Nagoya City University Academic Repository
学 位 の 種 類 博士 (薬学) 報 告 番 号 甲第1703号 学 位 記 番 号 第349号 氏 名 壁谷 知樹 授 与 年 月 日 平成 31 年 3 月 25 日 学位論文の題名 ヒト iPS 細胞を用いた小腸における薬物動態予測評価系の構築 論文審査担当者 主査: 湯浅 博昭 副査: 松永 民秀.青山 峰芳.井上 靖道
名古屋市立大学学位論文
ヒトiPS細胞を用いた小腸における薬物動態予測評価系の構築
平成 30 年度 (2019年3月)
名古屋市立大学大学院薬学研究科 臨床薬学分野
壁谷 知樹
一.本論文は2019年3月 名古屋市立大学大学院薬学研究科において審査されたものである。 主査 湯浅 博昭 教授 副査 青山 峰芳 教授 井上 靖道 准教授 松永 民秀 教授 二.本論文は、学術情報誌に掲載された次の報文を基礎とするものである。 【基礎となる報文】
1. Tomoki Kabeya, Wakana Matsumura, Takahiro Iwao, Masakiyo Hosokawa, Tamihide Matsunaga. Functional analysis of carboxylesterase in human induced pluripotent stem cell-derived enterocytes. Biochemical and Biophysical Research Communications, 486, 143-148 (2017).
2. Tomoki Kabeya, Shimeng Qiu, Momona Hibino, Mizuka Nagasaki, Nao Kodama, Takahiro Iwao, Tamihide Matsunaga. Cyclic AMP Signaling Promotes the Differentiation of Human Induced Pluripotent Stem Cells into Intestinal Epithelial Cells. Drug Metabolism and Disposition, 46, 1411-1419 (2018).
三.本論文の基礎となる研究は、松永 民秀 教授の指導のもとに名古屋市立大学大学院薬 学研究科において行われた。
略語一覧
2-ME β-mercaptoethanol 3MM chromatography paper 5-aza-2'-dc 5-aza-2'-deoxycytidine ABC ATP-binding cassette
BCRP breast cancer resistance protein BIO 6-bromoindirubin-3'-oxime BSA albumin Bovine F-Ⅴ
cAMP cyclic adenosine monophosphate CDX2 caudal type homeobox 2
CES carboxylesterase
c-MYC v-myc myelocytomatosis viral oncogene homolog CYP cytochrome P450
DAPI 4',6-diamidino-2-phenylindole
DAPT N-[(3,5-difluorophenyl)acetyl]-L-alanyl-2-phenylglycine-1,1-dimethylethylester DMEM Dulbecco’s modified Eagle’s medium
DMEM/F12 DMEM mixed 1:1 with Ham’s F-12 CNT concentrative nucleoside transporter DNMT DNA methyltransferase
DPP4 dipeptidyl peptidase 4 EGF epidermal growth factor
Fa fraction absorbed in humans
FABP2 intestinal fatty acid-binding protein FBS fetal bovine serum
FGF fibroblast growth factor 2
GAPDH glyceraldehyde-3-phosphate dehydrogenase GFR growth factor reduced
GSK glycogen synthase kinase HBSS Hanks' Balanced Salt Solution HEPES HEPES Buffer Solution
HPRT hypoxanthine phosphoribosyltransferase IBMX 3-Isobutyl-1-methylxanthine
iPS induced pluripotent stem ISX intestine specific homeobox KLF4 kruppel-like factor 4
KSR KnockOut serum replacement L-Glu L-glutamine
MDR1 multiple drug resistance 1 MEGX monoethylglycylxylidide
MEK mitogen-activated protein kinase kinase NEAA non-essential amino acid
OATP organic anion-transporting polypeptide OCT3/4 octamer transcription factor-3/4
Papp apparent permeability coefficient
PBS Dulbecco’s phosphate buffered saline without calcium, magnesium PEPT1 peptide transporter 1
P-gp p-glycoprotein PKA protein kinase a
PNPA p -nitrophenyl acetate
PXR pregnane X receptor
RPMI Roswell Park Memorial Institute SDS sodium dodecyl sulfate
SGLT1 sodium-glucose cotransporter 1 SI sucrase-isomaltase
SLC solute carrier transporter SULT sulfotransferase
SOX2 sex determining region Y-box 2 TEER transepithelial electrical resistance TGF transforming growth factor
i 目次 第一章 序論 1 第二章 ヒトiPS細胞由来小腸上皮細胞のCESの機能解析 4 2.1 緒言 4 2.2 実験方法 6 2.2.1 試薬 6 2.2.2 ヒトiPS細胞の培養 7 2.2.3 ヒトiPS細胞の小腸上皮細胞への分化 7 2.2.4 Caco-2細胞の培養 7 2.2.5 Real-Time RT-PCR 8 2.2.6 Western blotting法による解析 8 1) ポリアクリルアミドゲルの作製およびバッファーの調整 8 2) 泳動サンプルの調製 9 3) SDS-PAGE 9 2.2.7 加水分解能の評価 10 1) バッファーおよび試薬の調製 10 2) S9の調製 10 3) PNPAの加水分解反応 10 4) MEGXおよびAspirinの加水分解反応 11 5) 高速液体クロマトグラフィー(HPLC) 11 2.2.8 統計解析 11 2.3 実験結果 12 2.3.1 ヒトiPS細胞由来小腸上皮細胞とCaco-2細胞におけるCES発現 レベルの比較 12 2.3.2 ヒトiPS細胞由来小腸上皮細胞とCaco-2細胞におけるCES加水 分解活性の比較 13 1) PNPAの代謝 13 2) MEGXの代謝 14 3) Aspirinの代謝 15
ii 2.4 考察 16 2.5 小括 17 第三章 cAMPシグナルはヒトiPS細胞の小腸上皮細胞への分化誘導を促進する 18 3.1 緒言 18 3.2 実験方法 19 3.2.1 試薬 19 3.2.2 細胞の培養 20 3.2.3 ヒトiPS細胞から小腸上皮細胞への分化 20 3.2.4 Real-Time RT-PCR 21 3.2.5 免疫蛍光染色 21 3.2.6 CYP3A4誘導剤処理 22 3.2.7 薬物代謝酵素活性の測定 22 3.2.8 取り込み試験 22 3.2.9 双方向輸送試験 23 3.2.10 統計解析 24 3.3 実験結果 25 3.3.1 ヒトiPS細胞株Windyにおける小腸上皮細胞への分化誘導に対する 8-Br-cAMP、IBMXおよびForskolinの効果 25 3.3.2 8-Br-cAMPとIBMXを組み合わせた分化誘導法の効果 (8-Br-cAMP+IBMX群) 28 3.3.3 ヒトiPS細胞由来小腸上皮細胞のCYP3A4誘導能と 薬物代謝酵素活性 33 3.3.4 ヒトiPS細胞由来小腸上皮細胞のPEPT1およびOATP活性 36 3.3.5 ヒトiPS細胞由来小腸上皮細胞のP-gp活性 37 3.4 考察 38 3.5 小括 41
iii 第四章 ヒトiPS細胞から作製した小腸上皮細胞の薬物動態学的機能解析 42 4.1 緒言 42 4.2 実験方法 44 4.2.1 試薬 44 4.2.2 細胞の培養 44 4.2.3 ヒトiPS細胞から小腸上皮細胞への分化 45 4.2.4 Real-Time RT-PCR 46 4.2.5 免疫蛍光染色 46 4.2.6 薬物代謝酵素活性の測定 47 4.2.7 取り込み試験 47 4.2.8 双方向輸送試験 47 4.2.9 膜透過試験 48 4.2.10 電子顕微鏡での観察 49 4.3 実験結果 50 4.3.1 ヒトiPS細胞由来小腸上皮細胞の遺伝子およびタンパク質 発現解析 50 4.3.2 ヒトiPS細胞由来小腸上皮細胞の代謝酵素活性 52 4.3.3 ヒトiPS細胞由来小腸上皮細胞のトランスポーター活性 53 4.3.4 ヒトiPS細胞由来小腸上皮細胞の膜透過特性 55 4.4 考察 57 4.5 小括 58 第五章 総括 59 謝辞 60 引用文献 61
1
1. 序論
ヒト人工多能性幹(induced pluripotent stem:iPS)細胞は、レトロウイルスベクターを用い て体細胞に4種類の遺伝子、octamer transcription factor-3/4(OCT3/4)、sex determining region Y-box 2(SOX2)、Kruppel-like factor 4(KLF4)、v-myc myelocytomatosis viral oncogene homolog (c-MYC)を導入することによって、2007年に山中らにより樹立された1)。このヒトiPS細胞 は、免疫拒絶のない再生医療や細胞治療の他にも、稀少疾病等の疾患メカニズムの解明に繋 がる可能性もあることから、ヒトiPS細胞に関する研究が盛んに行われている。しかし、ヒ トiPS細胞の利用は再生医療や細胞治療への応用だけに限ったことではない。例えば、多分 化能を有するという特長を生かし、ある特定の細胞に分化させることにより医学研究や創薬 研究における研究材料として利用することも期待されている。 新薬の開発において、小腸における吸収過程を把握することは、創薬を効率よく進める上 で極めて重要である 。小腸上皮細胞には peptide transporter 1(PEPT1)やorganic anion-transporting polypeptide(OATP)といった薬の取り込みに関わるトランスポーターやP-glycoprotein(P-gp)やbreast cancer resistance protein(BCRP)といった薬の排出に関わるトラ ンスポーターなど多くの薬物トランスポーターが存在する2, 3)。また、cytochrome P450(CYP) 3A4に代表される様々なCYP分子種や、プロドラッグの変換に関わるカルボキシルエステラ ーゼ(CES)などの薬物代謝酵素が存在するため、小腸は経口投与された薬のバイオアベイ ラビリティを左右する重要な臓器である4–6)。現在、医薬品の薬物動態を把握するために、実 験動物、ヒト小腸ミクロソームおよび結腸がん由来の株化細胞であるCaco-2細胞などが使用 されている。しかしながら、実験動物とヒトとの間には種差があること、ミクロソームはグ ルクロン酸抱合以外の第Ⅱ相反応の代謝評価が困難であること、Caco-2細胞は正常細胞と薬 物の膜透過特性が異なることなど多くの課題がある。本来ならば、薬の吸収過程の評価に生 体の腸管組織を使用するのが望ましい。しかしながら生体の腸管組織は、生体外で機能を維 持したまま培養する技術が確立されていないため短命であることや、倫理的な問題もあり、 その入手は非常に困難となっている7–9)。ヒトにおける薬物動態を正確に予測するためには、 ヒトの体内での薬の吸収や代謝特性をできる限り模倣した評価系が必要であるが、現在の技 術では小腸での薬物吸収と代謝の両方を包括的に評価することは難しい。 Caco-2細胞は、生体の腸管組織の代替として薬物の消化管吸収過程の評価系として汎用さ れている。実際にCaco-2細胞は、P-gpやBCRPといった排出トランスポーターの試験や、膜透
2 過試験に利用されている10–13)。しかし、生体の小腸と比較してCaco-2細胞ではCES分子種の 発現パターンが異なっていることや、CYP3A4の発現が極めて低いことなど、評価系として 問題がある。さらに強固なタイトジャンクションなどが原因で、透過特性にも生体小腸と差 異があることが報告されている14–19)。2011年にSpenceらによって開発された技術として、腸 管幹細胞をMatrigelという細胞外マトリックスに包埋することで作製される腸管を構成する 細胞を含んだ三次元構造体であるオルガノイドが注目されている20)。腸管オルガノイドの内 部には微絨毛や吸収上皮細胞、杯細胞、分泌細胞などの腸細胞の存在も確認され、クリプト などの腸管特異的な構造を示すため、再生医療だけでなく、薬物動態への応用も期待されて いる21)。しかしながら、立体的な腸管オルガノイドでは、腸管の吸収過程を定量的に評価す ることは難しい。したがって、薬物動態試験へ応用するには二次元で単層膜を形成する小腸 上皮細胞が望まれている。 ヒト小腸における薬物動態を正確に予測する評価系がないために、ヒト初代小腸細胞と同 等の機能を有したヒトiPS細胞由来小腸上皮細胞の作製が期待されている。低コストかつ簡 便にヒト多能性幹細胞から腸管細胞を作製する方法として、低分子化合物を用いた分化方法 が報告された22–27)。Glycogen synthase kinase(GSK)-3β阻害剤である6-bromoindirubin-3'-oxime
(BIO)およびγ-secretase 阻害剤であるN-[(3,5-difluorophenyl)acetyl]-L-alanyl-2-phenylglycine-1,1-dimethylethylester(DAPT)を用いた方法は、分化に効果的な低分子化合物を用いている 点では有用である22)。しかしながら、Yangらが報告したように、Notchシグナルが不活性化
されてAtoh1を発現するようになった前駆細胞は分泌系細胞へと分化するため、この方法で 得られる細胞は分泌系細胞の発現が多いという特徴を持つ28)。また、その他にヒトiPS細胞
から小腸上皮細胞への分化誘導に、transforming growth factor(TGF)-β受容体阻害剤、DNA methyltransferase(DNMT)阻害剤、 mitogen-activated protein kinase kinase(MEK)阻害剤を 用いる方法も報告されている23, 24, 29)。この方法により分化させた小腸上皮細胞は、腸管に発
現する薬物代謝酵素の活性や、PEPT1およびBCRPのトランスポーター活性が認められた30)。
また、膜透過試験により得られたapparent permeability coefficient(Papp)とfraction absorbed in
humans(Fa)の間には良好な相関が認められ、ヒト消化管におけるFaを予測できる可能性が
示唆された。しかしながら、P-gpを介した輸送活性が認められなかったことや、主要な代謝 酵素であるCYP3A4の活性が低いなど機能的に不十分であった。さらにプロドラッグの代謝 に影響を与える重要なCESの機能の解析も未だ行われていないなどの問題もあった。
3 ヒトiPS細胞由来小腸上皮細胞を創薬研究に応用するためには、CESやCYPの代謝、薬物ト ランスポーターによる輸送、受動輸送による膜透過など小腸における重要な薬物動態学的な 機能解析が必須である。そこで本論文では、ヒトiPS細胞から分化誘導した小腸上皮細胞に おけるCESの機能解析を行い、その機能についてCaco-2細胞と比較し得られた知見について 論ずる。さらに、ヒトiPS細胞から分化誘導した小腸上皮細胞の分化成熟および機能の向上 に、cyclic adenosine monophosphate(cAMP)シグナルの活性化が有用であることおよび、確 立した新規の分化誘導法により作製された小腸上皮細胞の機能に関して、ヒト初代小腸細胞 およびCaco-2細胞と比較して得られた知見について論ずる。
4 2. ヒトiPS細胞由来小腸上皮細胞のCESの機能解析 2.1 緒言 医薬品開発において、薬の吸収、組織移行、あるいは代謝が原因となり開発が中断される ことがあるが、これを解決する方法のひとつにプロドラッグ化がある。生体に豊富に存在す る加水分解酵素である CES が、プロドラッグの生体内変換を制御する酵素として利用され ることが多い31–33)。CES はヒト CES1 酵素のアミノ酸配列に基づく相同性から、CES1 から
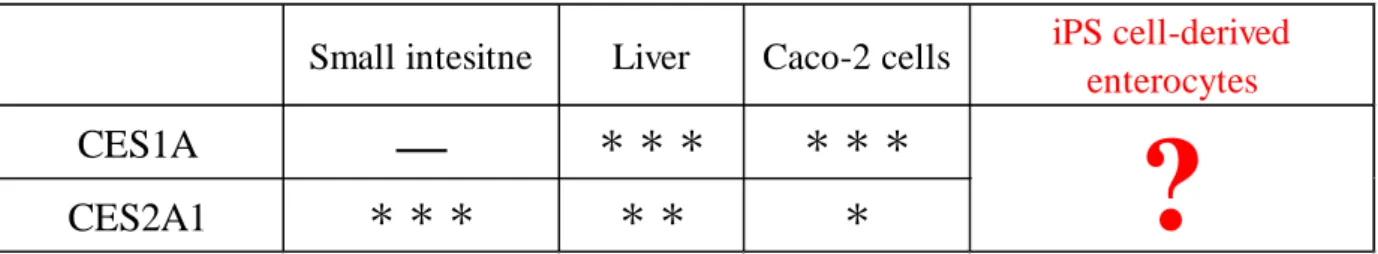
CES5 まで大きく 5 つのファミリーに分けられる34–36)。経口投与されたすべての薬物はまず 小腸を通過してから門脈を経て肝臓に至るが、CES の加水分解能には臓器差が存在するた め、小腸と肝臓において薬物の受ける代謝は異なる35)。小腸での CES による初回代謝はプ ロドラッグの生体内変換に大きく関与するが、ヒト小腸においては CES2A1 がその加水分 解のほとんどを担っており、CES1A はほとんど発現していない37)。CES 代謝予測モデルと しては実験動物やヒト小腸あるいは肝臓のミクロソームが用いられている。しかしながら、 実験動物では CES 発現パターンに種差があり、ミクロソームでは薬物吸収過程を評価でき ないといった問題がある。さらに、現在薬の吸収・排出の予測に汎用されている Caco-2 細 胞の CES 発現パターンは、ヒト小腸とは異なり、CES1A が高発現し CES2A1 の発現は低い ことが知られている(Table 1 参照)18, 19)。そのため、ヒト生体内における小腸での吸収およ
び CES による代謝を同一の系で評価できる薬物動態モデルは報告されていない。そこで本 研究ではヒト iPS 細胞由来小腸上皮細胞における CES 発現解析および機能解析を行い、in
vitro における CES による代謝の新規評価系として有用か検討した。
ヒト iPS 細胞由来小腸上皮細胞の CES 発現を Real-Time PCR 法および Western blotting 法 により確認した。ヒト iPS 細胞由来小腸上皮細胞および Caco-2 細胞から S9 を調製し、CES の各アイソフォームに対して非選択的な基質である p-nitrophenyl acetate(PNPA)4, 18)、CES1A
の基質である monoethylglycylxylidide(MEGX)および CES2A1 の基質である aspirin38)を使
用して、加水分解活性を評価した。その際に、CES1A および CES2A1 の阻害剤である digitonin および telmisartan を用いて39)、代謝活性への影響について評価した。
5
Table 1. Specific expression profile of CES isozymes in human tissue, Caco-2 cells and human iPS cell-derived enterocytes.
Small intesitne Liver Caco-2 cells iPS cell-derived
enterocytes
CES1A
—
*** ***6
2.2 実験方法 2.2.1 試薬
Aspirin、chlorpropamide、digitonin、Dulbecco’s modified Eagle’s medium(DMEM)high glucose、 DMEM-Ham’s F12(DMEM/F12)、ethanol(99.5%)、N,N,N',N',-tetramethyl-ethylendiamine、 N2 supplement、PD98059、p-nitrophenol、p-nitrophenyl acetate、sodium dodecyl sulfate(SDS)、 sucrose、5-aza-2'-deoxycytidine(5-aza-2'-dC)、NaCl、glyceraldehyde-3-phosphate dehydrogenase (GAPDH)抗体は和光純薬工業(大阪)より、L-glutamine(L-Glu)、non-essential amino acid (NEAA)はBiological Industries(コネチカット州クロムウェル、米国)より、Y-27632はFocus Biomolecules(ペンシルべニア州プリマス・ミーティング、米国)より、KnockOut serum replacement(KSR)、GlutaMaxはGIBCO(カリフォルニア州カールズバッド、米国)より、 A 83-01、acetonitrile、β-mercaptoethanol(2-ME)、gelatin、Trizma® Base(Tris)、MEGXは
SIGMA-Aldrich(ミズーリ州セントルイス、米国)より、accutase、Albumin Bovine F-Ⅴ(BSA)、 dimethylsulfoxide(DMSO)、telmisartanはナカライテスク(京都)より、mitomycin C(MMC) は協和発酵キリン(東京)より、penicillin–streptomycinはコスモバイオ(東京)よりfibroblast growth factor 2(FGF2)、activin A、epidermal growth factor(EGF)はPeproTech(ニュージャ ージー州ロッキーヒル、米国)より、Dulbecco’s phosphate buffered saline without calcium, magnesium(PBS)用錠剤はタカラバイオ(滋賀)より、fetal bovine serum(FBS)はニチレ イ(東京)より、HEPES Buffer Solution(HEPES)は同仁化学研究所(熊本)より、Growth Factor Reduced(GFR)MatrigelはBecton, Dickinson and Company(ニュージャージー州フラン クリンレイクス、米国)より、B27 serum free supplement、Pierce™ BCA Protein Assay Kit、 Roswell Park Memorial Institute(RPMI)+GlutaMax mediumはThermo Fisher Scientific(マサチ ューセッツ州ウォルサム、米国)より、chromatography paper(3MM)はWhatman(ペンシル ベニア州ピッツバーグ、米国)、human adult small intestinal total RNA from 5 donors、human adult liver tissue total RNA from 5 donorsはBiochain(カルフォルニア州ニューワーク、米国) より、CES1抗体はRockland(ペンシルベニア州ライムリック、米国)より、CES2抗体はR&D Systems(ミネソタ州ミネアポリス、米国)より、Agencourt RNAdvance Tissue KitはBeckman Coulter, Inc.(カリフォルニア州ブレア、米国)より、ReverTra Ace qPCR KitはTOYOBO(大 阪)より、KAPA SYBR Fast qPCR Kitは日本ジェネティクス(東京)より、Goat Anti-Rabbit IgG (HRP)、Goat Anti-Mouse IgG (HRP)、Donkey Anti-Goat IgG (HRP)はAbcam(ケンブリッ
7
ジ、英国)より購入した。その他の試薬はすべて市販の特級品を用いた。
2.2.2 ヒトiPS細胞の培養
ヒト胎児肺線維芽細胞から樹立されたヒトiPS細胞(Windy)は、国立成育医療研究センタ ー研究所の梅澤明弘博士よりご供与いただいた。ヒトiPS細胞は20% KSR、1% NEAA、2 mM L-Glu、0.1 mM 2-ME、5 ng/mL FGF2を含むDMEM/F12(ヒトiPS培地とする)を用いて、MMC 処理により不活化したマウス胎仔線維芽細胞上で培養した。培地交換は毎日行い、3–4 日毎 に霊長類ES/iPS 細胞用細胞剥離液を用いて剥離し、継代培養を行った。ヒトiPS細胞の小腸 上皮細胞への分化は、ヒトiPS細胞が培養ディッシュに対し、未分化コロニーの占める割合 が約70%になった状態で開始した。
2.2.3 ヒトiPS 細胞の小腸上皮細胞への分化
0.5% FBS、100 units/mL penicillin G、100 µg/mL streptomycin sulfate、100 ng/mL activin Aを 含むRPMI + GlutaMax mediumで48時間培養後、2% FBS、100 units/mL penicillin G、100 µg/mL streptomycin sulfate、100 ng/mL activin Aを含むRPMI + GlutaMax mediumで24時間培養するこ とで内胚葉に分化させた。その後、2% FBS、1% GlutaMax、100 units/mL penicillin G、100 µg/mL streptomycin sulfate、 250 ng/mL FGF2を含むDMEM/F12 mediumで96時間培養するこ とで小腸幹細胞に分化させた。Activin AおよびFGF2処理後の小腸幹細胞様細胞をaccutaseに て剥離し、あらかじめヒトiPS培地で30倍に希釈したGFR-Matrigel上に播種した。播種後は2% FBS、1% NEAA、2 mM L-Glu、100 units/mL penicillin G、100 µg/mL streptomycin sulfate、2% B27 supplement、1% N2 supplement、20 ng/mL EGFを含むDMEM/F12 mediumで19日間培養し た。分化開始14日目からPD98059を20 µM、5-aza-2'-dCを5 µM、A 83-01を0.5 µMになるよう に添加して小腸上皮細胞に分化させた。
2.2.4 Caco-2細胞の培養
Caco-2細胞(RCB0988)は、理化学研究所バイオリソースセンター(茨城)より入手し、 ゼラチンでコートした細胞培養ディッシュ上で、10% FBS、2 mM L-Glu、1% NEAA、100 units/mL penicillin G、100 μg/mL streptomycin sulfateを含むDMEM(high glucose)を用いて培 養した。コンフルエントになってから2–3日に一度培地交換をして14日以上培養したものを 実験に用いた。
8
2.2.5 Real-Time RT-PCR
Agencourt® RNAdvance™ Tissue Kitの取扱説明書に従い、ヒトiPS細胞由来小腸上皮細胞お
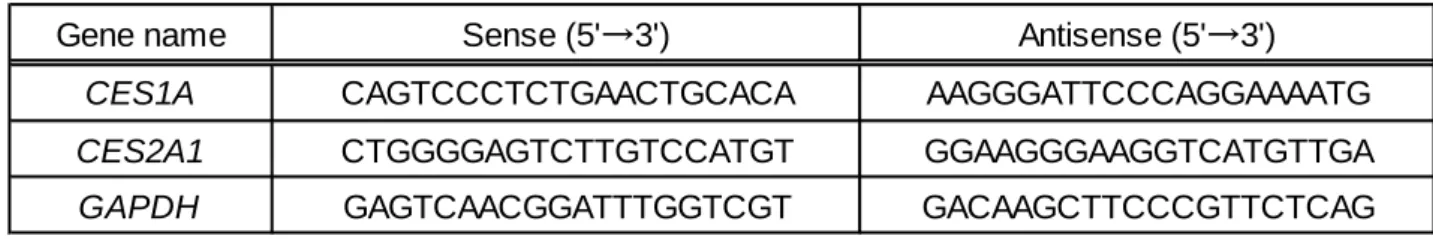
よびCaco-2細胞のRNAを抽出した。cDNA合成はReverTra Ace qPCR RT Master Mixを、Real-Time PCRの反応混合液はKAPA SYBR Fast qPCR Kitを、反応には7300 Real Mixを、Real-Time PCR System (Applied Biosystems、米国)を使用し、取扱説明書に従い行った。PCRプライマーは、Table 2に示したものを用いた。内在性コントロールとしてGAPDHを用いて補正した。コントロー ルとして、購入したhuman adult small intestineおよびhuman adult liverのtotal RNAから合成し たcDNAを用いた。
Table 2. Sequences of primers for real-time PCR analysis.
2.2.6 Western blotting法による解析 1)ポリアクリルアミドゲルの作製およびバッファーの調製 ゲルカセットにガラスプレートを2枚セットした。分離ゲルモノマー溶液を調製し(組成 はTable 3参照、6 mL/ゲル1枚)、コームをゲルカセットに差し込んだ下端より1 cm低い位置 まで分離ゲルモノマー溶液を注いだ。注入したモノマー溶液の上に直ちに70%エタノール溶 液を重層し、20分間ほど放置して重合させた。重層したエタノールを捨て精製水でよく洗浄 した。濃縮ゲルモノマー溶液を調製し(組成はTable 3参照、4 mL/ゲル1枚)、分離ゲル上の 水分を除いてから、濃縮ゲルモノマー溶液をショートガラスプレート上端まで重層し直ちに コームを静かに挿入した。30分間ほど放置して濃縮ゲルが重合した後、コームを抜き取りポ リアクリルアミドゲルとしてSDS-PAGEに用いた。
Gene name Sense (5'→3') Antisense (5'→3')
CES1A CAGTCCCTCTGAACTGCACA AAGGGATTCCCAGGAAAATG
CES2A1 CTGGGGAGTCTTGTCCATGT GGAAGGGAAGGTCATGTTGA
9 Table 3. Composition of gel solution for SDS-PAGE
2)泳動サンプルの調製
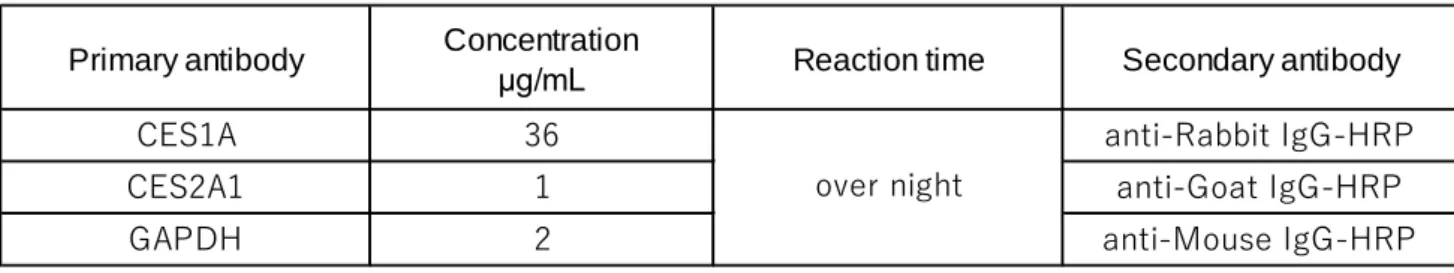
泳動サンプル用の細胞に、細胞培養ディッシュ1枚あたり400 µLのSDS sample bufferを加 え、セルリフターで剥離して細胞を1.5 mLサンプルチューブに回収した。超音波式ホモジナ イザーVP-050(タイテック、埼玉)で細胞を破砕した後、100℃で3分間煮沸して泳動サンプ ルとした。タンパク量の測定は、Pierce™ BCA Protein Assay Kitを用い、Synergy HTX マル チモードプレートリーダーとGen 5データ解析ソフトウェア(ともにBio Tek、米国)で吸光 度測定(570 nm)と解析を行った。 3)SDS-PAGE ミニプロティアンTetraセル(Bio-Rad、米国)の取扱説明書に沿って行った。0.02 A(ゲル 1枚あたり)、250 Vで90分電気泳動した。泳動後、ゲルカセットからゲルを取り出しtransfer buffer中で10分間振盪した。転写前にPVDFメンブレンをメタノールで軽くすすぎ、transfer buffer中で2回(1回にあたり10から15分間)振盪して洗浄した。転写装置Trans-Blot® SD Semi-Dry Transfer Cell(Bio-Rad、米国)に移した後、ゲル1枚あたり定電流0.11 Aで30分間かけて 転写した。転写を終えたメンブレンを5%スキムミルク中で20分間振盪した。一次抗体をTBS-Tで希釈し、パラフィルム上でメンブレンに転写されたタンパク質と反応させた。一次抗体 を反応させた後、メンブレンをTBS-Tで3回以上洗浄した。二次抗体をTBS-Tで5000倍に希釈 し、パラフィルム上で30分間、室温下で反応させた(一次抗体と二次抗体の対応については
Stacking gel monomer solution Final concentration 1.5 M Tris-HCL (pH 6.8, SDS 0.4%) 0.375 mol/L 30% (w/v) Acrylamide/Bis Mixed Solution (29:1) 3.75% (w/v)
Ammonium Peroxodisulfate 0.10%
N, N, N, N', -Tetramethyl-ethylendiamine 8.25 mol/L
Resolving gel monomer solution Final concentration 1.5 M Tris-HCL (pH 8.8, SDS 0.4%) 0.375 mol/L 30% (w/v) Acrylamide/Bis Mixed Solution (29:1) 10% (w/v)
Ammonium Peroxodisulfate 0.83%
10
Table 4参照)。ラップフィルム上にPierce® Western Blotting Substrateのdetection regent 1と detection regent 2を等量混ぜ、メンブレンをその混合液に1分間ほど浸して発光基質を結合さ せた。その後、ポリプロピレン製の透明シートの間にメンブレンを挟み、ルミノイメージア ナライザLAS-3000 mini(富士フイルム、東京)でタンパク質のバンドを検出した。
Table 4. Conditions of antibodies used in Western blotting.
2.2.7 加水分解能の評価
1)バッファーおよび試薬の調製
200 mM Na, Kリン酸bufferは200 mM KH2PO4溶液と200 mM Na2HPO4・12H2O溶液を混合し、
pH 7.4となるように調製した。1 M Tris-HCl bufferはpH 8.0となるように調製した。50 mM HEPES buffer(pH 7.6)はHEPESを精製水に溶解させて、HClを加えてpH 7.6に合わせてから 終濃度が50 mMとなるようにメスアップして調製した。 2)S9の調製 分化終了後のヒトiPS細胞由来小腸上皮細胞とCaco-2細胞を用いてそれぞれのS9を、Imai らの報告を元に以下の手順に従って調製した4, 18)。細胞をPBSで2回洗浄し、SET buffer(Table 5参照)で1.5 mLサンプルチューブに回収した。回収した細胞を超音波式ホモジナイザーVP-050で破砕した。9,000×g、4℃で20分間遠心し、上清のタンパク質を定量し加水分解能評価 のサンプルとして用いた。タンパク量の測定は、Pierce™ BCA Protein Assay Kitを用いた。 Table 5. Composition of SET buffer.
3)PNPAの加水分解反応
Hosokawaらの方法を参考にして40)、PNPA加水分解反応を行った。PNPAを加水分解反応時
Primary antibody Concentrationμg/mL Reaction time Secondary antibody
CES1A 36 anti-Rabbit IgG-HRP
CES2A1 1 anti-Goat IgG-HRP
GAPDH 2 anti-Mouse IgG-HRP
over night SET buffer sucrose 290 mmol/L EDTA 1 mmol/L Tris 50 mmol/L Final concentration
11
の終濃度が500 µMになるように精製水で希釈した。また、凍結保存しておいたS9を氷上で 解凍し、200 mMリン酸buffer(pH 7.4)で希釈し、37℃に設定したdry bath incubator(Major Science、米国)で15分間温めた。PNPA溶液、Tris-HCl buffer(pH 8.0)、S9溶液の順に、そ れぞれの容量比が18:1:1になるように1.5 mLサンプルチューブに加えてボルテックスし、 37℃条件下で加水分解反応を開始した。所定の時間経過後、氷冷したacetonitrileを反応溶液 と等量添加し、反応を停止させた。また反応開始前のプレインキュベーション時および加水 分解反応時に、終濃度がdigitoninは250 µM、telmisartanは200 µMとなるように添加した。阻 害剤処理を行わなかったvehicle群についてはDMSOを加えた。反応停止後、10,000×g、4℃で 5分間遠心し、上清を吸光度測定のサンプルとして用いた。Synergy HTXマルチモードプレー トリーダーとGen 5データ解析ソフトウェアで吸光度測定(405 nm)を行った。市販のp-nitrophenolで検量線を作成し、p-nitrophenolに加水分解された濃度を算出した。 4)MEGXおよびAspirinの加水分解反応 各S9溶液を50 mM HEPES buffer(pH 7.4)で最終濃度200–220 µg/mLになるように希釈し、 37℃で15分間温めた後、2.5 mM MEGXまたは500 μM aspirinを添加した。内部標準物質とし てchlorpropamideを使用し、これを含むacetonitrileを添加して反応を停止させた。PNPAの加 水分解反応と同様に阻害剤にはdigitoninおよびtelmisartanを使用した。反応停止後、10,000×g、 4℃で5分間遠心し、上清を測定用サンプルとして使用した。 5)高速液体クロマトグラフィー(HPLC) HPLCシステムには、LC-20ADポンプ、CTO-20Aカラムオーブン、SPD-20A検出器、LC solution(島津製作所、京都)およびInertsil ODS-2カラム(5 µm 4.6 × 150 mm)(ジーエルサ イエンス、東京)を使用した。移動相には10 mM phosphate buffer(pH 2.6)とacetonitrileを使 用した。2,6-xylidine(MEGXの代謝物)の測定には65:35の割合で、salicylic acid(aspirinの代 謝物)の測定には60:40の割合で混合し、1.0 mL/minの流速で測定を行った。カラム温度は 40℃、測定波長は210 nmとした。 2.2.8 統計処理 2群間の比較にはStudent’s t-test検定を用いた。多重の比較には一元配置分散分析を行った 後に、Bonferroni correctionを用いた。危険率5%未満を有意とした。
16
2.4 考察
CES1A mRNA およびタンパク質発現は、Caco-2 細胞と比較してヒト iPS 細胞由来小腸上 皮細胞において低かった。一方で CES2A1 mRNA およびタンパク質発現は同程度であった (Fig. 1A, B)。これらの結果から、ヒト iPS 細胞由来小腸上皮細胞における CES アイソフォ ームの発現パターンは、Caco-2 細胞よりもヒト小腸により類似していると示唆された。しか しながら、ヒト小腸と比較するとヒト iPS 細胞由来小腸上皮細胞の CES2A1 の発現量は低い ため、分化誘導法の改善が必要である(Fig. 1A)。
PNPA 加水分解試験において、p-nitrophenol 生成量はヒト iPS 細胞由来小腸上皮細胞より も Caco-2 細胞の方が大きかった(Fig. 2A, B)。しかしながら、Caco-2 細胞において CES1A 阻害剤および CES2A1 阻害剤の処理で、同程度に PNPA 加水分解活性が阻害された(Fig. 2D)。一方でヒト iPS 細胞由来小腸上皮細胞の PNPA 加水分解活性は、CES2A1 阻害剤の処 理によってより顕著に阻害された(Fig. 2C)。これらの結果から、ヒト iPS 細胞由来小腸上 皮細胞における PNPA 加水分解活性の主要な代謝酵素は CES2A1 であるのに対し、Caco-2 細胞では CES1A および CES2A1 の両方が CES による加水分解に関与していると示唆され た。この結果は CES1A および CES2A1 の mRNA およびタンパク質発現に相関していると考 えられる。
CES1A の基質 MEGX および CES2A1 の基質 aspirin を用いた加水分解試験においても同 様の結果が観察された。MEGX 代謝物は Caco-2 細胞で検出されたが、ヒト iPS 細胞由来小 腸上皮細胞では検出されなかった(Fig. 3)。一方で CES2A1 活性は、ヒト iPS 細胞由来小腸 上皮細胞および Caco-2 細胞において同程度であった(Fig. 4)。このことはヒト iPS 細胞由 来小腸上皮細胞は CES1A による加水分解活性をほとんど有さず、CES による加水分解は、 主に CES2A1 を介して行われることを示している。
CES1A 基質である lidocain は MEGX を経て 2,6-xylidine に代謝されることが報告されてい る41, 42)。本研究での予備的な検討において、lidocain よりも MEGX を用いることで、より多
17
CES1A 阻害剤である digitonin 添加によってのみ阻害されたこと、および MEGX の加水分解 が、CES1A の発現が極めて低く、CES2A1 の発現が Caco-2 細胞と同程度のヒト iPS 細胞由 来小腸上皮細胞において認められなかったことから、MEGX は lidocain と同様に CES1A の 基質として使用できると結論付けた。また本研究において CES 阻害剤を使用しても加水分 解 活 性を 完 全に 阻害で き なか っ た。 その原 因 とし て は、 S9 に存在する arylacetamide deacetylase、paraoxonase、butyrylcholinesterase などの CES 以外の加水分解酵素の寄与がある ためだと考えられる39, 43–45)。
本研究によって、ヒト iPS 細胞由来小腸上皮細胞において CES2A1 が CES1A よりも高い レベルで発現していることおよび、ヒト iPS 細胞由来小腸上皮細胞における CES を介した 加水分解反応では CES2A1 が主に寄与することが明らかとなった。これらの結果は、ヒト iPS 細胞由来小腸上皮細胞における CES アイソフォームの発現パターンおよび活性が、 Caco-2 細胞と比較してヒト小腸により類似していることを示唆している。したがって、ヒト iPS 細胞由来小腸上皮細胞は、ヒト消化管における薬物の CES を介した代謝および吸収を評価 するための新規システムとして有用である。 2.5 小括 本研究により、ヒト iPS 細胞由来小腸上皮細胞は、Caco-2 細胞よりも優れた小腸における CES 代謝の評価系として利用可能であることが示唆された。しかしながら、本研究では CES2A1 の発現レベルがヒト小腸よりも低いこと、検討した基質の数が少ないことに加えて、 基質の吸収を含めた実験の評価を行っていないことなどの残された課題については、今後さ らなる検討が必要である。
18 3. cAMPシグナルはヒトiPS細胞の小腸上皮細胞への分化誘導を促進する 3.1 緒言 非臨床試験の段階で経口バイオアベイラビリティ(Foral)を出来るだけ精度良く予測し、 薬効と安全性を確保することは、医薬品開発の成功の確率を上げる重要な因子の一つである。 Foralは、消化管での吸収率(Fa)、代謝回避率(Fg)および肝臓での代謝回避率(Fh)の積で 算出される。そのため、薬物動態における小腸の寄与は大きく、薬物代謝酵素および薬物ト ランスポーターを考慮した消化管吸収過程の評価の重要性が高まっている46, 47)。現在、ヒト 小腸の薬物動態における代謝と吸収を同時に予測する評価系の報告はなされていないため、 ヒト小腸と同等の機能を有したヒト iPS 細胞由来小腸上皮細胞の作製が期待されている。ヒ ト iPS 細胞から心筋細胞や神経細胞、肝細胞など様々な組織の細胞への分化に関して報告が されているが、小腸細胞への分化の報告は他の細胞と比較して少ない。これまでに様々な方 法でヒト iPS 細胞の分化誘導が行われているが、特に低分子化合物を用いた分化誘導法は、 低コストかつ安定して供給できる方法であるため、ヒト iPS 細胞から小腸上皮細胞への分 化に用いる方法として有用であると考えられる23–27)。しかしながら、小腸上皮細胞への分化 を促進する低分子化合物に関する知見は乏しく、どのような効果を持った化合物が有用であ るかということはよく分かっていない。そこで分化誘導に有用な化合物を見出すために、小 腸細胞への分化過程に選択的な作用を持つ化合物を添加するスクリーニングを行った。その 結果、cAMP シグナルの活性化がヒト iPS 細胞の小腸上皮細胞への分化誘導に効果的である ことを示す知見が得られた。
cAMP は ATP からアデニル酸シクラーゼを介して産生される。生成した cAMP は protein kinase A(PKA)を活性化させ、様々な生理機能を示す。これまでに、ヒト多能性幹細胞由 来肝細胞の分化成熟が cAMP シグナルによって促進されるという報告48)や、PKA シグナル が細胞分化のタイミングを制御するという報告 49)があるものの、小腸への分化誘導に関す る報告は未だなされていない。そこで本研究では、cAMP シグナルに着目し、cAMP アナロ グであり PKA 活性化剤である 8-bromo-cAMP(8-Br-cAMP)、phosphodiesterase(PDE)を阻 害し細胞内 cAMP 濃度を高める 3-isobutyl-1-methylxanthine(IBMX)およびアデニル酸シク ラーゼを活性化して細胞内 cAMP 濃度を高める forskolin を用いて、ヒト iPS 細胞から小腸 上皮細胞への分化誘導過程における、それら低分子化合物による影響を検討した。
19
3.2 実験方法 3.2.1 試薬
DMEM high glucose、DMEM/F12、N2 supplement、NEAA、PD98059、5-aza-2'-dC、forskolin、 は和光純薬工業(大阪)より、RPMI+GlutaMax medium、B27 serum free supplement、Alexa Fluor 488 (or 568) Goat Anti-Mouse IgG(H+L)、Alexa Fluor 488 (or 568) Goat Anti-Rabbit IgG(H+L)、 occludin 抗体は Thermo Fisher Scientific(マサチューセッツ州 ウォルサム、米国)より、GFR-Matrigel は Becton, Dickinson and Company(ニュージャージー州フランクリンレイクス、米 国)より、FGF2、activin A、EGF は PeproTech(ニュージャージー州ロッキーヒル、米国) より、A 83-01、IBMX は SIGMA-Aldrich(ミズーリ州セントルイス、米国)より、accutase、 DMSO はナカライテスク(京都)KSR、GlutaMax は GIBCO(カリフォルニア州カールズバ ッド、米国)より、TaKaRa BCA Protein Assay Kit、PBS 用錠剤はタカラバイオ(滋賀)より、 FBS はニチレイ(東京)より、Agencourt RNAdvance Tissue Kit は Beckman Coulter, Inc.(カ リフォルニア州ブレア、米国)より、ReverTra Ace qPCR Kit は TOYOBO(大阪)より、KAPA SYBR Fast qPCR Kit は日本ジェネティクス(東京)より、HNF-4α 抗体、GATA6 抗体、FOXA3 抗体、P-gp 抗体、BCRP 抗体、Alexa Fluor 488 Donkey Anti-Goat IgG(H+L)は Abcam(ケン ブリッジ、英国)、villin 抗体、PEPT1 抗体は Santa Cruz Biotechnology(テキサス州ダラス、 米国)より、fatty acid-binding protein 2(FABP2)抗体は R&D Systems(ミネソタ州ミネアポ リス、米国)より、human adult small intestinal total RNA from 5 donors は Biochain(カルフォ ルニア州ニューワーク、米国)より、8-Br-cAMP は Enzo Life Sciences(ニューヨーク州ファ ーミングデール、米国)、rhodamine 123 は Dojindo Laboratories(熊本)、[3H]-glycylsarcosine
は Moravek(カリフォルニア州ブレア、米国)より購入した。その他の試薬はすべて市販の 特級品を用いた。
20 3.2.2 細胞の培養 ヒトiPS細胞株610B1は、ヒトの臍帯血にOCT3/4、SOX2、KLF4、L-MYC、LIN28を、エピ ソーマルベクター(pCXLE)を用いて導入することで京都大学にて樹立されたものであり、 理化学研究所バイオリソースセンター(茨城)より入手した。第二章と同様な方法にてヒト iPS細胞株Windyおよび610B1を培養した。Caco-2細胞(RCB0988)は、理化学研究所バイオ リソースセンター(茨城)より入手し、第二章と同様な方法で培養した。なお本章では主に Windyを使用し、一部610B1を使用した。 3.2.3 ヒト iPS 細胞から小腸上皮細胞への分化 第二章と同様な方法でヒト iPS 細胞を小腸上皮細胞へ分化誘導した。分化誘導期間に 3 種 類の低分子化合物(終濃度 1 mM 8-Br-cAMP、0.5 mM IBMX、30 μM Forskolin)を添加し、 小腸上皮細胞への分化に及ぼす影響について検討した。また、それぞれの化合物の添加期間 は分化開始後 8–13 日および 13–26 日の 2 期間設定した。添加期間検討後の実験では、分化 開始後 8–14 日に 8-Br-cAMP を、14–26 日に IBMX を終濃度がそれぞれ 1 mM および 0.5 mM となるよう添加し(8-Br-cAMP+IBMX 群とする)、小腸上皮細胞への分化に及ぼす影響につ いて検討した。
21
3.2.4 Real-Time RT-PCR
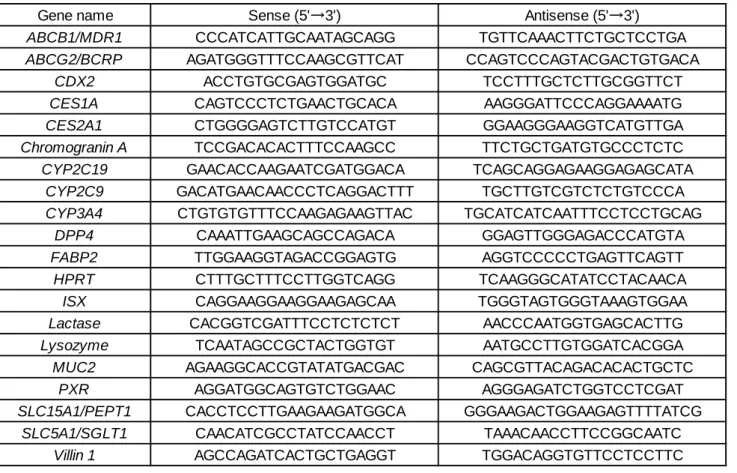
第二章と同様の方法にて行った。PCR プライマーは、Table 6 に示したものを用いた。内 在性コントロールとして hypoxanthine phosphoribosyltransferase(HPRT)を用いて補正した。 Table 6. Sequences of primers for real-time PCR analysis.
3.2.5 免疫蛍光染色
Villin、occludin、CDX2 および HNF4α の染色では、4℃条件下で 5 分間 methanol(-20℃) にて分化した細胞を固定透過処理した。PEPT1、P-gp 、FOXA3 および GATA6 の染色では、 細胞を 4%(w/v)パラホルムアルデヒド溶液中で 30 分間固定し、0.1% Triton X-100 溶液中 にて 5 分間室温で透過処理した。PBS で 3 回洗浄した後、5% FBS を含有する PBS にて室温 で 20 分間ブロッキングした。次いで細胞を一次抗体と共に室温で 60 分間インキュベートし た。一次抗体は以下のように希釈した:villin 1:100、occludin 1:100、PEPT1 1:50、P-gp 1:20、 CDX2 1:50、HNF4α 1:100、FOXA3 1:50、GATA6 1:100。細胞を PBS で洗浄し、Alexa Fluor
Gene name Sense (5'→3') Antisense (5'→3')
ABCB1/MDR1 CCCATCATTGCAATAGCAGG TGTTCAAACTTCTGCTCCTGA
ABCG2/BCRP AGATGGGTTTCCAAGCGTTCAT CCAGTCCCAGTACGACTGTGACA
CDX2 ACCTGTGCGAGTGGATGC TCCTTTGCTCTTGCGGTTCT
CES1A CAGTCCCTCTGAACTGCACA AAGGGATTCCCAGGAAAATG
CES2A1 CTGGGGAGTCTTGTCCATGT GGAAGGGAAGGTCATGTTGA
Chromogranin A TCCGACACACTTTCCAAGCC TTCTGCTGATGTGCCCTCTC
CYP2C19 GAACACCAAGAATCGATGGACA TCAGCAGGAGAAGGAGAGCATA
CYP2C9 GACATGAACAACCCTCAGGACTTT TGCTTGTCGTCTCTGTCCCA
CYP3A4 CTGTGTGTTTCCAAGAGAAGTTAC TGCATCATCAATTTCCTCCTGCAG
DPP4 CAAATTGAAGCAGCCAGACA GGAGTTGGGAGACCCATGTA
FABP2 TTGGAAGGTAGACCGGAGTG AGGTCCCCCTGAGTTCAGTT
HPRT CTTTGCTTTCCTTGGTCAGG TCAAGGGCATATCCTACAACA
ISX CAGGAAGGAAGGAAGAGCAA TGGGTAGTGGGTAAAGTGGAA
Lactase CACGGTCGATTTCCTCTCTCT AACCCAATGGTGAGCACTTG
Lysozyme TCAATAGCCGCTACTGGTGT AATGCCTTGTGGATCACGGA
MUC2 AGAAGGCACCGTATATGACGAC CAGCGTTACAGACACACTGCTC
PXR AGGATGGCAGTGTCTGGAAC AGGGAGATCTGGTCCTCGAT
SLC15A1/PEPT1 CACCTCCTTGAAGAAGATGGCA GGGAAGACTGGAAGAGTTTTATCG
SLC5A1/SGLT1 CAACATCGCCTATCCAACCT TAAACAACCTTCCGGCAATC
22
488 または 568 で標識された二次抗体と共に室温で 60 分間インキュベートした。その後、 細胞を1 μg/mL の 4′,6-diamidino-2-phenylindole(DAPI)と共に室温で 5 分間インキュベート した。細胞の観察は、LSM 510 共焦点顕微鏡(Carl Zeiss、ドイツ)もしくは Opperetta (PerkinElmer、米国)を使用した。
3.2.6 CYP3A4 誘導剤処理
薬物代謝酵素の誘導実験は 10 nM 1α,25-dihydroxyvitamin D3もしくは 40 µM rifampicin を 分化終了 48 時間前に添加した。
3.2.7 薬物代謝酵素活性の測定
分化終了後 40 µM phenacetin、50 µM bupropion、5 µM diclofenac、47.5 µM (S)-mephenytoin、 5 µM bufuralol、5 µM midazolam および 10 µM 7-hydroxycoumarin を含む培地と共に 37℃で 24 時間インキュベートした。必要に応じ CYP3A4 の阻害剤である ketoconazole を10 μM と なるように添加した。インキュベーション後、36 μL の反応培地を回収し、内部標準を含む 氷冷した acetonitrile を 24 μL 添加することによって反応を停止させた。代謝物は、UPLC– MS/MS を用いて測定した。CYP1A、CYP2B6、CYP2C9、CYP2C19、CYP2D6、CYP3A4/5、 UDP-glucuronosyltransferase(UGT)および sulfotransferase(SULT)活性は、O-de-ethylation of phenacetin、hydroxylation of bupropion、4′-hydroxylation of diclofenac、4′-hydroxylation of (S)-mephenytoin、1′-hydroxylation of bufuralol、1′-hydroxylation of midazolam、glucuronidation of 7-hydroxycoumarin および sulfation of 7-7-hydroxycoumarin の生成量を測定することによって算出 した。分化した細胞の総タンパク質量を TaKaRa BCA Protein Assay Kit を使用して測定した。
3.2.8 取り込み試験
取り込み試験は Iwao らの報告に従って行った30)。PEPT1 の基質として glycylsarcosine を、
23
Salt Solution(HBSS)(pH 6.0)で 20 分間プレインキュベートした。その後37℃で阻害剤の 存在下または非存在下、もしくは4℃で 60 分間、135 nM [3H]-glycylsarcosine を含有する HBSS
(pH 6.0)とインキュベートした。氷冷した HBSS(pH 6.0)を添加することによって取り込 みを停止させた。細胞を洗浄し、液をすべて取り除いた後、0.5% SDS を含有する 0.2 M NaOH で細胞を可溶化した。3 mL の Clear-sol I(Nakarai Tesque、京都)を細胞溶解液に加え、液体 シンチレーションカウンター(日立アロカメディカル、東京)により放射能を測定した。分 化した細胞の総タンパク質量を TaKaRa BCA Protein Assay Kit を使用して測定した。同様な 方法で OTAP の活性を測定した。OATP の活性を測定するための基質として 5 nM [3
H]-estrone-3-sulfate を、阻害剤として 200 µM sulfobromophthalein を使用した52)。HBSS(pH 7.4)を輸 送緩衝液として使用した。
3.2.9 双方向輸送試験
分化したヒト iPS 細胞由来小腸上皮細胞および Caco-2 細胞を cell culture insert 上に播種し た。経上皮電気抵抗(TEER)値は、Millicell ERS-2(Millipore、米国)を用いて測定した。 P-gp の基質として rhodamine 123 を、阻害剤として 2 μM PSC833 を使用した53, 54)。阻害剤の
存在下または非存在下で、HBSS(pH 7.4)で 20 分間プレインキュベートした。その後、100 μM rhodamine 123 を含有する HBSS(pH 7.4)を apical 側または basal 側に添加し、阻害剤の 存在下または非存在下で、120 分間 37℃でインキュベートした。測定用サンプルをレシーバ ーチャンバー側から回収した。Rhodamine 123 の濃度は、SYNERGY HTX(BioTek、米国) を用いて定量した。Rhodamine 123 の励起波長および蛍光波長は、それぞれ 485 および 535 nm とした。Pappは以下のように算出した30)。 Papp= dQ dt × 1 A×C0
ここで、dQ/dt は単位時間当たりに透過した化合物の量であり、A は cell culture insert の表面 積(0.33 cm2)であり、C
0はドナーチャンバー中の化合物の初期濃度である。Rhodamine 123
24 輸送の Pappで割ることによって算出した。
3.2.10 統計解析
2 群間の比較には Student’s t-test 検定を用いた。多重比較には一元配置の分散分析を行っ た後、Dunnett 法を用いた。その際の統計分析は、PASW Statistict 18 system software を用いて 行った。
25
3.3 実験結果
3.3.1 ヒト iPS 細胞株 Windy における小腸上皮細胞への分化誘導に対する 8-Br-cAMP、 IBMX および Forskolin の効果
8-Br-cAMP、IBMX もしくは forskolin を添加して分化させた小腸上皮細胞において、小腸 マーカーおよび薬物動態関連遺伝子の発現解析を行った。低分子化合物を処理していない control 群と比較して、分化開始後 8–13 日に 8-Br-cAMP または forskolin を添加した群では、 小腸マーカーである Villin155, 56)および FABP257)、薬物代謝酵素である CYP3A4、薬物トラン
ス ポ ー タ ー で あ る ATP-binding cassette sub-family B member 1/multiple drug resistance 1 (ABCB1/MDR1)および solute carrier family 15 member 1/PEPT1(SLC15A1/PEPT1)2)の mRNA
発現が有意に増加した。一方、分化開始後 8–13 日に IBMX を添加した群では、小腸マーカ ーおよび薬物動態関連遺伝子の mRNA 発現は増加する傾向にあったものの有意な差は認め られなかった。また、分化開始後 13–26 日に 8-Br-cAMP を添加した群では FABP2、 ABCB1/MDR1 、 SLC15A1/PEPT1 お よ び solute carrier family 5 member 1/sodium-glucose cotransporter 1(SLC5A1/SGLT1)58)、IBMX を添加した群では Villin1、intestine specific homeobox (ISX)59)、ABCB1/MDR1、SLC15A1/PEPT1 および SLC5A1/SGLT1、forskolin を添加した群
では FABP2、ISX、CYP3A4、ABCB1/MDR1、SLC15A1/PEPT1 および SLC5A1/SGLT1 の mRNA 発現が有意に増加した(Fig. 5)。さらに 8-Br-cAMP、IBMX もしくは forskolin を添加して 72 時間後に、腸の発生に必須の転写因子60)である caudal type homeobox 2(CDX2)61, 62)および
28
3.3.2 8-Br-cAMP と IBMX を組み合わせた分化誘導法の効果(8-Br-cAMP+IBMX 群)
3.3.1 の結果から 8-Br-cAMP は分化開始後 8–14 日および IBMX は分化開始後 14–26 日に 添加することにした。小腸マーカーおよび薬物動態関連遺伝子の発現解析を行った結果、8-Br-cAMP+IBMX 群は vehicle 群と比較して、小腸マーカーである Villin1、CDX2、dipeptidyl peptidase-4(DPP4)64)、FABP2 および ISX、CYP3A4 の発現に関与する核内受容体である pregnane X receptor (PXR)、薬物代謝酵素である CYP2C9、CYP2C19、CYP3A4 および CES2A1、 薬物トランスポーターである ABCB1/MDR1、SLC15A1/PEPT1、ATP-binding cassette transporter G2/BCRP(ABCG2/BCRP)および SLC5A1/SGLT1 の mRNA 発現が有意に増加した。また、 PXR および CES2A1 に関しては adult small intestine と同程度、Villin1、CDX2、DPP4、ISX、 CYP2C9、CYP2C19、ABCB1/MDR1、SCL15A1/PEPT1 および ABCG2/BCRP に関しては adult small intestine より高い値を示した。FABP2、CYP3A4 および SLC5A1/SGLT1 に関しては adult small intestine より低かったが、Caco-2 細胞よりも高い値を示した。また、肝臓で寄与の大 きい代謝酵素である CES1A の遺伝子発現量は有意に低下した。(Fig. 7)。さらに、小腸上皮 細胞マーカーの Lactase や Villin1 だけでなく、小腸を構成する様々な細胞の一つである杯細 胞やパネート細胞のマーカーである MUC2 や Lysozyme の遺伝子発現量も vehicle 群に比較 して有意に増加した(Fig. 8)。免疫蛍光染色により小腸マーカーおよび薬物トランスポータ ーのタンパク質発現について解析した結果、小腸マーカーである villin、薬物トランスポー ターである P-gp および PEPT1 の発現が認められた(Fig. 9)。
33
3.3.3 ヒト iPS 細胞由来小腸上皮細胞の CYP3A4 誘導能と薬物代謝酵素活性
CYP3A4 の誘導剤である 1α,25-dihydroxyvitamin D3もしくは rifampicin 添加したところ、
CYP3A4 mRNA 発現レベルおよび CYP3A4/5 代謝活性は、8-Br-cAMP および IBMX で分化 したヒト iPS 細胞由来小腸上皮細胞(8-Br-cAMP+IBMX 群)において、いずれの誘導剤によ っても有意に誘導された。Vehicle 群と比較して CYP3A4/5 活性は、1α,25-dihydroxyvitamin D3
もしくは rifampicin の処理により、それぞれ 1.8 倍または 1.3 倍増加した。さらにその活性 は CYP3A4 の阻害剤である ketoconazole によって有意に阻害された。一方 Caco-2 細胞にお いて、mRNA 発現レベルおよび代謝活性は1α,25-dihydroxyvitamin D3によってのみ有意に増
加した。Caco-2 細胞における CYP3A4/5 活性は、1α,25-dihydroxyvitamin D3で処理すること
によって 1.8 倍増加した(Fig. 11)。また 8-Br-cAMP+IBMX 群では vehicle 群と比較して、 CYP2C9、CYP2C19、CYP2D6、CYP3A4/5 および UGT の代謝活性はそれぞれ 78 倍、48 倍、 1.6 倍、3.7 倍、1.9 倍増加した(Table 7)。さらに、CYP1A および CYP2B6 の活性は有意に 減少した。
35
Table 7. Drug-metabolizing enzyme activities of CYP1A, CYP2B6, CYP2C9, CYP2C19, CYP2D6, CYP3A4/5, UGT, and SULT in the differentiated enterocytes.
At terminal differentiation, the differentiated cells were incubated with intestinal differentiation medium containing 40 µM phenacetin, 50 µM bupropion, 5 µM diclofenac, 47.5 µM (S)-mephenytoin, 5 µM bufuralol, 5 µM midazolam, and 10 µM 7-hydroxycoumarin for 24 h at 37°C. The supernatants were collected and the metabolites in the differentiated cells with or without 8-Br-cAMP and IBMX in combination were measured by UPLC–MS/MS. All data are presented as mean ± standard deviation (n = 4). Levels of statistical significance compared with the vehicle group: **P < 0.01.
Vehicle 8-Br-cAMP+IBMX
CYP1A Phenacetin Acetaminophen 81.5 ± 8.4 45.8 ± 7.6** 0.56 CYP2B6 Bupropion Hydroxybupropion 6.8 ± 0.9 4.0 ± 0.8** 0.59 CYP2C9 Diclofenac 4'-Hydroxydiclofenac 0.2 ± 0.2 15.5 ± 3.4** 77.5
CYP2C19 (S )-Mephenytoin 4'-Hydroxymephenytoin 0.6 ± 0.1 28.6 ± 4.4** 47.7
CYP2D6 Bufuralol 1'-Hydroxybufuralol 0.5 ± 0.1 0.8 ± 0.1** 1.6
CYP3A4/5 Midazolam 1'-Hydroxymidazolam 23.7 ± 2.6 88.2 ± 7.9** 3.72
UGT 7-Hydroxycoumarin glucuronide 2499 ± 220 4696 ± 539** 1.88
SULT 7-Hydroxycoumarin sulfate 9949 ± 3061 5744 ± 909* 0.58 7-Hydroxycoumarin
Ratio Enzymes Substrate Metabolite
Metabolic activities (pmol/24 h/mg protein)
38 3.4 考察 cAMP シグナルは、多能性幹細胞から肝細胞、膵臓β 細胞、血管内皮細胞への分化を促進 することが報告されている48, 66, 67)。しかしながら、小腸細胞への分化に関する報告はない。 本研究は、cAMP シグナルの活性化がヒト iPS 細胞由来小腸上皮細胞の分化および成熟に寄 与することをはじめて明らかにした。ヒト iPS 細胞由来小腸上皮細胞における小腸マーカー および薬物動態関連遺伝子の発現は、分化の際に 8-Br-cAMP、IBMX もしくは forskolin を添 加することによって著しく増加した(Fig. 5)。さらに 8-Br-cAMP と IBMX を組み合わせて 使用することで 8-Br-cAMP もしくは IBMX を単独で用いるよりも、小腸マーカーおよび薬 物動態関連遺伝子の発現が増加した。そのため、これら低分子化合物を組み合わせて分化誘 導することで、小腸上皮細胞への分化がより促進することが明らかとなった(Fig. 7)。また、 免疫染色によって小腸上皮細胞に発現する villin、PEPT1、および P-gp の発現がヒト iPS 細 胞由来小腸上皮細胞において観察された(Fig. 9)。さらに、8-Br-cAMP と IBMX を用いてヒ ト iPS 細胞株 610B1 を小腸上皮細胞に分化させた場合も同様に、小腸マーカーおよび薬物 動態関連遺伝子の発現が増加した(Fig. 10)。これらの結果は、cAMP シグナルの活性化はヒ ト iPS 細胞株の違いに関わらず、小腸上皮細胞への分化促進に有用であることを示唆してい る。小腸は薬物の代謝や吸収に関わる吸収上皮細胞だけでなく、粘液を分泌する杯細胞や腸 の免疫に関わるパネート細胞など様々な細胞で構成されており、それらの細胞はクリプトの 底部に存在する小腸幹細胞から派生することが知られている。ヒト iPS 細胞由来小腸幹細胞 の分化・成熟過程において、cAMP シグナルを活性化させることで、吸収上皮細胞、杯細胞 およびパネート細胞のマーカー遺伝子の発現増加が認められた(Fig. 8)。このことから、 cAMP シグナルの活性化は、ヒト iPS 細胞由来小腸幹細胞の小腸を構成する様々な細胞への 分化誘導を促進することが示唆された。そして 8-Br-cAMP、IBMX もしくは forskolin を添加 して 72 時間後に、腸の発生に必須の転写因子60)である CDX261, 62)および GATA663)の陽性細 胞率の有意な増加が認められたことから、cAMP シグナルの分化促進効果は CDX2 および GATA6 の増加が関与していることが示唆された(Fig. 6)。一方で cAMP は様々なシグナル
39
経路に関与する。例えば cAMP-PKA シグナルの活性化によってヒストンメチル化酵素であ る G9a の発現が促進され、未分化マーカーである OCT3/4 および Nanog の発現が抑制され る報告49)や、cAMP シグナルが PEPT1 の発現増加に関与しているといった報告68)もある。 そのため、本研究で用いた低分子化合物がどのような機序でヒト iPS 細胞から小腸上皮細胞 への分化を促進するのかについては十分に明らかにできていない。しかしながら、cAMP シ グナルはヒト iPS 細胞から小腸上皮細胞への分化において重要な役割を持つことが本研究 により明らかとなった。 さらに cAMP シグナルの活性化は、遺伝子の発現量を増加させただけでなく、薬物動態学 的機能も向上させた。生体のヒト小腸は複数の CYP を発現し、様々な薬を代謝することが 知られている5, 6)。ヒト小腸での薬物代謝に対する寄与が大きいとされる CYP2C9、CYP2C19、
CYP2D6 および CYP3A4/5 および UGT の代謝活性は、8-Br-cAMP と IBMX を使用すること によって著しく上昇した(Table 7)。この結果は、cAMP シグナルの活性化がヒト iPS 細胞 由来小腸上皮細胞の薬物代謝能の向上に寄与していることを示唆している。一方、ヒト小腸 での薬物代謝に対する寄与が小さいとされる CYP1A および CYP2B6 の活性は著しく低下し た。これらの結果から、ヒト iPS 細胞から小腸細胞への分化誘導期間に cAMP シグナルを活 性化させることで、小腸上皮細胞へ選択的に分化を促進し、肝臓など他の組織細胞への分化 は抑制されている可能性が示唆された。薬物代謝酵素の誘導能についてはヒト結腸がん由来 の LS-180 細胞や Caco-2 細胞では十分に評価できていないが、ヒト iPS 細胞由来腸管細胞に おいては PXR、vitamin D 受容体および glucocorticoid 受容体などの核内受容体を介した CYP3A4 の誘導が報告されている 27)。本研究においても、8-Br-cAMP と IBMX の組み合わ せによって分化させたヒト iPS 細胞由来小腸上皮細胞において、CYP3A4 mRNA 発現および CYP3A4/5 活性が1α,25-dihydroxyvitamin D3もしくは rifampicin によって誘導された。一方、
Caco-2 細胞では、1α,25-dihydroxyvitamin D3での処理で顕著な誘導が認められたが、rifampicin
では認められなかった 69)(Fig. 11A)。さらにヒト iPS 細胞由来小腸上皮細胞における
40 果は、PXR を含む核内受容体を介して行われる CYP3A4 誘導の研究において、ヒト iPS 細 胞由来小腸上皮細胞が Caco-2 細胞よりも有用な評価系であることを示唆している。しかし ながら、本研究で作製した細胞における薬物代謝酵素活性はまだ十分に高くはないため、薬 物動態研究にヒト iPS 細胞由来小腸上皮細胞を応用するには、小腸の分化により適した細胞 株を使用し、分化誘導法をさらに改善する必要がある。 ペプチドトランスポーターである SLC15A1/PEPT1 および有機アニオントランスポーター である SLCO2B1/OATP2B1 は小腸上皮細胞に発現し、管腔内から基質の取り込みに関与し ている2)。8-Br-cAMP および IBMX を用いて分化した細胞はこれらのトランスポーターを介 した輸送活性を有していた。さらに、PEPT1 の活性は 2 つの化合物によって著しく増加した (Fig. 12)。本研究室では以前に TGF-β 受容体、DNMT、および MEK 阻害剤を用いたヒト iPS 細胞由来小腸上皮細胞への分化誘導法と、作製された細胞における BCRP などの輸送機 能について報告している24, 30)。しかしながら、その細胞は十分な P-gp 活性を有していなか った。本研究において、以前に報告した 3 つの阻害剤に加えて 8-Br-cAMP および IBMX を 使用することによって、TEER 値は増加し、より頑強な密着結合を形成した(Fig. 13A, B)。 さらに 8-Br-cAMP および IBMX を使用し作製された細胞は P-gp 活性を有していた(Fig. 13C)。Caco-2 細胞において同じ基質を用いて行った輸送試験から得られた PEPT1 および OATP の活性は、それぞれ 3.7 および 0.07 pmol/h/mg protein であった。また、P-gp の活性は、 PSC833 の非存在下および存在下で rhodamine 123 の apical-to-basal Pappは 0.49 ± 0.05 および
0.56 ± 0.01(×10-6 cm/s)であった。一方、basal-to-apical Pappは PSC833 の非存在下および存
在下で、それぞれ 2.73 ± 0.26 および 0.63 ± 0.07(×10-6 cm/s)であった。これらから算出した
ER は、阻害剤である PSC833 によって 5.7 から 1.1 に減少した。以上の結果から 8-Br-cAMP および IBMX を使用して作製されたヒト iPS 細胞由来小腸上皮細胞は、Caco-2 細胞とほぼ 同等もしくはそれ以上の取り込みおよび排出輸送の機能を有することが示唆された。そのた め、ヒト iPS 細胞由来小腸上皮細胞は、腸管吸収過程における PEPT1、OATP、P-gp および BCRP を介した輸送評価系として有用である。
41 ヒト iPS 細胞から小腸上皮細胞への分化期間中に cAMP シグナルを活性化させることに よって、小腸マーカーおよび薬物動態関連遺伝子の発現を増加させ、薬物代謝酵素および薬 物トランスポーターの活性を向上させることを見出した。したがって、この新規分化誘導法 を用いて作製したヒト iPS 細胞由来小腸上皮細胞は、以前に報告された方法により分化させ た細胞よりも優れたヒト小腸における薬物動態予測系であると考えられる。本研究は cAMP シグナルの活性化がヒト iPS 細胞由来小腸上皮細胞への分化に有用であることをはじめて 明らかにした。 3.5 小括 本研究により cAMP シグナルは、ヒト iPS 細胞から小腸上皮細胞への分化誘導に関与する 重要なシグナル経路であることが示唆された。新規の分化誘導法で作製されたヒト iPS 細胞 由来小腸上皮細胞は Caco-2 細胞と比較して、同等以上の薬物代謝酵素活性およびトランス ポーター活性を有しているため、小腸における薬物の吸収と代謝を同時に評価可能な新規評 価系としてより優れていることが示唆された。しかしながら生体のヒト小腸と比較すると、 とりわけ CYP3A4 などの薬物代謝酵素の発現量や活性は不十分であるため、小腸への分化 により適した細胞株を使用することや、さらなる分化誘導法の改善が必須である。
42 4. ヒトiPS細胞から作製した小腸上皮細胞の薬物動態学的機能解析 4.1 緒言 経口投与されたほぼ全ての薬は、小腸から吸収される際に代謝酵素に曝露されるため、薬 物動態における小腸の寄与は大きく、代謝酵素およびトランスポーターを考慮した消化管吸 収過程の評価の重要性が高まっている2–6, 46, 47)。しかしながら、ヒト正常小腸上皮細胞はタ ーンオーバーが速く機能を維持したまま培養ができないため、創薬研究に用いるのはほぼ不 可能である7, 8, 9)。消化管吸収の評価系としてヒト結腸癌由来 Caco-2 細胞が汎用されている 10–13)が、正常な組織と比べて薬物代謝酵素の発現や医薬品の透過特性が異なる14–19)ため、医 薬品の消化管吸収を正確に予測することはできない。そこでヒト iPS 細胞から機能的な小腸 上皮細胞を作製し、薬物動態試験をはじめとした創薬研究で応用することが期待されている 22–27)。これまでのヒト iPS 細胞から腸管への分化に関する報告では、主要な代謝酵素である CYP3A4 の活性が低いことが問題となっている 17, 30)。さらに薬物動態の機能に関する詳細 な解析がされておらず、実際にどのくらいの機能を有しているのか、既存の評価系である Caco-2 細胞と比べてどれほど有用であるのかよくわかっていない。最近 Akazawa らが、ヒ ト iPS 細胞由来腸管オルガノイドから腸上皮細胞を単離して cell culture insert 上に播種し、 その細胞の薬物動態学的機能についての解析結果を報告している 17)。その報告では Caco-2 細胞と比較して、CES の代謝機能や、上皮細胞の単膜層を細胞間隙経路で透過する薬の吸収 予測精度が優れていることが示唆されている。しかしながら既存の報告と同様に、作製され た腸上皮細胞は CYP3A4 の代謝機能が低いため、CYP3A4 の基質の吸収予測の評価には有 用ではないと述べられている。 当研究室ではこれまでにいくつかヒト iPS 細胞から小腸上皮細胞への分化誘導法を報告 している。TGF-β 受容体、DNMT、MEK 阻害剤を用いて分化誘導した細胞では、小腸に発 現する薬物代謝酵素および薬物トランスポーター活性に加えて、膜透過試験によって得られ た Pappとヒト消化管における Faの間に良好な相関が認められた。その 3 つの阻害剤を使用
43 害し細胞内 cAMP 濃度を高める IBMX を組み合わせて使用することで、より一層小腸上皮 細胞への分化が促進することを報告している。そしてさらなる研究によって、8-Br-cAMP お よび IBMX を組み合わせて使用する方法よりも、アデニル酸シクラーゼを活性化して細胞 内 cAMP 濃度を高める forskolin を用いた方が、より小腸上皮細胞への分化を促進すること を見出した。本章では内胚葉分化に優れたヒト iPS 細胞株である FF-1 を、TGF-β 受容体、 DNMT、MEK 阻害剤の 3 つの阻害剤と forskolin を使用した新規の方法で分化誘導し、作製 された小腸上皮細胞の薬物動態学的機能について解析した。そして、その小腸上皮細胞が実 際にどのくらいの機能を有しているのかを調べるために、ヒト初代小腸細胞70)および Caco-2 細胞と比較して得られた知見について論ずる。