応用微生物学研究室雑誌会 No.1210 2017/06/23 M1 中村匡
大腸菌における酢酸代謝制御
背景 大腸菌は、過多炭素源を酢酸に変換して蓄積し、炭素欠乏となると蓄積した酢酸を代謝する。自身が蓄積した 酢酸により大腸菌の生育は阻害され、これは大腸菌を工業利用する時には好ましくない現象である。また、酢酸 の生成、排出によりバイオマス量の減少、組み換えタンパク質、低分子化合物の合成阻害も同時に引き起こす。 大腸菌の酢酸生成、消費の切り替え機構は未だ不明である。また、私が研究しているAcetobacter aceti、Acetobacter pasteurianusにおいてもエタノール条件下、エタノ ール&グルコース条件下において酢酸代謝切り替えが観察できる(1,2)。そのため、今回は大腸菌における酢酸代 謝制御についての研究への様々なアプローチを調べることにした。
本雑誌会では、酢酸代謝に関わる経路とその下流にあるTCA 回路の制御に注目した。一報目では酢酸代謝制 御と内部環境との関係、二報目では外部環境との関係について紹介する。
Fig. 0 (A) Central carbon metabolic pathway of A. aceti grown on ethanol (Sakurai et al., 2012)
Red arrows indicate that the genes mediating reactions were upregulated in cells compared with grown on glucose. Blue arrows indicate that these were downregulated.
2
Appied Microbiology and Biotechnology 98 (2014) 5131-5143 より
Pta-AckA 経路を破壊した場合、酢酸排出量は減少するが、比増殖速度(以下μ)、バイオマス収量(以下 YXS)は 低下し、副次的に乳酸、ギ酸の排出量が増加する。また、二番目に主要な酢酸生産の経路であるPoxB 経路を破 壊した場合も、好気条件下における生育効率が低下することがわかっている。これらの結果の通り、酢酸オーバ ーフローを防止するために単に酢酸生合成経路を除去しても好ましくない副作用が生じてしまう。 大腸菌はあらゆる比増殖速度条件下において、好気ならば常に酢酸を生産すること、低μ条件下では、酢酸は Pta-Acs 経路によって完全にリサイクルされるために酢酸オーバーフローが起こらないこと、が様々な研究グル ープによって主張されている。そこで更に本研究者らは、高μ条件下における酢酸排出は酢酸・グルコース同時 利用能力の損失と関係し、Acs の不活性化による Pta-Acs 経路の破壊が原因であるという仮説を立てた。 本研究では、Pta-Acs 経路と酢酸オーバーフローの関連性を更に深く調べることが目的である。そのために、 本研究者らはA-stat と呼ばれる連続培養法を選択し(3)、様々な特定のグルコース消費速度(以下 qglc)において、 E. coli K-12 BW251113 とその突然変異株の増殖を観察、解析した。
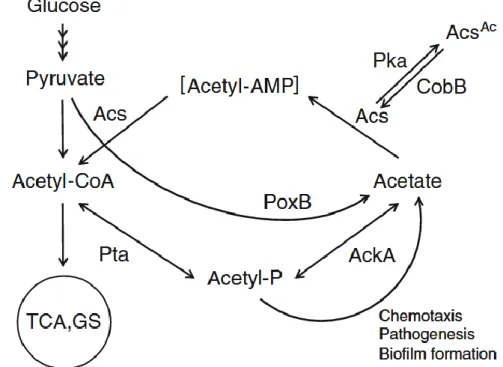
Fig. 1 Simplified overview of glucose metabolism, PTA-ACS node and regulation of Acs by Pka and CobB in E. coli. Acetate is mainly generated from: (a) acetyl-CoA by Pta and AckA enzymes via the high energy intermediate acetyl-phosphate (acetyl-P); (b) acetyl-P during different cellular regulation processes; (c) pyruvate by PoxB. Acetate in the PTA-ACS node is reactivated by recycling it back to acetyl-CoA by the high-affinity Acs enzyme. Activity of Acs is essential for functioning of the PTA-ACS node: protein lysine acetyltransferase (Pka) inactivates Acs by acetylation (AcsAc), while the NAD+-dependent protein deacetylase CobB reactivates Acs by deacetylating it. Pta phosphotransacetylase, AckA acetate kinase, Acs acetyl-CoA synthetase, PoxB pyruvate oxidase, TCA tricarboxylic acid cycle, GS glyoxylate shunt
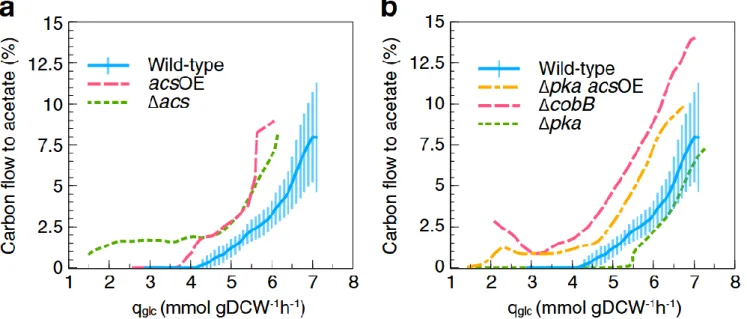
Acs の欠損、過剰発現による酢酸代謝への影響 (Fig. 2a)
WT では、qglcが4.2 mmol gDCW-1 h-1に達するまでは酢酸排出は見られなかったが、Δacsでは全てのqglc
値において酢酸排出が確認された。これは、大腸菌が低μ条件下でも酢酸を生成しているという仮説を裏付けて いる。また、ΔacsはWT よりも多くの炭素を酢酸に変換するため、最大 qglcはWT よりも低く、YXSの平均も
5%程少なかった。
Acs 過剰発現株(以下 acsOE)を作製し、WT と比較したところ、acsOE における酢酸オーバーフロー開始時の qglc値は低く、全てのqglc条件下においてより多くの炭素を酢酸として排出していた。また、Δacs同様、最大
qglcつまり最大μはWT よりも低かった。タンパク質の過剰発現が要因となり、ほとんどの Acs タンパク質が不
活性化したと推測された。
酢酸代謝におけるタンパク質のアセチル化の役割 (Fig. 2b)
Pka は Acs をアセチル化することで不活性化させ(4)、CobB は Acs を脱アセチル化することで活性化させる(5)。 また、Acs は酢酸と ATP の存在下で自己アセチル化が可能であり(6)、Acs タンパク質活性に関わるアセチル化 の重要性が伺える。したがって、アセチル化の酢酸代謝への影響を調べるために、ΔpkaとΔcobBが作製され た。 ΔcobBはアセチル化したAcs を脱アセチル化により再活性化させることができないため、全ての qglc値にお いてWT に比べて多くの酢酸を排出し、YXSは平均して28%低かった。また、Δacsと同様に炭素の酢酸へのフ ラックスはqglc値が4 mmol gDCW-1 h-1に達するまでは安定していた。 Δpkaにおいて、酢酸オーバーフロー開始が遅れたが、これは細胞内の不活性化Acs タンパク質の割合が減少 し、Pta-Acs 経路によるリサイクル率が高まったためと考えられた。
acsOE のpkaを破壊してΔpka acsOE を作製し、Acs の過剰発現が酢酸排出量の減少をもたらさないのは Acs タンパク質のほとんどが不活性化してしまっていることが原因である可能性を確認した。結果としては、Δpka acsOE は WT と比べて酢酸オーバーフロー開始が遅れることはなく、酢酸排出量も WT より多かった。
4
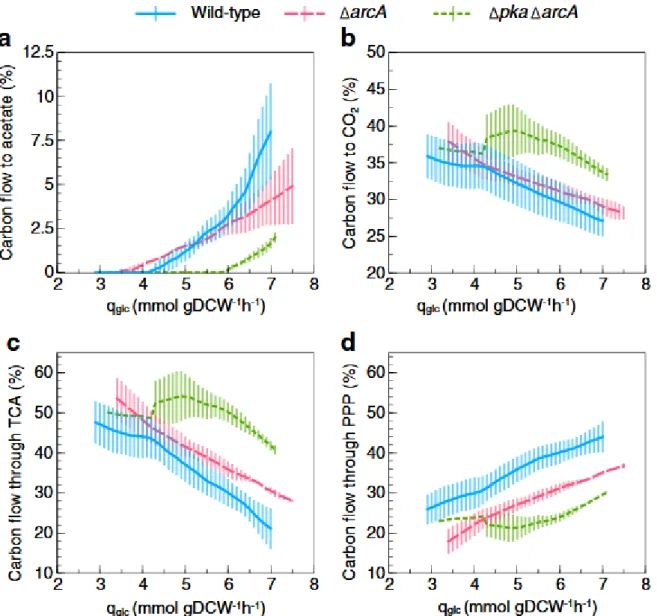
高レベルの活性化Acs と TCA サイクルが酢酸オーバーフローを強く低減させる (Fig. 3)
Acs の過剰発現は酢酸オーバーフローの緩和に繋がらなかったが、Δpkaにおいては酢酸オーバーフローの減 少が確認できたため、Δpka acsOE ほどではなくとも、Acs の発現レベルが少しでも高ければ、Δpkaならば更 に酢酸オーバーフローを減少させることができると考えられた。しかし、ΔpkaにおいてAcs の活性レベルが高 くても、下流の経路(例えば TCA サイクル)が制限されていると、リサイクルされたアセチル CoA が再び酢酸へ と流れる可能性がある。そこで、下流の経路を向上させる方法も検討された。
ArcA を欠失させると TCA 回路関連遺伝子の発現が促進され、更に WT と比較してacsの転写量が約2 倍に なる(7,8)。従ってΔpkaΔarcAが作製された。この二重ノックアウト株は酢酸オーバーフロー開始時のqglc値が
6 mmol gDCW-1 h-1の段階まで遅れ、WT に比べて酢酸に消費される炭素量は 4 分の 1 以下であった。
ΔpkaΔarcAは酢酸の代わりにCO2を多く生成していることが示された。これはarcAの欠失によってTCA
サイクルへの炭素流入の増加の結果であると考えられた。また、ΔpkaΔarcAではペントースリン酸回路(PPP) を経て生成されるCO2が減少し、酢酸代謝時のTCA サイクルと PPP とのバランスが示された。Pta-AckA 経路、
あるいはpoxBが破壊された株と異なり、ΔpkaΔarcA株はWT と同程度のμmaxを示し、WT と比較して約 5%
低いYXSしか示さなかった。
本研究者らは、ΔpkaΔarcAにおける酢酸オーバーフローの減少がarcAの欠失のみでも生じるかを確認する ためにΔarcAの生育を分析した。ΔarcAではWT よりも酢酸オーバーフローの開始が早かったため、ΔpkaΔ arcAにおける酢酸オーバーフローの減少はarcAの欠失だけでは説明できない。また、ΔarcAはWT に比べて 酢酸生成が遅く、最大qglcにおいて酢酸へと流れた炭素はWT よりも少ない。ΔpkaΔarcAと比べてΔarcAの
CO2生成量は少ないが、これはΔpkaΔarcAがΔarcAに比べてより高効率に酢酸をリサイクルし、より多くの
アセチルCoA を TCA サイクルに供給するためだと思われる。このことから、TCA サイクルの働きを向上させ る際のΔpkaの重要性が示された。
Fig. 3 Specific glucose consumption rate (qglc)-dependent carbon flow through metabolism in E. coli K-12 BW25113 wild type and its acetate overflow-reduced mutants. The percentage of carbon flow from glucose to acetate (a) and CO2 (b). The percentage of carbon flow from glucose through TCA cycle (c) and pentose phosphate pathway (PPP) (d) represented by suc and gnd fluxes, respectively, expressed as the C-molar percentage of flux through the reaction from the consumed carbon. Wild type (blue solid line), ΔarcA (pink dashed line), Δpka ΔarcA (green dotted line). Lines for each strain are best-fit splines of 13–43 glucose and product concentration measurements in each experiment. Error bars represent standard deviation of four and two independent A-stat experiments for wild type and ΔarcA, Δpka ΔarcA, respectively
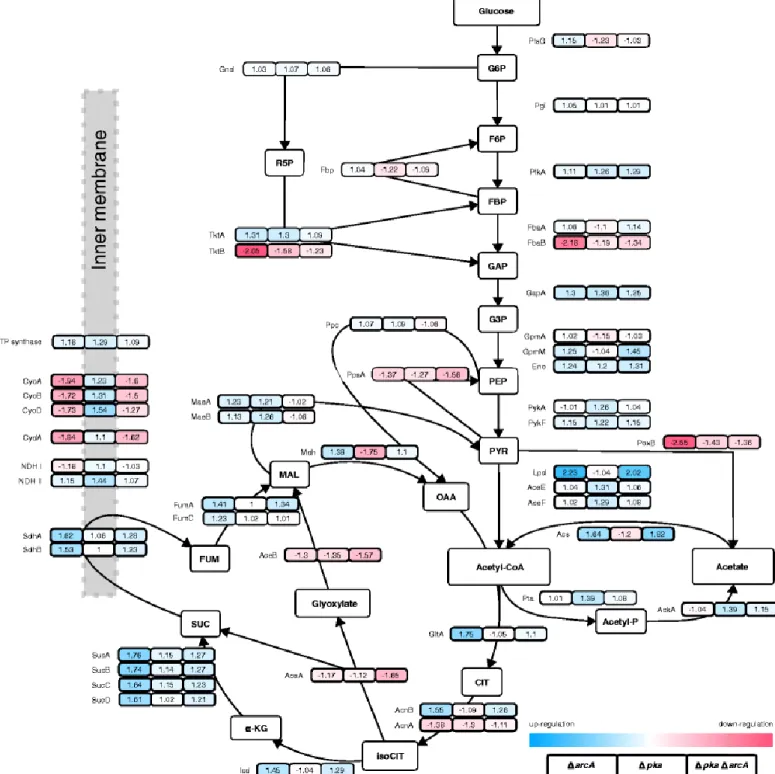
6 異なる遺伝子破壊株におけるプロテオーム解析 (Fig. 4)
Δpka、ΔarcA、ΔpkaΔarcAおよびWT についてプロテオーム解析を行った。
ΔarcAのTCA サイクルフラックスは qglc値が5 以下の時点でΔpkaΔarcAと比較して低かったが、TCA サ
イクルのタンパク質濃度はΔarcAで上昇している。これは翻訳後調節によって酵素の触媒速度が制御されたた めと考えられる。翻訳後調節によって触媒速度を変化させて代謝を調節する方法は大腸菌において一般的である という近年の研究と一致する。
また、arcAの欠損によりTCA サイクル関連遺伝子および呼吸鎖関連遺伝子の転写が 7 倍増加することが判明 している。しかし、TCA サイクル関連酵素の量は 1.5 倍程度にしか増加しなかった。さらにΔarcA、ΔpkaΔarcA についてシトクロムの発現の低下を確認した。
先行研究において、酢酸オーバーフローを減少させるためにはAcs のみならずグリオキシル酸経路の発現の増 加も必要であると判明している(9)。しかし本研究者らはΔpkaΔarcAにおいてグリオキシル酸経路タンパク質 AceA、AceB の発現の低下を確認したが、ΔpkaΔarcAは著しい酢酸オーバーフローの減少を示した。先行研究 と本研究では大腸菌株が異なる。先行研究では連続培養ではなく、これらの差異が原因である可能性が考えられ た。
Fig. 4 Proteome expression of E. coli K-12 BW25113 ΔarcA (left box), Δpka (middle box) and Δpka ΔarcA (right box) compared to wild type at μ = 0.4 h−1. Numbers represent fold change of protein concentration compared to wild type; down-regulation is shown in red and denoted by negative values, up-regulation in blue and by positive values. Colour intensity is proportional to fold change and scaled by the maximum and minimum protein concentration fold change on the figure. Proteins aligned on the
grey area are known to be associated with cell membrane. NDH I and ATP synthase represent the average fold change of NADH dehydrogenase and ATP synthase complex protein concentrations, respectively. Arrow heads show the presumed reaction direction in given conditions
8 Discussion
まず本研究者らは、Δacs、ΔcobBがWT と異なり低μ条件下でも酢酸を排出することを観察し、低μ条件下 においても酢酸が生合成されることを確認した。これは酢酸がPta-Acs 経路で完全にリサイクルされるため、大 腸菌の生育が遅い間に活性Acs が酢酸オーバーフローを減少させるという予測を支持する。更にΔacs、ΔcobB は酢酸排出が増加し、WT と比較して YXSが低かった。酢酸生成の増加による炭素のロスだけでは観察された
YXSの低さは説明できない。
活性型Acs の欠如が酢酸オーバーフローを引き起こす事実と、Acs の抑制によって qglcの増加に伴う酢酸蓄積
が発生するという仮説を考えると、Acs を高発現させれば酢酸オーバーフローを減少させることができると予測 できる。しかし、本研究によるとacsOE 株では酢酸オーバーフローの減少は見られなかった。更に acsOE の Acs が不活性化している可能性は、Δpka acsOE が WT よりも多くの酢酸蓄積を示したことで否定された。本研 究者らはAcs の過剰発現が Pta-Acs 経路による ATP の過剰消費を引き起こし、あるいは Acs タンパク質合成の ためにエネルギーを浪費させ、細胞に強い代謝負荷を発生させ、エネルギーバランスを崩していると推測した。 エネルギーバランスが崩れた結果、エネルギー生成目的のために低qglcにおいても酢酸を生成、排出したと考え られた。 本研究により、Δpkaでは酢酸オーバーフローが遅延したことで、Acs を抑制することで酢酸オーバーフロー が生じると考えられた。プロテオーム解析ではΔpkaはWT と比較して低い Acs 濃度を示し、中心炭素代謝に変 化は確認できなかった。よってΔpkaにおいて酢酸オーバーフローが減少した原因は、活性化Acs の量が増加し ていたためと思われた。更に本研究では、単純な遺伝子過剰発現だけではあらゆる条件で狙った効果を達成する ことはできず、下流の代謝経路と共にタンパク質の活性/不活性に関連する部分の発現レベルも微調整する必要が あることを示した。
ΔpkaΔarcAにおいてはシトクローム、グリオキシル酸経路関連酵素の発現は低く、また、TCA サイクル関 連酵素の高発現を示したΔarcAにおいてΔpkaΔarcAやWT と比較して高い酢酸排出が観察された。以上の結 果から、本研究者らは呼吸鎖の抑制、TCA サイクル代謝物、グリオキシル酸経路活性は酢酸オーバーフローと 無関係であると主張し、高濃度の活性化Acs による Pta-Acs 経路の酢酸リサイクル能の増加と TCA サイクルと の間のバランスが酢酸オーバーフローの制御に重要であると結論付けた。
一報目について
acsOE では酢酸オーバーフローの減少は見られず、Δpka acsOE においても酢酸オーバーフローが増加した ためにAcs の不活性化が原因という考えは本研究者らに否定された。しかし、Acs は自己アセチル化が可能であ るため、その部分の考察、実験をするべきだと感じた。また、acsOE において Acs の過剰発現がエネルギーを 浪費させ、エネルギーバランスを崩しているという仮説についてははっきりしないという印象を受けた。タンパ ク質の過剰発現が問題であり、Acs であることが問題でなければ、別の適当なタンパク質を過剰発現しても低 qglc
10 SCIENTIFIC REPORTS 7 (2017) 42135 より
グルコース過剰条件下で生育する大腸菌は酢酸を生成し、グルコースが完全に消費された後に酢酸を消費する (10)。また、グルコース過剰条件下では、大腸菌における主なグルコース輸送系である PTS の EIIA 成分はほと んど非リン酸化状態で存在するが、これはアデニリルシクラーゼを阻害する。結果としてcAMP レベルは低く、 acsの転写活性化に必要なcAMP 受容体タンパク質(CRP)も不活性のままである。Acs 発現が抑制されているた め、グルコースでの増殖期間中は酢酸消費が止まり、グルコースを完全に消費すると、酢酸消費が可能になる。 しかし、異化代謝抑制機構の一部が欠失した場合はグルコースと酢酸の同時消費が観察される。 このようにカタボライト抑制のために通常ではグルコースと酢酸の同時消費は起こらないと予想される。しか し最近の研究において、E. coli K-12 野生株がグルコース&酢酸混合条件下において対数期で酢酸を取り込み、 代謝することが観察されている。カタボライト抑制のためにAcs が発現しないことから、この観察結果は酢酸が グルコース過剰時に別の経路で依然として消費されていることを示唆している。よって、カタボライト抑制のみ が酢酸利用の可不可の条件ではない可能性が高い。
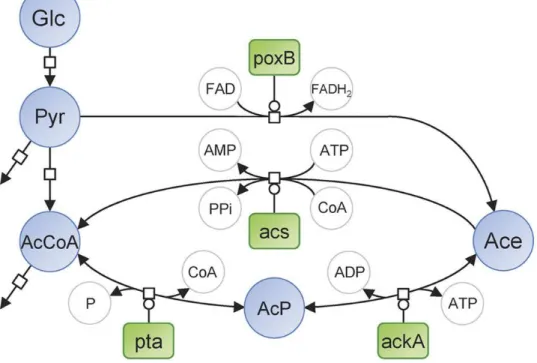
Figure 1: Representation of E. coli pathways involved in acetate metabolism, in Systems Biology Graphical Notation format (http://sbgn.org). Circles represent metabolites and rounded rectangles represent enzymes.
本研究の目的は、過剰グルコース条件下、つまりカタボライト抑制条件下で酢酸が消費される経路を明らかに すること、酢酸代謝の制御機構を解明することであった。そのために本研究者らは酢酸生成量と消費フラックス を個別に定量化し、代謝経路を同定するために13C ラベリング実験を行った。
グルコース条件下における酢酸のTCA サイクルにおける代謝 (Fig. 2) 13C 標識された酢酸をグルコース消費期、または酢酸消費期の培養液に添加した。酢酸消費期では、13C は全 ての中心炭素代謝産物において観察された。一方グルコース消費期では、TCA サイクル中間体において観察さ れ、解糖系の中間体では観察されなかった。これは、グルコース消費期において糖新生経路の活性が低いことと 一致する。これらの結果より、グルコース過剰条件下でも細胞外酢酸は取り込まれ、代謝されることが確認され た。グルコース消費時の酢酸蓄積は酢酸生成と消費のバランスから生じ、細胞と培地の間で双方向に酢酸を交換 していることがわかる。
Figure 2: Incorporation of 13C atoms in central metabolites after a pulse of 13C-acetate during exponential growth on glucose or during the acetate consumption phase. E. coli K-12 MG1655 was grown on M9 supplemented with 15 mM glucose. 13C-acetate was added at a final concentration of 1 mM in mid exponential phase (dark grey bars) or during the phase of net acetate consumption (light grey bars). Intracellular metabolites were collected before each isotopic switch and one hour after the addition of labeled acetate, and the molecular 13C-enrichments of central metabolites were quantified by mass spectrometry. Results are displayed as the ratio between the 13C-enrichments of metabolite before and after the pulse. Asterisk (*)
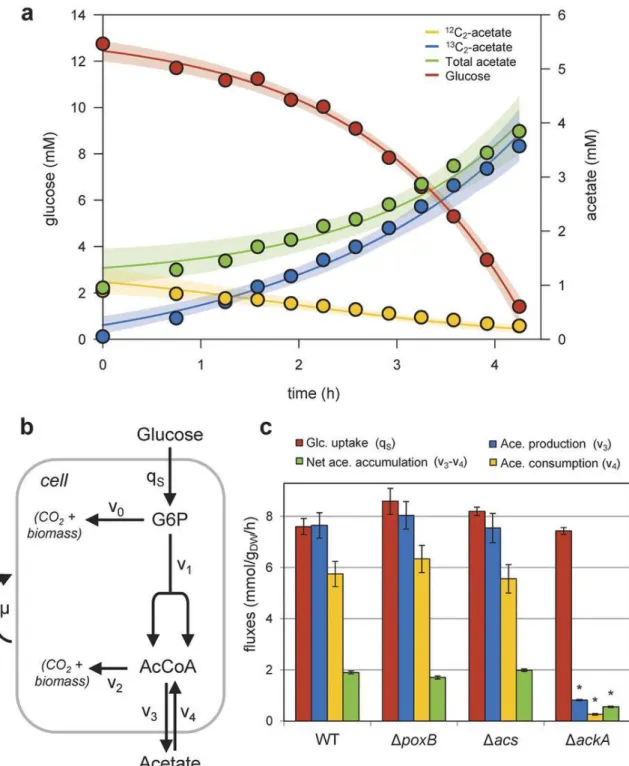
12 Pta-AckA 経路によって同時に行われる酢酸の生成と消費 (Fig. 3) グルコース過剰条件下における細胞-培地間の酢酸の移動の度合いを評価するために、酢酸生成と消費のフラッ クスを調査した。13C 標識された過剰グルコースと非標識の酢酸との二成分を含む最小混合培地で大腸菌を増殖 させた。増殖とグルコース消費に伴い、標識酢酸は増加したが、非標識酢酸は減少した(Fig. 3a)。そこで、酢酸 生成と消費をシミュレーションするモデルが開発された(Fig. 3b)。シミュレーションと上記の実験データによる と、単方向の酢酸生成、および消費量はそれぞれ7.7±0.5、5.7±0.5 mmol gDW-1h-1であり、正味の酢酸蓄積速 度2.2 mmol gDW-1h-1の3~4 倍に相当し、WT ではグルコース消費速度と酢酸生成速度の値が近いとわかった。
Pta-AckA 経路と Acs 経路はそれぞれ異なる補因子を必要とする。アセチル CoA-酢酸サイクルの補因子バラン スは酢酸生成、消費経路に依存するので、これらの経路は細胞内のエネルギーバランスに関わっていると言える。 よって、同時に行われる酢酸生成と消費の細胞内エネルギーへの影響を評価するために、各欠乏株において酢酸 フラックスを検証した(Fig. 3c)。WT、ΔacsおよびΔpoxBにおけるあらゆる酢酸フラックスは同等だった。し かしΔackAにおける正味の酢酸蓄積、生成、消費量はWT と比較して著しく減少した。これらの結果より、 Pta-AckA 経路が酢酸生成、消費に関与している一方、Acs も PoxB も今回の条件では酢酸消費に関与していな いことが判明した。Pta-AckA 経路のみでも十分に双方向の酢酸フラックスを維持可能であり、また、
Pta-AckA-Acs サイクルと異なり、Pta-AckA 経路の双方向プロセスのみならば ATP の浪費は無く、細胞内のエ ネルギーバランスを崩すことはないはずである。
Figure 3: Quantification of acetate consumption and production fluxes by dynamic 13C-metabolic flux analysis.
(a) Time-course profiles of glucose and total, labeled and unlabeled extracellular acetate concentrations. E. coli was grown on M9 supplemented with 15 mM 13C-glucose and 1 mM unlabeled acetate. The concentration of the four isotopomers of acetate was quantified by NMR every 30 minutes until the cells had consumed all the glucose (red circles). Unlabeled and 1,2-13C2-acetate are shown in yellow and blue, respectively. Concentrations of 1-13C1- and 2-13C1-acetate remained negligible (<0.1 mM) during the experiment. Total acetate concentration (in green) was calculated by summing the concentrations of all its isotopomers. Circles represent measurements, lines represent the best fit with their 95% confidence intervals (shaded areas). (b) Graphical representation of the model used to quantify acetate production and uptake fluxes. Details on this model
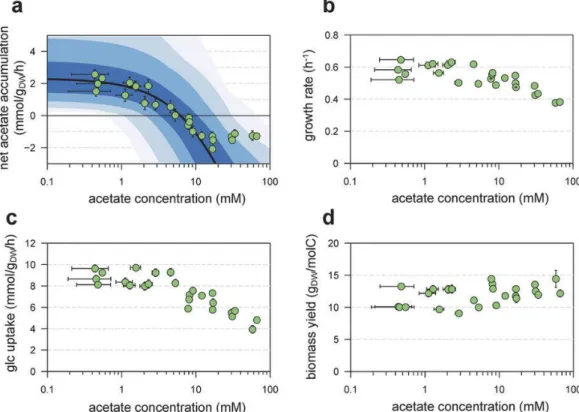
14 酢酸、グルコースの同時消費 (Fig. 4) Pta-AckA 経路は、細胞外酢酸濃度の増加をきっかけに酢酸生成の低下に向かう可能性があり、更には酢酸消 費に切り替わる可能性がある。この問題について定量的に調査するためにPta-AckA 経路の動力学的モデルを開 発し、広範囲の酢酸濃度条件について経路の動態のシミュレーションを繰り返した。実験による観察と同様、酢 酸濃度が低い培地において酢酸蓄積が起こると予測した。Pta-AckA フラックスは酢酸濃度増加に伴い減少し、 予測値では6 mM、多くとも 10 mM を超えると酢酸消費量が酢酸生成量を上回ることが予測された(Fig. 4a)。 グルコースと異なる濃度の酢酸を添加した最小培地で大腸菌を増殖させ、増殖速度、グルコース消費速度、酢 酸生成速度を実際に測定した(Fig. 4a,b,c)。増殖速度、グルコース消費速度は 8 mM 酢酸濃度まで安定であり、 その後酢酸濃度の増加につれて減少した。一方、正味の酢酸蓄積量は培地の酢酸濃度が1 mM を超えるとすぐに 低下し、また5.5 mM を超えると 0 になり、酢酸消費が酢酸生成を上回ったことを示した。これは動力学的モデ ルの予測値6 mM と近い。32 mM を超える酢酸濃度では、培地中の酢酸消費速度は安定した。これは細胞内酢 酸の飽和、あるいは他の代謝によるフィードバック阻害を示唆している。これらの結果より、グルコースと酢酸 は同時に消費されることを示した。 酢酸が高濃度の場合、細胞の生育が阻害される一方、バイオマス収量は減少しない(Fig. 4d)。これは Pta-AckA 経路の双方向のフラックスのみではエネルギーが大量に拡散されることはない、という事実を裏付けている。
Figure 4: In silico analysis of the Pta-AckA pathway, and experimental validation of model predictions. The steady-state flux through the Pta-Ack pathway was simulated for different acetate levels (panel a, black line), with kinetic parameters taken from the literature and concentrations of ADP, ATP, CoA, acetyl-CoA and phosphate set to experimental values. To investigate the robustness of these predictions, sensitivity analysis was carried out by uniformly sampling all concentrations and
parameters within ±10%, ±20%, ±30% and ±40% of their experimental values (dark blue to light blue, respectively). These predictions were compared to experimental values (green dots). Experimental growth rates (b), glucose uptake rates (c) and growth yields (d) for different acetate levels.
グルコース存在条件下の酢酸消費におけるPta-AckA 経路の役割 (Fig. 5)
Pta-AckA 経路がグルコース過剰条件下での酢酸消費に関係しているか確認するため、WT、Δacs、ΔpoxB およびΔackAにおいて異なる酢酸濃度条件の下、酢酸の正味の蓄積量、酢酸生成、消費フラックスを測定した。 WT、ΔpoxB、Δacsでは、酢酸濃度が8 mM 以上で正味の酢酸蓄積量が 0 以下になった。対照的に、ΔackA では酢酸生成、消費フラックスが減少し、酢酸蓄積は依然として観察された。これは、アセチル-P の AckA 非依 存的な酢酸への変換、あるいは酢酸が代謝副産物として生じる生合成経路が原因と考えられた。また、酢酸の生 成、消費についてAcs、PoxB は重要な役割を持たず、Pta-AckA 経路が不可欠であることを確認した。
Figure 5: Net acetate accumulation flux and unidirectional production and consumption fluxes in E. coli K-12 MG1655 WT, ΔackA, Δacs and ΔpoxB grown on glucose and different acetate concentrations. Dynamic 13C-metabolic flux analyses were
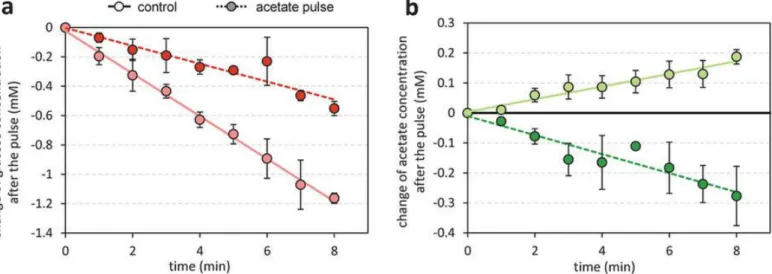
16 Pka-AckA 経路の代謝制御 (Fig. 6) 大腸菌をグルコース条件下で増殖させ、対数期中間で酢酸を添加し、その後のグルコース、酢酸濃度を測定し た。グルコース消費速度は、酢酸添加後に減少した。また、何も添加しない条件では酢酸濃度は上昇する一方、 酢酸を添加した場合は1~2 分後に酢酸濃度の減少が見られた。つまり、酢酸の添加は酢酸の生成と消費の急激な 逆転を誘発した。
Figure 6: Short-term response of the Pta-AckA flux to an acetate pulse. E. coli was grown on M9 supplemented with 15 mM glucose. Changes in concentration of glucose (a) and acetate (b) were measured in mid-exponential growth phase with (dark green line) or without (control experiment, light green line) addition of acetate (at a final concentration of 30 mM) in mid-exponential growth phase.
共基質としての酢酸 (Fig. 7)
酢酸が培地中に十分に高濃度に存在する場合、グルコースと酢酸を共基質として利用できることが判明した。 グルコン酸、フコースにおいても酢酸を共基質として利用できるか調べた。結果、グルコース同様、グルコン酸、 フコースからも酢酸が生成され、酢酸を基質として同時に使用した。よって、酢酸はその細胞外濃度のみに依存 して共基質になりうることが示された。
Figure 7: E. coli can co-consume acetate and glycolytic carbon sources. E. coli was grown on M9 supplemented with 15 mM gluconate or fucose. Growth rates, glycolytic substrate uptake rates and acetate accumulation rates were measured with or without acetate added initially to the medium (at a concentration of 32 mM). Asterisk (*) represents a p-value lower than 0.01 when compared to the same condition without acetate added initially to the medium.
Discussion
大腸菌は過剰グルコース条件下において、酢酸とグルコースを同時に消費できることが示された。この酢酸消 費の現象にacsやpoxBは寄与していない。本研究者らはPta-AckA 経路のみが過剰グルコース条件下のおける 酢酸生成、消費に寄与していることを示した。過剰グルコース条件下においてPta-AckA 経路の酢酸生成、消費 は基本的に酢酸の細胞外濃度により制御されている。本研究の実験データ、動態モデルによるシミュレーション から、細胞外酢酸濃度が閾値を超えると代謝が酢酸生成から酢酸消費に切り替わることが明らかになった。 Acs はカタボライト抑制によって制御される。これは Pta-AckA-Acs サイクルの働きを阻害し、余計なエネル ギーロスを抑える。一方、Pta-AckA は酢酸の細胞外濃度の変化に反応し、酢酸生成と消費のバランスを微調整 することができると判明した。Pta-AckA 経路の酢酸濃度の変化への応答時間は Acs など、段階的に発現を調節 するものよりもはるかに速い。この迅速な応答は特定の環境、条件における優位な生育をサポートしている可能 性がある。また、Pta-AckA-Acs サイクルによる酢酸同化は限られた炭素条件で酢酸を資化するには効果的だが、 高コストである。一方、炭素が十分に存在する条件では、Pta-AckA 経路は低コストで酢酸を資化できる。 Pta-AckA 経路の中間体であるアセチル-P は他のタンパク質をリン酸化、アセチル化することで細胞内の多く のプロセスを調節できることが知られている。つまり、酢酸の細胞外濃度の変化がPta-AckA 経路を通じてアセ チル-P 量を制御し、細胞内プロセスを調節している可能性がある。更に、酢酸利用可能性による代謝の微調整に ついてもアセチル-P を介している可能性が考えられる。 本研究により、細胞外の酢酸濃度が十分に高い場合、グルコース過剰条件下においても酢酸を消費することが 可能であると示されたが、これは糖枯渇後に酢酸が消費されるという従来の考え方とは対照的である。グルコー ス条件において、細胞外酢酸濃度が8 mM の時、酢酸生成は起こらない。この状態ではグルコース消費量、生育 速度は変わらず、酢酸生成がエネルギー節約、有害有機酸量の制御のためではないということが示された。実験 室においてグルコース条件で大腸菌を生育させる場合、一般的には酢酸の蓄積が少ない条件で行われ、これは酢 酸生成が優勢となる条件である。しかし実際には腸内の酢酸濃度は高く、8 mM の閾値をはるかに上回っている。 このような条件においては酢酸消費が優勢となる。従って、腸内において大腸菌は糖と酢酸を同時消費している 可能性が高い。 二報目について Fig. 4 の部分で Pta-AckA 経路の正味の酢酸蓄積量をわざわざ動力学的モデルでシミュレーションした本研究 者らの意図が理解できなかった。Pta-AckA 経路の中間体にアセチル-P が存在するので、細胞外の酢酸濃度がア セチル-P 量を変化させ、細胞内プロセスの調節に影響している可能性は高いと考えられた。細胞外酢酸とアセチ ル-P の関わる各細胞内プロセスの関係に注目してみてもいいかもしれない。また、今回は実験室環境と腸内環境 の違いを強く意識する結果となった。大腸菌に限らず、自然環境条件を意識することで細胞内動態の生化学的意 義の理解がより深まると思った。
18 まとめ
私のA. acetiの代謝制御に関する研究は、トランスクリプトーム解析のデータを軸にしている。しかし一報目
では、タンパク質の翻訳後修飾、転写量とタンパク質濃度の差異等、転写量観察だけでは及ばない部分へのアプ ローチが多かった。今後、既知の転写観察だけでは説明がつかない状況が起きうることを強く意識した。また、 pka(Acs アセチル化酵素遺伝子)はA. aceti、A. pasteurianusには存在しなかった。これは酢酸菌の酢酸代謝 能力の強さの裏付けであると考えられる。また、一報目ではarcAによるacs、TCA サイクルの制御を確認でき た。A. acetiの酢酸代謝に関わる遺伝子aarA、aarC付近にarcAを制御するsixAと推測される遺伝子が存在す
るので、A. acetiにおいてsixAの破壊、過剰発現により大腸菌と同じく有意義な結果が期待できるのでは、と考 えた。 二報目については、シミュレーションという手法は参考にできないが、各代謝フラックスに注目する点は興味 深かった。また、あらゆるアプローチで細胞外酢酸による応答を調べていたが、これは自分の研究においても似 たようなアプローチができるのではないか、と感じた。 参考文献
1. Sakurai, K., Arai, H., Ishii, M., & Igarashi, Y. (2011). Transcriptome response to different carbon sources in Acetobacter aceti. Microbiology, 157(3), 899-910.
2. Sakurai, K., Arai, H., Ishii, M., & Igarashi, Y. (2012). Changes in the gene expression profile of Acetobacter aceti during growth on ethanol. Journal of bioscience and bioengineering, 113(3), 343-348. 3. Paalme, T., Kahru, A., Elken, R., Vanatalu, K., Tiisma, K., & Raivo, V. (1995). The computer-controlled
continuous culture of Escherichia coli with smooth change of dilution rate (A-stat). Journal of microbiological methods, 24(2), 145-153.
4. Castaño‐Cerezo, S., Bernal, V., Blanco‐Catalá, J., Iborra, J. L., & Cánovas, M. (2011). cAMP‐CRP co ‐ordinates the expression of the protein acetylation pathway with central metabolism in Escherichia coli. Molecular microbiology, 82(5), 1110-1128.
5. Zhao, K., Chai, X., & Marmorstein, R. (2004). Structure and substrate binding properties of cobB, a Sir2 homolog protein deacetylase from Escherichia coli. Journal of molecular biology, 337(3), 731-741.
6. Barak, R., Prasad, K., Shainskaya, A., Wolfe, A. J., & Eisenbach, M. (2004). Acetylation of the
chemotaxis response regulator CheY by acetyl-CoA synthetase purified from Escherichia coli. Journal of molecular biology, 342(2), 383-401.
7. Covert, M. W., Knight, E. M., Reed, J. L., Herrgard, M. J., & Palsson, B. O. (2004). Integrating high-throughput and computational data elucidates bacterial networks. Nature, 429(6987), 92. 8. Vemuri, G. N., Altman, E., Sangurdekar, D. P., Khodursky, A. B., & Eiteman, M. A. (2006). Overflow
metabolism in Escherichia coli during steady-state growth: transcriptional regulation and effect of the redox ratio. Applied and environmental microbiology, 72(5), 3653-3661.
9. Shin, S., Chang, D. E., & Pan, J. G. (2009). Acetate consumption activity directly determines the level of acetate accumulation during Escherichia coli W3110 growth. J Microbiol Biotechnol, 19(10), 1127-1134. 10. Wolfe, A. J. (2005). The acetate switch. Microbiology and molecular biology reviews, 69(1), 12-50.