Control of root cap maturation and cell detachment by BEARSKIN transcription factors in Arabidopsis
10
0
0
全文
(2) RESEARCH ARTICLE. preferentially expressed in the root cap initials and promotes formative divisions. SMB is expressed in differentiated root cap cells and suppresses FEZ expression. SMB was independently identified in our root-specific activation tagging screen and overexpression of SMB confers root cap-like characteristics to the epidermis (Waki et al., 2013). Epidermal cells in the SMB-overexpressing roots underwent additional anticlinal divisions, which normally occur in the LRC but not in the epidermis, in the meristematic region. These root cap-like epidermal cells were eventually lost in the differentiation zone, possibly via programmed cell death (Waki et al., 2013). SMB belongs to the group I NAC transcription factors (Pereira-Santana et al., 2015). In Arabidopsis, group I NAC transcription factors also include VND/NST/SND proteins, which are known to promote secondary cell wall (SCW) synthesis in vascular cells, and the two BEARSKIN (BRN) transcription factors, BRN1 and BRN2. Previous GUS reporter analyses indicated that SMB, BRN1 and BRN2 are expressed in largely overlapping patterns in Arabidopsis roots; SMB is expressed in the entire region corresponding to the differentiated COL and LRC cells, whereas BRN1 and BRN2 expression appeared to be stronger in the COL and the flanking LRC than peripheral LRC (Bennett et al., 2010). Lossof-function smb mutants are defective in root cap dehiscence and cell patterning in the stem cell region. Mutants of a single BRN gene are normal, whereas brn1 brn2 double mutants are defective in cell detachment (Bennett et al., 2010). Overexpression of SMB, BRN1 and BRN2 induced SCW formation in the root vasculature, as did overexpression of VND/NST/SND members, suggesting that group I NAC transcription factors have a shared regulatory function (Bennett et al., 2010). However, as SCW synthesis does not occur in wild-type root caps, endogenous functions of SMB, BRN1 and BRN2 are expected to differ from those of the VND/NST/SND members (Bennett et al., 2010). Despite extensive studies of the genetic pathways controlling root cap differentiation, interaction between these pathways seems limited (Bennett et al., 2014). Nevertheless, the SMB/BRN1/BRN2 pathway appears to act after the cell specification step and therefore is more closely linked to the differentiation processes than are the other pathways (Bennett et al., 2014; Willemsen et al., 2008). In this study, we performed detailed expression analyses of SMB, BRN1 and BRN2, and identified their downstream target genes. Our data suggest that expression of BRN1 and BRN2 is tightly linked to cell positioning on the root surface. Furthermore, we found that SMB, BRN1 and BRN2 regulate the expression of genes involved in lipid metabolism, endomembrane organization and cell separation, cellular events that are closely associated with the classically documented characteristics of the outer root cap layers. A gene encoding glycosyl hydrolase 28 (GH28) polygalacturonase (PG) is directly activated by BRN1 and facilitates root cap detachment. Therefore, our analyses linked key regulators of root cap differentiation with a unique developmental feature of the root cap.. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. GUS staining (Bennett et al., 2010), which has low cellular resolution. To analyze the expression patterns of BRN1 and BRN2 in more detail, we generated transgenic plants that express a cellautonomous, nuclear-localized YFP-GUS (nYG) reporter (Waki et al., 2013) under the BRN1 and BRN2 promoters. Observations of YFP fluorescence by confocal laser scanning microscopy (CLSM) revealed that transcription of BRN1 and BRN2 is restricted to the outermost root cap layer (Fig. 1D,G). Although weak expression was detected in the layer directly beneath the outermost layer, no expression was detected further inside the root (Fig. 1D,G). This expression pattern was confirmed at the protein level by complementing brn1 brn2 double mutants with BRN1-GFP or BRN2-GFP expressed under the respective BRN promoter (Fig. S1B,C). These results suggest that the spatial expression pattern of BRN1 and BRN2 is distinct from that of SMB. The highly restricted expression of BRN1 and BRN2 in the outer root cap layers also suggests a specific role for BRN1 and BRN2 in the later phases of root cap differentiation. SMB modifies the spatial expression pattern of BRN1 and BRN2. Ectopic expression of SMB confers root cap-like characteristics to the epidermis of Arabidopsis roots (Waki et al., 2013). To link SMB functions to root cap characteristics, we performed microarray analysis using dexamethasone (DEX)-inducible SMBoverexpressing plants (35S-iSMB) (Waki et al., 2013). As described below, this analysis identified BRN1 and BRN2 among the genes upregulated by overexpression of SMB. BRN1 and BRN2 mRNA levels showed a 3.7- and 7.4-fold increase, respectively, compared with DEX-treated control plants, suggesting that SMB either directly or indirectly activates BRN1 and BRN2. To examine whether SMB modifies the spatial expression patterns of BRN1 and BRN2, we crossed the pBRN1-nYG and pBRN2-nYG reporter lines with Q2610-iSMB plants, which ectopically express SMB throughout the root meristematic region in a DEX-dependent manner (Waki et al., 2013). CLSM observation of DEX-treated plants revealed that ectopic expression of BRN1 and BRN2 was limited to the epidermal cells exposed to the root surface (Fig. 1F,I). The inner tissue layers, including the cortex and endodermis, did not express BRN1 and BRN2, even though SMB was expressed by the ubiquitous Q2610 promoter (Waki et al., 2013). The BRN1 and BRN2 reporter lines were also crossed with the loss-of-function smb mutants (Willemsen et al., 2008). Expression of BRN1 and BRN2 reporters was lost in the majority of LRC cells in the smb mutants, but was retained in the COL cells (Fig. 1E,H). By contrast, transcription from the SMB promoter did not respond to the altered SMB expression (Fig. 1B,C). These results indicate that BRN1 and BRN2 expression primarily depends on the cell being on the root surface, and requires SMB in the LRC.. The spatial expression patterns of SMB have been investigated by in situ hybridization, transcriptional reporter analysis and complementation of smb mutants with SMB-GFP fusion proteins expressed under the SMB promoter (Bennett et al., 2010; Waki et al., 2013; Willemsen et al., 2008). These experiments have consistently indicated that SMB is transcribed specifically in the differentiated root cap cells (Fig. 1A; Fig. S1A). Expression patterns of BRN1 and BRN2, however, have been investigated solely by whole-mount 4064. Although some NAC transcription factors are known to bind to specific DNA targets and to regulate gene transcription (Ernst et al., 2004), the regulatory capacity of SMB, BRN1 and BRN2 proteins has not been demonstrated. Notably, SMB, BRN1 and BRN2 show little conservation in the WQ box sequences that are responsible for activating transcription by the group I NAC protein NST3 (Bennett et al., 2010; Ko et al., 2007). To address this, we constructed plasmids to express either full-length or segments of SMB, BRN1 and BRN2 proteins fused with the yeast GAL4 DNAbinding domain (GAL4-BD) (Fig. 2). These plasmids were introduced into tobacco protoplasts together with a reporter. DEVELOPMENT. SMB, BRN1 and BRN2 activate gene transcription RESULTS Expression of BRN1 and BRN2 is restricted to the outer root cap layers.
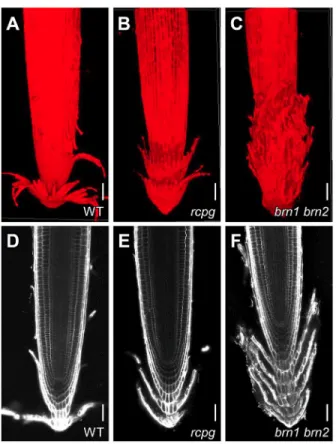
(3) RESEARCH ARTICLE. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. Fig. 1. Transcription of BRN1 and BRN2 is restricted to the outermost root cap layers. (A-C) Confocal images of a transcriptional reporter line of SMB. (A) Nuclear-localized YFP fluorescence indicates ubiquitous transcription of SMB in the differentiated root cap cells. (B,C) This transcription pattern is not affected by the loss-of-function smb mutation (B) or by SMB overexpression (DEX-treated Q2610iSMB) (C). (D-I) Confocal images of transcriptional reporter lines of BRN1 and BRN2. Transcription of BRN1 and BRN2 is restricted to the outermost root cap layers (D,G). The smb mutant lacks transcription of BRN1 and BRN2 in the LRC (white arrowheads in E and H), whereas SMB overexpression activates ectopic transcription of BRN1 and BRN2 in the epidermis (open arrowheads in F and I). BRN1 and BRN2 are ectopically expressed only in the epidermal cells exposed to the root surface. In each set of panels, boxed regions in the left panel are magnified in the center and right panels. Asterisk, QC; Epi, epidermis; COL, columella; LRC, lateral root cap. Scale bars: 200 µm, left panels; 20 µm, center and right panels.. plasmid that contained a firefly luciferase (fLuc) gene downstream of the GAL4-binding upstream activation sequence (UAS). Another plasmid harboring the CaMV35S promoter driving the renilla luciferase reporter (35S-rLuc) was co-introduced as an internal control. Measurement of the luciferase activities in the. transfected protoplasts revealed that the C-terminal domains of the three NAC factors can activate gene transcription, whereas the N-terminal region containing the NAC DNA-binding domain cannot (Fig. 2). These results indicate that SMB, BRN1 and BRN2 act as transcription activators. Fig. 2. The C-terminal domains of SMB, BRN1 and BRN2 activate gene transcription in plant cells. The structures of the fusion proteins (left). The bar graph indicates the transcription activation capacities of the fusion proteins measured by the dual luciferase assay with the value of the control (GAL4-BD alone) set to 1. Error bars indicate the s.d. calculated from five biological replicates. This graph shows a representative result of two experiments.. GAL4-BD. Control SMB -FL. NAC. SMB -N. NAC. SMB -C BRN1 -FL. NAC. BRN1 -N. NAC. BRN2 -FL. NAC. BRN2 -N. NAC. DEVELOPMENT. BRN1 -C. BRN2 -C VP16AD VP16-AD. 0. 5. 10. 15. 20. F-luc / R-luc 4065.
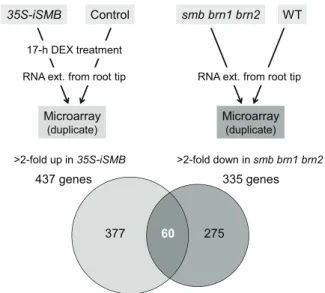
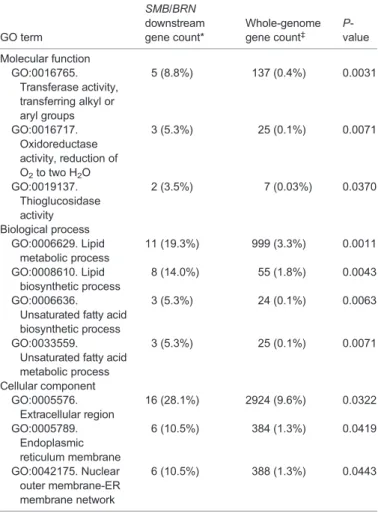
(4) RESEARCH ARTICLE. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. SMB, BRN1 and BRN2 regulate the expression of genes associated with root cap differentiation. 35S-iSMB. smb brn1 brn2. Control. WT. 17-h DEX treatment RNA ext. from root tip. RNA ext. from root tip. Microarray. Microarray. (duplicate). (duplicate). >2-fold up in 35S-iSMB. >2-fold down in smb brn1 brn2. 437 genes. 335 genes. 377. 60. 275. Fig. 3. Comparative microarray analyses to identify genes that act downstream of SMB, BRN1 and BRN2. In one experiment (left), transcriptomes of SMB-overexpressing roots (35S-iSMB) and control roots were compared, whereas in the other experiment (right), transcriptomes of the smb brn1 brn2 triple mutant and wild-type roots were compared. We identified 60 genes as candidates that act downstream of the three NAC transcription factors.. 4066. Fig. 4. RCPG is specifically transcribed in the detaching root cap cells in a manner that depends on BRN1 and BRN2, but not SMB. (A-D) Confocal images of the root tip of the transcriptional reporter lines of RCPG. RCPG is specifically transcribed in the detaching root cap cells in the wild type. This expression pattern is essentially the same in the smb mutant, but dramatically reduced in the smb brn1 brn2 and brn1 brn2 mutants. Insets in B-D indicate restoration of reporter expression in the cross between respective reporter lines with wild-type plants. Scale bars: 50 µm. (E) Measurement of endogenous RCPG mRNA levels in the wild-type and mutant root tips by RT-qPCR. Error bars indicate the s.d. from three independently pooled root tips. The graph shows a representative result of two experiments.. BRN1 and BRN2 activate a polygalacturonase gene in the outermost root cap layer. In addition to the lipid synthesis and endomembrane-associated genes described above, a gene encoding a GH28 family protein (At1G65570) was identified among the downstream genes. GH28 proteins are known to function as PGs in bacteria, fungi and plants (Abbott and Boraston, 2007; Cao, 2012; Sprockett et al., 2011). Arabidopsis mutants defective in GH28 PGs are compromised in developmentally regulated cell separation in pollen and carpel dehiscence (Ogawa et al., 2009; Rhee et al., 2003). Because cell detachment is a hallmark of mature root cap cells and the loss-offunction smb and brn1 brnn2 mutants are defective in root cap dehiscence (Bennett et al., 2010), we focused our analysis on the At1G65570 gene. The spatiotemporal expression pattern of At1G65570 was analyzed using transgenic plants harboring the 1.8 kb promoter of At1G65570 fused with the nYG reporter gene. CLSM observation of the root tip revealed tightly regulated transcription of At1G65570 in the outermost root cap layer (Fig. 4A). Based on this expression pattern, we named At1G65570 as ROOT CAP POLYGALACTURONASE (RCPG). The RCPG reporter was also introduced into the smb, brn1 brn2 and smb brn1 brn2 mutants. Observation of multiple independent lines revealed a dramatic reduction of RCPG expression in the brn1 brn2 and smb brn1 brn2 mutants (Fig. 4C,D), but not in the smb mutant (Fig. 4B). Crossing these reporter lines with wild-type plants restored reporter expression to comparable levels to that in the wild type (Fig. 4B-D, insets). Combined with the fact that reduced reporter expression was observed both in brn1 brn2 and smb brn1 brn2. DEVELOPMENT. To search for endogenous targets of SMB, BRN1 and BRN2, we performed transcriptome analyses of the root tip segments of 35SiSMB and a control line (35S-GVG) treated with DEX for 17 h (Fig. 3). We identified 437 genes exhibiting more than twofold upregulation in 35S-iSMB when compared with the control (Fig. 3). As mentioned above, BRN1 and BRN2 were found among the upregulated genes, indicating that genes upregulated by SMB via BRN1 and/or BRN2 are included in the 437 genes. To exclude genes artificially activated by SMB, BRN1 and BRN2 overexpression, comparable transcriptome analyses between the triple mutant smb brn1 brn2 and wild-type roots were performed in parallel. A total of 335 genes showing more than twofold downregulation in the smb brn1 brn2 triple mutant root tips were identified (Fig. 3). By extracting the overlap between the two comparative transcriptome analyses, 60 genes were identified as putative downstream targets of the three NAC factors (Fig. 3; Table S1). Transcriptional nYG reporter lines were generated for 11 of the 60 genes and CLSM observations revealed preferential expression of all 11 genes in the outer root cap layers (Fig. 4A; Fig. S2). This suggests that our combinatorial transcriptome approach successfully identified endogenous target genes. A gene ontogeny (GO) enrichment analysis of 57 annotated genes out of the 60 candidates by the AmiGO program (Carbon et al., 2009) revealed an enrichment of genes encoding enzymes (Table 1). Genes associated with lipid metabolism were highly enriched (Table 1). The GO enrichment analysis also indicated overrepresentation of extracellular proteins as well as ER and/or nuclear membrane-associated proteins (Table 1). As discussed below, both lipid synthesis and endomembrane rearrangement occur at the transition from the statocyte to the secretion cells in the COL lineage (Maitra and De, 1972; Sievers et al., 2002). Thus, our microarray and expression analyses suggest that the three NAC factors regulate a broad range of subcellular events associated with root cap differentiation..
(5) RESEARCH ARTICLE. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. Table 1. GO enrichment analysis of genes acting downstream of SMB, BRN1 and BRN2. GO term Molecular function GO:0016765. Transferase activity, transferring alkyl or aryl groups GO:0016717. Oxidoreductase activity, reduction of O2 to two H2O GO:0019137. Thioglucosidase activity Biological process GO:0006629. Lipid metabolic process GO:0008610. Lipid biosynthetic process GO:0006636. Unsaturated fatty acid biosynthetic process GO:0033559. Unsaturated fatty acid metabolic process Cellular component GO:0005576. Extracellular region GO:0005789. Endoplasmic reticulum membrane GO:0042175. Nuclear outer membrane-ER membrane network. SMB/BRN downstream gene count*. Whole-genome gene count‡. Pvalue. 5 (8.8%). 137 (0.4%). 0.0031. 3 (5.3%). 25 (0.1%). 0.0071. 2 (3.5%). 7 (0.03%). GFPer-35S-GVG) were generated and grown on DEX-containing media. CLSM observation of the root tips revealed a dramatic change in the cell adhesion properties in the root cap: in roots overexpressing RCPG, the outermost root cap cells sloughed off individually (Fig. 5B,C), whereas the control root cap cells detached as a layer (Fig. 5A). This observation strongly suggests that RCPG promotes cell separation. Interestingly, cell separation was not ectopically induced in the inner root cap layers, even though strong expression of the co-induced GFP reporter indicated that GVGmediated overexpression occurred throughout the root cap (Fig. 5B). This indicates that RCPG-mediated cell separation requires unknown factors and/or conditioning that specifically occurs in the outermost root cap layer.. 0.0370. RCPG facilitates root cap removal 11 (19.3%). 999 (3.3%). 0.0011. 8 (14.0%). 55 (1.8%). 0.0043. 3 (5.3%). 24 (0.1%). 0.0063. 3 (5.3%). 25 (0.1%). 0.0071. 16 (28.1%). 2924 (9.6%). 0.0322. 6 (10.5%). 384 (1.3%). 0.0419. 6 (10.5%). 388 (1.3%). 0.0443. *Among a total of 57 genes. ‡ Among a total of 30,471 genes.. mutants, these results suggest that the reduced expression of the reporter was caused by the loss of BRN1 and/or BRN2 functions, and was not due to the position of the T-DNA insertion or background mutations unrelated to brn1 and brn2. We confirmed this notion by measuring endogenous RCPG transcript levels by reverse transcription quantitative PCR (RT-qPCR) (Fig. 4E). Taken together, our data indicate that expression of RCPG does not require SMB functions, but depends on BRN1 and/or BRN2.. To examine whether RCPG activities are necessary for the detachment of root cap cells, we isolated and characterized a TDNA insertion mutant of RCPG (GABI_100C05, hereafter called the rcpg mutant) (Fig. S4A). RT-PCR analysis of the root tip segments of homozygous rcpg mutants indicated no accumulation of RCPG mRNA corresponding to the third exon, and no splicing of the second intron harboring the T-DNA (Fig. S4B), suggesting that the expression and/or functions of RCPG proteins, if any, derived from the rcpg allele were compromised. Although the growth of the homozygous rcpg mutants was indistinguishable from that of the wild type, observations of the roots of 5-day-old seedlings revealed incomplete removal of detaching root cap layers in the rcpg mutant (Fig. 6). Although this rcpg defect was qualitatively similar to that of the brn1 brn2 mutant, it was less conspicuous in the rcpg mutant (Fig. 6). Detaching root cap layers of the rcpg and brn1 brn2 mutants exhibited a closed, bowl-like shape, as opposed to a flat plate-like shape seen in the wild-type roots (Fig. 6). As described below, this phenotype was rescued with RCPG-RFP proteins expressed under the RCPG promoter (Fig. 7). These results suggest that the mutation in RCPG led to a change in the morphology of the detaching root cap layers and to their incomplete separation from the root. Considering the dramatically reduced expression of RCPG in the brn1 brn2 mutants, these observations indicate that RCPG acts downstream of BRN1 and/or BRN2 and facilitates the removal of the outermost root cap layer. RCPG localizes to the apoplast. Consistent with the predicted role of RCPG in the degradation of cell wall pectins, the pSORT program predicted that the RCPG. GH28 proteins have been extensively studied in bacteria and fungi, and their catalytic mechanism and reaction selectivity between endo- and exo-PG have been elucidated at the atomic level (Abbott and Boraston, 2007; Shimizu et al., 2002; van Pouderoyen et al., 2003). Our modeling analyses indicate that RCPG folds into a βhelical structure with an open-ended substrate-binding cleft typical of endo-PGs (Fig. S3A-C) (Abbott and Boraston, 2007). Both the substrate-binding residues and the catalytic Asp residues are perfectly conserved in RCPG (Fig. S3D) (Armand et al., 2000; Pages et al., 2000; Shimizu et al., 2002). These residues are not only conserved in sequence but also in the superposition of the RCPG model with an experimentally determined bacterial PG structure (Fig. S3E,F) (Shimizu et al., 2002), supporting the notion that RCPG functions as an endo-PG. To test whether RCPG can promote cell separation in the root cap, DEX-inducible overexpression lines of RCPG (UAS-RCPG/UAS-. Fig. 5. Overexpression of RCPG alters the behavior of the detaching root cap cells. (A) A confocal image of the RCPG overexpression line grown without the DEX inducer. The outermost root cap cells detach as a layer (arrows). (B,C) Confocal images of a root overexpressing RCPG. Cells in the outermost layer detach independently by cleavage at the longitudinal walls (arrowheads). Image shown in C is a reconstructed 3D view of the root tip, colored according to the depth across the root diameter for easy recognition. Scale bars: 50 µm.. 4067. DEVELOPMENT. RCPG promotes cell separation.
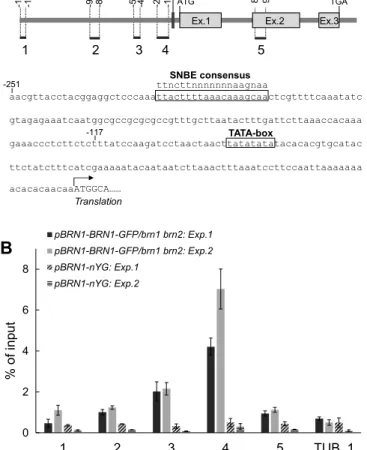
(6) RESEARCH ARTICLE. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. Fig. 7. RCPG-RFP fusion proteins localize to the apoplast and complement the defects of the rcpg mutant. (A,B) A median confocal section of a pRCPG-RCPG-RFP root. RFP fluorescence is specifically detected in the space surrounding the detaching root cap cells (arrowheads). (B) A magnified view of the boxed region in A. (C-F) Expression of RCPG-RFP is associated with the reversion of detaching root cap morphology from the closed bowl-like (F) to the flat plate-like (E) shape, which is typical of rcpg mutants (D) and the wild type (C), respectively. Green indicates FDA-positive viable cells. Scale bar: 50 µm.. protein is targeted to the extracellular space. To confirm this, we generated a construct to express RCPG-RFP fusion proteins driven by the RCPG promoter ( pRCPG-RCPG-RFP) and introduced this construct into the rcpg mutants. CLSM observations of the resulting transgenic plants revealed RFP fluorescence in the apoplasts surrounding the outermost root cap cells (Fig. 7A,B). RFP fluorescence appeared as a band surrounding the detaching root cap layer (Fig. 7B). In segregating T2 progeny, individuals with RCPG-RFP fluorescence had detaching root cap layers exhibiting a flat plate-like morphology, as seen in the wild type (Fig. 7C,E). Those that lacked RFP fluorescence, however, remained to have a closed bowl-like shape, as seen for the rcpg mutant (Fig. 7D,F). These results confirmed that functional RCPG-RFP proteins localized to the apoplast, consistent with the predicted role of RCPG in the degradation of cell wall pectins. BRN1 directly binds to the RCPG promoter. NAC transcription factors regulate gene transcription by binding to specific sequences in their target promoters (Ernst et al., 2004). Spatially overlapping expression of RCPG with BRN1 and BRN2, loss of RCPG expression in the brn1 brn2 mutant, and the ability of BRN1 and BRN2 to activate gene transcription collectively suggest a mechanism in which BRN1 and BRN2 directly bind to the RCPG promoter to activate RCPG expression in the detaching root cap cells. To test this possibility, chromatin immunoprecipitation followed by quantitative PCR (ChIP-qPCR) was performed using 4068. DISCUSSION Expression patterns of BRN1 and BRN2 are different from that of SMB. Although the smb brn1 brn2 triple mutants were strongly affected both in cell separation and stem cell divisions, the brn1 brn2 double mutants exhibited a weak cell separation defect (Bennett et al., 2010). Whole-mount GUS staining suggested that SMB, BRN1 and BRN2 had similar expression patterns in the root cap (Bennett et al., 2010). However, by using a cell-autonomous reporter and CLSM observations, we found that the expression of BRN1 and BRN2 is tightly restricted to the outer root cap layers. Complementation of the brn1 brn2 mutants with the BRN1-GFP and BRN2-GFP fusion proteins expressed under the respective BRN promoters also supported the conclusion that functional expression of BRN1 and BRN2 is limited to the outer root cap layers. The apparently broader expression of BRN1 and BRN2 found in the previous study is likely attributable to the use of diffusive GUS staining and the wholemount set up. Although expression of BRN1 and BRN2 was ectopically activated in the epidermis by the overexpression of SMB, the surface-specific expression was still maintained in spite of the ubiquitous expression of SMB. This indicates that expression of. DEVELOPMENT. Fig. 6. The rcpg and brn1 brn2 loss-of-function mutants fail to separate from detaching root cap layers. (A-C) Reconstructed 3D views of the wildtype (A), rcpg mutant (B) and brn1 brn2 mutant (C) roots. (D-F) Median confocal sections of the wild-type (D), rcpg mutant (E) and brn1 brn2 mutant (F) roots. Roots were observed without lifting from the agar surface. The detaching root cap layers of the rcpg and brn1 brn2 mutants retain a closed bowl-like shape (B,C), whereas those of the wild-type root are flattened (A). Scale bars: 50 µm.. the aforementioned pBRN1-BRN1-GFP plants that complemented the brn1 brn2 mutant (Fig. S1B). Purification of chromatin fractions from the root tip cells using anti-GFP antibodies followed by qPCR measurement revealed significant enrichment of DNA fragments covering 117-251 bp upstream of the first ATG of RCPG, demonstrating that BRN1 directly binds to the proximal promoter region of RCPG (Fig. 8B). This region contains a consensus sequence for the secondary wall NAC-binding element (SNBE) (TTnCTTnnnnnnnAAGnAA), a binding site for the VND transcription factors (Fig. 8A). Taken together, our ChIP-qPCR analysis suggests that BRN1 and, by inference, BRN2 directly control the expression of RCPG in the outermost root cap layer, and thereby promotes its dehiscence..
(7) RESEARCH ARTICLE. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. 860 951. ATG. Ex.1. 1. 2. 3. TGA. Ex.2. 4. Ex.3. SMB, BRN1 and BRN2 regulate a broad range of subcellular events associated with root cap differentiation. 5. SNBE consensus ttncttnnnnnnnaagnaa aacgttacctacggaggctcccaaattacttttaaacaaagcaactcgttttcaaatatc. -251. gtagagaaatcaatggcgccgcgcgccgtttgcttaatactttgattcttaaaccacaaa -117 TATA-box gaaaccctcttctctttatccaagatcctaactaacttatatatatacacacgtgcatac ttctatctttcatcgaaaaatacaataatcttaaactttaaatccttccaattaaaaaaa acacacaacaaATGGCA…… Translation pBRN1-BRN1-GFP/brn1 brn2: Exp.1. B. pBRN1-BRN1-GFP/brn1 brn2: Exp.2. 8. pBRN1-nYG: Exp.1 pBRN1-nYG: Exp.2. % of input. 6. 4. 2. 0. 1. 2. 3. 4. Therefore, a mechanism that senses outer cell positioning is required and the resulting positional information should be eventually converted to the transcriptional control of the BRN1 and BRN2 promoters.. 5. TUB_1. Fig. 8. BRN1 directly binds to the proximal promoter region of RCPG. (A) The RCPG gene structure. Boxes indicate exons. Regions analyzed by ChIP-qPCR (B) are shown by bars numbered 1-5 together with the distance from the first ATG. The TATA-box element predicted by the TSSP-TCM program (Shahmuradov et al., 2005) and the consensus sequence for the secondary wall NAC-binding element (SNBE) are boxed in the sequence below the diagram. (B) qPCR measurements of RCPG genomic fragments coimmunoprecipitated with the BRN1-GFP protein. A transcriptional reporter line of BRN1 ( pBRN1-nYG) was used as a negative control. A region in the unrelated TUB1 gene was used to set the background amplification level. Results from two independent experiments are shown. Error bars indicate s.d. from three qPCR measurements (technical replicates).. BRN1 and BRN2 is strictly dependent on surface positioning, as well as on a second input that is associated with root cap identity. Although SMB is a good candidate for a factor that mediates the second input, the persistent expression of BRN1 and BRN2 in the COL of smb mutants suggests the existence of as yet unknown factors that promote BRN1 and BRN2 expression in COL. In view of the widely accepted concept of position-dependent tissue patterning and its functional significance in the Arabidopsis root (Petricka et al., 2012), mechanisms that underlie surface-specific gene expression are of great interest. In the Arabidopsis shoot, cells in the outermost L1 layer have a clonal lineage and L1-specific gene expression is maintained by the binding of the HD-ZIP IV transcription factors ARABIDOPSIS THALIANA MERISTEM LAYER1 (ATML1) and PROTODERMAL FACTOR 2 (PDF2) to the promoters of target genes as well as to the promoter of PDF2 itself (Abe et al., 2003). Although mechanisms regulating surfacespecific gene expression have not been reported for the roots, a lineage-dependent mechanism is not expected to operate in the root cap, as cells constituting the root cap continuously turn over.. SMB, BRN1 and BRN2 constitute the group I NAC transcription factors together with the VND and NST/SND members that control SCW synthesis in vascular cells (Pereira-Santana et al., 2015). Based on the finding that SMB, BRN1 and BRN2 activate SCW synthesis in the root vasculature (as do the VND proteins), group I NAC proteins appear to possess a shared activation capacity for SCW synthesis (Bennett et al., 2010). However, this overexpression phenotype is likely artificial (Bennett et al., 2010). In this study, we adopted a comparable transcriptome approach to search for endogenous targets of SMB, BRN1 and BRN2. A GO enrichment analysis of the identified genes revealed a bias towards enzymecoding genes, especially those involved in lipid synthesis. Reporter analyses indicated that most of these enzyme-coding genes are specifically expressed in the outer root cap layers. In a classical ultrastructural observation of Medicago root caps, lipid bodies were found to develop as root cap cells mature and then to disappear in the detaching cells (Maitra and De, 1972). It has been proposed that in the outer root cap layers, starch stored in the amyloplasts is rapidly converted to lipids for energy storage and then consumed in the outermost root cap cells. In this regard, BRN-dependent activation of lipid synthesis in the outer root cap layers likely provides a molecular basis for a root cap maturation process. In addition, our GO enrichment analysis revealed an overrepresentation of nuclear- and ER membrane-associated proteins. COL cells are known to undergo dramatic rearrangement of the subcellular compartments during the transition from statocytes to secretion cells (Sievers et al., 2002). Therefore, we suspect that the identified membrane-associated proteins function in this transition of COL differentiation, though this hypothesis remains to be tested by future mutant analyses. BRN1 directly controls a gene that encodes a cell wall modification enzyme. Our transcriptome analysis identified a number of extracellular proteins that act downstream of the three NAC factors and we decided to perform functional analysis of one of these, RCPG. RCPG belongs to the GH28 family of PGs, which occur in bacteria, fungi and plants. In plants, some GH28 proteins are known to degrade cell wall pectins and thereby promote cell separation. For example, Arabidopsis with mutations in QUARTET2 (QRT2) and QRT3 are defective in the separation of the four daughter cells produced from the microspore mother cell (Preuss et al., 1994). Similarly, mutants of ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE1 (ADPG1) and ADPG2 exhibit incomplete dehiscence of siliques and anthers. Recombinant ADPG1, ADPG2 and QRT3 proteins showed PG activities (Ogawa et al., 2009; Rhee et al., 2003). These reports highlight the essential role of GH28 proteins in pectin degradation and developmentally regulated cell separation in plants. GH28 proteins have been extensively characterized structurally for their catalytic mechanisms, reaction processivity and exo/endo selectivity (Armand et al., 2000; Pages et al., 2000; Shimizu et al., 2002; van Pouderoyen et al., 2003). Our modeling study indicated that RCPG shares a conserved protein folding with known GH28 endo-PGs, as well as perfectly conserved positioning of amino acid residues that 4069. DEVELOPMENT. -251 -117. -507 -430. -986 -875. -1749 -1682. A.
(8) RESEARCH ARTICLE. MATERIALS AND METHODS Plant materials. Arabidopsis thaliana (L.) Heynh accession Col-0 was used as the wild type. smb (smb-3, SALK_143526), brn1 (brn1-1, SALK_151986), brn2 (brn2-1, SALK_151604), 35S-iSMB, Q2610-iSMB and pSMB-nYG lines have been described previously (Bennett et al., 2010; Waki et al., 2013; Willemsen et al., 2008). Seeds of the rcpg mutant (GABI_100C05) were obtained from the Arabidopsis Biological Resource Center (Columbus, OH, USA) and backcrossed with wild-type plants. The T-DNA insertion was found in the second intron (1415 bp downstream of the first ATG) with deletion of the flanking 25 bp region rather than in the third exon [as annotated in the GABI-Kat database (https://www.gabi-kat.de/) (Fig. S4A)]. The transgenic lines described below were in the wild-type Col-0 background, unless noted otherwise.. 4070. Generation of transgenic plants. Nuclear-localized YFP-GUS (nYG) reporter constructs of BRN1, BRN2 and the genes identified in the microarray analyses were constructed by amplifying the promoter regions of the respective genes by PCR from wildtype genomic DNA using the primers listed in Table S2. PCR fragments were digested with restriction enzymes at the restriction sites incorporated at the end of the primers and inserted into the pBI-Kan-nlsYG plasmid (Waki et al., 2011). The RCPG reporter construct was introduced into the smb, brn1 brn2 and smb brn1 brn2 mutant backgrounds in addition to wild-type Col-0. To express GFP-fusion proteins of SMB, BRN1 and BRN2 under the native promoters, genomic DNA fragments consisting of the promoter, coding region and introns were amplified by PCR from the wild-type genomic DNA with the primers listed in Table S2. PCR fragments were digested at the SalI and BamHI sites incorporated in the primers and inserted into pBIB-4xGly-sGFP-NosT, which harbors the coding sequence of the 4xGly linker and GFP (S65T) (Niwa et al., 1999) followed by the nopaline synthase (Nos) terminator region. The resulting constructs were introduced into the smb or brn1 brn2 mutants. For DEX-inducible overexpression of RCPG, a genomic fragment encompassing the entire RCPG-coding sequence and the introns was amplified from the wild-type genomic DNA with the primers listed in Table S2. PCR products were digested with ApaI and SpeI, and inserted into the pBIB-UAS-NosT plasmid (Waki et al., 2011). The resulting pBIB-UASRCPG construct was introduced into the host plants harboring pBIN-UASGFPer-35S-GVG (Waki et al., 2013). For expression of RCPG-RFP fusion proteins by the RCPG promoter, DNA fragments of the RCPG promoter and gene body regions (full coding region plus introns) were separately amplified from the wild-type genomic DNA with the primers listed in Table S2. The resulting fragments were sequentially inserted into the pDONR P2R_P3-tagRFP-OcsT plasmid harboring tagRFP (Merzlyak et al., 2007) and the octopine synthase terminator (OcsT) using Gateway technology (Life Technologies). The assembled insert was then transferred into the bialaphos-resistant binary vector pBm43GW (Karimi et al., 2005). Transcription activation assay. Plasmids that express either full-length or a segment of the NAC polypeptide and the yeast GAL4-BD were constructed by amplifying the corresponding parts of the NAC cDNA fragments by PCR using the primers listed in Table S2. PCR fragments were digested with BamHI and EcoRI at the sites incorporated at the end of each primer and inserted into p35S-GAL4BD (Waki et al., 2011). The transcription activation domain of Herpes VP16 was used as a positive control (Waki et al., 2011). A transcription activation assay was performed with the Dual Luciferase Assay System (Promega) using protoplasts prepared from the tobacco (Nicotiana tabacum L.) BY-2 cell cultures as described previously (Waki et al., 2011). Microarray analysis. Total RNA was extracted from about 1 cm of root tip segments using the RNeasy Plant Mini Kit (Qiagen). First-strand cDNA synthesis and preparation of Cy3- and Cy5-labeled cRNAs were performed with the Low Input Quick Amp Labeling Kit (Agilent Technologies). Labeled cRNAs were hybridized with the Agilent 4x44K Arabidopsis Gene Expression Microarray Ver.4.0 (Agilent Technologies) in a two-color format with dye-swapped biological duplicates. Microscopy. Confocal laser scanning microscopy (CLSM) was carried out with a Nikon C2 confocal microscope. Roots were stained with 10 µM of propidium iodide unless noted otherwise. Fluorescein diacetate (FDA) staining was performed by soaking the roots in a solution containing 2 µg/ml FDA. RNA extraction and RT-PCR analysis. Total RNA was extracted from the ∼5 mm root tip segments of 5-day-old seedlings using the RNeasy Plant Mini Kit (Qiagen). First-strand cDNA was synthesized using the PrimeScript RT Reagent Kit with gDNA Eraser (Takara Bio). RT-qPCR was performed with the primers listed in Table S2. DEVELOPMENT. are known to be important for catalysis and substrate recognition. Functional RCPG-RFP fusion proteins were localized to the apoplast, and overexpression of RCPG promoted cell separation. Expression of RCPG depends on BRN1 and/or BRN2, and BRN1 directly binds to the RCPG promoter. Taken together, these results strongly suggest that RCPG degrades cell wall pectins in the outermost root cap layers and thereby promotes cell separation. Although our results are consistent with the role of RCPG in root cap dehiscence, the loss-of-function rcpg mutant phenotype was relatively weak, and the outermost root cap layer still dehisced in the rcpg mutant. The Arabidopsis genome contains a total of 67 genes encoding putative pectin lyases (Cao, 2012) and several of these genes appear to be expressed in the root cap (Brady et al., 2007). Although RCPG is the only GH28 member among the identified 60 genes, it is possible that root cap separation is preconditioned by NAC-independent PG activities. Alternatively, efficient cell separation can be achieved by the modification of multiple classes of cell wall components. In this regard, it should be noted that CELLULASE3 (CEL3) and CEL5 genes, which encode putative cellulases, are specifically expressed in the outer root cap layers, and a weak phenotype is postulated for cel5 mutants (del Campillo et al., 2004). Our transcriptome analyses indicated a reduction in the expression of CEL3 and CEL5 in the smb brn1 brn2 triple mutant, as was suggested previously (Bennett et al., 2010), and weak upregulation of these genes by SMB overexpression. Although the roles of cellulase have not been reported in other instances of developmentally regulated cell dehiscence, it is possible that some degree of cellulose decomposition facilitates cell detachment. Another intriguing possibility is that RCPG primarily controls the shape of the detaching root cap layer and thereby facilitates its removal. In the rcpg mutant, the detaching root cap layers have a closed bowl-like shape as opposed to the flat plate-like shape seen in the wild type. It has been reported that in mutants of compromised pectin biosynthesis, root cap cells detached as individual cells instead of cell layers (Durand et al., 2009). A similar phenomenon was also observed in our RCPG-overexpressing lines. These observations suggest that the integrity of the pectin matrix is important for connecting the detaching root cap cell layers. RCPG may preferentially separate cells at their longitudinal walls and thereby promote the efficient removal of the root cap layer. In summary, our study provides a molecular link between the key regulators of root cap differentiation and the cellular events underlying root cap dehiscence. A catalog of genes acting downstream of the NAC transcription factors in the root cap will serve as an important reference, not only to identify molecular components associated with the previously described root cap differentiation process, but also to establish previously unknown cellular events occurring in the root cap differentiation process.. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331.
(9) with the SYBR Premix Ex Taq (Takara Bio). Measurements were normalized to the levels of ACTIN3 transcript. Molecular modeling. The three-dimensional model of RCPG was calculated at the SWISSMODEL site (http://swissmodel.expasy.org/). The crystal structure of the endopolygalacturonase I from Stereum purpureum complexed with two galacturonate molecules (PDB code: 1KCD, chain A) was used as template. Structural images were drawn using the PyMOL program (https://www. pymol.org/). ChIP-qPCR. Seedlings of pBRN1-BRN1-GFP plants (in the brn1 brn2 mutant background) were grown on nutrient agar plates for 5 days. To increase the fraction of root cap-derived cells, seedlings were transferred to fresh plates containing 25 µM indole acetic acid to induce lateral root formation. After 3 days, root tip segments with numerous short lateral roots were harvested and subjected to chromatin purification. ChIP experiments were carried out as previously described (Kim et al., 2014) using the anti-GFP antibody (1/100; ab290, Abcam). The ChIP fractions were used as template to measure bound DNA fragments by real-time PCR using the primers listed in Table S2. Acknowledgements We are grateful to Tom Bennett and Ben Scheres for the smb, brn1 and brn2 seeds; and to Masako Kanda and Emi Murata for technical assistance.. Competing interests The authors declare no competing or financial interests.. Author contributions M.K., S.-Y.H., A.I., J.-M.K., S.M. and K.N. conducted the experiments; M.K., J.-M.K., M.S., S.M. and K.N. designed the experiments; and M.K. and K.N. wrote the paper.. Funding This work was supported by Japan Society for the Promotion of Science KAKENHI grants [JP25113007, JP15K14548 and JP21570042 to K.N.].. Data availability Microarray data obtained in this study have been deposited in the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/) under the accession number GSE86443.. Supplementary information Supplementary information available online at http://dev.biologists.org/lookup/doi/10.1242/dev.142331.supplemental. References Abbott, D. W. and Boraston, A. B. (2007). The structural basis for exopolygalacturonase activity in a family 28 glycoside hydrolase. J. Mol. Biol. 368, 1215-1222. Abe, M., Katsumata, H., Komeda, Y. and Takahashi, T. (2003). Regulation of shoot epidermal cell differentiation by a pair of homeodomain proteins in Arabidopsis. Development 130, 635-643. Armand, S., Wagemaker, M. J. M., Sanchez-Torres, P., Kester, H. C. M., van Santen, Y., Dijkstra, B. W., Visser, J. and Benen, J. A. E. (2000). The active site topology of Aspergillus niger endopolygalacturonase II as studied by site-directed mutagenesis. J. Biol. Chem. 275, 691-696. Bennett, T., van den Toorn, A., Sanchez-Perez, G. F., Campilho, A., Willemsen, V., Snel, B. and Scheres, B. (2010). SOMBRERO, BEARSKIN1, and BEARSKIN2 regulate root cap maturation in Arabidopsis. Plant Cell 22, 640-654. Bennett, T., van den Toorn, A., Willemsen, V. and Scheres, B. (2014). Precise control of plant stem cell activity through parallel regulatory inputs. Development 141, 4055-4064. Brady, S. M., Orlando, D. A., Lee, J.-Y., Wang, J. Y., Koch, J., Dinneny, J. R., Mace, D., Ohler, U. and Benfey, P. N. (2007). A high-resolution root spatiotemporal map reveals dominant expression patterns. Science 318, 801-806. Cannesan, M. A., Durand, C., Burel, C., Gangneux, C., Lerouge, P., Ishii, T., Laval, K., Follet-Gueye, M.-L., Driouich, A. and Vicré -Gibouin, M. (2012). Effect of Arabinogalactan proteins from the Root Caps of Pea and Brassica napus on Aphanomyces euteiches Zoospore chemotaxis and germination. Plant Physiol. 159, 1658-1670.. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. Cao, J. (2012). The pectin lyases in Arabidopsis thaliana: evolution, selection and expression profiles. PLoS ONE 7, e46944. Carbon, S., Ireland, A., Mungall, C. J., Shu, S., Marshall, B., Lewis, S., AmiGO Hub and Web Presence Working Group. (2009). AmiGO: online access to ontology and annotation data. Bioinformatics 25, 288-289. del Campillo, E., Abdel-Aziz, A., Crawford, D. and Patterson, S. E. (2004). Root cap specific expression of an endo-beta-1,4-D-glucanase (cellulase): a new marker to study root development in Arabidopsis. Plant Mol. Biol. 56, 309-323. Dolan, L., Janmaat, K., Willemsen, V., Linstead, P., Poethig, S., Roberts, K. and Scheres, B. (1993). Cellular organisation of the Arabidopsis thaliana root. Development 119, 71-84. Driouich, A., Follet-Gueye, M.-L., Vicre-Gibouin, M. and Hawes, M. (2013). Root border cells and secretions as critical elements in plant host defense. Curr. Opin. Plant Biol. 16, 489-495. Durand, C., Vicre-Gibouin, M., Follet-Gueye, M. L., Duponchel, L., Moreau, M., Lerouge, P. and Driouich, A. (2009). The organization pattern of root border-like cells of Arabidopsis is dependent on cell wall homogalacturonan. Plant Physiol. 150, 1411-1421. Ernst, H. A., Olsen, A. N., Skriver, K., Larsen, S. and Lo Leggio, L. (2004). Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 5, 297-303. Fendrych, M., Van Hautegem, T., Van Durme, M., Olvera-Carrillo, Y., Huysmans, M., Karimi, M., Lippens, S., Guerin, C. J., Krebs, M., Schumacher, K. et al. (2014). Programmed cell death controlled by ANAC033/SOMBRERO determines root cap organ size in Arabidopsis. Curr. Biol. 24, 931-940. Hawes, M. C., Gunawardena, U., Miyasaka, S. and Zhao, X. (2000). The role of root border cells in plant defense. Trends Plant Sci. 5, 128-133. Karimi, M., De Meyer, B. and Hilson, P. (2005). Modular cloning in plant cells. Trends Plant Sci. 10, 103-105. Kim, J.-M., To, T. K., Tanaka, M., Endo, T. A., Matsui, A., Ishida, J., Robertson, F. C., Toyoda, T. and Seki, M. (2014). Highly reproducible ChIP-on-chip analysis to identify genome-wide protein binding and chromatin status in Arabidopsis thaliana. Methods Mol. Biol. 1062, 405-426. Ko, J.-H., Yang, S. H., Park, A. H., Lerouxel, O. and Han, K.-H. (2007). ANAC012, a member of the plant-specific NAC transcription factor family, negatively regulates xylary fiber development in Arabidopsis thaliana. Plant J. 50, 1035-1048. Maitra, S. C. and De, D. N. (1972). Ultrastructure of root cap cells : formation and utilization of lipid. Cytobios 5, 111-118. Merzlyak, E. M., Goedhart, J., Shcherbo, D., Bulina, M. E., Shcheglov, A. S., Fradkov, A. F., Gaintzeva, A., Lukyanov, K. A., Lukyanov, S., Gadella, T. W. J. et al. (2007). Bright monomeric red fluorescent protein with an extended fluorescence lifetime. Nat. Methods 4, 555-557. Niwa, Y., Hirano, T., Yoshimoto, K., Shimizu, M. and Kobayashi, H. (1999). Noninvasive quantitative detection and applications of non-toxic, S65T-type green fluorescent protein in living plants. Plant J. 18, 455-463. Ogawa, M., Kay, P., Wilson, S. and Swain, S. M. (2009). ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE1 (ADPG1), ADPG2, and QUARTET2 are Polygalacturonases required for cell separation during reproductive development in Arabidopsis. Plant Cell 21, 216-233. Pages, S., Heijne, W. H. M., Kester, H. C. M., Visser, J. and Benen, J. A. E. (2000). Subsite mapping of Aspergillus niger endopolygalacturonase II by site-directed mutagenesis. J. Biol. Chem. 275, 29348-29353. Pereira-Santana, A., Alcaraz, L. D., Castano, E., Sanchez-Calderon, L., Sanchez-Teyer, F. and Rodriguez-Zapata, L. (2015). Comparative genomics of NAC transcriptional factors in angiosperms: implications for the adaptation and diversification of flowering plants. PLoS ONE 10, e0141866. Petricka, J. J., Winter, C. M. and Benfey, P. N. (2012). Control of Arabidopsis root development. Annu. Rev. Plant Biol. 63, 563-590. Pi, L., Aichinger, E., van der Graaff, E., Llavata-Peris, C. I., Weijers, D., Hennig, L., Groot, E. and Laux, T. (2015). Organizer-derived WOX5 signal maintains root columella stem cells through chromatin-mediated repression of CDF4 expression. Dev. Cell 33, 576-588. Preuss, D., Rhee, S. Y. and Davis, R. W. (1994). Tetrad analysis possible in Arabidopsis with mutation of the QUARTET (QRT) genes. Science 264, 1458-1460. Rhee, S. Y., Osborne, E., Poindexter, P. D. and Somerville, C. R. (2003). Microspore separation in the quartet 3 mutants of Arabidopsis is impaired by a defect in a developmentally regulated polygalacturonase required for pollen mother cell wall degradation. Plant Physiol. 133, 1170-1180. Sarkar, A. K., Luijten, M., Miyashima, S., Lenhard, M., Hashimoto, T., Nakajima, K., Scheres, B., Heidstra, R. and Laux, T. (2007). Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 446, 811-814. Shahmuradov, I. A., Solovyev, V. V. and Gammerman, A. J. (2005). Plant promoter prediction with confidence estimation. Nucleic Acids Res. 33, 1069-1076. Shimizu, T., Nakatsu, T., Miyairi, K., Okuno, T. and Kato, H. (2002). Active-site architecture of endopolygalacturonase I from Stereum purpureum revealed by. 4071. DEVELOPMENT. RESEARCH ARTICLE.
(10) RESEARCH ARTICLE. Waki, T., Hiki, T., Watanabe, R., Hashimoto, T. and Nakajima, K. (2011). The Arabidopsis RWP-RK protein RKD4 triggers gene expression and pattern formation in early embryogenesis. Curr. Biol. 21, 1277-1281. Waki, T., Miyashima, S., Nakanishi, M., Ikeda, Y., Hashimoto, T. and Nakajima, K. (2013). A GAL4-based targeted activation tagging system in Arabidopsis thaliana. Plant J. 73, 357-367. Wang, J.-W., Wang, L. J., Mao, Y. B., Cai, W. J., Xue, H. W. and Chen, X. Y. (2005). Control of root cap formation by MicroRNA-targeted auxin response factors in Arabidopsis. Plant Cell 17, 2204-2216. Willemsen, V., Bauch, M., Bennett, T., Campilho, A., Wolkenfelt, H., Xu, J., Haseloff, J. and Scheres, B. (2008). The NAC domain transcription factors FEZ and SOMBRERO control the orientation of cell division plane in Arabidopsis root stem cells. Dev. Cell 15, 913-922.. DEVELOPMENT. crystal structures in native and ligand-bound forms at atomic resolution. Biochemistry 41, 6651-6659. Sievers, A., Braun, M. and Monshausen, G. B. (2002). The root cap: structure and function. In Plant Root: The Hidden Half (ed. Y. Waisel, A. Eshel and U. Kafkafi), pp. 33-47. New York: CRC Press. Sprockett, D. D., Piontkivska, H. and Blackwood, C. B. (2011). Evolutionary analysis of glycosyl hydrolase family 28 (GH28) suggests lineage-specific expansions in necrotrophic fungal pathogens. Gene 479, 29-36. van Pouderoyen, G., Snijder, H. J., Benen, J. A. and Dijkstra, B. W. (2003). Structural insights into the processivity of endopolygalacturonase I from Aspergillus niger. FEBS Lett. 554, 462-466. Vicre, M., Santaella, C., Blanchet, S., Gateau, A. and Driouich, A. (2005). Root border-like cells of Arabidopsis. Microscopical characterization and role in the interaction with rhizobacteria. Plant Physiol. 138, 998-1008.. Development (2016) 143, 4063-4072 doi:10.1242/dev.142331. 4072.
(11)
図

+2
関連したドキュメント
熱力学計算によれば、この地下水中において安定なのは FeSe 2 (cr)で、Se 濃度はこの固相の 溶解度である 10 -9 ~10 -8 mol dm
Two grid diagrams of the same link can be obtained from each other by a finite sequence of the following elementary moves.. • stabilization
The mGoI framework provides token machine semantics of effectful computations, namely computations with algebraic effects, in which effectful λ-terms are translated to transducers..
Standard domino tableaux have already been considered by many authors [33], [6], [34], [8], [1], but, to the best of our knowledge, the expression of the
An example of a database state in the lextensive category of finite sets, for the EA sketch of our school data specification is provided by any database which models the
A NOTE ON SUMS OF POWERS WHICH HAVE A FIXED NUMBER OF PRIME FACTORS.. RAFAEL JAKIMCZUK D EPARTMENT OF
H ernández , Positive and free boundary solutions to singular nonlinear elliptic problems with absorption; An overview and open problems, in: Proceedings of the Variational
Keywords: Convex order ; Fréchet distribution ; Median ; Mittag-Leffler distribution ; Mittag- Leffler function ; Stable distribution ; Stochastic order.. AMS MSC 2010: Primary 60E05
