Title
カニクイザルの精巣毒性モデル構築及び精巣毒性に関連す
るmiRNAに関する研究( 本文(Fulltext) )
Author(s)
櫻井, 健
Report No.(Doctoral
Degree)
博士(獣医学) 乙第148号
Issue Date
2016-09-26
Type
博士論文
Version
ETD
URL
http://hdl.handle.net/20.500.12099/55525
※この資料の著作権は、各資料の著者・学協会・出版社等に帰属します。カニクイザルの精巣毒性モデル構築及び精巣毒性に関連する
miRNA
に関する研究
2016 年
岐阜大学大学院連合獣医学研究科
目次
緒言 ... 1 第1 章 次世代シークエンスによるカニクイザル精巣 miRNA の解析 ... 6 1. 序論 ... 6 2. 実験材料及び実験方法 ... 7 3. 実験結果 ... 9 4. 考察 ... 10 5. 小括 ... 12 第2 章 カニクイザルにおける EGME 誘発精巣毒性モデルの構築及び miRNA プロファイ ルの評価 ... 15 1. 序論 ... 15 2. 実験材料及び実験方法 ... 16 3. 実験結果 ... 21 4. 考察 ... 23 5. 小括 ... 26 第3 章 カニクイザルの精巣温浴障害モデルにおける miRNA プロファイルの評価 ... 32 1. 序論 ... 32 2. 実験材料及び実験方法 ... 33 3. 実験結果 ... 37 4. 考察 ... 39 5. 小括 ... 42 総括 ... 50 謝辞 ... 52 文献 ... 53略語一覧
本文及び図表で用いた略語は以下の通りである。 1,3-DNB: 1,3-dinitrobenzene
A/G: albumin/globulin ratio
ALB: albumin
ALP: alkaline phosphatase
ALT: alanine aminotransferase AST: aspartate aminotransferase BCL-2: B-cell leukemia/lymphoma 2 bp: base pair BTB: blood-testis barrier Ca: calcium CBZ: carbendazim CK: creatine kinase Cl: chloride CRE: creatinine
D.BIL: direct bilirubin
EGME: ethylene glycol monomethyl ether
GLB: globulin
GLU: glucose
HB: hemoglobin
HE: hematoxylin-Eosin
HT: hematocrit
ICH: International Council on Harmonisation of Technical Requirements for Registration of Pharmaceuticals for Human Use
IP: inorganic phosphorus
K: potassium
KO: knock out
LDH: lactate dehydrogenase
MAA: methoxy acetic acid
MCH: mean corpuscular hemoglobin
MCHC: mean corpuscular hemoglobin concentration
MCV: Mean corpuscular volume
miRNA: microRNA
Na: sodium
PCR: polymerase chain reaction PDCD4: programmed cell death protein 4
PLT: platelet count
PTEN: phosphatase and tensin homolog RBC: red blood cell count
RET: reticulocyte count
rpm: reads per million
RT-qPCR: reverse transcription-quantitative real-time- polymerase chain reaction
SD: standard deviation
T.BIL: total bilirubin T.CHO: total cholesterol
TE: tris-EDTA
TG: triglyceride
T.PRO: total protein
TH treatment: testicular hyperthermia treatment
UN: urea nitrogen
WBC: white blood cell count
緒言
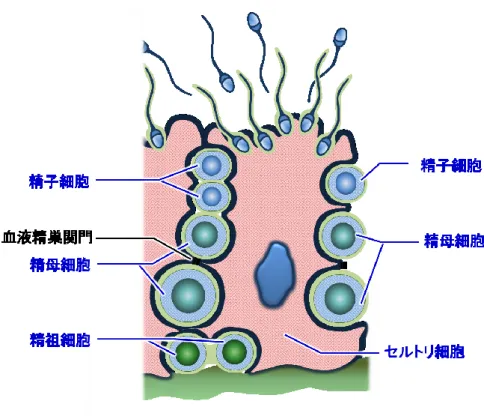
精巣は,雄性生殖機能に必須である精子形成及び性ホルモン産生を担う,左右 1 対の器官 である。精巣の重要な役割の 1 つである精子形成は,個体の性成熟に伴い精細管で開始さ れ,精粗細胞,精母細胞,精子細胞を経て最終的に精子が形成される(Figure 1)。精子成熟 過程において,これら精細胞を支持するセルトリ細胞は,各精細胞への栄養供給とともに, 下垂体からの卵胞刺激ホルモンの刺激を受けてインヒビン分泌をするなど,精子形成に重 要な役割を果たしている。隣接するセルトリ細胞間には,密着結合帯からなる血液精巣関 門(Blood-Testis Barrier:BTB)が形成され,精細胞を基底側と管腔側で区切り,減数分裂 以降の精細胞を血液成分から隔離している。減数分裂により半数体となった精細胞は免疫 学的に非自己と認識される恐れがあるが,血液精巣関門により自己の免疫系から隔絶され ているため免疫応答の影響を受けないことが知られている。精巣の間質に位置するライデ ィッヒ細胞は,下垂体からの黄体刺激ホルモンの刺激を受け,精子形成に必須なテストス テロンを主体として,アンドロステンジオンなど数種の性ホルモンを合成・分泌する。 医薬品開発における精巣毒性は,ヒトで精子形成不全あるいは不妊に直結する重篤な副作 用の一つである。従来の非臨床安全性試験における精巣毒性評価法としては,精巣容量測 定,精液検査,性ホルモン測定,及び病理学的検査などが挙げられる。しかしながら,上 述のように精巣では種々の細胞及び性ホルモンが精子形成および性ホルモン産生において 複雑に関与しており,精巣毒性のメカニズムや対応策を考察することは容易ではない。精 巣毒性評価では,主にラットを中心とした齧歯類が汎用されているが,精子形成(12)及びホ ルモン制御(27)を含めた生理学的な類似性の観点から,サル類がヒト精巣毒性のリスクアセ スメントにおいて最も適切な動物種であることが期待される。サル類の中でも,カニクイ ザルは最も一般的に非臨床安全性試験で用いられる動物種である。カニクイザルの精巣は ヒト同様に体壁から離れた状態で陰嚢内部に位置し,これはヒトを含む霊長類で共通の特 徴である。また,形態学的にカニクイザルの精子頭部はヒトと同様に円形であるが,ラットなどの齧歯類では鎌型を呈するというヒトとの相違点が認められる。精子形成のホルモ
ン制御に関してもラットと比較してカニクイザルがヒトと近いことが示唆されている(27)。
例えば,長期のテストステロン投与による精子形成障害は,ヒトとカニクイザルで類似し
ているが,ラットでは障害細胞の種類が異なることが知られている(27)。また,近年増加傾
向にあるバイオ医薬品開発のための非臨床安全性試験においては,International Council on
Harmonization of Technical Requirements for Registration of Pharmaceuticals For Human Use(ICH)
の「バイオテクノロジー応用医薬品の非臨床における安全性評価に関するガイドライン(S6 ガイドライン)」によって,被験物質が薬理学的活性を示すような動物種で実施することが 明記されており,交差種での毒性試験に重点が置かれている。抗体などのバイオ医薬品を 用いた非臨床安全性試験では,遺伝的にヒトに近縁であるサル類のみに交差性を示す場合 があるため,カニクイザルが汎用されている(7)。このようにカニクイザルの精巣毒性に関す る研究はその必要性が増しているが,その研究報告は齧歯類を用いた研究と比較して非常 に少ないのが現状である。 これまでに,精巣毒性評価のための検査法及びバイオマーカーについて数多くの研究が報 告されているが(11, 22, 31),十分な感度及び特異性を示すバイオマーカーは現在のところ見出 されていない。カニクイザルで使用可能な精巣毒性評価法のうち,精巣容量測定は簡便で あるものの精巣毒性の検出感度が低いことが問題となる。また精液検査は生理学的変動が 大きいため精巣毒性の検出感度が低く,また障害精子が精巣から精液として射出されるま でに時間差が生じることを考慮に入れる必要がある。ホルモン依存的な精巣毒性の評価に は性ホルモンの測定が有用であるが,精細胞の直接障害による精巣毒性では変動が見られ ない場合があるため検出感度が必ずしも高いとは言えない。また種交差性の問題から,カ ニクイザルでの測定はテストステロンなど比較的低分子量のステロイドホルモンを除いて は困難である。病理組織学的検査は精巣毒性評価の最も標準的な検査であるが,屠殺を伴 うため,使用動物数が制限されるカニクイザルを用いた検討では経時的変化の検討は困難 な場合が多い。従って,新規の精巣特異的バイオマーカー及び精巣障害の発生機序を解析
するためのツールの確立が望まれている。 MicroRNA(miRNA)は,20~25 塩基程度のタンパク質をコードしない 1 本鎖 RNA であ り,標的となるmRNA の主に 3’非翻訳領域の一部に相補的に結合し,分解または翻訳を抑 制することによりタンパク質の発現を調節する(3)。精巣においても,miRNA の産生に必須 である Dicer のコンディショナル KO マウスが不妊を示すことから,精子形成においても miRNA が必須であることが示されている(37)。一般に miRNA の塩基配列には種間差が小さ いという特徴を有するが(3, 17),一部のmiRNA にはミスマッチ配列がみられ,また特定の動 物種のみに存在する miRNA もある。また,miRNA の変動は,種々の病態の発生に関連す ることが報告されている(3)。毒性領域においても,臓器特異的に発現しているmiRNA が臓 器障害時に漏出し血中や尿中などで検出される事例が報告されていることから,臓器障害 マーカーとしての有用性が期待されている。例えば,急性肝毒性の際に血中で上昇がみら
れるmiR-122 及び miR-192 や(40, 46),心毒性の際に血中で上昇がみられるmiR-1,miR-208,
及びmiR-499 などがその例である(35)。さらに,miRNA は細胞の分化・増殖,免疫,アポト
ーシスなどホメオスタシスの制御に大きく関わっていることが知られている。miR-21 は, programmed cell death protein 4(PDCD4),phosphatase and tensin homolog(PTEN)などの多 くの腫瘍抑制因子を負に制御することが知られており,このことから腫瘍発生に関与する 腫瘍関連miRNA として認識されている(1, 28)。このように特定の役割を有するmiRNA の変 動を捕捉することにより,臓器毒性の発生機序を解析する上で一助となる。以上から, miRNA はバイオマーカーとして,また毒性発生機序の解析ツールとして有用と考えられる が,毒性領域におけるカニクイザルのmiRNA に関する報告は著者の調べる限り皆無である。 本研究では,カニクイザルの精巣毒性及びmiRNA に焦点をあて,薬剤誘発性精巣毒性モデ ルを構築するとともに,精巣毒性に関連するmiRNA を探索した。第 1 章では,塩基配列を 直接解析可能な次世代シークエンス法を用いてカニクイザルの主要臓器ごとにmiRNA シー クエンスを実施し,カニクイザルの miRNA 塩基配列を確認するとともに,精巣特異的
miRNA を検索した。第 2 章では,代表的精巣毒性物質であるエチレングリコールモノメチ ルエーテル(ethylene glycol monomethyl ether:EGME)を用いて薬物誘発性精巣障害モデル
を確立するとともに,精巣障害時に変動する精巣及び血漿中miRNA について検討した。第
3 章では,精巣温浴障害モデルを用いて精巣障害時に変動する miRNA を検討した。総括で
は,第 1 章から第 3 章までの結果に基づいて,カニクイザルを用いた精巣毒性評価及び精
第
1 章 次世代シークエンスによるカニクイザル精巣 miRNA の解析
1. 序論 近年,miRNA 研究が急速に進展し,悪性腫瘍をはじめとした様々な疾患に関与している 可能性が示唆され,これらmiRNA が疾患バイオマーカー候補として注目されている(48)。ま た,臓器特異的miRNA は臓器障害マーカーとなり得ることが報告されており,毒性分野へ の応用も期待されている。例えば,肝臓に高発現がみられるmiR-122 及び miR-192 はアセ トアミノフェン誘発肝毒性発症時に血中濃度が上昇することや(40, 46),心筋に比較的高発現 することが知られている miR-1,miR-208,miR-499 は急性心毒性モデルにおいて血中レベ ルが上昇することが報告されている(35, 43)。精巣においては,ラットでmiR-34 や miR-449 が 高発現し,かつ臓器特異性が高いことが報告されているが,カニクイザルにおける発現パ ターンについては過去に解析されていない。さらに,ヒトやラットを含めた数種の動物で はmiRNA のシークエンス解析が進展し(30, 47),膨大なmiRNA の塩基配列が決定され公開さ れているが(20),カニクイザルで塩基配列が明らかとなっているmiRNA は限定されている。miRNA の解析方法としては,1 種の miRNA に対して実施する reverse transcription- quantitative real-time-polymerase chain reaction(RT-qPCR)法と多種の miRNA を同時に解析可
能なマイクロアレイが汎用されている。しかし,これらの方法では既知のmiRNA 配列に対 して相補的なプライマーあるいはプローブを設計することが必要であり,塩基配列が未知 のmiRNA ではその発現量を確認することができない。一方で,シークエンス解析では未知 の塩基配列を有するmiRNA を解析することができるが,これまでの技術では上述した 2 法 と比較して検査に費やす時間が非常に長いことが欠点であった。しかしながら近年では, 次世代シークエンス技術の進歩により,シークエンス解析の速度が劇的に向上した。例え ば,旧法であるキャピラリー電気泳動のサンガーシークエンス法では,ヒト全ゲノム解析 には約13 年を費やしたが,次世代シークエンス法では約 10 日間での解析が可能となった。
結果として,未知の塩基配列決定やmiRNA の発現量解析にも次世代シークエンス法が応用 され,報告数は年々増加している。次世代シークエンス法では塩基配列を直接読み取るた め,ほとんどのmiRNA の塩基配列が不明であるカニクイザルの miRNA 解析には適した方 法と考えられる。また,次世代シークエンス解析によりmiRNA の塩基配列を一度決定すれ ば,その後はRT-qPCR やマイクロアレイを使用することで解析がより効率的になると考え られる。 そこで本章では,健常カニクイザルの精巣を含めた主要7 臓器から miRNA を含めた total RNA を抽出し,次世代シークエンス法により網羅的にカニクイザル miRNA の塩基配列情 報を取得するとともに,カニクイザルの精巣特異的なmiRNA について特定を試みた。 2. 実験材料及び実験方法 本実験は,第一三共株式会社の動物実験委員会の承認下(動物実験承認番号:A1200207) で実施した。 2. 動物 2.1. 動物 健常な雌雄各2 例,計 4 例のカニクイザル(中国産,雄 2 例,雌 1 例,8-11 歳齢,体重 2.8-6.1 kg;インドネシア産,雌 1 例,5 歳齢,体重 3.2 kg)を用いた。動物は,温度 24ºC, 湿度60%,照明 12 時間/日(7:00-19:00),照度 150-300 ルクス,換気回数 10-15 回/ 時間に設定した試験室にて,ステンレス製ケージ内に個別飼育した。給餌は,カニクイザ ル用固形飼料(PS,オリエンタル酵母工業株式会社,東京)を 1 日 1 回約 100 g 与えた。給 水は,水道水を給水ノズルから自由に摂取させた。 2.2. 臓器の採材 4 例のカニクイザルに,20 mg/kg の塩酸ケタミン(ケタラール®筋注用500 mg,第一三共
株式会社,東京)及び0.5 mg/kg のキシラジン塩酸塩(セラクタール®,バイエルメディカ ル株式会社,大阪)を筋肉内投与して全身麻酔を施し,頸動脈からの放血により安楽死し た。安楽死後は速やかに心臓(左心室壁),肺(左肺後葉),小腸(空腸及び回腸),肝臓(外 側左葉),脾臓,腎臓(皮質),及び精巣(実質のみ)の各臓器・器官を1 臓器あたり 50- 100 mg で 2 検体ずつ分取して,速やかに液体窒素で急速凍結し,RNA 抽出まで -80ºC で保 管した。 2.3. RNA 抽出
臓器サンプルは,miRNeasy Mini Kit(QIAGEN, Venlo, Netherlands)を用いて,定法に従い
small RNA を含む total RNA を抽出した。抽出した RNA の品質は,Agilent 2100 バイオアナ
ライザ(Agilent Technologies, Santa Clara, CA, USA)を用いて確認した。各臓器について,4
例から採材したtotal RNA を等量混合して,解析用の total RNA とした。精巣については雄
2 例から採材した total RNA を等量混合し,解析用の total RNA とした。これら total RNA は 送付時まで -80ºC で保管した。
2.4. 次世代シークエンス解析 2.4.1. 塩基配列及び発現情報の取得
2.3. 項により調製した total RNA を北海道システム・サイエンス株式会社に送付し,下記
の概要にて次世代シークエンス解析を実施した。RNA の品質は,分光光度計(Nanodrop,
Thermo Fisher Scientific Inc., Waltham, MA, USA)による RNA 濃度測定,並びに Agilent 2100
バイオアナライザ(Agilent Technologies)による RIN(RNA Integrity Number)及びリボソー
ムRNA 比(28S/18S)の測定により確認した。各臓器より採材した 1 μg の Total RNA から,
TruSeq small RNA Sample Prep Kit(Illumina Inc., San Diego, CA, USA)を用いてシークエンス
ライブラリを調製した。次世代シークエンス解析は,HiSeq(Illumina Inc.)を用いて,100 base
ースコールを行い,蛍光純度によってフィルタリングした。さらに,リード配列に含まれ るアダプター配列を除去し,解析用リードデータとした。 2.4.2. データ解析 リードデータの解析は北海道システム・サイエンス株式会社及び第一三共株式会社トラ ンスレーショナルメディシン部で実施した。臓器ごとに同一配列の出現回数を集計しリー ドカウントとして算出した。集計した配列データごとに,miRBase
(
http://www.mirbase.org/
)
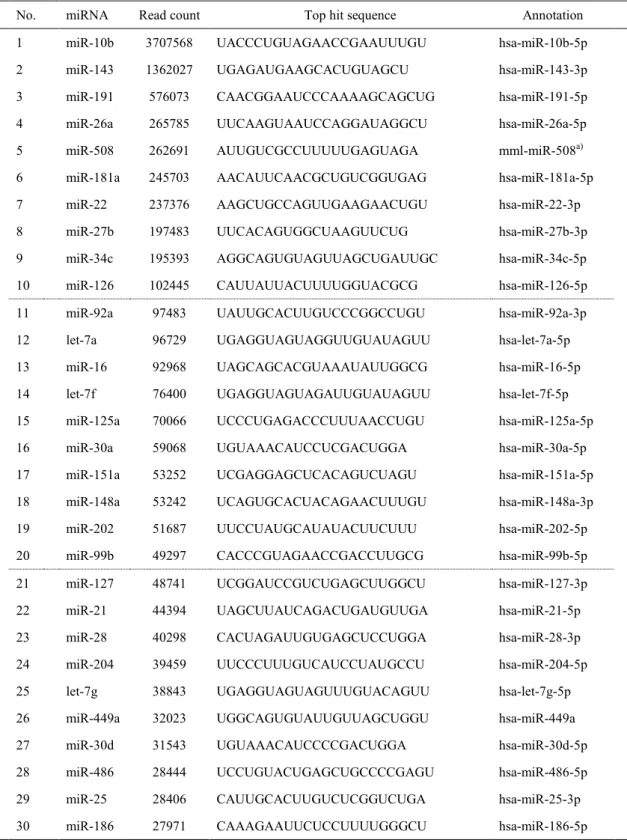
に登録された既知配列に対してBLASTN 解析を実施し,アノテーションを付与した。カウ ントデータは,-3p 及び-5p の合計を miRNA としてカウントし,精巣に由来する上位 30 種の成熟miRNA を抽出した。上位 30 種の成熟 miRNA の塩基配列は、miRBase に登録されて
いるヒトの相同配列と比較した。また,臓器ごとに総リードカウント数で標準化した reads
per million(rpm)を算出し,miRNA の臓器発現を比較した。精巣特異的 miRNA について
は,精巣で 500 rpm 以上の発現を示し,かつ精巣中での発現割合が全臓器の合計に対して 90%以上を占める miRNA を精巣特異的 miRNA として選別した。 2.5. 統計学的解析 有意差検定は実施しなかった。 3. 実験結果 3.1. 次世代シークエンス解析
次世代シークエンス解析により約5 万の small RNA 断片を検出した。BLASTN 解析の結
果,約1000 の成熟 miRNA を検出した。精巣に由来するリードカウント数上位 30 種の成熟
miRNA を Table 1 に示した。また,精巣を含む全 7 臓器の総 rpm に対する精巣 rpm の比率
巣発現比率は,miR-34c で 91.1%,miR-202 で 99.2%,miR-449a で 94.1%,及び miR-508 で 98.2%であった。またそれぞれの miRNA のうち最もリードカウント数が高い配列は, miR-34c-5p,miR-202-5p,miR-449a,及び miR-508-3p であった。一方で,miR-34c 及び miR-449a
は,肺での発現比率が全7 臓器の合計 rpm に対してそれぞれ 8.0%及び 5.5%と,軽微な発現 が認められた。ヒトの配列に対する相同性を確認した結果,miR-508 以外についてはヒトの 配列に対し100%の相同性が認められた。カニクイザルの miR-508 の塩基配列については, ヒトの塩基配列に対し 2 塩基のミスマッチがみられたのに対し,アカゲザルの塩基配列に 対しては100%一致していた。なお,5’又は 3’末端に±2 塩基の配列差異が認められ,isomiR と考えられる配列も各miRNA に認められた。 4. 考察 本章では,成熟カニクイザルの精巣を含めた主要 7 臓器について,次世代シークエンス 技術を用いて網羅的miRNA 発現解析を実施し,約 1000 の成熟 miRNA を同定することに成 功した。精巣において発現量が上位であった30 種の miRNA を集計した結果,ヒトの精巣 でも高発現していることが報告されているlet-7a,let-7f,miR-34c,miR-202,及び miR-508 については,カニクイザルの精巣でも同様に高発現していることが確認された(47)。これら 上位30 種の miRNA の塩基配列のうち,miR-508 ではヒトの相同配列と 2 塩基のミスマッチ が認められたものの,他の29 種の miRNA はヒトの配列と完全に一致しており,カニクイ ザルとヒトの間で塩基配列の相同性が極めて高いことが示唆された。この結果から,一部 の配列についてはミスマッチ配列に注意して評価する必要があるものの,カニクイザルの miRNA 解析にはヒト用のマイクロアレイが応用可能であると考えられた。またヒトとのミ スマッチが認められたmiRNA については,今回明らかになった配列を参考に RT-qPCR の プライマーを作成することで,個別に発現検討することが可能となると考えられた。なお, miRNA の 5’又は 3’末端に±2 塩基の配列差異が認められる isomiR(32)が本実験においても
確認されたが,この配列の多様性の意義は不明であった。
次に,精巣及び他の主要 6 臓器の miRNA 発現を比較した結果,miR-34c,miR-202,
miR-449a,及び miR-508 が精巣特異的に高発現することが確認された。さらに前述した miRNA の中で,最もカウント数が高い塩基配列を選別した結果,miR-34c-5p,miR-202-5p, miR-449a,及び miR-508-3p の 4 種が,カニクイザルの精巣特異的な miRNA であることを
確認した。miR-34c-5p 及び miR-449a はマウスにおいてアポトーシス及び細胞増殖に関与す
ること(9, 23, 25, 44),また両miRNA の double KO マウスでは精子形成不全により不妊となるこ
とが報告されている(44)。このことから,これらのmiRNA が精子形成に重要な役割を果たし
ており,精巣毒性モデルにおけるバイオマーカー候補として有用となるだけではなく,薬
物による精巣毒性メカニズムを解析するためのマーカーとしても有用となる可能性が示唆
された。一方で,miR-34c 及び miR-449a は肺においても軽度な発現が認められた。両 miRNA
のdouble KO マウスでは,気管から肺で正常と比較して繊毛が極端に短いことが報告されて おり,両miRNA はカニクイザルにおいても繊毛形成に関与すると考えられる(44)。miR-202-5p の精巣における役割についてはこれまでのところ報告されていないが,1,3-ジニトロベンゼ ン(1,3-DNB)又はカルベンダジム(CBZ)投与による精巣毒性惹起ラットにおいて血中 miR-202-5p が上昇することが報告されていることから(11),精巣毒性を評価する上での血中 マーカー候補となる可能性が示唆された。また,miR-508-3p の生理学的な機能についても これまでに報告されていないが,4 種の精巣特異的 miRNA の中で精巣での発現量が最も高 かったことを鑑みると,有望なバイオマーカー候補となる可能性が示唆された。これら 4 種のmiRNA は,いずれも精巣特異性が高く,かつヒト精巣中でも高発現することが確認さ れていることから(47),ヒトにおける精巣毒性を推測する上で有用なトランスレーショナル バイオマーカーとなる可能性が示唆された。しかしながら前述の結果は,いずれも正常動 物における発現解析の結果であり,今後バイオマーカーならびに精巣毒性メカニズムの解 析ツールとして有用となるかを確認するためには,精巣毒性を惹起した動物における発現 の変動を確認する必要があると考えられた。
5. 小括 カニクイザルのmiRNA の塩基配列及び精巣特異的 miRNA を特定するため,精巣を含め た主要7 臓器の次世代シークエンス解析を実施した。その結果,カニクイザルの精巣 miRNA の塩基配列を初めて明らかにした。カニクイザルmiRNA の塩基配列は一部の配列を除いて ヒトmiRNA の塩基配列と一致していたことから,カニクイザルの miRNA 解析にはヒトの miRNA 解析プラットフォームを利用可能であると考えられた。また,カニクイザルの精巣
特異的miRNA として,miR-34c-5p,miR-202-5p,miR-449a,及び miR-508-3p を見出した。
これらのmiRNA はヒト精巣においても高発現が認められ,精巣機能に重要な役割を果たし
ていると考えられる。以上から,本研究で選抜した精巣特異的miRNA は,カニクイザルの
精巣毒性バイオマーカー及びヒトへのトランスレーショナルマーカーとして有力な候補と
TABLE 1. Top 30 miRNAs in the testis of cynomolgus monkeys as determined by next generation
sequencing
No. miRNA Read count Top hit sequence Annotation 1 miR-10b 3707568 UACCCUGUAGAACCGAAUUUGU hsa-miR-10b-5p 2 miR-143 1362027 UGAGAUGAAGCACUGUAGCU hsa-miR-143-3p 3 miR-191 576073 CAACGGAAUCCCAAAAGCAGCUG hsa-miR-191-5p 4 miR-26a 265785 UUCAAGUAAUCCAGGAUAGGCU hsa-miR-26a-5p 5 miR-508 262691 AUUGUCGCCUUUUUGAGUAGA mml-miR-508a)
6 miR-181a 245703 AACAUUCAACGCUGUCGGUGAG hsa-miR-181a-5p 7 miR-22 237376 AAGCUGCCAGUUGAAGAACUGU hsa-miR-22-3p 8 miR-27b 197483 UUCACAGUGGCUAAGUUCUG hsa-miR-27b-3p 9 miR-34c 195393 AGGCAGUGUAGUUAGCUGAUUGC hsa-miR-34c-5p 10 miR-126 102445 CAUUAUUACUUUUGGUACGCG hsa-miR-126-5p 11 miR-92a 97483 UAUUGCACUUGUCCCGGCCUGU hsa-miR-92a-3p 12 let-7a 96729 UGAGGUAGUAGGUUGUAUAGUU hsa-let-7a-5p 13 miR-16 92968 UAGCAGCACGUAAAUAUUGGCG hsa-miR-16-5p 14 let-7f 76400 UGAGGUAGUAGAUUGUAUAGUU hsa-let-7f-5p 15 miR-125a 70066 UCCCUGAGACCCUUUAACCUGU hsa-miR-125a-5p 16 miR-30a 59068 UGUAAACAUCCUCGACUGGA hsa-miR-30a-5p 17 miR-151a 53252 UCGAGGAGCUCACAGUCUAGU hsa-miR-151a-5p 18 miR-148a 53242 UCAGUGCACUACAGAACUUUGU hsa-miR-148a-3p 19 miR-202 51687 UUCCUAUGCAUAUACUUCUUU hsa-miR-202-5p 20 miR-99b 49297 CACCCGUAGAACCGACCUUGCG hsa-miR-99b-5p 21 miR-127 48741 UCGGAUCCGUCUGAGCUUGGCU hsa-miR-127-3p 22 miR-21 44394 UAGCUUAUCAGACUGAUGUUGA hsa-miR-21-5p 23 miR-28 40298 CACUAGAUUGUGAGCUCCUGGA hsa-miR-28-3p 24 miR-204 39459 UUCCCUUUGUCAUCCUAUGCCU hsa-miR-204-5p 25 let-7g 38843 UGAGGUAGUAGUUUGUACAGUU hsa-let-7g-5p 26 miR-449a 32023 UGGCAGUGUAUUGUUAGCUGGU hsa-miR-449a 27 miR-30d 31543 UGUAAACAUCCCCGACUGGA hsa-miR-30d-5p 28 miR-486 28444 UCCUGUACUGAGCUGCCCCGAGU hsa-miR-486-5p 29 miR-25 28406 CAUUGCACUUGUCUCGGUCUGA hsa-miR-25-3p 30 miR-186 27971 CAAAGAAUUCUCCUUUUGGGCU hsa-miR-186-5p
Data of read count was obtained from a pooled sample of 2 mature male animals. a) mml-miR-508 is a previous name of mml-miR-508-3p.
FIGURE 2. Tissue specificity of several miRNAs in cynomolgus monkeys.
(A) miR-508-3p, (B) miR-34c-5p, (C) miR-202-5p, and (D) miR-449a. Y-axis represents reads per million. Liver: Left lateral lobe, Heart: Left ventricular wall, Lung: Left caudal lobe, Kidney: Cortex, Small intestine: Jejunum and ileum.
第
2 章 カニクイザルにおける EGME 誘発精巣毒性モデルの構築及び miRNA
プロファイルの評価
1. 序論 ラットにおいては,カドミウム,1.3.-DNB,ドキソルビシン等,数多くの化学物質及び薬 剤による精巣毒性モデルが報告されている。一方でカニクイザルを用いた薬剤誘発性精巣 毒性モデルについては,これまでのところ報告が無いのが現状である。エチレングリコールモノメチルエーテル(ethylene glycol monomethyl ether:EGME)は, 印刷インキ及び塗料の溶剤,並びにジェット燃料の氷結防止等として今日でも幅広く使用
されている工業用品の1 つである(2, 19)。一方でEGME は,マウス(33),ラット(14),ハムスタ
ー(34),モルモット(21),ウサギ(29),及びヒト(19)等,多種の哺乳動物に対して単回あるいは 4
日間反復など比較的短期間の経口投与で精巣毒性を惹起することが報告されており,精巣
毒性を評価するためのモデル化合物として利用されている。EGME による精巣毒性は,そ
の代謝物であるメトキシ酢酸(methoxy acetic acid:MAA)がパキテン期精母細胞を標的と
することによるもので,ホルモン非依存的な障害であることが報告されている(15)。また EGME による精巣毒性発生機序として,酸化ストレスによるアポトーシス誘導が関与して いることも報告されているが(2, 16, 21),詳細なメカニズムはこれまでのところ不明である。ま た,精巣以外にもリンパ造血器抑制を惹起することが知られているが(19),同様に代表的な ホルモン非依存的精巣毒性物質であるドキソルビシンやシクロフォスファミド等の細胞障 害性抗癌剤と比較すると,他臓器への影響は軽度である。これらの報告から,EGME はカ ニクイザルにおいても全身臓器への影響が軽度でありながら,確実に精巣毒性を惹起する モデル化合物であることが期待される。 福島らは,ラットにEGME を投与した精巣毒性モデルにおいて,精巣中 miR-449a,miR-92a,
miR-320,miR-134,及び miR-188 などの miRNA の発現量が変動していたことを報告した(16)。
の標的であるB-cell leukemia/lymphoma 2(BCL-2)は精巣中で発現上昇することが確認され た。BCL-2 はアポトーシス抑制を促すことから,障害を受けた精細管では新たな精細胞の 産生を促すため BCL-2 が活性化している可能性が考えられている。これらの報告から, miRNA が精巣毒性の発現機序解析におけるツール及びバイオマーカーとして有用となる可 能性が示されているが,カニクイザルの精巣毒性に関連するmiRNA の報告はこれまでのと ころ皆無である。 第 1 章では,カニクイザルの精巣を含めた主要臓器を検体として次世代シークエンス解 析を行い,精巣特異性が高い 4 種の miRNA(miR-34c-5p,miR-202-5p,miR-449a,及び miR-508-3p)を選別した。これらは,カニクイザルにおいて精巣毒性を検出するバイオマー カーとしての可能性が期待されるが,実際にカニクイザルに精巣毒性を惹起した際の挙動 は不明である。そこで本研究では,非臨床安全性試験に一般的に用いられるカニクイザル を用いて,EGME 誘発精巣毒性モデルの確立を試みた。さらに、精巣毒性に伴い変動する miRNA を見出すことを目的とし,EGME をカニクイザルに反復経口投与した後に,精巣及 び血漿中miRNA をマイクロアレイ及び RT-qPCR を用いて網羅的に解析した。 2. 実験材料及び実験方法 本実験は,第一三共株式会社の動物実験委員会の承認下で(動物実験承認番号: A1200207 及びA1201239)実施した。 2.1. 動物 健常な成熟雄性カニクイザル6 例(中国産,3 例;インドネシア産,3 例,8-10 歳齢,体 重4.3-6.9 kg)を用いた。動物は,温度 24ºC,湿度 60%,照明 12 時間/日(7:00-19:00), 照度150-300 ルクス,換気回数 10-20 回/時間に設定した試験室にて,ステンレス製ケ ージ内で個別飼育した。給餌は,カニクイザル用固形飼料(PS;オリエンタル酵母工業株
式会社,東京)を1 日 1 回約 100 g 与えた。給水は,水道水を給水ノズルから自由に摂取さ せた。 2.2. EGME 誘発精巣毒性モデルの作成および miRNA 測定 EGME(2-メトキシエタノール,和光純薬工業株式会社,大阪)を注射用水(株式会社大 塚製薬,徳島)で希釈して60 mg/mL の EGME 溶液を調製し,雄 3 例にディスポーザブル シリンジ(テルモシリンジ,テルモ株式会社,東京)及びカテーテル(SF-SE1435R,テル モ株式会社)を用いて5 mL/kg の容量で 1 日 1 回,4 日間反復強制経口投与した。初回投与 日をDay 1 と規定した。なお EGME の投与用量については,前試験として成熟雄性カニク イザル2 例を用いた用量設定試験(未発表)を行ったところ,最大耐量が 300 mg/kg であっ たことからこの用量に設定した。
EGME 投与の全身臓器への影響を確認するため,投与前(Day -7)及び Day 5 に末梢血約 3.1 mL を大腿静脈から採血し,完全血球計算,血液化学検査,及び血中テストステロン濃 度の測定を実施した。また,精巣毒性に関連する血漿中miRNA を検討する目的で,Day 5 に末梢血1.2 mL を採血して miRNA 測定に供した。さらに,Day 5 に実施した採血の後に 25.9 mg/kg のペントバルビタールナトリウム(ソムノペンチル注®,共立製薬株式会社,東京) を静脈内投与して全身麻酔を施し,麻酔下で頸動脈及び腋窩動静脈からの放血により安楽 死した。その後,左右精巣を採材してそれぞれ病理組織学的検査及びmiRNA 測定に使用し た。安楽死までの間は,動物の一般状態を毎日観察した。なお,残りの 3 例は媒体対照群 として,EGME 溶液の代わりに注射用水を投与し,同様の解析を実施した。 2.3. 完全血球計算及び血液化学検査 EGME 投与による全身への影響を確認する目的で,2.2.項で採取した末梢血のうち 1 mL を, EDTA-2 カリウム(和光純薬工業株式会社)15%溶液が 10 μL 添加された採血容器(アシス トツーポジションチューブ,ザルスタット株式会社,東京)に注入した後,自動血球計数
装置(ADVIA2120i,シーメンスヘルスケア・ダイアグノスティクス株式会社,東京)を用
いて完全血球計算を実施した。また,同様に2.2.項で採取した末梢血のうち 0.6 mL を,ヘ
パリンリチウムがコーティングされたマイクロテイナ®(Becton, Dickinson and Company,
Franklin Lakes, NJ, USA)に注入し,遠心分離(3,000 回転/分,10 分間,4ºC)して血漿を分
離した後に,自動分析装置(TBA-2000FR,東芝メディカル株式会社,栃木)を用いて血液 化学検査を実施した。項目を以下に示す。 ・完全血球計算: RBC,HB,HT,MCH,MCV,MCHC,RET,WBC,WBC Diff,PLT ・血液化学検査: AST,ALT,ALP,LDH,CK,T.BIL,D.BIL,T.CHO,TG,GLU,T.PRO, ALB,GLB,A/G,UN,CRE,Ca,IP,Na,K,Cl 2.4. 血中テストステロン濃度の測定 2.2.項で採取した末梢血のうち 1.5 mL を,ヘパリンリチウムがコーティングされたマイク
ロテイナ®(Becton, Dickinson and Company)に注入し,遠心分離(10,000 回転/分,5 分間,
4ºC)して血漿を分離し,測定まで -80°C で保存した。測定はテストステロン測定キット (DELFIA® Testosterone Reagents,PerkinElmer, Inc., MA, USA)を用いて測定した。
2.5. 病理組織学的検査 左側精巣はホルマリン・ショ糖・酢酸固定液で固定後,パラフィン包埋及びヘマトキシリ ン・エオジン(HE)染色を実施し,光学顕微鏡で観察した。精細胞数の減少については, 重篤度に応じて1+(軽度)~3+(重度)にスコア化して評価を行った。 2.6. miRNA マイクロアレイ解析 精巣毒性により変動する精巣及び血漿中miRNA を調べるため,miRNA マイクロアレイ解 析によりEGME 投与後の精巣及び血漿中 miRNA を網羅的に検索し,対照群と比較した。
2.6.1. 採血及びサンプル処理
2.2.項で採取した末梢血を,EDTA がコーティングされたマイクロテイナ®(Becton,
Dickinson and Company)に注入し,遠心分離(10,000 回転/分,5 分間,4ºC)により血漿 500 µL を分離した後,送付まで -80ºC で保管した。また採取した右精巣の一部(50~100 mg)
を液体窒素で急速凍結し,送付まで -80ºC にて保管した。
2.6.2 Total RNA 抽出
血液サンプルについては,凍結状態のまま三菱化学メディエンス株式会社(現株式会社
LSI メディエンス,鹿島)に送付し,3D-Gene® RNA extraction reagent from liquid sample(東
レ株式会社,東京)を用いてTotal RNA を抽出した。精巣サンプルについては,凍結状態の
まま東レ株式会社に送付し,miRNeasy Mini Kit(QIAGEN)を用いて Total RNA を抽出した。
抽出したRNA の品質については,Agilent 2100 バイオアナライザ(Agilent Technologies)を
用いて確認した。抽出サンプルについては -80ºC で保管し,その後東レ株式会社にて miRNA
マイクロアレイ解析に供した。
2.6.3 miRNA マイクロアレイ解析
抽出した血漿及び精巣由来の RNA サンプルを,miRCURY LNA Array miR labeling kit
(Exiqon A/S, Vedbaek, Denmark)を用いて Hy5 でラベリングした後,3D-Gene® Human miRNA
Oligo chip(東レ株式会社)にハイブリダイズし,3D-Gene® Scanner(東レ株式会社)で検出
した。3D-Gene® Human miRNA Oligo chip 上のプローブのアノテーション及びシークエンス
はmiRBase(http://www.mirbase.org/,Version 19.0)に従った。miRNA マイクロアレイ解析
は東レ株式会社で実施した。
miRNA のうち,EGME 投与群で対照群よりも発現量が上昇(> 1.50 倍)又は低下(< 0.66
倍)し,かつ全例でpresent コールを示した miRNA を,変動 miRNA として選別した。さら
> 4.0 倍の上昇又は< 0.50 倍の低下を示し,かつ signal intensity が対照群の全例で 100 を超え ているmiRNA を選別した。 2.7. 定量的 PCR(RT-qPCR)解析 miRNA マイクロアレイ解析によって発現変動がみられた miRNA については,RT-qPCR によりmiRNA の発現を確認した。 2.7.1. Total RNA 抽出
精巣10 mg あたり 700 µL の QIAzol Lysis Reagent(QIAGEN)を加え,ホモジナイザー
(Heidolph DIAX 900,Sigma-Aldrich St. Louis, MO, USA)で約 1 分間ホモジナイズした後に
室温で5 分間静置した。次いで,ホモジネート 500 µL を 1.5 mL マイクロチューブに分取し,
5 µL の外部スパイク溶液及び 100 µL のクロロホルム(和光純薬工業株式会社)を添加し,
ボルテックスミキサーで15 秒間,さらに激しく 15 秒間転倒混和した後,室温で 3 分間静
置した。サンプルを遠心分離(12,000 回転/分,15 分間,4ºC)した後,水層(上層)をピ
ペットで採取し,チューブに移した。次いで採取した水層の1.5 容の 100% ethanol(和光純
薬工業株式会社)を添加・混合し,RNeasy Mini Spin Column(QIAGEN)により total RNA
を回収し,使用時まで -80ºC で保管した。
2.7.2. 逆転写反応
TaqMan MicroRNA Reverse Transcription Kit(Thermo Fisher Scientific Inc.)を用いて逆転写
反応を行った。すなわち,5 µL の total RNA サンプルに 10 µL の RT master mix 溶液(0.15 µL
の100 mM dNTP,1 µL の 50 U/µL MµltiScribe Reverse Transcriptase,1.5 µL の 10x Reverse Transcription Buffer,0.19 µL の 20 U/L RNase inhibitor,3 µL の RT primer mix[0.75 µL/miRNA]:, 4.16 µL の Nuclease-free water)を混和して氷上で 5 分間静置した後,サーマルサイクラー (GeneAmp PCR System 9700,Thermo Fisher Scientific Inc.)を用いて 16ºC で 30 分間,42ºC
で30 分間,85ºC で 5 分間反応させ,cDNA サンプルを作製した。cDNA サンプルは定量的 PCR に使用するまで -80ºC で保管した。
2.7.3. RT-qPCR による精巣中 miRNA 量測定
2 µL の 5 倍希釈の cDNA サンプル又は検量線用の cDNA サンプルを,TaqMan Fast Advanced MasterMix,TaqMan MicroRNA Assay 及び Applied Biosystems 7900HT Fast Real Time PCR System(Applied Biosystems)を用いて,miR-34b-5p,miR-449a 及び miR-238(外部標準)の miRNA 発現量を real-time PCR 法により相対定量で測定した。サイクル条件については,50ºC で2 分間,95ºC で 20 秒間反応後,95ºC で 1 秒間,60ºC で 20 秒間を 45 サイクルとした。 miRNA 量については,検量線を用いた相対定量法により定量化した。測定対象となる miRNA の発現量は,外部標準である miR-238 の発現量を用いて補正した。 2.8. 統計学的解析 血液・血液化学パラメータ,血漿中テストステロン濃度,miRNA マイクロアレイ,及び 定量的PCR の各測定値は,測定時点ごとに群別の平均値±標準偏差(SD)で解析した。媒 体対照群とEGME 投与群の平均値の有意差検定は,F 検定により等分散が確認された場合 はStudent の t 検定を,不等分散の場合は Aspin-Welch の t 検定を実施した。有意水準は 5% とした。 3. 実験結果 3.1. 一般状態,体重,及び摂餌量 一般状態の観察では,EGME 投与群の 3 例中 2 例で,吐物が Day 3 の投与前に認められ
た。また,EGME 投与群では Day 3 又は Day 4 以降に全例で摂餌量が低下し,Day 5 では全
-11.6%)減少した。いずれの例においても活動性には異常は認められなかった。対照群で は,一般状態,体重,及び摂餌量ともに投与前後で変化は認められなかった。
3.2. 完全血球計算及び血液化学検査
EGME 投与群では対照群と比較して,リンパ球数及び reticulocyte count(RET)の減少が 認められた。また血液中のalanine aminotransferase(ALT)値,total bilirubin(T.BIL)値,direct bilirubin(D.BIL)値の軽度上昇,ならびに inorganic phosphorus(IP)値の上昇が認められた。 その他の項目には,投薬が起因したと思われる異常は認められなかった。 3.3. 血中テストステロン濃度 結果をFigure 3 に示す。試験期間を通じて EGME 投与群と対照群の間に有意な差はみら れなかった。 3.4. 病理組織学的検査 病理組織学的検査により精巣で認められた代表的な所見をFigure 4 に示す。 EGME 投与群の 3 例では,全例で重度の精母細胞の変性及び細胞数の減少,並びに軽度 から中等度の精細管の空胞変性が認められ,1 例では中等度の円形精子細胞数の減少が認め られた。病理組織学的な重篤度ついては,3 例の間で顕著な差は認められなかった。なお精 細管の空胞変性については,精細管を構成するどの細胞に認められるのかを特定するには 至らなかった。対照群には前述の変化は認められなかった。 3.5. miRNA マイクロアレイ解析 EGME 投与群において,対照群よりも発現量が上昇(>1.50 倍)又は低下(<0.66 倍)し たmiRNA を選別したところ,血漿中には発現上昇した miRNA は認められず,逆に 326 種 のmiRNA で発現低下が認められた。精巣では,347 種の miRNA で発現上昇が認められた
のに対し,16 種の miRNA で発現低下が認められた(Figure 5)。精巣中で発現低下した miRNA
には,精巣特異的なmiRNA である miR-34c-5p 及び miR-449a が含まれていた。また血漿中
で発現低下が認められた326 種の miRNA のうち,138 種の miRNA では精巣において発現
上昇も同時に認めた。さらにEGME 投与群の精巣において発現上昇が顕著(> 4.0 倍)であ
ったmiRNA は 18 種であったのに対し,発現低下が顕著(< 0.50 倍)であった miRNA は 7
種であった(Table 2)。
3.6. RT-qPCR 解析
EGME 投与群の精巣では,miR-34b-5p 及び miR-449a のいずれにおいても,対照群と比較 して発現量が有意に低値を示した(Figure 6)。なお前述の miRNA 発現量については,対照 群及びEGME 群のいずれにおいても個体間における顕著な差は認められなかった。 4. 考察 本章では,カニクイザルを用いた薬剤誘発性精巣毒性モデルを確立した。またカニクイ ザルにおいて精巣毒性に伴って変動するmiRNA を特定するため,代表的な精巣毒性物質で あるEGME を成熟雄性カニクイザルに 4 日間反復経口投与し,血漿中及び精巣中における miRNA の変動をマイクロアレイによる網羅的解析法及び RT-qPCR 法により検討した。 EGME 投与群では,病理組織学的に精母細胞の変性・消失及び円形精子細胞の消失が認 められた。また精細管の空胞化が認められたが,この変化はセルトリ細胞の空胞化による ものと考えられた。本実験でみられた精巣の組織学的変化は,EGME 投与ラットにおける 所見と類似しており(16),EGME はカニクイザルにおいても精巣毒性を惹起することが確認 された。また前述の変化がテストステロンの変動を伴わずに認められたことから,EGME 投与群における精巣毒性はホルモン非依存的な変化であると考えられた。なお,本実験で は摂餌低下に伴い最大約 12%の体重減少が認められたが,一般状態には重篤な変化が認め られなかった。また,血液学的検査の結果,EGME 投与群では軽度な肝障害及びリンパ造
血器抑制を疑わせる所見が認められた。しかしながらこれらの異常は,精巣における異常
と比較すると軽微であり,精巣毒性モデルとして許容できる範囲の異常であると考えられ
た。
本研究で実施されたmiRNA マイクロアレイ解析の結果,EGME の 4 日間反復投与によ
り血漿中及び精巣で計550 種の miRNA に発現量の変動が認められた。精巣では 347 種の
miRNA で発現上昇が,16 種の miRNA で発現低下が認められた。これらの miRNA には,
第1 章で確認された精巣特異的 miRNA のうち,miR-34c-5p 及び miR-449a の 2 種類が含ま
れていた。さらにEGME 投与群の精巣では,18 種の miRNA で発現上昇(>4.0 倍)が,ま
た7 種の miRNA で発現低下(<0.50 倍)が認められた(Table 2)。これらのうち精巣特異性
が高いmiR-449a を含む 2 種の miRNA については,定量的 PCR 解析でも有意な発現低下が
認められた(Figure 6)。miR-449a,miR-34b-5p,及び miR-34c-5p は,miR-34/449 ファミリ
ーに属し,マウスでは精母細胞(減数分裂細胞)に高発現がみられることが知られている(39)。
また,miR-34b-5p 及び miR-34c-5p は異なる miRNA であるが,両 miRNA の塩基配列は 23
塩基中 3 塩基のみのミスマッチであり,役割及び組織分布についても重複していることが
知られている(39)。本試験において,EGME 投与群でパキテン期精母細胞の消失が観察され
たことを考慮すると,miR-449a 及び miR-34b-5p の精巣中での発現低下は減数分裂細胞であ
る精母細胞の障害を反映していることが示唆された。miR-34/449 はアポトーシス・細胞増
殖に関連すること,miR-34b のターゲットは histone deacetylase,sirtuin 1(SIRT1)を介して P53 を活性化すること,あるいは cyclin-dependent kinase 6 や E2F を down regulate し E2F pathway を阻害することで cell cycle arrest を引き起こすことが知られている(5, 41, 45)。miR-449a
もまた,histone deacetylase 1 あるいは SIRT1 を介し,miR-34b と同様に P53 を活性化するこ
とが報告されている(24, 36)。また,福島らはEGME を投与したラット精巣中で miR-449a の低
下及びBCL-2 の発現上昇が認められることを報告している(16)。これらの報告から,本試験
でみられた精巣中におけるmiR-449a 及び miR-34b-5p の発現低下により,P53 の活性化が抑
ある。しかしカニクイザルにおいては,精子形成における前述のシグナル伝達経路につい て未だ不明な点が多く,更なる検討が必要と考えられる。 第1 章で明らかにしたカニクイザルの精巣特異的 miRNA のうち精巣中 miR-202-5p 及び miR-508 については本実験条件下では変動が認められなかった。miR-202-5p については,ヒ ト精巣中ではセルトリ細胞に高発現することが報告されている(10)。今回,EGME 投与によ るカニクイザル精巣毒性モデルでは,セルトリ細胞に局在すると考えられる空胞変性は認 められたものの,精巣miR-202-5p に変動が認められなかったことから,セルトリ細胞の機 能に対する EGME の影響が軽微であった可能性が示唆された。miR-508 については,第 1 章においてヒト相同配列と 2 塩基のミスマッチが存在することが確認されているため,本 実験で用いられたマイクロアレイ解析では正しく検出されなかった可能性が考えられる。 また本研究では,カニクイザルに由来するmiR-508-3p の塩基配列を基にカスタムプライマ ーを設計したものの,品質確認において正常な増幅がみられなかったため,miR-508-3p の 正確な発現量は解析できなかった。この理由については未だ不明であり,今後は多方面か らの検討が必要であると考えられた。 血漿中 miRNA についてマイクロアレイ解析を行った結果,miRNA の発現上昇は認めら れなかったが,326 種の miRNA で発現低下が認められた(Figure 5)。発現変動が認められ たmiRNA は,いずれもその役割が報告されていない miRNA か,あるいはカニクイザルの 精巣において特異的に発現するmiRNA とは異なるものであった。したがって本試験系では, 精巣中で発現変動したmiRNA を,血漿を用いて捕捉することはできなかった。この理由の 一つとして,精巣が他の主要臓器(心臓,肺,肝臓,消化器など)と比較し小器官である ため,総細胞数及び総miRNA 数が少なく,血漿中への寄与率が低かった可能性が考えられ た。
5. 小括 カニクイザルにEGME を 4 日間反復経口投与し,精母細胞の変性・消失及び円形精子細 胞の消失を特徴とする精巣毒性モデルを確立した。同モデルでは血漿中のmiRNA に変動は みられなかったが,精巣においてはmiR-34b-5p 及び miR-449a の発現低下が認め,精母細胞 障害を反映すると考えられた。また前述した2 種の miRNA は,カニクイザルにおける精巣 毒性を示唆するバイオマーカーの候補になると考えられた。
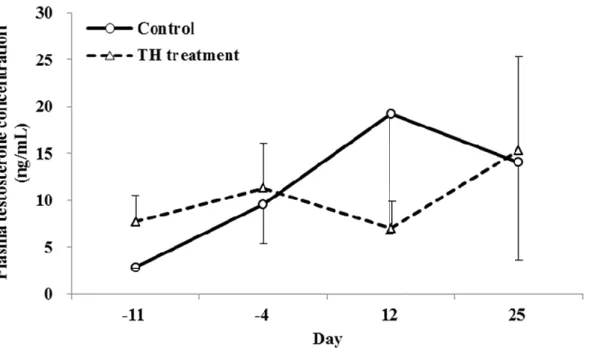
FIGURE 3. Effect of EGME on plasma testosterone levels in cynomolgus monkeys.
Testosterone levels were measured before treatment (Days -7) and on Day 5. Data are shown as the mean ± SD (n = 3 / group).
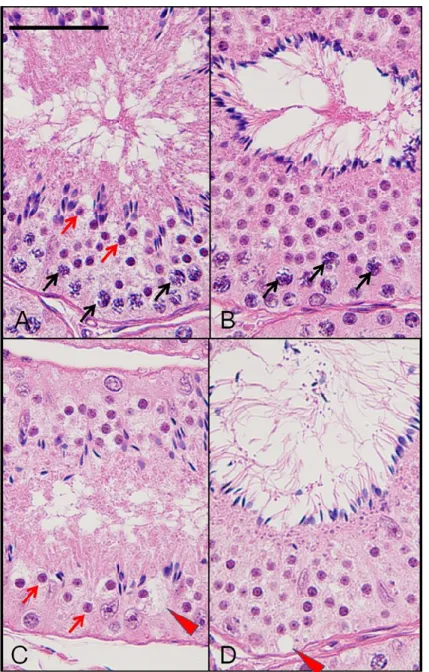
FIGURE 4. Representative histopathological changes in the testis of cynomolgus monkey treated
with vehicle (A and B) or EGME at 300 mg/kg (C and D). All specimens with HE staining represent the seminiferous tubules in different morphological stages. A and C represent stage 1 to 4, while B and D represent stage 5 to 6. EGME treatment decreased the number of pachytene spermatocytes during stage 1 to 6 (C and D) compared with the concurrent control group (A and B). The number of round spermatid decreased during stage 1 to 4 after EGME treatment (C). Vacuolation in
seminiferous tubule (red arrowheads) was also observed in animals with EGME treatment (C and D).
Black and red arrows show morphologically normal pachytene spermatocytes and round spermatids, respectively. Bar indicates 50 m.
FIGURE5. Venn diagrams showing overlapping up- or down- regulated miRNA between plasma
and the testis after EGME treatment. These miRNAs were determined based on the criterion of >1.5 fold for up-regulation or <0.66 fold for down-regulation compared with the concurrent control group.
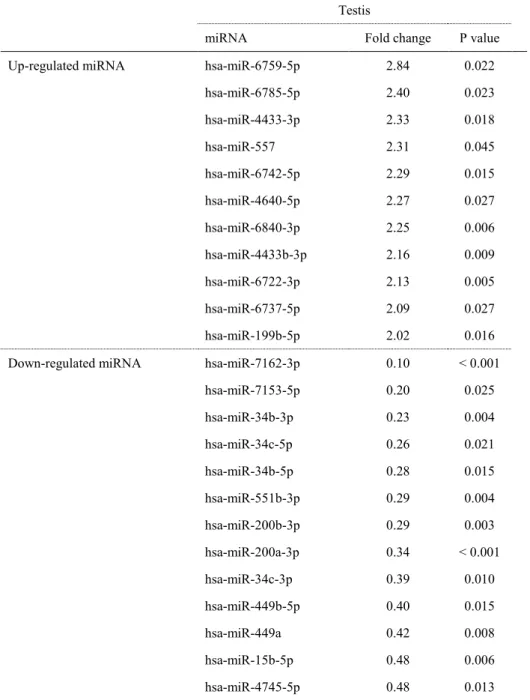
TABLE 2. Effect of EGME on testicular miRNAs in cynomolgus monkeys
Testis
miRNA Fold change P value Up-regulated miRNA hsa-miR-4465 9.82 0.022 hsa-miR-4747-3p 8.40 0.005 hsa-miR-5585-3p 6.70 0.003 hsa-miR-665 6.19 < 0.01 hsa-miR-3619-3p 6.07 0.001 hsa-miR-4257 5.76 0.002 hsa-miR-614 5.43 < 0.01 hsa-miR-658 4.88 0.017 hsa-miR-4739 4.86 < 0.01 hsa-miR-4730 4.55 0.002 hsa-miR-642a-3p 4.51 0.005 hsa-miR-4697-5p 4.48 0.002 hsa-miR-675-5p 4.48 < 0.01 hsa-miR-365a-5p 4.42 0.001 hsa-miR-3196 4.32 0.002 hsa-miR-663a 4.30 0.001 hsa-miR-3178 4.17 0.039 hsa-miR-642b-3p 4.01 0.002 Down-regulated miRNA hsa-miR-5685 0.06 0.049 hsa-miR-34c-3p 0.41 0.044 hsa-miR-449a 0.45 0.0498 hsa-miR-93-5p 0.47 0.058 hsa-miR-425-5p 0.48 0.059 hsa-miR-449b-5p 0.48 0.061 hsa-miR-34b-5p 0.49 0.074 P values were calculated by Student's t-test or Aspin-Welch's t-test.
FIGURE 6. Effect of EGME treatment on miRNA expression in the testis in cynomolgus monkeys
Down-regulations of miR-449a and miR-34b-5p were confirmed by quantitative RT-PCR. The expression level of each miRNA in EGME-treated group expressed relative to that of the control group as 1.00. Data are shown as mean ± SD (n = 3 / group). The asterisk (*) shows the statistical significance (P < 0.05) by t-test.
第
3 章 カニクイザルの精巣温浴障害モデルにおける miRNA プロファイルの評
価
1. 序論 第2 章では,カニクイザルの EGME 誘発精巣毒性モデルを作出し,EGME 投与により精 巣において miR-34b-5p 及び miR-449a の発現が低下することを示した。しかし,前述した miRNA の発現低下は EGME 投与時に限って認められる反応であり,精巣毒性を直接反映す る現象ではない可能性も考えられる。そのため,EGME 投与とは異なる方法でカニクイザ ルに精巣障害を惹起し,miRNA 発現を詳細に検討することで,前述の miRNA が精巣毒性 マーカーとして妥当であるかを検証する必要があると考えた。また,EGME 投与モデルで は血漿中におけるmiRNA の発現上昇が認められなかったが,第 1 章で発見したカニクイザ ルの精巣特異的miRNA を含め,他の手法により作成された精巣毒性モデルにおける血漿中 miRNA の挙動をより詳細に解析する必要があると考えた。 近年,Lue らは,カニクイザルの精巣を 43ºC の温水中に 1 日 1 回 30 分間,6 日間連続で 浸漬することによりパキテン期精母細胞及び円形精子細胞のアポトーシスを特徴とした精 巣障害が惹起されることを報告した(26)。本モデルでは,温浴開始6~8 週後に精液中の精子 がほぼ消失し,その後回復することが知られている(26)。精巣で産生された精子が精巣上体 で貯留された後,射精されるまでに約1~2 週間の時間差を有することから,温浴開始から 約4~6 週目に精巣障害が最も重篤となる可能性が推測される。また,精巣温浴モデルの最 大の利点は,精巣に限局的な障害を誘発することが可能である点にある。薬物誘発性の精 巣毒性モデルでは,EGME で報告されている血液毒性や肝毒性(19),及びニトロベンゼンで 報告されているメトヘモグロビン血症(4)などのように,他の臓器に対する毒性を併発する。 以上のことから精巣温浴モデルは,精巣障害に特異的なバイオマーカーの探索を行う上で 最適なモデルであると考えられる。本章では,精巣障害に関連するmiRNA を発見し,精巣毒性機序を解析するためのツール としてのmiRNA の有用性を検討するため,カニクイザル精巣温浴モデルを用いて精巣中及 び血漿中におけるmiRNA の変動を,miRNA マイクロアレイ解析又は RT-qPCR により検討 した。 2. 実験材料及び実験方法 本実験は,第一三共株式会社の動物実験委員会の承認下で(動物実験承認番号:A1302542) 実施した。 2. 動物 2.1. 動物 健常な成熟雄性カニクイザル6 例(インドネシア産,7-10 歳齢,体重 4.3-6.6 kg)を用 いた。動物は,温度24ºC,湿度 60%,照明 12 時間/日(7:00-19:00),照度 150-300 ルク ス,換気回数10-20 回/時間に設定した試験室にて,ステンレス製ケージ内で個別飼育し た。給餌は,カニクイザル用固形飼料(PS-A,オリエンタル酵母工業株式会社,東京)を 1 日1 回,約 100 g 与えた。精巣温浴実施日にはさらにリンゴ 1/2 個を給餌した。給水は,塩 素添加水を給水ノズルから自由に摂取させた。 2.2. 精巣温浴障害モデルの作出 精巣温浴の方法には,Lue らの報告(26)で用いられた方法に一部修正を加えた方法を用いた。 3 例のカニクイザルに塩酸ケタミン(ケタラール®筋注用 500 mg,第一三共株式会社)10 mg/kg を筋肉内投与した後,43℃に設定した恒温槽の温水中に陰嚢を浸し 30 分間静置した。 対照群の3 例についても同様に塩酸ケタミンを筋肉内投与し,空の恒温槽に陰嚢を 30 分間 静置した。精巣温浴を行った初日をDay 1 と規定し,5 日間連続で温浴を実施した。血漿中
血約3.2 mL を大腿静脈から採血した。また,末梢血の一部は,全身臓器への影響及び性ホ ルモンへの影響を確認する目的で,それぞれ血液化学パラメータ及び血中テストステロン
濃度の測定に供した。さらに,Day 26 に塩酸ケタミン(ケタラール®筋注用500 mg)の 10
mg/kg を筋肉内投与した後に,イソフルラン(Escain®,Mylan N.V.,PA,USA)を濃度 1.0
~1.5%で吸入させて全身麻酔を施し,左側精巣直上の陰嚢を切開して精巣を露出させた。 精管及び精巣動静脈を 2 ヵ所で結紮した後に結紮糸間を切断し,精巣及び精巣上体を無菌 的に採材した。摘出した精巣は速やかに短軸方向に2 分割し,一方を病理組織学的検査に, 他方をmiRNA 解析に使用した。試験期間中,一般状態は毎日観察した。 2.3. 血液化学検査 2.2.項で採取した末梢血のうち 0.6 mL を,ヘパリンリチウムがコーティングされたマイク
ロテイナ®(Becton, Dickinson and Company)に注入し,遠心分離(3,000 回転/分,10 分間,
4ºC)して血漿を分離した後に,自動分析装置(TBA-2000FR,東芝メディカル株式会社) を用いて血液化学検査を実施した。項目を以下に示す。 血液化学検査:AST,ALT,ALP,LDH,CK,T.BIL,D.BIL,T.CHO,TG,GLU,T.PRO, ALB,GLB,A/G,UN,CRE,Ca,IP,Na,K,Cl 2.4. 血中テストステロン濃度の測定 2.2.項で採取した末梢血のうち 1.4 mL を,ヘパリンリチウムがコーティングされたマイク
ロテイナ®(Becton, Dickinson and Company)に注入し,遠心分離(10,000 回転/分,5 分間,
4ºC)して血漿を採取し,測定まで -80℃で保存した。測定にはテストステロン測定キット (DELFIA® Testosterone Reagents;PerkinElmer, Inc.)を用いた。
2.5. 病理組織学的検査
及びHE 染色を実施し,光学顕微鏡で観察した。精細胞数の減少については,重篤度に応じ て1+(軽度)~3+(重度)にスコア化して評価を行った。 2.6. miRNA マイクロアレイ解析 精巣毒性により精巣中で発現量が変動する miRNA を網羅的に検索するため,miRNA マ イクロアレイ解析を実施して対照群と比較した。 2.6.1. Total RNA 抽出 2.2.項で採取した左側精巣の一部(50~100 mg)を速やかに液体窒素で凍結し,total RNA の抽出まで -80℃にて保管した。凍結した精巣サンプルを株式会社 LSI メディエンスに送付
し,miRNeasy Mini Kit(QIAGEN)を用いて Total RNA を抽出した。抽出した RNA の品質
は,Agilent2100 バイオアナライザ(アジレントテクノロジー株式会社)を用いて確認した。
2.6.2. miRNA マイクロアレイ解析
抽出した精巣由来のRNA サンプルを,第 2 章の 2.6.3.項と同様の方法により miRNA マイ
クロアレイ解析を実施した。対照群と比較して温浴群で発現量が上昇(> 2.00 倍)又は低下
(< 0.50 倍)し,かつ present コールを示した miRNA を変動 miRNA として抽出した。
2.7. RT-qPCR 解析 miRNA マイクロアレイ解析によって発現変動がみられた精巣 miRNA について発現確認 を行うと共に,精巣毒性のマーカー候補である血漿中miRNA の有用性について評価を行う ため,RT-qPCR 解析を実施した。 2.7.1. Total RNA 抽出及び逆転写反応 2.2.項で採取した末梢血のうち 1.2 mL を,EDTA-2K 添加マイクロテイナ(Becton,
Dickinson and Company)に注入後に氷冷した。遠心分離機(10,000 回転/分,5 分間,4ºC) を用いて500 µL の血漿サンプルを得た。サンプルは RNA 抽出まで冷凍庫で -80ºC で保存 した。Total RNA の抽出は第 2 章の 2.6.1.項に準じて実施した。また,精巣については,2.6.1. 項で抽出したtotal RNA をそのまま使用した。なお,逆転写反応は,第 2 章の 2.7.2.項に準 じて実施した。 2.7.2. Pre-Amplification
2.7.1.項で得られた 2.5 µL の cDNA に,preAmp reaction mix(12.5 µL の 2x TaqMan PreAmp Master Mix,3.75 µL の PreAmp Primer Pool[10 µL の 20x TaqMan MicroRNA assays(2.6.3.項
に示す各 miRNA)を加え,最終液量が 1000 µL となるよう 1x TE で希釈],6.25 µL の
Nuclease-free water)を添加した。氷上で 5 分間静置した後,サーマルサイクラー(GeneAmp PCR System 9700,Thermo Fisher Scientific Inc.)を用いて,95ºC で 10 分間,55ºC で 2 分間, 72ºC で 2 分間反応後,95ºC で 15 秒間及び 60ºC で 4 分間を 12 サイクル実施して,最後に 100ºC で 10 分間反応させた。サンプルは RT-qPCR に使用するまで -80ºC で保管した。 2.7.3. RT-qPCR による miRNA 量測定 精巣のmiRNA マイクロアレイ解析により温浴群で発現変動が認められた miRNA のうち, 第1章および第2 章の結果を考慮し,miR-449a,miR-34b-5p,及び miR-34c-5p の 3 種につ いてRT-qPCR を実施した。血漿については,上述の 3 種の miRNA のうち、miRNA マイク ロアレイ解析により温浴群で発現量が最も低下したmiR-34c-5p,及び第 1 章で精巣特異的 な発現が確認されたmiR-202-5p に限定して RT-qPCR を実施した。 定量的PCR 解析は,第 2 章 2.6.3.項に準じて実施した。精巣中 miRNA は対照群に対する 相対値,血漿中miRNA は個体ごとの投与前値(Day -11 及び-4)に対する相対値をそれぞれ 算出した。
2.8. 統計学的解析 血液化学パラメータ,血漿中テストステロン濃度,miRNA マイクロアレイデータ,及び RT-qPCR の各測定値は,測定時点ごとに群別の平均値±標準偏差(SD)で示した。対照群 と温浴群間における平均値の有意差検定にはF 検定を用い,等分散の場合は Student の t 検 定を,不等分散の場合はAspin-Welch の t 検定を実施した。有意水準は 5%とした。 3. 実験結果 3. 動物 3.1. 一般状態,体重,及び摂餌量 試験期間中,対照群及び温浴群ともに軟便及び麻酔の影響と考えられる吐物が散見され たが,体重や摂餌量には変化がみられなかった。 3.2. 血液化学検査 試験期間を通じて,血液化学パラメータには顕著な変化が認められなかった。 3.3. 血中テストステロン濃度 結果をFigure 7 に示す。試験期間を通じて温浴群と対照群の間に有意な差はみられなかっ た。 3.4. 病理組織学的検査 精巣で認められた個体別の病理組織学的所見をTable 3 に示し,代表的な変化を Figure 8 に示す。 温浴群の 3 例では,全例で精母細胞,円形精子細胞,及び伸長精子細胞数の減少が認め られた。精母細胞及び円形精子細胞の減少は軽度から中等度であり,伸長精子細胞数の減 少は 3 例全例ともに重度であった。また,温浴群の全例で精母細胞及び精子細胞の変性像
が認められた。対照群では前述の変化は認められなかった。
3.5. miRNA マイクロアレイ解析
温浴群の精巣では,対照群と比較して 11 種の miRNA 量が上昇(>2.00 倍)し,13 種の
miRNA 量が低下(<0.50 倍)していた(Table 4)。発現量が低下した miRNA のうち,miR-34/449
ファミリーに属する miR-34b-3p,miR-34b-5p,miR-34c-3p,miR-34c-5p,miR-449a,及び
miR-449b-5p は,対照群と比較して温浴群で 0.23-0.42 倍の低下を示した。一方で,精巣特
異的miRNA である miR-202-5p の発現量には,2 群間で差は認められなかった。miR-508-3p
については,本マイクロアレイ解析では検出できなかった。
3.6. RT-qPCR 解析
精巣中miRNA の測定結果を Table 5 及び Figure 9 に,血漿中 miRNA の測定結果を Figure
10 にそれぞれ示した。
精巣では温浴群でmiR-34b-5p 及び miR-34c-5p の発現量が対照群よりも低値を示した(P <
0.05)。miR-449a の発現量には群間有意差は認められなかったが,対照群と比較して約 13% の低値を示した(Figure 9)。温浴群の 3 例における個体別の miR-34b-5p 及び miR-34c-5p 発
現量は,共に動物番号005 が最も低値を示したのに対し,動物番号 006 は最も高値を示し
た。血漿では,対照群におけるmiR-34c-5p 量の変動幅が約 4 倍と大きかったため,温浴群
では対照群の変動範囲を超える顕著な上昇は観察されなかった。miR-202-5p についても,
4. 考察 本章ではカニクイザルに 5 日間連続で精巣温浴を施したところ,全例で伸長精子細胞, 精母細胞,及び円形精子細胞数の減少を特徴とした精巣毒性が認められた。カニクイザル に精巣温浴を施すと,まずパキテン期精母細胞及び円形精子細胞を中心にアポトーシスが 誘導されることが過去に報告されている(26)。カニクイザルの精子形成サイクルが42 日間で あることを考慮すると(12),温浴開始時点でパキテン期精母細胞及び円形精子細胞であった 精細胞が,4 週間後には伸長精子細胞へと分化したことが推定された。したがって,本試験 において伸長精子細胞を主体とした細胞数の減少が認められたことについては,温浴によ りパキテン期精母細胞及び円形精子細胞が障害されたことを反映しているものと推察され た。 Lue らが報告した精巣温浴障害モデルでは,血漿中テストステロン濃度の減少が 2/3 例で みられているが,同値の変動幅は最大でも約 4 倍程度であり,精巣障害が最も顕著であっ た動物では同値の減少が認められなかったことが報告されている(26)。一般的にマカク属の サルでは,テストステロンのパルス状分泌が認められ,分泌期と非分泌期とでは同値に 4 倍程度の変動が認められることが知られている(38)。このことから Leu らの温浴モデルで認 められた同値の変動幅は,テストステロンの分泌パターンの変化に影響されていた可能性 が推察されている。実際に精巣温浴群では,対象群と比較して血漿中テストステロン濃度 に差が認められなかったことから,本試験に用いた精巣温浴モデルにおけるテストステロ ンへの影響は軽微であった可能性が示唆された。また本試験で作出した精巣温浴モデルで は,試験期間を通じて一般状態,体重及び摂餌量,並びに血液化学パラメータに変化が認 められなかったことから,本温浴条件では他臓器への影響を与えることなく,ホルモン非 依存的な精巣障害を惹起していた可能性が示唆された。 カニクイザルの精巣を用いたmiRNA マイクロアレイ解析により,精巣温浴群では対照群 と比較して11 種の miRNA 量の上昇及び 13 種の miRNA 量の低下が認められた。発現量の