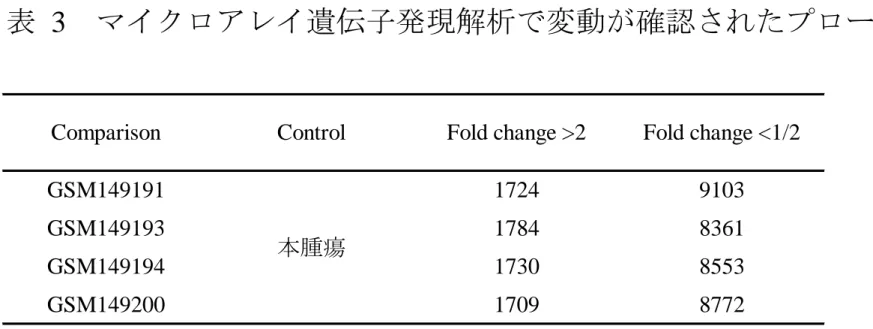
本症例に対して、マイクロアレイによる遺伝子発現解析を施行し、NCBI の Gene Expression Omnibus (GEO) に GSE6481 として登録されている PTH 非産 生 MFH [42] の中の 4 症例 (GSM149191, 149193, 149194, 149200) と比較した。
凍結保存していた本腫瘍から RNA を抽出し、250 ng をビオチンで標識した。
ビオチン標識 cRNA 10 μg を、GeneChip 3’IVT Express Kit User Manual に従い Human Genome U133 Plus 2.0 Array を使用して遺伝子特異的プローブにハイブ リ ダ イ ゼ ー シ ョ ン さ せ 、 各 プ ロ ー ブ の シ グ ナ ル 値 を Affymetrix GeneChip Command Console Software (AGCC) を使用して算出した。本腫瘍のマイクロアレ イデータのスケーリングを GSE6481 データのそれに合わせるために、シグナル 値のトリム平均が 100 になるように Affymetrix Expression Console Software (Expression Console) を使用してデータを正規化した (MAS 5.0 アルゴリズム)。 GSE6481 の Affymetrix Human Genome U133A Array に共通する Probe Set ID を抽出した。得られたプローブに対して、発現比を Log2 (PTH 非産生比較検体
/ PTH 産生対照検体) に基づいて算出した。発現比が 2 倍以上もしくは 1/2 倍
以下のプローブセットを抽出した。各比較において、発現上昇が観察された遺 伝子リストと発現抑制が観察された遺伝子リストを抽出し、4 比較分の合計 8 グループを作成した。変動遺伝子の抽出により得られたグループに含まれる遺 伝子の生物学的特徴を知るために、Gene Ontology (GO) term の出現頻度による 解析を行った。まず、各グループに含まれる遺伝子の annotation (NCBI Entrez Gene の Gene ID) を元に、それぞれの遺伝子が持つ GO の全 term を決定した。
8
次に、グループ毎に GO term の出現頻度を集計し、各 term の出現頻度が NCBI Entrez Gene に記載されている GO term を持つ遺伝子全体での term の出現頻 度と比べ有意であるかの検定を行い p-value を算出した。
9
第3章 結果
第1節 腫瘍細胞での PTH タンパク質の検出
摘出腫瘍の一部を培地 DMEM (0.1 % BSA 添加) 中で 2 時間インキュベー ションし、その上清の whole-PTH 濃度を測定したところ 4830 pg/mL (基準範囲
10-39) と高値で、腫瘍が PTH を産生していたことが示唆された (図 2 A)。さ
らに、本腫瘍が実際に PTH を産生していることを別の方法で確認する目的で、
ホルマリン固定したパラフィン包埋切片を抗 PTH 抗体を用いて免疫組織化学 染色を行った (図 2 B-D)。紡錘形の腫瘍細胞と巨大な核を持つ腫瘍細胞の細胞 質が、抗 PTH 抗体により染色された (図 2 B)。陽性対照の副甲状腺主細胞は 同じ抗体で染色された (図 2 C) のに対し、本腫瘍の腫瘍細胞は正常マウスの IgG で染色されなかった (図 2 D)。このことから、実験室での培養条件下でな くても、本腫瘍が実際に PTH タンパク質を産生していることが確認できた。
第2節 腫瘍細胞での PTH mRNA の過剰発現
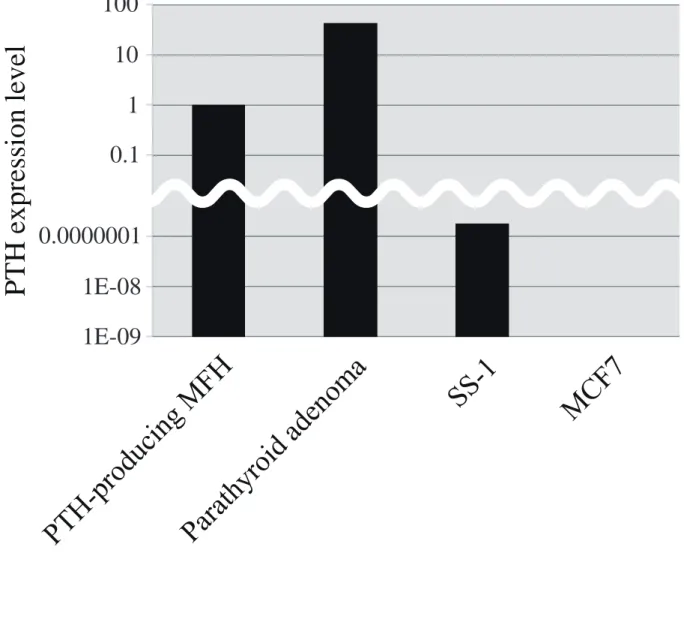
本症例における異所性 PTH タンパク質の産生が、PTH の mRNA の増加に よるものなのか、あるいは PTH タンパク質の安定化によるものなのかを区別す る目的で、本腫瘍における PTH mRNA の量を qRT- PCR により測定した。RNA 抽出後、逆転写酵素反応を行い、定量的 PCR にかけた。GAPDH との相対的発 現量で算出した (図 3)。本腫瘍における PTH の mRNA 量は、陽性対照の SS-1 細胞のそれよりもはるかに多く、ヒト副甲状腺腺腫の mRNA 量と比肩しうるレ ベルであった。本腫瘍における PTH mRNA の過剰発現は、マイクロアレイ解 析でも確認された (表 3)。
10
第3節 PTH 遺伝子の構造解析
異所性 PTH 産生卵巣癌症例において、PTH 遺伝子とその上下流における再 構成と増幅が PTH 産生に関係している症例があることから [17]、PTH 遺伝子 の遺伝子再編成と塩基配列変化を検討した。まず、PTH 遺伝子のプロモーター 領域を含む上流の大規模な再構成がないかどうかを調べるために、PTH 遺伝子 の転写開始点から 21 キロ塩基対上流の領域を、それぞれ両端が重なり合う 4
つの DNA 断片として、PCR 増幅を試みた。4 つとも、サイズの変化やバンド
の消失を伴わずに増幅することができた (図は示さず)。従って、少なくともひ とつのアレル上の PTH 遺伝子プロモーター領域は遺伝子再編成のような大き な構造変化はないようであった。
次に、PTH mRNA のコード領域および非コード領域の変異が mRNA を安定
させた可能性を検討する目的で、腫瘍由来の PTH cDNA の塩基配列を決定した が変異は発見されなかった。さらに、転写因子の結合性を変化させ、PTH 過剰 産生に寄与する可能性を検討するために、本腫瘍のゲノム DNA からプロモー ターを増幅し、それぞれ塩基配列を決定した。cDNA とプロモーター領域 (5.7 キロ塩基対) に変異は検出できなかった (図 4)。
以上から、PTH 遺伝子のプロモーターを含めた領域の構造異常は見いだせな かった。
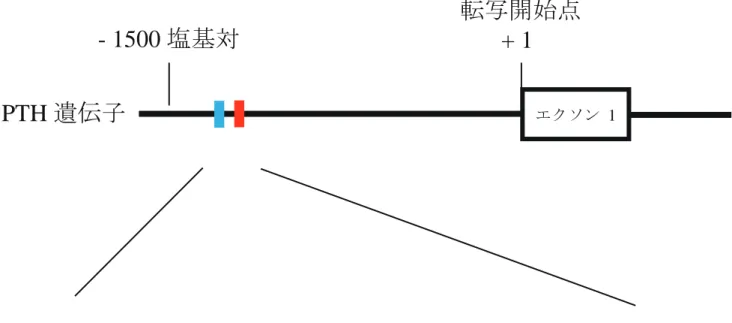
第4節 PTH 遺伝子プロモーター領域 CpG 部位のメチル化解析
PTH 産生膵神経内分泌癌症例において PTH 遺伝子の低メチル化が報告され ており [18]、本症例での PTH 遺伝子のプロモーター領域の低メチル化による 転写活性化の可能性を検討する目的で、CpG ジヌクレオチドのメチル化状態を バイサルファイト法で解析した。本腫瘍と副甲状腺腺腫からのゲノム DNA を
11
バイサルファイト処理し、2 か所の CpG 部位を含む 232 塩基対の断片を PCR 増幅後、ベクターにクローン化し、塩基配列を決定した (図 5)。副甲状腺腺腫
では、2 か所とも CpG 部位が脱メチル化されていたのに対して、本腫瘍では、
2 か所ともにメチル化状態であった。このことから、PTH 遺伝子プロモーター
領域の CpG ジヌクレオチドの低メチル化によって、本腫瘍における異所性
PTH 産生が起こった可能性は低いことが示唆された。
第5節 網羅的遺伝子発現解析
今までの結果から、PTH mRNA の過剰は認めるけれども、PTH 遺伝子の構造 変異もなく、プロモーター領域の低メチル化もなかったことが明らかとなった。
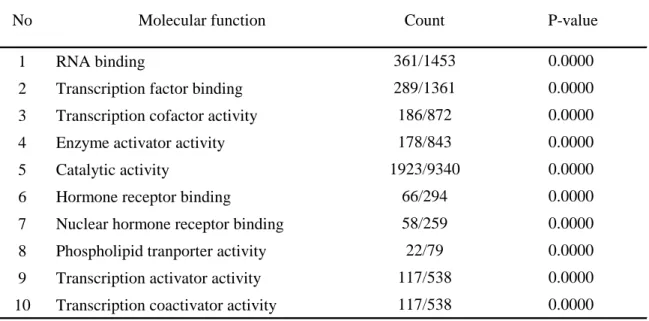
次の可能性として、本腫瘍におけるグローバルな転写のパターンの変化によっ て、PTH 遺伝子の転写活性化がもたらされたという仮説をたてた。PTH 遺伝子 の転写を高める転写因子を網羅的に把握する目的で、マイクロアレイによる網 羅的遺伝子発現解析を施行した。対照として PTH 非産生 MFH 4 症例 (市川論 文) を選択した。本腫瘍は、PTH 非産生 MFH と比較して、8000 以上のプロー ブで遺伝子発現が上昇し、1700 以上のプローブで遺伝子発現が抑制されていた (表 3)。発生部位が後腹膜腔である GSM149194 と本腫瘍とを比較して、本腫瘍 で活性化されていた発現比が上位の 10 種の中に、第 2 番目に PTH があり (表 4)、また他の PTH 非産生 MFH との比較でも同様に PTH 遺伝子の高発現 を認めた。GO 解析で、いずれの PTH 非産生 MFH 症例との比較でも本腫瘍 において活性化されていたもしくは抑制されていた遺伝子群をみると、多くの 細胞学的な部位において多くの生物学的な機能が修飾されており、特定の細胞 内機能を抽出し、本腫瘍を特徴づけることは困難であった (表5-7)。ただし、分 子機能の GO 解析において、発現上昇上位群の中に RNA 結合や転写因子とい
12
った、DNA もしくは RNA に結合するタンパク質の発現上昇が認められた (表 7) ことから、PTH 遺伝子が転写的に活性化された、あるいは PTH mRNA がタ ンパク質により安定化する可能性が示唆された。
そのことをふまえ、副甲状腺の発生に関係する転写因子 (Hoxa3, Pbx1, Pax1, Pax3, Pax9, Eya1, Six1, Tbx1, Gcm2, Gata3) [43]、ヒトPTHプロモーター領域に結 合する転写活性化因子 (Sp1, Sp3, NF-Yα, NF-Yβ, NF-Yγ) [44]、PTH の 3’ 非翻訳 領域に結合し PTH の mRNA の安定性に関係する遺伝子群 (AUF1, Unr, KSRP,
Pin1) [45] に着目し、マイクロアレイのデータを精査した (図 6 A-C)。副甲状腺
発生関連遺伝子群 (図 6 A) は、どの PTH 非産生MFH と比較しても、PTH 産 生 MFH において、Gcm2 の発現低下を認めた以外は、顕著な発現の差を認め なかった。Gcm2 は副甲状腺発生の主たる調節遺伝子 [46] であるが、PTH 産 生 MFH だけでなく PTH 非産生 MFH でもシグナル値は極めて小さかった。
一方、ヒト PTH プロモーター領域に結合する転写遺伝子群の中で、Sp3 と NF-Yβ の発現量が、どの PTH 非産生 MFH と比較しても増加していた (図 6 B)。Sp3 と NF-Yβ とは協同して、PTH プロモーターを活性化する (文献 44 の 図 3) ことが知られている。PTH の mRNA の安定性に影響する因子群では、
AUF1、Unr の発現が PTH 産生腫瘍において増加していた (図 6 C)。AUF1 も Unr の発現が PTH の mRNA に結合し、安定化させる機能がある [45]。Sp タ ンパク質も NF-Y タンパク質も p300/CBP や PCAF といったヒストンアセチ ル化酵素活性を有する転写因子と結合・動員して [47-49]、近傍のクロマチンを 開いた構造にする機能があるので、本腫瘍において、p300/CBP や PCAF を含 めたヒストンアセチル化酵素の発現を精査した (図 6 D)。PTH 非産生MFH と 比較して、PTH 産生 MFH では、p300、PCAF、GCN5、Hat1 の発現上昇が見 られた。
13
第4章 考察
異所性 PTH 産生腫瘍の新規症例
高 Ca クリーゼ症を呈し、しかも、PTHrP でなく PTH を異所性に産生する 腫瘍に起因するという極めて珍しい症例を報告する。今回の症例については、(i) 腫瘍の存在に依存した高 Ca 血症 (図 1)、(ii) 摘出腫瘍の培養上清中の PTH の 検出 (図 2 A)、(iii) 腫瘍の PTH 抗体による免疫化学染色 (図 2 B)、(iv) qRT-PCR 法およびマイクロアレイ解析による PTH の発現上昇 (図 3、図 6) の 結果から、PTH の mRNA 量が高まることによって、本腫瘍が異所性に PTH を 産生したと推論した。
副甲状腺以外の組織から発生した腫瘍による異所性 PTH 産生の報告は極め て稀で、1988 年以降で真の異所性 PTH 産生腫瘍は 23 例しかない (表 8)
[17-39]。おそらく、PTHrP と異なり、PTHは副甲状腺組織特異的に発現するよ
うに厳格にその発現が制御されているためであろう。副甲状腺組織以外の組織
が PTH を異所性に産生する機序解明は、患者の予後改善の面からも副甲状腺の
発生・分化の理解を深める面からも追究されるべき研究課題である。
PTH 産生の機序について
本腫瘍では PTH 遺伝子とその上流に変異はなく、CpG サイトの低メチル化 もみられず、本腫瘍における異所性 PTH 産生にはこれまで報告されてきたもの とは別の機構が原因と考えられた。マイクロアレイによる網羅的遺伝子発現解 析により (図 6)、本腫瘍において、PTH 遺伝子発現を正に制御するタンパク質 の発現上昇を見出した。具体的には、Sp3、NF-Yβ、AUF1、Unr、p300、PCAF で ある。PTH 遺伝子のプロモーター -178~+21 塩基対領域内に Sp タンパク質の
14
結合部位が 1 つ、NF-Y 結合部位が 2 つ存在する。Sp3 と NF-Yβ を SL2 細 胞に外来性に導入後、上述のプロモーター領域にルシフェラーゼをつないだレ ポーターでアッセイすると、プロモーター活性が協同的に高まる [44]。Sp タン パク質や NF-Y タンパク質と結合する p300/CBP や PCAF といったヒストン アセチル化酵素 [47-49] の発現も本腫瘍で上昇していた。これらの結果から、
本腫瘍において、発現上昇した Sp3 タンパク質および NF-Yβ タンパク質が PTH 遺伝子のプロモーター領域に結合し、p300/CBP や PCAF のヒストンアセ チル化酵素を動員し、近傍のコアヒストンのアセチル化を高め、クロマチン構 造を開いた状態にすることで、PTH 遺伝子の発現上昇を促進したと推察した。
こういう状況に加え、転写された PTH の mRNA の 3’ 非翻訳領域に、RNA 結 合タンパク質である AUF1 タンパク質、Unr タンパク質が結合し、PTH の
mRNA を安定化することで、PTH の発現上昇に貢献していると推測された。
本症例の異分化がんとしての可能性
本症例のがん発生メカニズムとして、原発組織が副甲状腺に分化を遂げた結 果、PTH を過剰産生した可能性が考えられる。しかしながら、副甲状腺の発生・
分化に重要な Gcm2 の発現は、本症例においてはシグナル値が小さく、PTH 非 産生 MFH と比較しても低下していた (図 6 A)。さらに、副甲状腺の腫瘍化に 関係する遺伝子 [50] のうち、PRAD1、HRPT2 および Menin の発現は本腫瘍 では副甲状腺腺腫や MCF7 と比較して顕著な差はなく、RET の発現は本腫瘍 で観察されなかった (データ示さず)。以上の実験結果から、本症例が、副甲状 腺に分化した後に腫瘍化した可能性は、低いと考えられた。
15
MFH という組織型から見た本症例の特異性
本症例は MFH で、PTH 産生源としては初の報告である。MFH は「病理組 織学的に細胞起源が不明で、他の肉腫としては分類できない高悪性度多形成肉 腫」であることを特徴とする [51]。
MFH は成人の悪性軟部腫瘍の中で最も頻度が高い (26.7 - 40 %)。MFH の原 発部位は、四肢が最多で、後腹膜原発は比較的少ない (4 - 16 %) [52]。後腹膜原
発 MFH は、自覚症状に乏しく発見が遅れ、多くは 10 cm 以上の腫瘍径でみつ
かり、外科的切除の成績も良くない [53]。MFH の標準的な化学療法は確立され ていない。本症例は、PTH 産生による症状が出現したことによって、10 cm 以 下の腫瘍径にも関わらず、比較的早期に発見できた、と考える。
WHO 分類では、免疫組織化学や遺伝子診断技術の進歩もあり、2002 年に粘 液線維肉腫、多形型平滑筋肉腫、未分化多形肉腫などに再分類され、2013 年に は未分化多形肉腫がさらに未分化 / 未分類肉腫 (undifferentiated unclassified
sarcoma) という名称に改められ MFH の腫瘍名は除外された。今後、MFH と
いう名称は次第に使用されなくなるが、本症例を診療・診断した時代の分類と、
これまでの MFH に関する研究結果を引用したことなどもふまえ、本論文では MFH という名称を一貫して使用した。
16
第5章 結論
異所性 PTH 産生 MFH を初めて報告した。PTH 遺伝子上流の再構成や PTH 遺伝子プロモーター領域の変異、CpG 部位の低メチル化はみられなかった。マ イクロアレイによる網羅的遺伝子発現解析により、本腫瘍において、PTH プロ モーター領域に結合する転写因子である NF-Yβ と Sp3、さらにそれらと結合す るヒストンアセチル化酵素である p300、CBP、PCAF の発現が上昇していたこ とが明らかとなった。また、PTH の mRNA の非翻訳領域に結合し、mRNA を 安定化する AUF1、Unr の発現も増加していた。クロマチン構造を開いた状態 にして、PTH 遺伝子の転写が高められると同時に、PTH の mRNA が安定化を 受けて、PTH 遺伝子の過剰発現が導かれたと推測された。
17
謝辞
稿を終えるにあたり、本研究の機会を与えて下さり、全般的なご指導をいた だきました防衛医科大学校総合臨床部、田中祐司教授に深甚なる感謝の意を表 します。
本研究を実施するにあたり、研究方法および論文作成に直接ご指導をいただ きました帝京大学医学部生化学講座、岡崎具樹教授、飯塚眞由准教授、安達三 美准教授に深い感謝を申し上げます。
また、研究に際し貴重なご助言ご指導を下さいました防衛医科大学校総合臨 床部、大渕康弘准教授、廣岡伸隆准教授に感謝申し上げます。
18
引用文献
1. Parma J, Duprez L, Van Sande J, Cochaux P, Gervy C, Mockel J, Dumont J, Vassart G, Somatic mutations in the thyrotropin receptor gene cause hyperfunctioning thyroid adenomas. Nature 1993;365: 649-51.
2. Parma J, Duprez L, Van Sande J, Hermans J, Rocmans P, Van Vliet G, Costagliola S, Rodien P, Dumont JE, Vassart G, Diversity and prevalence of somatic mutations in the thyrotropin receptor and Gs alpha genes as a cause of toxic thyroid adenomas. J Clin Endocrinol Metab 1997;82: 2695-701.
3. Suarez HG, du Villard JA, Caillou B, Schlumberger M, Parmentier C, Monier R, gsp mutations in human thyroid tumours. Oncogene 1991;6: 677-9.
4. Modlin IM, Champaneria MC, Bornschein J, Kidd M, Evolution of the diffuse neuroendocrine system--clear cells and cloudy origins. Neuroendocrinology 2006;84:
69-82.
5. Strewler GJ, Humoral manifestations of malignancyWilliams Textbook of Endocrinology. Philadelphia: Saunders, 2002:1834-56.
6. Baylin SB, Mendelsohn G, Ectopic (inappropriate) hormone production by tumors:
mechanisms involved and the biological and clinical implications. Endocr Rev 1980;1: 45-77.
7. Baylin SB, Herman JG, Graff JR, Vertino PM, Issa JP, Alterations in DNA methylation: a fundamental aspect of neoplasia. Adv Cancer Res 1998;72: 141-96.
8. Wiesener MS, Seyfarth M, Warnecke C, Jurgensen JS, Rosenberger C, Morgan NV, Maher ER, Frei U, Eckardt KU, Paraneoplastic erythrocytosis associated with an inactivating point mutation of the von Hippel-Lindau gene in a renal cell carcinoma.
Blood 2002;99: 3562-5.
9. Rad FH, Ulusakarya A, Gad S, Sibony M, Juin F, Richard S, Machover D, Uzan G, Novel somatic mutations of the VHL gene in an erythropoietin-producing renal carcinoma associated with secondary polycythemia and elevated circulating endothelial progenitor cells. Am J Hematol 2008;83: 155-8.
10. Gong K, Zhang N, Zhang K, Na Y, The relationship of erythropoietin overexpression with von Hippel-Lindau tumour suppressor gene mutations between hypoxia-inducible factor-1alpha and -2alpha in sporadic clear cell renal carcinoma.
Int J Mol Med 2010;26: 907-12.
11. Newell-Price J, King P, Clark AJ, The CpG island promoter of the human proopiomelanocortin gene is methylated in nonexpressing normal tissue and tumors and represses expression. Mol Endocrinol 2001;15: 338-48.
12. Newell-Price J, Proopiomelanocortin gene expression and DNA methylation:
19
implications for Cushing's syndrome and beyond. J Endocrinol 2003;177: 365-72.
13. Case Records of the Massachusetts General Hospital. Case 27461. N Engl J Med 1941;225: 789-91.
14. Grill V, Rankin W, Martin TJ, Parathyroid hormone-related protein (PTHrP) and hypercalcaemia. Eur J Cancer 1998;34: 222-9.
15. Stewart AF, Clinical practice. Hypercalcemia associated with cancer. N Engl J Med 2005;352: 373-9.
16. Clines GA, Mechanisms and treatment of hypercalcemia of malignancy. Curr Opin Endocrinol Diabetes Obes 2011;18: 339-46.
17. Nussbaum SR, Gaz RD, Arnold A, Hypercalcemia and ectopic secretion of parathyroid hormone by an ovarian carcinoma with rearrangement of the gene for parathyroid hormone. N Engl J Med 1990;323: 1324-8.
18. VanHouten JN, Yu N, Rimm D, Dotto J, Arnold A, Wysolmerski JJ, Udelsman R, Hypercalcemia of malignancy due to ectopic transactivation of the parathyroid hormone gene. J Clin Endocrinol Metab 2006;91: 580-3.
19. Demura M, Yoneda T, Wang F, Zen Y, Karashima S, Zhu A, Cheng Y, Yamagishi M, Takeda Y, Ectopic production of parathyroid hormone in a patient with sporadic medullary thyroid cancer. Endocr J 2010;57: 161-70.
20. Abe Y, Makiyama H, Fujita Y, Tachibana Y, Kamada G, Uebayashi M, Severe hypercalcemia associated with hepatocellular carcinoma secreting intact parathyroid hormone: a case report. Intern Med 2011;50: 329-33.
21. Iguchi H, Miyagi C, Tomita K, Kawauchi S, Nozuka Y, Tsuneyoshi M, Wakasugi H, Hypercalcemia caused by ectopic production of parathyroid hormone in a patient with papillary adenocarcinoma of the thyroid gland. J Clin Endocrinol Metab 1998;83: 2653-7.
22. Koyama Y, Ishijima H, Ishibashi A, Katsuya T, Ishizaka H, Aoki J, Endo K, Intact PTH-producing hepatocellular carcinoma treated by transcatheter arterial embolization. Abdom Imaging 1999;24: 144-6.
23. Nakajima K, Tamai M, Okaniwa S, Nakamura Y, Kobayashi M, Niwa T, Horigome N, Ito N, Suzuki S, Nishio S, Komatsu M, Humoral hypercalcemia associated with gastric carcinoma secreting parathyroid hormone: a case report and review of the literature. Endocr J 2013;60: 557-62.
24. Ohira S, Itoh K, Shiozawa T, Horiuchi A, Ono K, Takeuchi H, Hosoda W, Konishi I, Ovarian non-small cell neuroendocrine carcinoma with paraneoplastic parathyroid hormone-related hypercalcemia. Int J Gynecol Pathol 2004;23: 393-7.
25. Uchimura K, Mokuno T, Nagasaka A, Hayakawa N, Kato T, Yamazaki N, Kobayashi
20
T, Nagata M, Kotake M, Itoh M, Tsujimura T, Iwase K, Lung cancer associated with hypercalcemia induced by concurrently elevated parathyroid hormone and parathyroid hormone-related protein levels. Metabolism 2002;51: 871-5.
26. Wong K, Tsuda S, Mukai R, Sumida K, Arakaki R, Parathyroid hormone expression in a patient with metastatic nasopharyngeal rhabdomyosarcoma and hypercalcemia.
Endocrine 2005;27: 83-6.
27. Yoshimoto K, Yamasaki R, Sakai H, Tezuka U, Takahashi M, Iizuka M, Sekiya T, Saito S, Ectopic production of parathyroid hormone by small cell lung cancer in a patient with hypercalcemia. J Clin Endocrinol Metab 1989;68: 976-81.
28. Strewler GJ, Budayr AA, Clark OH, Nissenson RA, Production of parathyroid hormone by a malignant nonparathyroid tumor in a hypercalcemic patient. J Clin Endocrinol Metab 1993;76: 1373-5.
29. Botea V, Edelson GW, Munasinghe RL, Hyperparathyroidism, hypercalcemia, and calcified brain metastatic lesions in a patient with small cell carcinoma demonstrating positive immunostain for parathyroid hormone. Endocr Pract 2003;9:
40-4.
30. Rizzoli R, Pache JC, Didierjean L, Bürger A, Bonjour JP, A thymoma as a cause of true ectopic hyperparathyroidism. J Clin Endocrinol Metab 1994;79: 912-5.
31. Bernini M, Bacca A, Casto G, Carli V, Cupisti A, Carrara D, Farnesi I, Barsotti G, Naccarato AG, Bernini G, A case of pheochromocytoma presenting as secondary hyperaldosteronism, hyperparathyroidism, diabetes and proteinuric renal disease.
Nephrol Dial Transplant 2011;26: 1104-7.
32. Bhattacharya A, Mittal BR, Bhansali A, Radotra BD, Behera A, Cervical paraganglioma mimicking a parathyroid adenoma on Tc-99m sestamibi scintigraphy.
Clin Nucl Med 2006;31: 234-6.
33. Buller R, Taylor K, Burg AC, Berman ML, DiSaia PJ, Paraneoplastic hypercalcemia associated with adenosquamous carcinoma of the endometrium. Gynecol Oncol 1991;40: 95-8.
34. Chen L, Dinh TA, Haque A, Small cell carcinoma of the ovary with hypercalcemia and ectopic parathyroid hormone production. Arch Pathol Lab Med 2005;129: 531-3.
35. Kandil E, Noureldine S, Khalek MA, Daroca P, Friedlander P, Ectopic secretion of parathyroid hormone in a neuroendocrine tumor: a case report and review of the literature. Int J Clin Exp Med 2011;4: 234-40.
36. Mahoney EJ, Monchik JM, Donatini G, De Lellis R, Life-threatening hypercalcemia from a hepatocellular carcinoma secreting intact parathyroid hormone: localization by sestamibi single-photon emission computed tomographic imaging. Endocr Pract
21
2006;12: 302-6.
37. Morita SY, Brownlee NA, Dackiw AP, Westra WH, Clark DP, Zeiger MA, An unusual case of recurrent hyperparathyroidism and papillary thyroid cancer. Endocr Pract 2009;15: 349-52.
38. Nielsen PK, Rasmussen AK, Feldt-Rasmussen U, Brandt M, Christensen L, Olgaard K, Ectopic production of intact parathyroid hormone by a squamous cell lung carcinoma in vivo and in vitro. J Clin Endocrinol Metab 1996;81: 3793-6.
39. Vacher-Coponat H, Opris A, Denizot A, Dussol B, Berland Y, Hypercalcaemia induced by excessive parathyroid hormone secretion in a patient with a neuroendocrine tumour. Nephrol Dial Transplant 2005;20: 2832-5.
40. Ishikawa M, Kimura K, Tachibana T, Hashimoto H, Shimojo M, Ueshiba H, Tsuboi K, Shibuya K, Yoshino G, Establishment and characterization of a novel cell line derived from a human small cell lung carcinoma that secretes parathyroid hormone, parathyroid hormone-related protein, and pro-opiomelanocortin. Hum Cell 2010;23:
58-64.
41. Soule HD, Vazguez J, Long A, Albert S, Brennan M, A human cell line from a pleural effusion derived from a breast carcinoma. J Natl Cancer Inst 1973;51: 1409-16.
42. Nakayama R, Nemoto T, Takahashi H, Ohta T, Kawai A, Seki K, Yoshida T, Toyama Y, Ichikawa H, Hasegawa T, Gene expression analysis of soft tissue sarcomas:
characterization and reclassification of malignant fibrous histiocytoma. Mod Pathol 2007;20: 749-59.
43. Grigorieva IV, Thakker RV, Transcription factors in parathyroid development:
lessons from hypoparathyroid disorders. Ann N Y Acad Sci 2011;1237: 24-38.
44. Alimov AP, Park-Sarge OK, Sarge KD, Malluche HH, Koszewski NJ, Transactivation of the parathyroid hormone promoter by specificity proteins and the nuclear factor Y complex. Endocrinology 2005;146: 3409-16.
45. Naveh-Many T, Minireview: the play of proteins on the parathyroid hormone messenger ribonucleic Acid regulates its expression. Endocrinology 2010;151:
1398-402.
46. Günther T, Chen ZF, Kim J, Priemel M, Rueger JM, Amling M, Moseley JM, Martin TJ, Anderson DJ, Karsenty G, Genetic ablation of parathyroid glands reveals another source of parathyroid hormone. Nature 2000;406: 199-203.
47. Currie RA, NF-Y is associated with the histone acetyltransferases GCN5 and P/CAF.
J Biol Chem 1998;273: 1430-4.
48. Salsi V, Caretti G, Wasner M, Reinhard W, Haugwitz U, Engeland K, Mantovani R, Interactions between p300 and multiple NF-Y trimers govern cyclin B2 promoter
22
function. J Biol Chem 2003;278: 6642-50.
49. Huang W, Zhao S, Ammanamanchi S, Brattain M, Venkatasubbarao K, Freeman JW, Trichostatin A induces transforming growth factor beta type II receptor promoter activity and acetylation of Sp1 by recruitment of PCAF/p300 to a Sp1.NF-Y complex.
J Biol Chem 2005;280: 10047-54.
50. Sharretts JM, Simonds WF, Clinical and molecular genetics of parathyroid neoplasms. Best Pract Res Clin Endocrinol Metab 2010;24: 491-502.
51. Fletcher CD, Pleomorphic malignant fibrous histiocytoma: fact or fiction? A critical reappraisal based on 159 tumors diagnosed as pleomorphic sarcoma. Am J Surg Pathol 1992;16: 213-28.
52. Weiss SW, Enzinger FM, Malignant fibrous histiocytoma: an analysis of 200 cases.
Cancer 1978;41: 2250-66.
53. Kearney MM, Soule EH, Ivins JC, Malifnant fibrous histiocytoma: a retrospective study of 167 cases. Cancer 1980;45: 167-78.
23
表 1 入院時検査所見
Albumin (g/dL) BUN (mg/dL) Creatinine (mg/dL) Calcium (mg/dL) Phosphate (mg/dL) Magnesium (mg/dL) Intact-PTH (pg/mL) PTHrP (pmol/L)
1, 25 dihydroxyvitamin D3 (pg/mL)
BUN, blood urea nitrogen; PTHrP, parathyroid hormone related protein.
56 2.43 17.6 4.6
<1.0 6.8
基準範囲
1.9-2.5 3.9-4.9 8.0-20.0
0.5-1.0 8.4-10.4
2.5-4.5
10-65
<1.0 20-60 3.7
532 3.4
24
表 2 PCR で使用したプライマーの配列
PTH GGGTCTGCAGTCCAATTCAT CAGATTTCCCATCCGATTTT
Gcm2 AGTTGATTCCTTGTCGAGGGCACA TGTTGCTGAAATGACCACTGCTGTC
PRAD1 TCCTCTCCAAAATGCCAGAG TGAGGCGGTAGTAGGACAGG
HRPT2 TTCCTGCAGCTACCACCTCT GGTCCCAGTCTTGAGGCATA
Menin GACCCACTCACCCTCTACCA CCCGGCAGTAGTTGTAGTCC
RET ACAGGGGATGCAGTATCTGG CCTGGCTCCTCTTCACGTAG
GAPDH GCACCGTCAAGGCTGAGAAC TGGTGAAGACGCCAGTGGA
PTH の cDNA 解析 TCAGCATCAGCTACTAACATACCTG GGTAGAAGAGAGGTTTAAAAGTGCAA
バイサルファイト解析 AAGTGAGGTTAATTTAGAGTGGAGAA AAACAACTTTACCTTCCCACCA Forward (左側 5'、右側 3') Reverse (左側 5'、右側 3')
25
表 3 マイクロアレイ遺伝子発現解析で変動が確認されたプローブ数
GSM149191 GSM149193 GSM149194 GSM149200
Comparison Control Fold change >2 Fold change <1/2
本腫瘍
1724 1784 1730 1709
9103 8361 8553 8772
26
表 4 GSM149194 との比較で活性化もしくは 抑制された遺伝子群
活性化上位群
抑制上位群
Chitinase 3-like 1 (cartilage glycoprotein-39) Parathyroid hormone
Chemokine (C-C motif) ligand 20
Cell adhesion molecule with homology to L1CAM Protein tyrosine phosphatase, receptor type, D Coiled-coil domain containing 88A
Glycophorin A (MNS blood group)
Tumor necrosis factor receptor superfamily, member 11b Interleukin 26
Podoplanin 4982
55801 10630
-11.16 -11.07 -10.34 -10.07 -9.70
-9.66 -8.97 -8.79 -8.72 -8.69 6364
10752 5789 55704
2993
Entrez gene link Gene title Fold change
1116 5741
Neurotrophic tyrosine kinase, receptor, type 2 Chromosome 10 open reading frame 116 Glutamate receptor, metabotropic 1
Aldo-keto reductase family 1, member B10 (aldose reductas Centromere protein T
Glutamate receptor, ionotropic, AMPA 1 Hypothetical MGC13053
Interleukin 20 receptor, alpha
Epoxide hydrolase 1, microsomal (xenobiotic) Family with sequence similarity 182, member B
6.59 6.57 6.49 4915
10974 2911 57016 80152 2890 84796 53832 2052 728882
7.48 7.02 6.89 6.73 6.68
Entrez gene link Gene title Fold change
9.23 7.91
27
表 5 全ての比較で活性化もしくは抑制された GO biological process 上位群
活性化上位群
抑制上位群
1 Cellular metabolic process 2 Primary metabolic process 3 Metabolic process
4 Macromolecule metabolic process
5 Cellular macromolecule metabolic process 6 Biopolymer metabolic process
7 Cellular biopolymer metabolic process 8 Cellular protein metabolic process 9 Protein metabolic process
10 Nitrogen compound metabolic process
1353/6401 2018/9505
0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 3056/14668
3317/16065
No Biological process Count
2298/10738 1155/5380
P-value 3051/14724
2605/12330 2378/11180 2503/11772
1 Positive regulation of muscle cell differentia 2 Regulation of synapse organization
3 Cognition
4 Regulation of synaptogenesis
5 Regulation of synapse structure and activity 6 Digestion
7 Response to light stimulus
8 Regulation of calcium ion transport 9 Dendrite morphogenesis
10 Anion transport
P-value 14/113
9/58 11/91 20/219
No Biological process Count
28/361 22/281 11/102 25/318
0.0000 0.0000 0.0000 0.0001 0.0001 0.0003 0.0004 0.0004 0.0004 0.0005 11/72
77/1136
28
表 6 全ての比較で活性化もしくは抑制された GO cellular component 上位群
活性化上位群
抑制上位群
1 Membrane-bounded organelle
2 Intracellular membrane-bounded organelle 3 Nucleolus
4 Nuclear body 5 Cell part 6 Cell
7 Ribonucleoprotein complex 8 Nuclear speck
9 PML body
10 Organelle membrane
46/193 808/3998
0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 3377/16054
641/2850
No Cellular component Count
242/922 77/310
P-value 3381/16084
143/591 5065/24992 5065/24993
1 Ion channel complex 2 Postsynaptic membrane 3 Contractile fiber 4 Contractile fiber part 5 Actin cytoskeleton 6 Chloride channel complex 7 Cytoskeletal part
8 Myofibril 9 Sarcomere 10 Stress fiber
18/232 9/109
0.0000 0.0000 0.0000 0.0001 0.0001 0.0002 0.0006 0.0007 0.0014 0.0054 31/353
25/304
No Cellular component Count
121/2059 21/280
P-value 37/371
22/263 45/711 13/86
29
表 7 全ての比較で活性化もしくは抑制された GO molecular function 上位群
活性化上位群
抑制上位群
1 RNA binding
2 Transcription factor binding 3 Transcription cofactor activity 4 Enzyme activator activity 5 Catalytic activity
6 Hormone receptor binding
7 Nuclear hormone receptor binding 8 Phospholipid tranporter activity 9 Transcription activator activity 10 Transcription coactivator activity
P-value 361/1453
178/843 1923/9340
66/294
No Molecular function Count
58/259 22/79 117/538 117/538
0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 289/1361
186/872
1 Ligand-gated channel activity 2 Ligand-gated ion channel activity 3 Transporter activity
4 Neurotransmitter binding 5 Molecular transducer activity 6 Signal transducer activity
7 Transmembrane transporter activity
9 Passive transmembrane tranporter activity 10 Channel activity
P-value 27/299
19/149 216/3727 216/3727
No Molecular function Count
106/1646 Substrate-specific transmembrane
transporter activity
8 98/1521
58/766 58/766
0.0000 0.0000 0.0000 0.0000 0.0000 0.0000 0.0000
0.0000 0.0000 27/299
134/2122
0.0000
30