健康・栄養食品研究 Vol. 17 No. 1 2020
doi: 10.20618/jhnfa.17.1_18
Regular Article
The protective effects of bilberry extract and its main constituents against
blue light-emitting diode (LED) light-induced damage in
human corneal epithelial cells
Emi Ooe*, Tomohiro Yako*, Yoshiki Kuse*, Tetsuya Sogon**,
Shinsuke Nakamura*, Masamitsu Shimazawa*, Hideaki Hara*
, †(Received: April 9, 2020; Accepted:May 19, 2020; Published online: July 1, 2020)
The cornea is located on the surface of the eyeball and is constantly activated with light and various physical stimuli. Hu-mans are exposed to UV and other light with various wavelengths during daily life. Light ranging from 380 nm to 530 nm com-prises high-energy visible light that is present in sunlight, fluorescent light, and light-emitting diodes (LEDs). In this report, we investigated the effects of bilberry extract and its anthocyanin constituents on corneal epithelial cell damage induced by blue LED light. Human corneal epithelial (HCE-T) cells were in the presence of bilberry extract or its anthocyanin constitu-ents followed by exposure to 2,000 lux of blue LED light. We found that bilberry extract or its anthocyanin constituconstitu-ents (delph-inidin 3-glucoside and cyanidin 3-glucoside) reduced cell death, reactive oxygen species (ROS) production and cell dehydroge-nase activity. In addition, bilberry extract inhibited blue LED light-induced changes in the mitochondrial membrane potential and caspase-3/7 activity, both of which are associated with apoptosis. These findings suggested that bilberry extract or its constituent anthocyanins have protective effects against blue LED light-induced cell damage by reducing ROS production. Key words: blue light-emitting diode light, corneal epithelial cells, reactive oxygen species, bilberry, anthocyanin
ヒト角膜上皮細胞における青色発光ダイオード(LED)
光誘起損傷に対するビルベリー抽出物および
その主成分の保護効果
大江 絵美*,矢古宇 智弘*,久世 祥己*,荘厳 哲哉**,
中村 信介*,嶋澤 雅光*,原 英彰*
, † 角膜は眼球表面に存在し,光をはじめ様々な刺激を常に受けている.我々の目は日常生活において,紫外線やその他の様々な 波長の光にさらされている. 380 nmから530 nmの範囲の光は,太陽光,蛍光灯,および発光ダイオード(LED)に存在する光 で,エネルギーレベルが高いことから「高エネルギー可視光線」と呼ばれている.本論文では,青色LED光によって誘発される 角膜上皮細胞障害に対するビルベリー抽出物とそのアントシアニン成分の影響を調査した.ヒト角膜上皮細胞(HCE-T)をビル ベリー抽出物またはその含有アントシアニンとともにプレインキュベートした後,2,000ルクスの青色LED光に曝露した.ビルベ リー抽出物またはそのアントシアニン成分(デルフィニジン3-グルコシドおよびシアニジン3-グルコシド)は,細胞死,活性酸素 種(ROS)の産生および細胞代謝活性を低下させることが明らかになった.さらに,ビルベリー抽出物は,ミトコンドリア膜電位† Corresponding author (E-mail: [email protected])
* Molecular Pharmacology, Department of Biofunctional Evaluation, Gifu Pharmaceutical University, 1–25–4 Daigakunishi, Gifu 501–1196, Japan
** Wakasa Seikatsu Co. Ltd., 22 Naginataboko-cho, Shijo-Karasuma, Shimogyo-ku, Kyoto, Japan * 岐阜薬科大学 生体機能解析学大講座 薬効解析学研究室 〒501–1196 岐阜県岐阜市大学西1–25–4 ** 株式会社わかさ生活 〒600–8008 京都府京都市下京区四条烏丸長刀鉾町22
とカスパーゼ-3/7活性の青色LED光による変化を抑制した.この結果から,ビルベリー抽出物またはその構成アントシアニンは, ROS生成を減少させることにより,青色LED光誘発角膜上皮細胞障害に対する保護効果を有することが示唆された.
キーワード: 青色発光ダイオード光,角膜上皮細胞,活性酸素,ビルベリー,アントシアニン
Journal of Nutritional Food, 17(1), 18–33, 2020
I. Introduction
A light-emitting diode (LED) is a light source that is replacing conventional illumination and is widely used in lighting, television, personal com-puters and liquid crystal displays. Compared to a conventional light source, the LED lamp has sev-eral advantages, including less heat generation, longer lifespan and superior energy efficiency.
Light contains various colors and energies de-pending on the wavelength. Wavelengths span-ning from 380 nm to 750 nm comprise visible light and can be recognized by the human eye. Among the visible spectrum, light harboring a wave-length of 380 nm to 530 nm is high-energy visible light and is associated with the pathogenesis of age-related macular degeneration and retinitis pigmentosa.1, 2) Furthermore, recent reports have
explored the relationship between blue LED light and corneal dysfunction in both in vivo and in vitro models.3, 4) These blue LED light-induced
cor-neal impairments can lead to dry eye syndrome. Blue LEDs are known to emit quite a large amount of high-energy visible light.5–7) Moreover,
in our previous studies, we found that exposure to blue LED light induced reactive oxygen spe-cies (ROS) and oxidative stress in photoreceptor cells.8, 9) It has also been shown that oxidative
stress induced cell death in retinal pigment cells and photoreceptor cells.10–12) On the other hand,
the effects of blue light on the health of corneal cells are less clear.13) The cornea is located on the
ocular surface, and is directly exposed to visible blue light, which can contribute to photophobia
and ocular pain.14)
In previous reports, it was suggested that UV-B irradiation enhances ROS production in corneal epithelial cells and causes inflammation and tis-sue damage.13, 15–20) Thus, we hypothesized that
blue LED light irradiation may induce oxidative stress in corneal cells as well as UV-B.
Bilberry (Vaccinium myrtillus L.) is a member of the Ericaceae family of plants that grow in northern European forests. Bilberry extract con-tains 15 types of anthocyanins21, 22) and has
an-tioxidant activity.23) Previous reports have
dem-onstrated that anthocyanins are the molecules responsible for bilberrys main pharmacological effects, which include antioxidant activity24–26)
and free radical scavenging activity.27) Bilberry
extract has been used to treat several eye condi-tions and may prevent glaucoma and the forma-tion of cataracts.28) Furthermore, bilberry extract
has been reported to improve visual function in mouse models of endotoxin-induced uveitis (EIU).29) Bilberry extract has also been used
in clinical trials to treat eye fatigue induced by acute video display terminal (VDT) loads.30) We
previously reported that bilberry extract has a neuroprotective effect against retinal neuronal damage induced by N-methyl-D-aspartic acid in mice.31) We also demonstrated that bilberry
ex-tract inhibited angiogenesis in a mouse model of oxygen-induced retinopathy.32) Moreover, in in
vitro models, bilberry extract exhibited protec-tive effects against photoreceptor cell damage induced by UV-A irradiation or blue LED light ex-posure.9, 33, 34) However, there are limited studies
addressing the pharmacological effects of bilberry extract on the ocular surface.
In this study, we demonstrated that bilberry extract exhibited protective effects against blue LED light-induced corneal epithelial cell dam-age in an in vitro model. There have been few reports of protective agents against corneal cell damage caused by blue LED light exposure. Since blue LED light is increasingly used in common everyday applications, the search for compounds that prevent blue LED light-induced cell damage would be beneficial for maintaining the health of corneal epithelial cells.
II. Materials and methods 1. Materials
Bilberry extract ( VMA 25/37 (Lot: B0216- 09302) sold by Omnica Co., (Tokyo, Japan)) was supplied by Wakasa Seikatsu Co., Ltd (Kyoto, Ja-pan). Bilberry extract was extracted by extracting the relevant compounds from fruits with ethanol and then removing the solvent by concentration.
The rate of aglycone in the bilberry extract used in the experiment is attached. (Appendix 1). From the HPLC analysis chart, the type of antho-cyanin was identified based on the retention time of each peak.23) Furthermore, the content was
cal-culated from the peak area of HPLC and the mo-lecular weight of each anthocyanin. In this study, among the anthocyanins that can be obtained as standard products, experiments were conducted with three types of anthocyanins, starting from the one with the highest proportion of glucoside in the extract used in the experiment.
Delphinidin-3-glucoside (D3G), cyanidin-3-glu-coside (C3G), and malvidin-3-glucyanidin-3-glu-coside (M3G) were purchased from Tokiwa Phytochemical Co., Ltd. (Chiba, Japan). Hoechst 33342, prop-idium iodide (PI), 5-(and-6)-chloromethyl-2,7-dichlorodihydrofluorescein diacetate acetyl ester
(CM-H2DCFDA) and the JC-1 Mitochondrial Membrane Potential Assay Kit were purchased from Thermo Fisher Scientific Inc. (Waltham, MA, USA). N-Acetyl-L-cysteine (NAC) was pur-chased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). The Cell Counting Kit-8 (CCK-8) was obtained from Dojindo Laboratories (Kumamoto, Japan). The Caspase-Glo® 3/7 Assay
was purchased from Promega Co. (Madison, WI, USA). Antibodies directed against the following proteins were obtained from Cell Signaling Tech-nology (Beverly, MA, USA): rabbit phosphorylated p38 mitogen-activated protein kinase (MAPK), rabbit total p38 MAPK, rabbit phosphorylated extracellular signal-regulated kinase (ERK) and rabbit total ERK. Antibody directed against mouse β-actin was purchased from Sigma-Aldrich (St. Louis, MO, USA). SV-40 transformed human corneal epithelial cells (HCE-T; RCB No. 2280) were obtained from RIKEN BioResource Center Cell Bank (Tsukuba, Ibaraki, Japan).
2. Cell culture
HCE-T cells were maintained in Dulbeccos modified Eagles Medium (DMEM)/F-12 (Wako, Osaka, Japan) supplemented with 0.5% dimethyl sulfoxide, 5 µg/mL insulin, 2.75 µg/mL transfer-rin, 3.35 ng/mL selenium, 10 ng/mL recombinant human epidermal growth factor, and 5% fetal bovine serum (FBS). Cells were cultured at 37°C in a humidified atmosphere of 5% CO2. Cell
pas-sage was performed by trypsinization every 2 or 3 days.
3. Exposure of HCE-T cells to blue LED light in vitro
The Blue LED light device has aligning many LED light sources. The device conditions were ap-plied according to previous reports.8, 34–36) In this
with blue LED light at 2,000 lux illuminances. Blue LED light exposure was performed in a CO2
incubator under a humidified atmosphere of 5% CO2 at 37°C.
1) Test reagent application conditions
HCE-T cells were seeded at a density of 1×104 cells/100 µL on 96-well plates and cultured at 37°C for 24 h. The medium was then exchanged for 1% FBS-DMEM/Hams F12. After 30 min, the cells were treated with bilberry extract, D3G, C3G, M3G, or NAC, respectively. NAC is a posi-tive control that has already been reported to sup-press blue LED light-induced retinal photorecep-tor cell damage.8) The cells were then incubated
for 1 h and subsequently exposed to 2,000 lux of blue LED light for 24 h.
2) Blue LED light-induced cell death assay At the end of the incubation period, Hoechst 33342 (λex=360 nm; λem>490 nm) and PI (λex=
535 nm; λem>617 nm) were added to the culture
medium (16.2 and 1.5 μM, respectively) and then the cells were incubated for 30 min. Next, the cells were photographed through fluorescence filters (Olympus Co., Tokyo, Japan) for Hoechst 33342 (U-MWU) and PI (U-MWIG) with a charge-coupled device camera (DP30BW; Olympus Co., Tokyo, Japan). The cell mortality rate was calculated from the number of PI-positive cells as a percentage of the number of Hoechest 33342-positive cells. 3) Measurement of cellular ROS production
fol-lowing blue LED light exposure
Intracellular ROS production induced by blue LED light exposure of HCE-T cells was deter-mined using CM-H2DCFDA. At the end of the
blue LED light exposure period, CM-H2DCFDA
was added to the culture medium at a final con-centration of 10 μM and the cells were incubated at 37°C for 1 h. The 96-well plate was then loaded into a plate holder in a fluorescence spectropho-tometer. The reaction was performed at 37°C, and
fluorescence was measured at 488 nm excitation and 525 nm emission. The number of cells was de-termined by Hoechst 33342 staining and used to calculate ROS production per cell.37)
4) Measurement of cell viability following blue LED light exposure
The cell viability was measured using the CCK-8 assay for 2 h and the absorbance was measured at 492 nm (reference wavelength, 660 nm) using a spectrophotometer (Varioskan; Thermo Fisher Scientific, Waltham, MA, USA).
5) Mitochondrial membrane potential assay The mitochondrial membrane potential was measured with a JC-1 Mitochondrial Membrane Potential Assay Kit according to the manufactur-er s protocol. Aftmanufactur-er blue LED light exposure, next, the cells were incubated with 10 µg/mL JC-1 at 37°C for 15 min. Images were collected using a flu-orescence microscope (BZ-X710; Keyence, Osaka, Japan), which detects cells mainly containing JC-1 aggregates (excitation/emission=540/605 nm) and apoptotic cells mainly containing JC-1 mono-mers (excitation/emission=480/510 nm).
6) Caspase-3/7 activation assay
3/7 was measured with a Caspase-Glo®3/7 Assay according to the manufacturer s
instructions. After the blue LED light exposure, Caspase-Glo® 3/7 Reagent was added at a 1 : 1
ratio to the cell sample volume and the cells were incubated for 1 h at 37°C. The luminescence of each sample was measured with a microplate reader (Varioskan Flash 2.4; Thermo Fisher Sci-entific., Waltham, MA, USA).
4. Immunoblot analysis
After the light exposure, the cells were washed with phosphate buffered saline, and lysed using radioimmunoprecipitation assay (RIPA) buffer con-taining 1% protease inhibitor and phosphatase in-hibitor cocktails 2 and 3 (Sigma-Aldrich, St. Louis,
MO, USA). The lysates were then centrifuged at 12,000g for 20 min. Protein concentrations were determined using a BCA Protein Assay Kit (Ther-mo Fisher Scientific., Waltham, MA, USA) with bovine serum albumin as the standard. An equal volume of protein sample and sample buffer was mixed, separated on 5–20% SDS-polyacrylamide gels, and transferred to polyvinylidene difluoride membranes (Immobilon-P; EMD Millipore Corpo-ration, Billerica, MA, USA). For immunoblotting experiments, the following primary antibodies were used: rabbit anti-phospho-p38 MAPK, rab-bit anti-p38 MAPK, rabrab-bit anti-phospho-ERK, rabbit anti- ERK (all used at a 1 : 1000 dilution) and mouse anti-β-actin (used at a 1 : 5000 dilu-tion). Subsequently, the membranes were incu-bated with secondary antibodies: horseradish peroxidase (HRP)-conjugated goat anti-rabbit or goat anti-mouse (both used at a 1 : 2000 dilution; Thermo Fisher Scientific., Waltham, MA, USA). Immunoreactive bands were visualized using Im-munoStar® LD (Wako Pure Chemical Industries,
Osaka, Japan) and an LAS-4000 Luminescent Im-age Analyzer (Fuji Film Co., Ltd., Tokyo, Japan). β-actin was used as a loading control.
5. Statistical analysis
Data are presented as the mean±S.E.M. Sta-tistical comparisons were made using one-way analysis of variance followed by Student s t-tests, Tukeys tests or Dunnett s Multiple Comparison tests. A value of p<0.05 was considered to indi-cate statistical significance.
III. Results
1. Effects of bilberry extract, D3G, C3G, or M3G on blue LED light-induced cell death of hu-man corneal epithelial cells
Firstly, we investigated the effects of bilberry extract, anthocyanins or NAC on blue LED
light-induced human corneal epithelial cell death. We counted the number of cells exhibiting PI fluores-cence and expressed the death rate relative to the number of cells that showed Hoechst 33342 fluo-rescence. The cell damage induced by blue LED light exposure was inhibited by the addition of NAC (a positive control: 1mM). NAC (1mM) also suppressed corneal epithelial cell damage due to blue LED light damage as well as retinal photo-receptor damage. Furthermore, bilberry extract, D3G or C3G at 3–10 μM significantly inhibited HCE-T cell death in a concentration-dependent manner (Figure 1). On the other hands, M3G had no significant effect on blue LED light-induced cell death in HCE-T cells. In addition, it has been confirmed by single administration that each extract and compound does not show cytotoxic-ity at experimental concentrations (Figure 1B–D; showed on right end of each graph).
2. Bilberry extract, D3G or C3G each reduced the levels of blue LED light-induced ROS in HCE-T cells
In many previous reports, it has been suggested that one of the main causes of cytotoxicity induced by blue LED light exposure is intracellular ROS accumulation.3, 8, 10, 38) Thus, we examined blue
LED light-induced ROS production in HCE-T cells grown in the presence of bilberry extract or its an-thocyanin constituents. The levels of intracellular ROS, which were measured by the conversion of CM-H2DCFDA to a fluorescent product upon ex-posure to ROS, were increased by blue LED light exposure. In addition, intercellular ROS produc-tion was increased by blue LED light exposure. Bilberry extract, D3G or C3G each significantly reduced the blue LED light-induced ROS produc-tion in HCE-T cells. In contrast, M3G had no significant effect on blue LED light-induced ROS production in HCE-T cells (Figure 2).
3. The protective effects of bilberry extract, D3G or C3G against reduced cellular dehy-drogenase activity induced by blue LED light exposure
We investigated the effects of bilberry extract,
D3G, C3G or M3G on the blue LED light-induced reduction in cellular dehydrogenase activity in HCE-T cells. Cell dehydrogenase activity is the result of measuring dehydrogenase activity in living cells using CCK-8.9) We found that blue
Fig. 1 Effects of bilberry extract or its anthocyanin constituents on blue LED light-induced cell death in HCE-T cells
HCE-T cells were preincubated in the (A) absence or in the presence of (B), bilberry extract, (C), D3G, (D), C3G, or (E), M3G prior to blue LED light exposure for 24 h. (A–E) shows the representative fluorescence mi-croscopy of Hoechest 33342 and PI in the cells. The number of cells exhibiting PI fluorescence was counted and positive cells were expressed as the percentage of PI-positive to Hoechst 33342-positive cells. Data are represented as the mean±S.E.M. (n=5 or 6). ##p<0.01 vs. control; **p<0.01, *p<0.05 vs. the vehicle (Dun-nett s Multiple Comparison test or Student s t-test). The scale bar represents 50 µm. C, control; V, vehicle; D3G, delphinidin 3-glucoside; C3G, cyanidin 3-glucoside; M3G, malvidin 3-glucoside; N,N-acetyl-L-cysteine.
LED light exposure induced a significant reduc-tion in cell dehydrogenase activity. Preincubareduc-tion of the cells with bilberry extract (10 µg/mL), D3G (10 μM) or C3G (3 and 10 μM) significantly pre-vented the reduction in cellular dehydrogenase activity induced by blue LED light, in contrast, M3G had no significant effect (Figure 3). NAC also suppressed corneal epithelial cell damage due to blue LED light damage, and we confirmed by single administration that bilberry extract did not show cytotoxicity at the concentrations we tested (Figure 3A).
4. The protective effects of bilberry extract on blue LED light-mediated perturbation of mi-tochondrial membrane potential
We investigated the effects of bilberry extract on blue LED light-induced changes in mitochon-drial membrane potential in HCE-T cells. It has been reported that blue LED light exposure per-turbs the mitochondrial membrane potential in vitro in a blue LED light-induced retinal degen-eration model.8) Therefore, we investigated the
mitochondrial membrane potential in HCE-T cells exposed to blue LED light. JC-1 dye was used for Fig. 2 Effects of bilberry extract or its anthocyanin constituents on blue LED light-induced ROS production in
HCE-T cells
HCE-T cells were in the presence of (A), bilberry extract, (B), D3G, (C), C3G or (D), M3G prior to blue LED light exposure for 24 h. Intracellular ROS levels were then determined by measuring the fluorescence of CM-H2DCFDA. Data are represented as the mean±S.E.M. (n=6 or 12). ##p<0.01 vs. control; **p<0.01 vs. the vehicle (Dunnett s Multiple Comparison test or Student s t-test). C, control; V, vehicle; D3G, delph-inidin 3-glucoside; C3G, cyanidin 3-glucoside; M3G, malvidin 3-glucoside; N,N-acetyl-L-cysteine. N-acetyl-L
this study. In cells with high JC-1 aggregates, aggregates of JC-1 (red) accumulate and in apop-totic cells monomers of JC-1 (green) accumulate. Control cells were stained red. Blue LED light exposure increased the number of apoptotic cells, and 1 h of pretreatment with 10 µg/mL of bilberry
extract significantly increased the ratio of red-stained, healthy cells (Figure 4A).
Fig. 4 Effects of bilberry extract on blue LED light-induced mitochondrial dysfunction and cas-pase-3/7 activation
HCE-T cells were in the presence of bilberry extract followed by exposure to blue LED light for 1 h. Next, mitochondrial dysfunction was assessed using the JC-1 Mitochondrial Membrane Potential Assay Kit. (A) Repre-sentative images showing JC-1 stained cells. The healthy cells mainly contain JC-1 ag-gregates (red) as indicated by the arrowhead and apoptotic or unhealthy cells mainly con-tain JC-1 monomers (green). Quantitative analysis revealed the ratio of red fluorescence (Hoechest 33342) in whole cells. (B) HCE-T cells were preincubated with bilberry extract followed by exposure to blue LED light for 1 h. Next, caspase-3/7 activity was monitored using the Caspase-Glo® 3/7 Assay. Data are
represented as the mean±S.E.M. (n=5 or 6). ##p<0.01 vs. control; **p<0.01, *p<0.05 vs. the vehicle (Student s t-test). The scale bar represents 50 µm. C, control; V, vehicle; Bil-berry, bilberry extract (10 µg/mL).
Fig. 3 Effects of bilberry extract or its anthocyanin constituents on blue LED light-induced HCE-T cell dehydrogenation
HCE-T cells were in the presence of (A) bil-berry extract or (B) D3G, C3G or M3G and cell viability was assessed by immersing the cells in CCK-8 for 2 h at 37°C. Data are repre-sented as the mean±S.E.M. (n=6). ##p<0.01 vs. control; **p<0.01, *p<0.05 vs. the vehicle (Dunnett s Multiple Comparison test or Stu-dent s t-test). C, control; V, vehicle; Bilberry ext., bilberry extract; D3G, delphinidin 3-glu-coside; C3G, cyanidin 3-glu3-glu-coside; M3G, mal-vidin 3-glucoside; NAC, N-acetyl-L-cysteine.
5. Bilberry extract inhibited the blue LED light-induced activation of caspase-3/7
We evaluated the caspase-3/7 activity using the Caspase-Glo® 3/7 Assay. Our results showed that
the caspase-3/7 activity was increased upon blue
LED light exposure and prior incubation with 10 µg/mL of bilberry extract significantly pre-vented the blue LED light-induced activation of caspase-3/7 (Figure 4B).
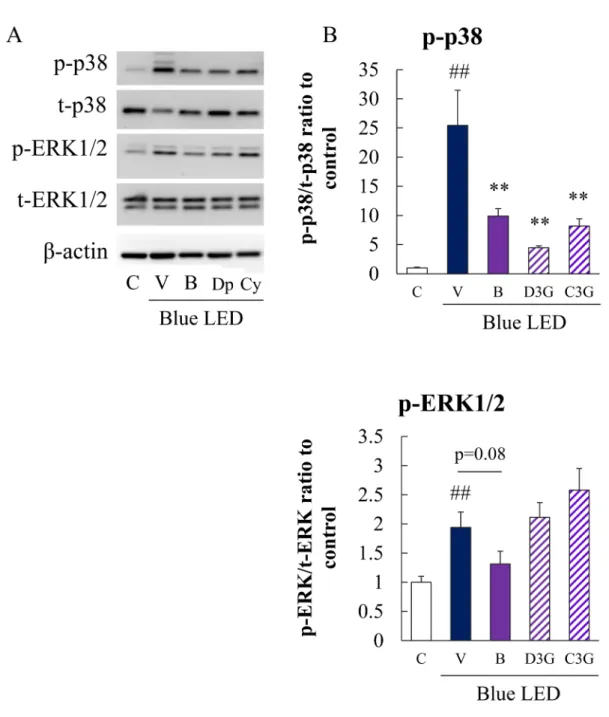
Fig. 5 Effects of bilberry extract or its anthocyanin constituents on blue LED light-induced activation of p38 MAPK and ERK 1/2 in HCE-T cells
HCE-T cells were in the presence of bilberry extract or its anthocyanin constituents followed by exposure to blue LED light for 24 h. Next, protein lysates were analyzed by immunoblotting using antibodies directed against the corresponding proteins. (A) Activation of p38 MAPK and ERK 1/2 was detected by immunoblot-ting. (B) Quantitative analysis of phospho-p38 MAPK and phospho-ERK1/2 protein levels. Data are repre-sented as the mean±S.E.M. (n=6 or 9). ##p<0.01 vs. control; **p<0.01, *p<0.05 vs. the vehicle (Tukey s tests or Student s t-test). C, control; V, vehicle; B, bilberry extract 10 µg/mL ; Dp and D3G, delphinidin 3-glu-coside 10μM; Cy and C3G, cyanidin 3-glu3-glu-coside 10μM.
6. Bilberry extract, D3G or C3G each inhibited blue LED light-induced phosphorylation of p38 in HCE-T cells
We performed immunoblot analysis to investi-gate the effects of bilberry extract or its anthocy-anin constituents on the phosphorylation of stress response proteins. Blue LED light exposure in-creased the phosphorylation of p38 and ERK 1/2. Bilberry extract, D3G or C3G each significantly inhibited p38 phosphorylation induced by blue LED light exposure. Additionally, bilberry extract inhibited the phosphorylation of ERK 1/2 induced by blue LED light exposure. In contrast, D3G or C3G had no effect on the phosphorylation status of ERK 1/2 (Figure 5). In addition, C3G tended to increase phosphorylation of ERK 1/2 (no signifi-cant difference).
IV. Discussion
The ocular surface, which consists of the tear film, conjunctiva and cornea, functions as a free radical barrier. Dry eye is the main eye disorder related to a deficiency in antioxidant defense by the tear film.38–40) In dry eye patients with
Sjögren syndrome, oxidative stress markers in-crease and correlate with severity.23, 41) Moreover,
useful treatments for dry eye include antioxidant
therapy, which suppresses inflammation in exper-imental and clinical cases of dry eye.42–47)
In this study, we investigated the protective effects of bilberry extract or its anthocyanin constituents against blue LED light-induced oxi-dative stress in human corneal epithelial cells. Bilberry extract, which contains anthocyanins, other polyphenols and flavonoids is known to have antioxidant and anti-inflammatory effects, which are associated with both a reduction in ROS and an increase in antioxidant enzymes.48) In a
previ-ous report, bilberry extract or its anthocyanin constituents suppressed murine photoreceptor cell damage induced by blue LED light exposure. This suppression was mediated by the attenu-ation of ROS production and regulattenu-ation of both p38 MAPK activation and caspase-3/7 activity.9)
We investigated whether blue LED light exposure induces cell damage in corneal epithelial cells and whether bilberry extract exhibits protective effects against this damage. It is known that blue LED light exposure induces damage to both retinal photoreceptor cells and corneal epithelial cells.3, 4, 8–10, 34–36) Since bilberry extract exhibits
protective effects for both cell types, we specu-lated that bilberry extract and respecu-lated compounds may be promising treatments for eye diseases in-Table 1 Analysis result of bilberry extract
Blueberry (comp % m/m) Blueberry dried (comp % m/m) Extracted Bilberry dried Water 84.6 <0.5 4.2 Protein 0.6 4.22 1.5 Fat 0.6 4.22 0.6 Sugar 6.05 42.5 0.1
Carbohydrates & Fiber 4.9 34.4 37.4
org Acids 1.37 9.63 —
Titrable Acids (as Citric acid) — — 2.9
Minerals 0.3 2.11
Ash — — 0.2
Anthocyanins 0.4 2.81 37
Phenols other than Anthos** — — 16
duced by blue LED light exposure.
First, in this study, blue LED light was directly irradiated from below the cell. Thereby, it is considered that light absorption by the drug can be ignored. Blue LED light exposure generated HCE-T cell damage and increased intracellular ROS production (Figures 1–3). The cell dysfunc-tion induced by blue LED light exposure was suppressed by bilberry extract, D3G, C3G or the positive control compound, NAC. It has been re-ported that these drugs not only have direct free radical scavenging activity but also suppress ROS production.31) These compounds also increase the
amount of intracellular antioxidant enzymes.48)
On the other hand, bilberry extract (10 µg/mL), D3G (10 μM) or C3G (10 μM) each significantly inhibited ROS production induced by blue LED light exposure. In the cell death assay using both PI fluorescence and Hoechst 33342 fluorescence, all of these compounds showed cytoprotective effects at concentrations lower than the concen-tration (bilberry extract (3 µg/mL), D3G (3 μM) or C3G (3 μM)) at which ROS production was inhibited. On the other hand, M3G did not sup-press corneal epithelial cell damage caused by blue LED irradiation (Figures 1 and 3). This is considered to be due to the low ability of M3G to suppress ROS production. Compared to D3G and C3G, M3G has a larger number of methoxy groups (-OCH3) and a smaller number of hydroxyl
groups (-OH).23) In a previous report, it has been
found that the radical scavenging effect increases as the number of coordinated hydroxyl groups increases.49) M3G also suppressed the amount
of ROS produced by blue LED irradiation, but it was not significant (Figure 2). This suggests that M3G did not show a cyto-protective effect. Conse-quently, our data indicated that these compounds possess HCE-T cell protective activity against blue LED light exposure by additional
mecha-nisms other than free radical scavenging activity. One hypothesis is that these compounds suppress the unfolded protein response (UPR). A previous report has shown that bilberry extract exhibited protective effects against blue LED light-induced UPR in retinal photoreceptor cells.34)
Further-more, bilberry extract exhibited protective effects against the optic nerve crush-induced UPR in retinal ganglion cells.50) Further studies are
re-quired to understand the mechanism of UPR sup-pression mediated by bilberry extract.
In the present study, blue LED light exposure induced caspase-3/7 activation and disrupted the mitochondrial membrane potential, which cor-related with HCE-T cell death (Figure 4). We also investigated cell damage using the CCK-8 assay, which reflects mitochondrial function (Figure 3). It has been reported that disturbance of the mitochondrial membrane potential by UV light exposure is the cause of caspase-3/7 activation in the corneal epithelial cell line HCE-T.51) Blue
LED light also induces mitochondrial damage in corneal epithelial cells. It has been observed that caspase-3/7 activation occurs through the release of cytochrome c from mitochondria.52) Bilberry
extract attenuated the mitochondrial dysfunction and suppressed the activation of caspase-3/7 (Fig-ure 4), both of which were induced by blue LED light exposure.
ROS induces MAPK activation, and MAPK modulates inflammation, cell death and other cellular processes.53) It has been shown that
vis-ible light exposure activated p38 MAPK in pho-toreceptor cells,8, 54) and it was revealed that blue
LED light exposure also activated p38 MAPK in corneal epithelial cells in this study. In addition, it has been reported that p38 MAPK is an apop-tosis-related factor, which is mainly activated by superoxide anions under oxidative stress condi-tions.55) The superoxide anion radical scavenging
activity of bilberry extract56) may contribute to
the suppression of p38 MAPK activation induced by blue LED light. Corneal epithelial cells are sensitive to oxidative stress associated with lipid peroxidation.57) UV-B, 4-hydroxynonenal (a
reac-tive lipid peroxidation end product) and osmotic pressure activated p38 MAPK and ERK 1/2.57–59)
Activation of ERK 1/2 has been shown to be in-volved in corneal epithelial wound healing.60)
Ex-posure of HCE-T cells to blue LED light induced the activation of ERK 1/2, but this activation was not suppressed by the addition of bilberry extract, D3G or C3G. Bilberry extract inhibited ROS pro-duction induced by blue LED light exposure and suppressed apoptotic signaling, as examined by the phosphorylation status of p38. Interestingly, bilberry extract did not affect the activation of ERK 1/2, which is involved in corneal epithelial cell wound healing. Thus, these findings indicat-ed that the protective effects of bilberry extract or its anthocyanin constituents on HCE-T cells ex-posed to blue LED light were independent of the ERK pathway.
Furthermore, we considered that the involve-ment of components other than anthocyanin contained in bilberry extract. We showed the analysis data of general bilberry extract to the Supplemental information 2. We have confirmed that bilberry extract contains dietary fiber, car-bohydrates and some polyphenols as components other than anthocyanins. We considered that the effect was small because each of the other poly-phenols was a minor amount (About one thou-sandth). In addition, we considered that the effect of bilberry extract was anthocyanin and exam-ined its action, as we thought that dietary fiber and carbohydrate also had little effect on ROS scavenging ability.
In conclusion, the present study showed that exposure of HCE-T cells to blue LED light
in-duced ROS production, which resulted in corneal epithelial cell damage. We also found that bilber-ry extract or its anthocyanin constituents, D3G or C3G exhibited protective effects against blue LED light-induced HCE-T cell damage.
Conflict of interest
Dr. Hideaki Hara received financial support from Wakasa Seikatsu Co., Ltd (Kyoto, Japan) as part of a collaborative research project.
References
1) Glazer-Hockstein C, Dunaief JL: Could Blue Light– Blocking Lenses Decrease the Risk of Age-Related Macular Degeneration? Retina, 26(1), 1–4, 2006. 2) Margrain T, Boulton M, Marshall J, Sliney DH:
Do blue light filters confer protection against age-related macular degeneration? Prog. Retin. Eye Res., 23(5), 523–531, 2004.
3) Lee HS, Cui L, Li Y, Choi JS, Choi J-H, Li Z, Kim GE, Choi W, Yoon KC: Influence of Light Emitting Diode-Derived Blue Light Overexposure on Mouse Ocular Surface. PLoS One, 11(8), e0161041, 2016. 4) Lee J-B, Kim S-H, Lee S-C, Kim H-G, Ahn H-G, Li
Z, Yoon KC: Blue light-induced oxidative stress in human corneal epithelial cells: protective effects of ethanol extracts of various medicinal plant mixtures.
Invest. Ophthalmol. Vis. Sci., 55(7), 4119–4127,
2014.
5) Grimm C, Wenzel A, Williams T, Rol P, Hafezi F, Remé C: Rhodopsin-mediated blue-light damage to the rat retina: effect of photoreversal of bleaching.
Invest. Ophthalmol. Vis. Sci., 42(2), 497–505, 2001.
6) Roehlecke C, Schumann U, Ader M, Knels L, Funk RHW: Influence of blue light on photoreceptors in a live retinal explant system. Mol. Vis., 17, 876–884, 2011.
7) Behar-Cohen F, Martinsons C, Viénot F, Zissis G, Barlier-Salsi A, Cesarini JP, Enouf O, Garcia M, Picaud S, Attia D: Light-emitting diodes (LED) for domestic lighting: Any risks for the eye? Prog. Retin.
Eye Res., 30(4), 239–257, 2011.
8) Kuse Y, Ogawa K, Tsuruma K, Shimazawa M, Hara H: Damage of photoreceptor-derived cells in culture induced by light emitting diode-derived blue light.
Sci. Rep., 4(1), 5223, 2015.
Shima-zawa M, Hara H: Protective effects of bilberry and lingonberry extracts against blue light-emitting di-ode light-induced retinal photoreceptor cell damage in vitro. BMC Complement. Altern. Med., 14(1), 120, 2014.
10) Nakamura M, Kuse Y, Tsuruma K, Shimazawa M, Hara H: The Involvement of the Oxidative Stress in Murine Blue LED Light-Induced Retinal Damage Model. Biol. Pharm. Bull., 40(8), 1219–1225, 2017. 11) Sanvicens N, Gómez-Vicente V, Masip I, Messeguer
A, Cotter TG: Oxidative Stress-induced Apoptosis in Retinal Photoreceptor Cells Is Mediated by Calpains and Caspases and Blocked by the Oxygen Radical Scavenger CR-6. J. Biol. Chem., 279(38), 39268– 39278, 2004.
12) Cai J, Nelson KC, Wu M, Sternberg P Jr., Jones DP: Oxidative damage and protection of the RPE. Prog.
Retin. Eye Res., 19(2), 205–221, 2000.
13) Kennedy M, Kim KH, Harten B, Brown J, Planck S, Meshul C, Edelhauser H, Rosenbaum JT, Armstrong CA, Ansel JC: Ultraviolet irradiation induces the production of multiple cytokines by human corneal cells. Invest. Ophthalmol. Vis. Sci., 38(12), 2483– 2491, 1997.
14) Moulton EA, Becerra L, Borsook D: An fMRI case report of photophobia: Activation of the trigeminal nociceptive pathway. Pain, 145(3), 358–363, 2009. 15) Estil S, Olsen WM, Huitfeldt HS, Haaskjold E:
UVB-induced formation of (6-4) photoproducts in the rat corneal epithelium. Acta Ophthalmol. Scand., 75(2), 120–123, 2009.
16) Cullen AP: Photokeratitis and Other Phototoxic Ef-fects on the Cornea and Conjunctiva. Int. J. Toxicol., 21(6), 455–464, 2002.
17) Andley UP, Fritz C, Morrison AR, Becker B: The role of prostaglandins E2 and F2 alpha in ultraviolet radiation-induced cortical cataracts in vivo. Invest.
Ophthalmol. Vis. Sci., 37(8), 1539–1548, 1996.
18) Cejková J, Stípek S, Crkovská J, Ardan T, Pláteník J, Cejka C, Midelfart A: UV Rays, the prooxidant/an-tioxidant imbalance in the cornea and oxidative eye damage. Physiol. Res., 53(1), 1–10, 2004.
19) Cejková J, Stípek S, Crkovská J, Ardan T, Midelfart A: Reactive oxygen species (ROS)-generating oxidas-es in the normal rabbit cornea and their involvement in the corneal damage evoked by UVB rays. Histol.
Histopathol., 16(2), 523–533, 2001.
20) Rogers CS, Chan L-M, Sims YS, Byrd KD, Hinton DL, Twining SS: The effects of sub-solar levels of UV-A and UV-B on rabbit corneal and lens epithelial
cells. Exp. Eye Res., 78(5), 1007–1014, 2004.
21) Baj A, Bombardelli E, Gabetta B, Martinelli EM: Qualitative and quantitative evaluation of vaccinium myrtillus anthocyanins by high-resolution gas chro-matography and high-performance liquid chromatog-raphy. J. Chromatogr. A, 279, 365–372, 1983. 22) Nakajima J, Tanaka I, Seo S, Yamazaki M, Saito
K: LC/PDA/ESI-MS Profiling and Radical Scaveng-ing Activity of Anthocyanins in Various Berries. J.
Biomed. Biotechnol., 2004(5), 241–247, 2004.
23) Ogawa K, Sakakibara H, Iwata R, Ishii T, Sato T, Goda T, Shimoi K, Kumazawa S: Anthocyanin Com-position and Antioxidant Activity of the Crowberry (Empetrum nigrum) and Other Berries. J. Agric.
Food Chem., 56(12), 4457–4462, 2008.
24) Salvayre R, Nègre A, Affany A, Lenoble M, Douste-Blazy L: Protective effect of plant flavonoids, analogs and vitamin E against lipid peroxidation of mem-branes. Prog. Clin. Biol. Res., 280, 313–316, 1988. 25) Viljanen K, Kylli P, Kivikari R, Heinonen M:
Inhibi-tion of Protein and Lipid OxidaInhibi-tion in Liposomes by Berry Phenolics. J. Agric. Food Chem., 52(24), 7419–7424, 2004.
26) Viljanen K, Kylli P, Hubbermann E-M, Schwarz K, Heinonen M: Anthocyanin Antioxidant Activity and Partition Behavior in Whey Protein Emulsion. J.
Ag-ric. Food Chem., 53(6), 2022–2027, 2005.
27) Kähkönen MP, Heinonen M: Antioxidant Activity of Anthocyanins and Their Aglycons. J. Agric. Food
Chem., 51(3), 628–633, 2003.
28) Head KA: Natural therapies for ocular disorders, part two: cataracts and glaucoma. Altern. Med. Rev., 6(2), 141–166, 2001.
29) Miyake S, Takahashi N, Sasaki M, Kobayashi S, Tsubota K, Ozawa Y: Vision preservation during retinal inflammation by anthocyanin-rich bilberry extract: cellular and molecular mechanism. Lab.
In-vest., 92(1), 102–109, 2012.
30) Ozawa Y, Kawashima M, Inoue S, Inagaki E, Suzuki A, Ooe E, Kobayashi S, Tsubota K: Bilberry extract supplementation for preventing eye fatigue in video display terminal workers. J. Nutr. Health Aging, 19(5), 548–554, 2015.
31) Matsunaga N, Imai S, Inokuchi Y, Shimazawa M, Yokota S, Araki Y, Hara H: Bilberry and its main constituents have neuroprotective effects against retinal neuronal damage in vitro and in vivo. Mol.
Nutr. Food Res., 53(7), 869–877, 2009.
32) Matsunaga N, Tsuruma K, Shimazawa M, Yokota S, Hara H: Inhibitory actions of bilberry
anthocy-anidins on angiogenesis. Phytother. Res., 24(S1) (Suppl 1), S42–S47, 2010.
33) Ogawa K, Tsuruma K, Tanaka J, Kakino M, Ko-bayashi S, Shimazawa M, Hara H: The Protective Effects of Bilberry and Lingonberry Extracts against UV Light-Induced Retinal Photoreceptor Cell Dam-age in Vitro. J. Agric. Food Chem., 61(43), 10345– 10353, 2013.
34) Ooe E, Kuse Y, Yako T, Sogon T, Nakamura S, Hara H, Shimazawa M: Bilberry extract and anthocyanins suppress unfolded protein response induced by expo-sure to blue LED light of cells in photoreceptor cell line. Mol. Vis., 24, 621–632, 2018.
35) Ooe E, Tsuruma K, Kuse Y, Kobayashi S, Shima-zawa M, Hara H: The involvement of ATF4 and S-opsin in retinal photoreceptor cell damage induced by blue LED light. Mol. Vis., 23, 52–59, 2017.
36) Hiromoto K, Kuse Y, Tsuruma K, Tadokoro N, Kaneko N, Shimazawa M, Hara H: Colored lenses suppress blue light-emitting diode light-induced damage in photoreceptor-derived cells. J. Biomed.
Opt., 21(3), 035004, 2016.
37) Tanaka J, Nakanishi T, Ogawa K, Tsuruma K, Shimazawa M, Shimoda H, Hara H: Purple Rice Ex-tract and Anthocyanidins of the Constituents Protect against Light-Induced Retinal Damage in Vitro and in Vivo. J. Agric. Food Chem., 59(2), 528–536, 2011. 38) Choy CKM, Cho P, Benzie IFF: Antioxidant Content
and Ultraviolet Absorption Characteristics of Hu-man Tears. Optom. Vis. Sci., 88(4), 507–511, 2011. 39) Uchino Y, Kawakita T, Miyazawa M, Ishii T,
Onou-chi H, Yasuda K, Ogawa Y, Shimmura S, Ishii N, Tsubota K: Oxidative stress induced inflammation initiates functional decline of tear production. PLoS
One, 7(10), e45805, 2012.
40) Uchino Y, Kawakita T, Ishii T, Ishii N, Tsubota K: A new mouse model of dry eye disease: oxidative stress affects functional decline in the lacrimal gland.
Cor-nea, 31(Suppl 1.), S63–S67, 2012.
41) Wakamatsu TH, Dogru M, Matsumoto Y, Kojima T, Kaido M, Ibrahim OMA, Sato EA, Igarashi A, Ichi-hashi Y, Satake Y, Shimazaki J, Tsubota K: Evalu-ation of lipid oxidative stress status in Sjögren syn-drome patients. Invest. Ophthalmol. Vis. Sci., 54(1), 201–210, 2013.
42) Järvinen RL, Larmo PS, Setälä NL, Yang B, Engb-lom JR, Viitanen MH, Kallio HP: Effects of oral sea buckthorn oil on tear film Fatty acids in individuals with dry eye. Cornea, 30(9), 1013–1019, 2011.
43) Cavet ME, Harrington KL, Vollmer TR, Ward KW,
Zhang J-Z: Anti-inflammatory and anti-oxidative effects of the green tea polyphenol epigallocatechin gallate in human corneal epithelial cells. Mol. Vis., 17, 533–542, 2011.
44) Gillespie K, Kodani I, Dickinson DP, Ogbureke KUE, Camba AM, Wu M, Looney S, Chu TC, Qin H, Bisch F, Sharawy M, Schuster GS, Hsu SD: Effects of oral consumption of the green tea polyphenol EGCG in a murine model for human Sjogren s syndrome, an autoimmune disease. Life Sci., 83(17-18), 581–588, 2008.
45) Li Z, Choi J-H, Oh H-J, Park S-H, Lee J-B, Yoon KC: Effects of eye drops containing a mixture of omega-3 essential fatty acids and hyaluronic acid on the ocu-lar surface in desiccating stress-induced murine dry eye. Curr. Eye Res., 39(9), 871–878, 2014.
46) Kangari H, Eftekhari MH, Sardari S, Hashemi H, Salamzadeh J, Ghassemi-Broumand M, Khabazk-hoob M: Short-term consumption of oral omega-3 and dry eye syndrome. Ophthalmology, 120(11), 2191–2196, 2013.
47) Brignole-Baudouin F, Baudouin C, Aragona P, Rolando M, Labetoulle M, Pisella PJ, Barabino S, Siou-Mermet R, Creuzot-Garcher C: A multicentre, double-masked, randomized, controlled trial assess-ing the effect of oral supplementation of omega-3 and omega-6 fatty acids on a conjunctival inflammatory marker in dry eye patients. Acta Ophthalmol., 89(7), e591–e597, 2011.
48) Milbury PE, Graf B, Curran-Celentano JM, Blum-berg JB: Bilberry (Vaccinium myrtillus) antho-cyanins modulate heme oxygenase-1 and glutathione S-transferase-pi expression in ARPE-19 cells. Invest.
Ophthalmol. Vis. Sci., 48(5), 2343–2349, 2007.
49) Noda Y, Kaneyuki T, Mori A, Packer L: Antioxidant activities of pomegranate fruit extract and its antho-cyanidins: delphinidin, cyanidin, and pelargonidin.
J. Agric. Food Chem., 50(1), 166–171, 2002.
50) Nakamura O, Moritoh S, Sato K, Maekawa S, Mu-rayama N, Himori N, Omodaka K, Sogon T, Naka-zawa T: Bilberry extract administration prevents retinal ganglion cell death in mice via the regulation of chaperone molecules under conditions of endo-plasmic reticulum stress. Clin. Ophthalmol., 11, 1825–1834, 2017.
51) Nishida T, Kuse Y, Mochizuki K, Shimazawa M, Yama moto T, Hara H: Protective effects of fluoroqui-nolones on UV-induced damage of cultured ocular cell lines. Eur. J. Pharmacol., 806, 59–66, 2017. 52) Li Z, Jo J, Jia J-M, Lo S-C, Whitcomb DJ, Jiao S,
Cho K, Sheng M: Caspase-3 activation via mitochon-dria is required for long-term depression and AMPA receptor internalization. Cell, 141(5), 859–871, 2010. 53) Junttila MR, Li S-P, Westermarck J:
Phosphatase-mediated crosstalk between MAPK signaling path-ways in the regulation of cell survival. FASEB J., 22(4), 954–965, 2008.
54) Sun M-H, Pang J-HS, Chen S-L, Kuo P-C, Chen K-J, Kao L-Y, Wu JY, Lin KK, Tsao YP: Photoreceptor protection against light damage by AAV-mediated overexpression of heme oxygenase-1. Invest.
Oph-thalmol. Vis. Sci., 48(12), 5699–5707, 2007.
55) Aruoma OI, Halliwell B, Hoey BM, Butler J: The an-tioxidant action of N-acetylcysteine: its reaction with hydrogen peroxide, hydroxyl radical, superoxide, and hypochlorous acid. Free Radic. Biol. Med., 6(6), 593–597, 1989.
56) Ogawa K, Oyagi A, Tanaka J, Kobayashi S, Hara H: The protective effect and action mechanism of Vac-cinium myrtillus L. on gastric ulcer in mice.
Phyto-ther. Res., 25(8), 1160–1165, 2011.
57) Zheng R, Po I, Mishin V, Black AT, Heck DE, Laskin
DL, Sinko PJ, Gerecke DR, Gordon MK, Laskin JD: The generation of 4-hydroxynonenal, an electrophilic lipid peroxidation end product, in rabbit cornea or-gan cultures treated with UVB light and nitrogen mustard. Toxicol. Appl. Pharmacol., 272(2), 345–355, 2013.
58) Black AT, Gordon MK, Heck DE, Gallo MA, Laskin DL, Laskin JD: UVB light regulates expression of antioxidants and inflammatory mediators in human corneal epithelial cells. Biochem. Pharmacol., 81(7), 873–880, 2011.
59) Kim Y-H, Oh TW, Park E, Yim N-H, Park KI, Cho WK, Ma JY: Anti-Inflammatory and Anti-Apoptotic Effects of Acer Palmatum Thumb. Extract, KIOM-2015EW, in a Hyperosmolar-Stress-Induced In Vitro Dry Eye Model. Nutrients, 10(3), 282, 2018.
60) Yin J, Yu F-SX: ERK1/2 mediate wounding- and G-protein-coupled receptor ligands-induced EGFR ac-tivation via regulating ADAM17 and HB-EGF shed-ding. Invest. Ophthalmol. Vis. Sci., 50(1), 132–139, 2009.