大阪青山大学紀要 2012 5巻 17−20 J. Osaka Aoyama University, 2012. vol.5, 17-20
原 著
ゲルろ過クロマトグラフィーによって認められる
低分子量グリアジンの解離・会合
団 野 源 一 *
大阪青山大学健康科学部健康栄養学科Association of low molecular weight gliadins detected by gel
fi ltration chromatography
Genichi DANNO
Department of Health and Nutrition, Faculty of Health Science, Osaka Aoyama University
Summary Whole gliadins were separated on a Sephadex G-100 column with 0.1 M acetic acid or 0.01 M aluminum lactate buffer (pH 3.1) as an elution solvent. When a sample of about 95 mg protein was loaded on the column and eluted with 0.1 M acetic acid, the fraction eluted at the void volume of the column was shown to be high molecular weight gliadin by analysis of their electrophoretic patterns on sodium dodecyl sulfate polyacrylamide gel. The main peak eluted at the near-column volume was low molecular weight gliadins (and gliadins, MW about 30,000). A similar elution profi le had been reported by Hebner and Wall. In contrast, when 0.01 M aluminum lactate buffer (pH 3.1) was used as an elution solvent, the main peak was eluted at the position of the molecular weight of about 60,000. From these results, we consider that the low molecular weight gliadins associate to form dimers in 0.01 M aluminum lactate buffer (pH 3.1).
*E-mail: [email protected] 〒562-8580 箕面市新稲2-11-1
緒 論
グリアジンは、70%エタノール可溶性の小麦タン パク質である。小麦粉に水を加えて混捏すると、小麦 粉中のグリアジンとグルテニンが相互作用して粘弾性 を示すグルテンを形成する。グルテンの示す性質のう ち、その粘性の部分にグリアジンが寄与していること が知られている1,2)。 グリアジンは、70%エタノールに溶解する小麦粉の たんぱく質であり、分子量やアミノ酸組成の似かよっ た多種類のたんぱく質の混合物である。電気泳動にお ける移動度にもとづいて、α−、β−、γ−、ω−グ リアジンに分類されている3,4)。グリアジンは、セファ デックスG-100カラムを用いたゲルろ過法で3つの グループに分けられている5,6)。分子量約30,000の 低分子量グリアジン(α、β、γ-グリアジン)と分 子量約100,000の高分子量グリアジン及びその分子量 約70,000のω−グリアジンである。α−、β−、γ−、 ω−グリアジンは何れも一本のペプチド鎖からなるた んぱく質である。高分子量グリアジンは複数のペプチ ド鎖が鎖間ジスルフィド結合(S-S結合)で連結した たんぱく質である7)。本研究では、セファデックス G-100カラムによるグリアジンのゲルろ過クロマトグ ラフィーの結果が、用いた溶媒によって著しく変化す ることから、低分子量グリアジンの会合を見出したの で報告する。18 団野 源一
実験結果及び考察
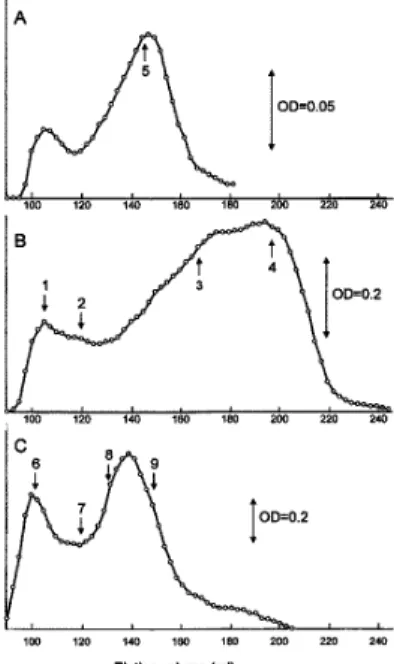
1.ゲルろ過クロマトグラフィー セファデックスG-100カラム(2.6×48 cm)を用 いたグリアジンのゲルろ過クロマトグラフィーの結果 をFig. 1 に、その溶出フラクションのSDS-PAGEを Fig. 2に示している。溶出用溶媒に0.1 M酢酸を用い て、95 mgのグリアジンをカラムに注入したときの溶 出曲線をFig. 1Bに示す。排除容積(V0)において溶 出されるピークとカラム容積(Vt)近くで溶出され る主ピークが検出された。BietzとWall7)はセファデッ クスG-100を用いたゲルろ過クロマトグラフィーで 同様な結果を報告している。溶出用溶媒に0.1 M酢酸 を用いて、15 mgのグリアジンをカラムに注入したと きの溶出曲線は、95 mg注入したときの溶出曲線と著 しく異なった(Fig. 1A)。95 mgのグリアジンを注入 したときの主ピークの溶出容積(195ml)が、15 mg の注入では140 ml付近で溶出された。同様な結果は、 Fig. 1Bの主ピークの再クロマトグラフィーでも認め られた(データ非掲載)。 溶出用溶媒に0.01 M乳酸アルミニウム緩衝液(pH 3.1)を用いて、100 mgのグリアジンをカラムに注入 したときの溶出曲線をFig. 1Cに示す。グリアジンの 注入量が多いにも関わらず、Fig. 1Aと同様の溶出曲 線が得られた。実験方法
1.試料 カナダ・ウエスタンレッド・スプリング小麦 ( Columbus )のテストミル粉( 60% extraction )を n-ブタノールで数回抽出、続いてn-ヘキサンで洗浄 した後風乾して調製したものを脱脂小麦粉として本実 験に用いた。酢酸(特級試薬)、ラウリル硫酸ナトリ ウム(SDS、99%)は、ナカライテスク(株)より購 入した。乳酸アルミニウム(一級試薬)は和光純薬か ら購入した。 2.グリアジンの調製 脱脂小麦粉から、Jonesらの方法8)に従ってグル テンを分離した。グルテンを30倍量の70%エタノー ル(v/v)に分散し、数時間攪拌してグリアジンを抽 出した。遠心分離法により不容物を除去して得たエタ ノール抽出液に1M水酸化ナトリウムを加えてpHを 6.5に調整したのち、冷蔵庫に一夜放置し、生じた沈 殿物を遠心分離法により除去した。上澄み液をヴィス キングチューブ(透析用セルロースチューブ)にいれ、 0.01 M酢酸に対して透析して含まれているエタノー ルを除去した透析内液をパーバポレーション法9)に より濃縮した後、凍結乾燥した。 3.ゲルろ過クロマトグラフィー ゲルろ過クロマトグラフィーは、0.1 M酢酸または 0.01 M乳酸アルミニウム緩衝液(pH 3.1)の二種類 の溶出用溶媒を用いて行った。セファデックスG-100 カラム(ゲルサイズ2.6 × 48 cm)を溶出用溶媒で平 衡化した。溶出用溶媒に溶解した試料グリアジンの8 ml(総たんぱく質15∼100 mg)をカラムに注入した。 流速は20 ml/hrとなるようにペリスタポンプで制御 し、溶出液はフローセルを用いた連続吸収記録計をも ちいて280 nmでの吸光度をモニターした。なお、溶 出用溶媒はオンライン脱気装置(Erma ERC 3312)を 用いて脱気し、試料溶液はMillipore fi lter(0.45 nm) で予めろ過したものをカラムに注入した。 4.SDS-ポリアクリルアミドゲル電気泳動(SDS-PAGE) SDS-PAGEは、10%ポリアクリルアミドゲル(0.75 mm厚スラブ)を使用し、Laemmliの不連続緩衝液系10) を用いて行った。ゲルは、0.04%コマジー・ブリリ アント・ブルーR250(10%三塩化酢酸に溶解)に、 35℃20時間浸漬して染色し、7%酢酸で脱染色した。Fig. 1. Gel fi ltration chromatography on a Sephadex G-100 column (2.6×
48 cm) of whole gliadin.
A: A protein of 15 mg in a volume of 8 ml was applied to the column and eluted with 0.1 M acetic acid. B: A protein of 95 mg in a volume of 8 ml was applied to the column and eluted with 0.1 M acetic acid. C: A protein of 100 mg in a volume of 8 ml was applied to the column and eluted with 0.01 M aluminum lactate buffer (pH 3.1). The arrows 1 to 9 correspond to the sample taken in Fig.2.
19
ゲルろ過クロマトグラフィーによって認められる低分子量グリアジンの解離・会合
J. Osaka Aoyama University, 2012. vol.5 カラムから溶出されるフラクションのたんぱく質組 成をSDS-PAGEにより検討した。Fig. 2の左の図は 還元していない試料の結果であり、右の図はメルカプ トエタノールによってたんぱく質のジスルフィド結合 を切断した試料の結果である。排除容積(V0)にお いて溶出されるピーク(Fig. 1B-1)は、SDS-PAGE から分子間ジスルフィド結合をもつ高分子量グリアジ ンと見なすことができる。Fig. 1B-3、4 はメルカプ トエタノールの存在の有無にかかわらず泳動パターン は変わらず、鎖間ジスルフィド結合をもたないことを 示している。カラム容積(Vt)近くで溶出される主ピー ク(Fig. 1B-4)は、単一鎖のポリペプチドからなるタ ンパク質である低分子量グリアジンと見なすことがで きる。Fig. 1A-5及びFig. 1C- 8、9も鎖間ジスルフィ ド結合は認められず、低分子量グリアジンと同じ泳動 パターンを示した。 れた。Fig. 1A及びFig. 1Cの主ピークは、先に示し たように鎖間ジスルフィド結合の存在を示さないの で、分子量30,000の低分子量グリアジンが2分子会 合したダイマーとなっていることを示唆している。
Fig. 2. SDS-polyacrylamide gel electrophoresis for samples from the fractions obtained in Fig. 1.
Left: without mercaptoethanol (2ME). Right: with 2ME. Numbers 1 through 9 correspond to the fractions in Fig. 1.
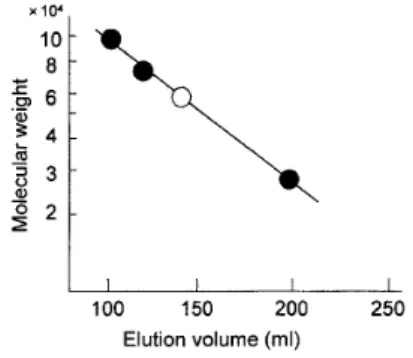
Fig. 3. Relationship between elution volume and molecular weight.
●:High molecular weight gliadin (MW 100,000), ω-gliadin (MW 70,000) and low molecular weight gliadin (MW 30,000) obtained in Fig. 1B. ○:
The peak obtained in Fig. 1A and 1C.
2.分子量 たんぱく質の注入量が少ないとき、または溶出用溶 媒として乳酸アルミニウムを用いたとき、ゲルろ過の 溶出曲線において主ピークの溶出位置が大きくシフ トすることが認められた。Fig. 1Aの主ピークの分子 量をゲルろ過法によって推定した。Fig. 1B-1、1B-2
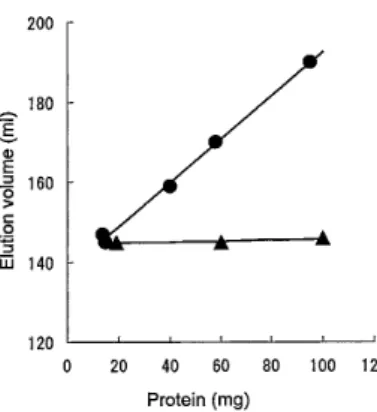
および1B-4を、Bietz and Wall7)のデータに準じて、 それぞれ高分子量グリアジン、ω−グリアジン、及び 低分子量グリアジンとみなした。Fig. 1B-1の分子量 を100,000、Fig. 1B-2の 分 子 量 を70,000、Fig. 1B-4 の分子量を30,000としてプロットした結果をFig. 3 に示す。15 mgの注入では140 ml付近で溶出された 主ピーク(Fig. 1A-5)の分子量は約60,000と推定さ 3.グリアジンの注入量と溶出容積 カラムに注入するたんぱく質量と低分子量グリアジ ンの溶出容積の関係をFig. 4に示す。 0.1 M 酢酸を溶 出用溶媒として用いたとき、低分子量グリアジンの溶 出容積は注入したタンパク質量の増加に伴いほぼ直線 的に増加することが認められた。注入タンパク質15 mgでは低分子量グリアジン(α-、β-、γーグリアジン) は二量体として、95 mgでは単量体となっていること を示している。低分子グリアジンは、0.1 酢酸中で単 量体と二量体が平衡状態を保ち、タンパク質量が少な いときは二量体の方に、タンパク質量が多くなると単 量体の方にシフトするものと推察される。他方、0.01 M乳酸アルミニウム緩衝液(pH 3.1)を溶出用溶媒と したときは、タンパク質量の影響が認められず、主と して低分子グリアジンは二量体として存在することを 示している。グリアジンの主要成分である低分子グリ アジンが環境条件によって解離・会合をおこなうこと は、小麦粉からグルテンの形成に、またグルテンの加 工適性に何らかの影響を与えているものと推察した。
20 団野 源一
文 献
1) Kasarda DD, Nimmo CC, Kohler GO, Proteins and the amino acid composition of wheat fractions, in Wheat chemistry and technology, 2nd ed. Y. Pomeranz. Ed. Am. Assoc. Cereal Chem., St Paul. MN, 1971, 227-99.
2) 団野源一, 小麦タンパク質の構造と物性、食品
タンパク質の科学―科学性質と食品物性.山内 文男編(株)食品資材研究会.1983, p71-103.
3) Joones, R. W., Taylor, N. W. and Senti, F. R. Electrophoresis and wheat gluten, Arch Biochem Biophys. 1959, 84, 363-76.
4) Woychik JH, Boundy JA, Dimler RJ, Starach gel electrophoresis of wheat gluten proteins with concentrated urea, Arch. Biochem. Biophys.1961,
94, 477-82.
5) Mecham DK, Kasarda DD, Qualset CO, Genetic aspects of wheat gliadin proteins, Biochem Genet. 1978, 16, 831-53.
6) Bietz JA, Wall JS, Wheat gluten subunits:
molecular weights determined by sodium dodecyl sulfate-polyacryl amide gel electrophoresis, Cereal Chem. 1972, 49, 416-30.
7) Bietz JA, Wall JS, Identity of high molecular weight gliadin and ethanol-soluble glutenin subunits fo wheat: relation to gluten structure, Cereal Chem. 1980, 57, 415-21.
8) Jones RW. Babcock GE, Senti RR, Electrophoresis and fractionation of wheat gluten, Arch Biochem Biophys. 1959, 84, 363-76.
9) 岩永貞昭, 山下仁平, 佐野 朗, 笹川 滋.タン パク質の抽出,溶解度を利用した分別法:生化
学実験講座1, タンパク質の化学Ⅰ‐分離生成‐,
日本生化学会編, 東京化学同人, 1976, 11-98
10) Laemmll UK, Cleavage of structural proteins during the assembly o f the head baacteriopgage T4, Nature. 1970, 227, 684.
Fig. 4. Relationship between amount of protein applied on a column and elution volume of low molecular weight gliadin.
●:0.1 M acetic acid used as an elution solvent, ▲:0.01 M aluminum lactate buffer (pH 3.1) use as an elution solvent.