九州大学学術情報リポジトリ
Kyushu University Institutional Repository
セレコキシブはWnt標的遺伝子の発現を抑制すること により骨芽細胞の分化を阻害する
永野, 晃弘
九州大学大学院歯学府
https://doi.org/10.15017/26331
出版情報:Kyushu University, 2012, 博士(歯学), 課程博士 バージョン:
権利関係:
セレコキシブは
Wnt 標的遺伝子の発現を抑制することにより 骨芽細胞の分化を阻害する
2012 年
永 野 晃 弘
九州大学大学院歯学府歯学専攻
指導教員
九州大学大学院歯学研究院 口腔機能修復学講座 歯周病学分野
前田 勝正 教授
研究指導者
九州大学大学院医学研究院 生体情報科学講座 臨床薬理学分野
笹栗 俊之 教授
対 象 論 文
本研究の一部は下記の学術雑誌に投稿中である。
Celecoxib inhibits osteoblast maturation by suppressing the expression of the Wnt target genes
Akihiro Nagano, Fumi Takahashi-Yanaga, Etsuko Matsuzaki,
Kentaro Jingushi, Tatsuya Yoshihara, Sachio Morimoto, Katsumasa Maeda, and Toshiyuki Sasaguri
Bone, November 28, 2012, submitted
略 語 一 覧
ALP:alkaline phosphatase APC:adenomatous polyposis coli COX:
cyclooxygenaseEDTA:ethylenediamine tetraacetic acid
GAPDH:glyceraldehyde-3-phosphate dehydrogenase GSK-3:glycogen synthase kinase 3
LRP:low-density lipoprotein receptor-related protein mRNA:messenger RNA
PGE
2:prostaglandin E2PI
:propidium iodideRT-PCR:reverse transcription-polymerase chain reaction Runx2:runt-related transcription factor 2
SDS:sodium dodecyl sulfate TCF:T-cell factor
TCF7L2:T-cell-specific transcriptional factor 7-like 2 UV:ultraviolet
目 次
要 旨・・・・・・・・・・・・・・・・・・・・・・ 1
緒 言・・・・・・・・・・・・・・・・・・・・・・ 2
材 料 と 方 法・・・・・・・・・・・・・・・・ 4
結 果・・・・・・・・・・・・・・・・・・・・・・ 9
考 察・・・・・・・・・・・・・・・・・・・・・・18
謝 辞・・・・・・・・・・・・・・・・・・・・・・20
引 用 文 献・・・・・・・・・・・・・・・・・・21
1
要 旨
非 ス テ ロイ ド 性抗 炎 症薬 (NSAIDs)は 骨 折の 治 癒 を阻 害 する こ とが 知 ら れ て い る 。我 々 は 、cyclooxygenase-2 選択 的 NSAIDs で ある セ レ コ キ シ ブ が 、 骨 芽 細 胞に お いて そ の 分 化 に 重要 な 役割 を 果た し て いる Wnt/-カテ ニ ン シグ ナ ル 伝 達経 路 を阻 害 する こ と を報 告 して き た 。そ こ で 、本 研究 で はセ レ コ キシ ブ が 骨 芽細 胞 様細 胞 株 MC3T3-E1 の 成熟 に与 え る 影響 を 検討 し た。セ レコ キ シ ブ は cyclinB1 の 発現 を 低 下さ せ るこ と によ り MC3T3-E1 の 細胞 周 期 を G2/M 期 で 停止 し た 。Wnt/-カテ ニ ンシ グナ ル 伝 達経 路 の主 要 な転 写 因 子で あ る T-cell-specific transcription factor 7-like 2 の タ ン パク 質 発現 は セレ コ キ シブ に よ り 時間・濃 度 依存性 に 低 下し た 。セ レコキ シ ブ は骨 芽 細胞 の 代表 的 分 化マ ー カ ー であ りWnt 標的 遺 伝 子 で あ る alkaline phosphatase の タン パ ク 質 発 現 を 、少 な く とも そ の一 部 は プロ モ ータ ー の T-cell factor 結合 部位 を 介 して 、 時 間・濃 度 依 存性 に 抑制 し た 。セ レ コキ シ ブは ま た 、骨 芽 細胞 の 分化 に 必須 で Wnt 標 的遺 伝 子 の一つ で あ る 転 写 因子 runt- related transcription factor 2 も 、そ の プ ロモ ー ター 活 性 を 低下 さ せる こ とで タ ン パク 質 発現 を 抑制 し た 。さ ら に セ レコ キ シブ は 骨芽 細 胞 によ る 石灰 化 を阻 害 し た 。以上 の 結果か ら 、セ レ コ キ シ ブは Wnt 標 的遺 伝 子 の発 現 を抑 え るこ と に より 骨 芽細 胞 の成 熟 を 阻 害 し 、 これ が NSAIDs 投 与に よ る骨 折 の治癒 遅 延 のメ カ ニズ ム のひ と つ であ る 可 能 性が 示 唆さ れ た。
2
緒 言
Wnt/-カテニンシグナル伝達経路は胚形成や成熟した組織の恒常性維持に重要な役 割を担っている(1-3)。 この経路は、分泌タンパク質であるWntが共役受容体である low-density lipoprotein receptor-related protein 5 or 6(LRP5/6)と7回膜貫通型受 容体である Frizzled に結合することで活性化される。 Wnt が受容体に結合すると - カテニンが核内に集積し、 T-cell factor(TCF)/lymphoid enhancer-binding factor と共同して標的遺伝子の発現を調節する(4, 5)。Wntシグナル伝達経路のなかでも、
このように-カテニンが関与する経路をWnt/-カテニンシグナル伝達経路と呼ぶ。こ のシグナル伝達経路は骨芽細胞の分化にも関係していることが報告されている。 例え
ばLRP5 の機能が消失するとヒトやマウスにおいて骨密度が低下することが報告され
(6-8)、 逆に恒常的な活性化型LRP5変異体により骨密度が高くなることが示された
(8-10)。さらにいくつかの Wnt ファミリータンパク質 (Wnt 1, Wnt 2, Wnt 3a) は 骨芽細胞の前駆細胞株(C2C12, C3H10T1/2, ST2)において alkaline phosphatase
(ALP)活性を促進することが示された(6, 11)。また、 Wnt/-カテニンシグナル伝 達経路は骨折の治癒において活性化され、骨の再生において重要な役割を果たしている ことが報告されている(12)。
セレコキシブは選択的 cyclooxygenase-2(COX-2)阻害の非ステロイド性抗炎症
薬(NSAIDs)として開発され、関節炎の治療において消炎・鎮痛薬として用いられ
ている。また、セレコキシブは家族性大腸腺腫症の治療にも用いられ、がんの化学療法 薬としての可能性も研究されている(13, 14)。我々やその他の研究者は、その作用機 序ががん細胞における Wnt/-カテニンシグナル伝達経路の抑制であることを報告して いる(15-17)。一方、選択的COX-2 阻害剤を含む NSAIDs は、生体内において骨形
3
成や骨折の治癒を阻害することが報告されている(18-21)。
そこで、本研究ではマウス由来骨芽細胞様細胞株 MC3T3-E1 を用い、セレコキシブ が骨芽細胞に及ぼす影響について、特にセレコキシブの効果と Wnt/-カテニンシグナ ル伝達経路の関連に着目して検討を行い、セレコキシブが Wnt 標的遺伝子の発現を抑 制することにより骨芽細胞の分化を阻害することを見出した。
4
材 料 と 方 法
1. 試薬および抗体
TOPflash(TCFレポータープラスミド)とFOPflash(TCFレポータープラスミド
のネガティブコントロール)はUpstate Biotechnology社から購入した。抗GAPDH モノクローナル抗体はAbcam社から購入した。抗transcription factor 7-like 2
(TCF7L2)モノクローナル抗体はMILLIPORE社から購入した。抗ALPモノクロー
ナル抗体はAbD Serotec社から購入した。抗runt- related transcription factor 2
(RUNX2)モノクローナル抗体と抗cyclin B1モノクローナル抗体はCell Signaling Technology社から購入した。抗cyclin D1ポリクローナル抗体はSanta Cruz
Biotechnology社から購入した。セレコキシブ(分子式:C17H14F3N3O2S、分子量:381.37、
下図参照)はPfizer社から供与された。
2. 細胞培養
細胞は、37℃、5%CO2 濃度のインキュベーター中で培養した。MC3T3-E1 細胞は
10%ウシ胎児血清、 100 U/ml ペニシリンG、100 g/mlストレプトマイシン存在下の
-Minimum Essential Medium Eagle(Sigma 社)にて培養した。
3. 細胞数の計測
MC3T3-E1 細胞は 2.0×104 個/well で、24 穴プレートに播種し、各濃度(10M, 30M, 50M, 100M)のセレコキシブ存在下または非存在下で標記した時間培養した。
5
細胞はトリプシン/EDTA(Invitrogen 社)処理で回収し、細胞数を TC10(BIO-RAD 社)を用いて計数した。
4. フローサイトメトリー解析
細胞は 50 g/ml propidium iodide (PI)、 0.1% sodium citrate、 0.1% Triton X-100 を含む低張性溶液に懸濁した。 PI によって染色したサンプル(1×105 細胞)は Becton-Dickinson FACSCalibur(Becton-Dickinson 社)を用いて蛍光を測定した。
5. ALP 活性の測定
MC3T3-E1 細胞は2.0×104 個/wellで、24 穴プレートに播種し、セレコキシブ(50
M)を添加した。ALP 活性の測定には、基質としてp-ニトロフェニルリン酸(Wako 社)
を用いた。方法を簡略化して述べると、回収した細胞を、0.1 M エタノールアミン
(pH10.5)、150 mM 塩化ナトリウム(Wako 社)、2 mM 塩化マグネシウム(Wako 社)、2.5 mg/ml p-ニトロフェニルリン酸を含む溶液で溶解し、 37 ℃で 60 分保温 した後、波長 405nm で吸光度を測定した。 ALP 活性測定の結果は、各タンパク質 濃度で補正し、グラフ化した。
6. ウエスタンブロッティング法
ウェスタンブロット法は以前に述べた方法で行った(21)。サンプルは12% SDS polyaclylamidegel electrophoresis によって分離し、その後 polyvinylidene difluoride メンブレンにセミドライ方式(1時間、12V)で転写した。免疫反応性タンパク質は検 出試薬(LumiGLO; Cell Signaling Technology 社)によって視覚化した。濃度測定に はNIH Image Jソフトウエアを用いた。
6 7. 定量的RT-PCR法
トリゾール試薬(Invitrogen 社)を用いてトータル RNA の抽出を行った。cDNA は3 μg のtotal RNAからFirst Strand cDNA Synthesis Kit(Roche Diagnostics K.K.
社)を用いて合成した。定量的PCRは SYBR Green RT-PCR Kit (Takara Bio社)
と Thermal Cycler Dice Real Time System(Takara Bio社)を用いて行った。使用し たPCRのコンディションを以下に示す。 95°C 10 秒; 引き続き 95°C 5 秒, 60°C 30 秒、 72°C 30秒、これを40 サイクル; その後 95°C 15秒, 60°C 30 秒、95°C 15 秒で 終了した. GAPDH を内部コントロールとして使用した。発現量は ΔΔ Ct 値から計算 した。プライマーはGenBank database に基づいて設計した。使用したプライマーを 以下に示す。 ALP (forward: 5′-GTT GCC AAG CTG GGA AGA ACA C-3′; reverse:
5′-CCC ACC CCG CTA TTC CAA AC-3′);GAPDH (forward: 5′-TGT GTC CGT CGT GGA TCT GA-3′; reverse: 5′-TTG CTG TTG AAG TCG CAG GAG-3′)
8. RT-PCR 法
トリゾール試薬(Invitrogen 社)を用いてトータル RNA の抽出を行った。1 g の トータル RNAを用いてRUNX2およびGAPDH mRNA量をRT-PCRによって解析した。
以下のプライマーを用いた。RUNX2 (forward: 5′-TCC TAT CTG AGC CAG ATG ACA TCC-3′; reverse: 5′-CCG GTC TCC CCC GGG TAC C-3′)、GAPDH (forward: 5′-ACC ACA GTC CAT GCC ATC AC-3′; reverse: 5′-TCC ACC ACC CTG TTG CTG TA-3′)
PCR サイクル数はRUNX2; 33サイクル、GAPDH; 23 サイクルである。
9. レポータープラスミドの作成
野生型および変異型 ALP プロモーターは以前作製したものを使用した(22)。簡単に述 べると、ヒトゲノムDNAからPCR法によって、ヒトALP遺伝子の5’上流領域(開始コド
7
ンを基点とした-1059/+249 bpを含む領域)を増幅し、ホタルルシフェラーゼレポーターベ ク ター(pGL3-Basic)に 組込ん だ。ま た、これ を鋳型 として TCF 結 合部位 変異 を
(-1023/-1017;ATCAGAG)に変異を導入した。RUNX2プロモーターはヒトゲノムDNA からPCR法によって、ヒトRUNX2遺伝子の5’上流領域(-1557/+31bpを含む領域)を増 幅し、ホタルルシフェラーゼレポーターベクター(pGL3-Basic) に組込んだ。
10. ルシフェラーゼレポーター活性測定
細胞には、レポーターベクター(TOPflash、FOPflash, 野生型または変異型ALP pGL-3、な らびにRUNX2 pGL-3)と内部コントロールとしてpRL-SV40(Toyo Ink Manufacturing Co.
社)をLipofect Amine Plus reagent(Invitrogen 社)を用いて導入した。ルシフェラーゼ 活性をルミノメーター(Lumat LB 9507, Berthold technologies 社)にて測定した。また、
同時にウミシイタケルシフェラーゼ活性を測定し、補正した。
11. von Kossa 染色
骨芽細胞による石灰化の評価にはvon Kossa染色を用いた。24 穴プレートに播種し た細胞がコンフルエントに達した後、 セレコキシブ(50 M)の存在下または非存在 下で 14 日間培養した。細胞は、10 %パラホルムアルデヒドを用いて室温で 30 分間 固定し、 5 %硝酸銀水溶液を入れて 10 分間 UV 下に曝して銀を黒色化させ、 5 % チオ硫酸ナトリウム水溶液で 2 分間処理した。染色後BIOREVO 顕微鏡 (キーエンス 社)を用いて細胞を撮影し、カルシウム沈着部分の測定を行った。
13. 統計
結果は平均値± S.E.で示した。値間の差は Student’s t-test もしくは one-way ANOVA
8
with Bonferroni post-hoc tests(GraphPad Prism 5.0, GraphPad Software)を用いて統 計的に解析した。P value < 0.05を統計学的に有意であるとした。
9
結 果
1. セレコキシブの MC3T3-E1 に対する増殖抑制作用
セレコキシブは、これまで様々な細胞株において増殖を抑制することが報告されてき た(24, 25)。はじめに、セレコキシブがMC3T3-E1 の増殖に及ぼす効果について検討 した。図 1A に示すように、セレコキシブは細胞増殖を時間・濃度依存性に抑制した。
我々は、ヒト大腸癌細胞株において、細胞周期の G0/G1 期から S 期の移行に必要な タンパク質である cyclinD1 の発現を、セレコキシブが抑制することを報告している
(15)。そこで、セレコキシブが細胞周期の分布に与える影響を分析するためにフロー サイトメトリー解析を行った。大腸癌細胞とは異なり、セレコキシブを添加した細胞で は24時間後に G0/G1 期と S 期の細胞数が減少し、 G2/M 期の細胞数が増加した。
このことからセレコキシブは MC3T3-E1 において G0/G1 期ではなく G2/M 期で細 胞周期を拘束することが示唆された(図 1B)。次に、G2/M 期の移行に必要なタンパク 質である cyclinB1 に与える影響を検討した。図 1C に示したように、セレコキシブ はMC3T3-E1 において cyclinD1 だけでなく cyclinB1 の発現も顕著に抑制していた。
A
0 24 48 72
0 10 20
30 control
10M 30M 50M 100M
*
**
*** ***
***
***
*** *** ***
Time (h) cell number (x104 cells/well)
10
B
C
図1 セレコキシブが MC3T3-E1 に与える増殖抑制作用
(A)増殖曲線。 MC3T3-E1細胞は24 穴プレートに播種し、種々の濃度のセレコキ シブを添加した。横軸に示した時間にtrypsin/EDTA処理によって回収して、細胞数を測 定した。(B)フローサイトメトリー。MC3T3-E1細胞をセレコキシブ(50 M)と24時 間培養後、trypsin/EDTA処理によって回収した。細胞をPIで染色後、核内蛍光を測定し た。様々な細胞周期にある細胞をパーセントで表示している。(C)cyclinへの影響。
MC3T3-E1細胞をセレコキシブ(50 M)の有無で24 時間培養し、ウエスタンブロッ
ト法にて cyclinD1, cyclinB1 のタンパク質発現を調べた。値は0時間に対するパーセン テージを示し、3回の実験結果の平均値± S.E.で表示した。*P<0.05; **P<0.01; ***P<0.001 vs. control.
PI Fluorescence control
G0/G1 (%) 71.1 3.3 S (%) 11.8 0.8
G2/M (%) 19.6 1.8
cell number
PI Fluorescence celecoxib (50M)
G0/G1 (%) 47.2 6.7 G2/M (%) 40.4 7.2 S (%) 6.2 1.2
cell number
11
2. セレコキシブは Wnt/-カテニンシグナル伝達経路を阻害する
以前、ヒト大腸癌細胞株においてセレコキシブが Wnt/-カテニンシグナル伝達経 路の主要な転写因子であるTCF7L2 の発現を抑制することを見出したので、今回セレ コキシブがMC3T3-E1 細胞において TCF7L2 に及ぼす影響について検討した。図 2Aおよび2B に示したようにセレコキシブは TCF7L2 の発現を時間・濃度依存性に 顕著に抑制した。次に、セレコキシブが Wnt/-カテニンシグナル伝達経路に与える影 響を評価するため、Wnt/-カテニンシグナル伝達経路に特異的な転写因子であるTCF 依存性の転写活性に及ぼす影響を検討した。TCF の結合部位を有した TOPflash と TCF 結合部位に変異が導入された FOPflsh を用い、セレコキシブの効果を検討した ところ、図 2C に示したように TOPflash の活性はコントロールの細胞では 24 時間 後増加したが、セレコキシブを添加した細胞では活性の増加は認められなかった。また、
FOPflash に関しては影響を及ぼさなかった。これらの結果から、セレコキシブは
TCF7L2 のタンパク質の発現を低下させることで、Wnt/-カテニンシグナル伝達経路
を抑制することが示唆された。
A
0 50 100 150 200 250
Time(h) 0 24 48 72
**
**
control
celecoxib(50M)
*
TCF7L2/GAPDH (% of control)
12
B
C
図 2 セレコキシブによるWnt/-カテニンシグナル伝達経路の阻害
セレコキシブが TCF7L2 の発現に与える影響の経時的変化(A)と濃度依存性(B)。(A)
MC3T3-E1 細胞を表示された時間セレコキシブ(50 M)と培養した。(B)MC3T3-E1 細胞を様々な濃度のセレコキシブと72時間培養した。回収したサンプルは抗TCF7L2抗体
と抗 GAPDH 抗体を用いてウエスタンブロット法にて解析し、バンドを定量化した。(C)
TCF依存性転写活性への影響。TOPFlash、FOPFlashを MC3T3-E1 細胞にトランス フェクションし、コントロールの細胞とセレコキシブ (50 M) を加えた細胞を 24 時 間培養した。値は 0時間に対するパーセンテージを示し、3 回の実験結果の平均値± S.E.
で表示した。*P<0.05; **P<0.01vs. control
0 50 100 150 200 250
celecoxib (M) Time (h)
- - 10 30 50
0 72
*
control celecoxib
*
TCF7L2/GAPDH (% of control)
13
3. セレコキシブの TCF/LEF 結合部位を介した ALP プロモーター活性の抑制作用 これまでに、Wnt/-カテニンシグナル伝達経路はALP の発現を促進することが報告 されているため(11, 22)、 セレコキシブの ALP に及ぼす影響についても検討した。
図 3A に示したように、セレコキシブは ALP 活性を時間依存性に顕著に抑制した。
図 3B 、3C に示したように、セレコキシブはALP のタンパク質発現を時間・濃度依 存性に顕著に抑制した。また、セレコキシブは ALP の mRNA の発現も抑制した (図
3D)。 次に、ALP レポータープラスミドを用いてセレコキシブが ALP プロモーター
活性に与える影響を検討した。図 3E に示したようにコントロールの細胞においては 24 時間後 ALP プロモーター活性は増加したが、セレコキシブ (50 M)を添加した 細胞では顕著に低下した。この実験に使用したALP の遺伝子プロモーター
(-1059/+249 bp)では TCF 結合部位(-1023/-1017 bp)が 1 箇所のみ存在した。
そこで、セレコキシブが TCF結合部位に与える影響の解析をするため、この部位に変 異を導入したレポータープラスミドを導入したところ、セレコキシブはルシフェラーゼ 活性に顕著な影響を及ぼさなかった。これらの結果から、セレコキシブによるALP の タンパク質量の低下とmRNAおよびプロモーター活性の低下の程度に差はあるものの、
セレコキシブによるTCF 結合部位を介したプロモーター活性の抑制が、ALP タンパ ク質発現低下の原因の一つであることが示唆された。
A
0 24 48 72
0 50 100 150 200 250
control
celecoxib (50 M)
* ***
Time (h)
ALP activity (units/L)
14
B C
D E
図 3 セレコキシブが ALP に及ぼす影響
(A)ALP 活性。MC3T3-E1細胞をセレコキシブ(50 M)存在下、非存在下で24 時 間培養し、 ALP 活性を測定した。(B)タンパク質発現への 経時的影響。MC3T3-E1 細胞にセレコキシブ(50 M)を添加し、表示した時間培養した。(C)タンパク質発現(濃 度依存性)。MC3T3-E1細胞に図に示した各濃度のセレコキシブを添加し72時間培養した。
回収したサンプルは抗ALP抗体と抗GAPDH抗体を用いてウエスタンブロットにて解析し、
バンドを定量化した。(D)mRNA発現。MC3T3-E1細胞をセレコキシブ(50 M)存在 下、非存在下で24 時間培養し、 RNAを抽出した。ALPmRNA発現は定量的PCRで 評価した。(E)プロモーターアッセイ。MC3T3-E1 細胞に野生型または変異型ALP プ ロモーターを含むコンストラクトとpRL-SV40を導入した。24時間培養後、セレコキシブ
(50 M)を添加し、24時間培養した。値は0時間に対するパーセンテージを示し、3回 の実験結果の平均値± S.E.で表示した。*P<0.05; ***P<0.001vs. control.
0 100 200 300
* ***
Time(h) 0 6 12 24
control
12
celecoxib
ALP/GAPDH (% of control)
0 100 200 300
celecoxib (M) Time (h)
- - 10 30 50
0 24
*** *** ***
ALP/GAPDH (% of control)
15 4. セレコキシブの RUNX2 の発現抑制作用
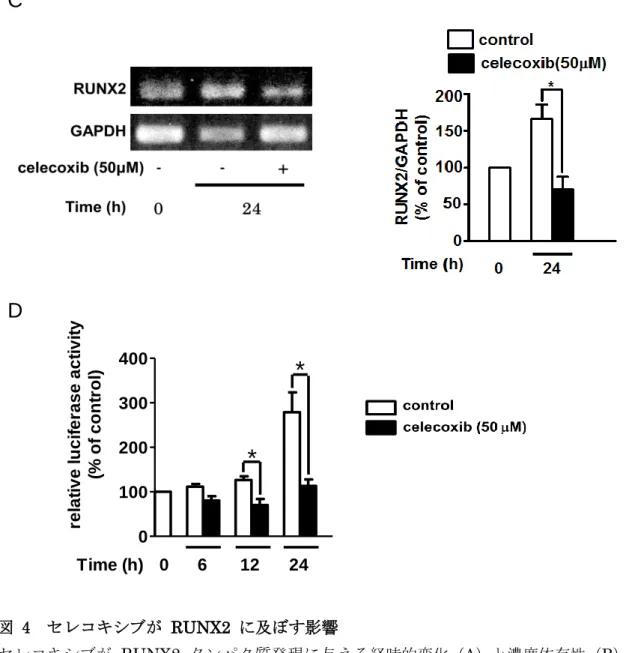
次に、セレコキシブが Wnt 標的遺伝子の 1 つで、骨芽細胞の分化に必須の転写因 子である RUNX2 (26-28)に及ぼす影響について検討した。 図 4A と4B に示した ように、セレコキシブは RUNX2 の発現を時間・濃度依存性に顕著に抑制し、図 4C に 示したようにRUNX2 の mRNA の発現も抑制した。さらに、セレコキシブは RUNX2 のプロモーター活性を著しく抑制した。これらの結果から、セレコキシブはプロモータ ー活性を低下させることで、 RUNX2 の発現を低下させることが示唆された。
A
B
0 50 100 150
celecoxib(M) - - 10 30 50 Time(h) 0 24
***
***
RUNX2/GAPDH (% of control)
16
C
D
図 4 セレコキシブが RUNX2 に及ぼす影響
セレコキシブが RUNX2 タンパク質発現に与える経時的変化(A)と濃度依存性(B)。
(A)MC3T3-E1細胞を表示された時間セレコキシブ(50 M)と培養した。(B)MC3T3-E1 細胞に、図に示す各濃度のセレコキシブを添加し、72時間培養した。回収したサンプルは
抗RUNX2抗体と抗GAPDH抗体を用いてウエスタンブロットにて解析し、バンドを定量
化した。(C)mRNA発現に与える影響。MC3T3-E1細胞をセレコキシブ(50 M)存 在下、非存在下で24 時間培養し、 RNAを抽出した。RUNX2mRNA発現はRT-PCR 法で評価した。(D)プロモーターアッセイ。MC3T3-E1細胞にRUNX2プロモーターを 含むコンストラクトとpRL-SV40を導入した。24時間の培養後、セレコキシブ(50 M)
を添加し、24時間培養した。値は0時間に対するパーセンテージを示し、3回の実験結果 の平均値± S.E.で表示した。*P<0.05; **P<0.01 ; ***P<0.001vs. control.
0 100 200 300
400
*
*
Time (h) relative luciferase activity (% of control)
0 6 12 24
17 5. セレコキシブの石灰化抑制作用
次に、von Kossa 染色を用いて、セレコキシブが骨芽細胞の石灰化に及ぼす影響を 検討した。図 5 に示したように培養して 14 日後コントロール群では多くのカルシウ ムの沈着を認めたが、セレコキシブを添加した群では石灰化は顕著に抑制された。この 結果からセレコキシブは骨芽細胞の石灰化を抑制することが示唆された。
図 5 セレコキシブの骨芽細胞の石灰化への影響
von Kossa 染色。24 穴プレートに播種した細胞がコンフルエントに達した後、セレコ
キシブ(50 M)存在下、非存在下で14 日間培養した。石灰化沈着物をvon Kossa染 色で可視化した。3回の実験結果の平均値± S.E.で表示した。***P<0.001vs. control.
18
考 察
本研究において、セレコキシブが TCF7L2 の発現を抑制することにより、 Wnt/- カテニンシグナル伝達経路を抑制すること、また、ALP 並びにRUNX2 のプロモータ ー活性の抑制がこれらのタンパク質発現を低下させる一因であることを見出した。
Wnt/-カテニンシグナル伝達経路はこれまで ALP 並びに RUNX2 の発現に関与して
いることが示されてきた (11, 22, 26)。 RUNX2 に関しては セレコキシブ によるタ ンパク質の発現抑制作用とmRNA発現抑制およびプロモーター活性の抑制効果が相関 しているため、セレコキシブによる Wnt/-カテニンシグナル伝達経路の阻害が
RUNX2 タンパク質発現抑制に大きく関与していることが示唆される。ところが ALP
の場合は、セレコキシブを添加して 24 時間後に抑制した割合は、タンパク質発現では
80.1 %であったが、プロモーター活性は 26.5 % と小さかった。残念ながら現時点では、
この抑制効果の違いを説明することはできないため、さらなる研究により明らかにする 必要がある。
COX-2 はアラキドン酸を、細胞の増殖や、血管新生、アポトーシスの誘導を抑制す
るプロスタグランジンに変化させる酵素である。従って、 NSAIDs のWnt/-カテニ ンシグナル伝達経路の阻害作用は、 COX-2 の活性を抑える作用によるものと考えられ てきた。 Castellone らは PGE2 が受容体に結合することで、-カテニンをリン酸化 する複合体の 1 つである Axin に結合する細胞質内の G タンパク質共役受容体を活 性化し、その結果分解を免れた-カテニンが核内に集積すると報告した(29) 。その ため、 NSAIDs による プロスタグランジンE2(PGE2)の合成の抑制が Wnt/-カテ ニンシグナル伝達経路の抑制を誘導したと報告された (30) 。 しかし、COX-2 を 発現していない細胞や、 COX-2 を欠損させた腫瘍においてもセレコキシブは抗癌作用
19
を示すことが報告された (24, 31)。 実際に、我々は以前 COX-2 を発現していない ヒト大腸癌細胞株 HCT-116 においてセレコキシブが TCF7L2 の分解を誘導するこ とを報告した(16)。 一方、 MC3T3-E1 細胞 は COX-2 遺伝子を発現し、炎症性刺 激がなくても PGE2 を生成する(32)。 従って PGE2 の減少は MC3T3-E1 細胞内 部において セレコキシブが Wnt/-カテニンシグナル伝達経路を抑制することに関係 するかもしれない。
本研究では、セレコキシブがWnt/-カテニンシグナル伝達経路を抑制することで、
骨芽細胞の成熟を抑制することを見出した。NSAIDs が生体内において骨形成や骨折 の治癒を阻害することが報告(18-21)と考え合わせると、セレコキシブの使用により、
生体内で骨芽細胞の成熟を阻害し、骨形成や骨折の治癒を妨げる可能性がある。
20
謝 辞
稿を終えるにあたり、懇篤なるご指導ご校閲をいただいた、歯学研究院歯周病学分野 前田 勝正 教授に深い感謝の意を表します。
また、実験に際しまして様々なご助言、ご援助いただきました、医学研究院臨床薬理学 分野 笹栗 俊之 教授、医学研究院医学教育学部門 高橋 富美 講師、歯学研究院歯 周病学分野 松﨑 英津子 助教に深く感謝いたします。
さらに、研究生活のご指導、ご助言いただき、あたたかく見守ってくださいました、
歯周病学分野の皆様、臨床薬理学分野の皆様、および研究生活を支えて下さったすべて の皆様に心から感謝いたします。
21
引 用 文 献
1. Clevers H. Wnt/-catenin signaling in development and disease. Cell 127 (2006) 469-480.
2. Nelson WJ, Nusse R. Convergence of Wnt, -catenin, and cadherin pathways.
Science 303 (2004) 1483-1487.
3. Moon RT, Bowerman B, Boutros M, Perrimon N. The promise and perils of Wnt signaling through -catenin. Science 296 (2002) 1644-1646.
4. Miller JR, Hocking AM, Brown JD, Moon RT. Mechanism and function of signal transduction by the Wnt/-catenin and Wnt/Ca2+pathways. Oncogene 18 (1999) 7860-7872.
5. Behrens J, von Kries JP, Kühl M, Bruhn L, Wedlich D, Grosschedl R, Birchmeier W. Functional interaction of -catenin with the transcription factor LEF-1. Nature 382 (1996) 638-642.
6. Gong Y, Slee RB, Fukai N, Rawadi G, Roman-Roman S, Reginato AM, Wang H, Cundy T, Glorieux FH, Lev D, Zacharin M, Oexle K, Marcelino J, Suwairi W, Heeger S, Sabatakos G, Apte S, Adkins WN, Allgrove J, Arslan-Kirchner M, Batch JA, Beighton P, Black GC, Boles RG, Boon LM, Borrone C, Brunner HG, Carle GF, Dallapiccola B, Paepe AD, Floege B, Halfhide ML, Hall B, Hennekam RC, Hirose T, Jans A, Juppner H, Kim CA, Keppler-Noreuil K, Kohlschuetter A, LaCombe D, Lambert M, Lemyre E, Letteboer T, Peltonen L, Ramesar RS, Romanengo M, Somer H, Steichen-Gersdorf E, Steinmann B, Sullivan B, Superti-Furga A, Swoboda W, van den Boogaard MJ, Van Hul W, Vikkula M, Votruba M, Zabel B, Garcia T, Baron R, Olsen BR, Warman ML.
LDL receptor-related protein 5 (LRP5) affects bone accrual and eye
22 development. Cell 107 (2001) 513-523.
7. Kato M, Patel MS, Levasseur R, Lobov I, Chang BH, Glass II DA, Hartmann C, Li L, Hwang TH, Brayton CF, Lang RA, Karsenty G, Chan L.
Cbfa1-independent decrease in osteoblast proliferation, osteopenia, and persistent embryonic eye vascularization in mice deficient in Lrp5, a Wnt coreceptor. J. Cell. Biol. 157 (2002) 303-314.
8. Cui Y, Niziolek PJ, MacDonald BT, Zylstra CR, Alenina N, Robinson DR, Zhong Z, Matthes S, Jacobsen CM, Conlon RA, Brommage R, Liu Q, Mseeh F, Powell DR, Yang QM, Zambrowicz B, Gerrits H, Gossen JA, He X, Bader M, Williams BO, Warman ML, Robling AG. Lrp5 functions in bone to regulate bone mass. Nat Med 17 (2011) 684-691.
9. Boyden LM, Mao J, Belsky J, Mitzner L, Farhi A, Mitnick MA, Wu D, Insogna K, Lifton RP. High bone density due to a mutation in LDL-receptor-related protein 5. N. Engl. J. Med. 346 (2002) 1513-1521.
10. Little RD, Carulli JP, Del Mastro RG, Dupuis J, Osborne M, Folz C, Manning SP, Swain PM, Zhao S, Eustace B, Lappe MM, Spitzer L, Zweier S,
Braunschweiger K, Benchekroun Y, Hu X, Adair R, Chee L, FitzGerald MG, Tulig C, Caruso A, Tzellas N, Bawa A, Franklin B, McGuire S, Nogues X, Gong G, Allen KM, Anisowicz A, Morales AJ, Lomedico PT, Recker SM, Van Eerdewegh P, Recker RR, Johnson ML. A mutation in the LDL
receptor-related protein 5 gene results in the autosomal dominant high-bone-mass trait. Am. J. Hum. Genet. 70 (2002) 11-19.
11. Rawadi G, Vayssiere B, Dunn F, Baron R, Roman-Roman S. BMP-2 controls alkaline phosphatase expression and osteoblast mineralization by a Wnt
23
autocrine loop. J. Bone Miner. Res. 18 (2003) 1842-1853.
12. Macsai CE, Foster BK, Xian CJ. Roles of Wnt signalling in bone growth, remodelling, skeletal disorders and fracture repair. J. Cell. Physiol. 215 (2008) 578-587
13. Dannenberg AJ and Subbaramaiah K. Targeting cyclooxygenase-2 in human neoplasia: rationale and promise. Cancer Cell 4 (2003) 431-436.
14. Steinbach G, Lynch PM, Phillips RK, Wallace MH, Hawk E, Gordon GB, Wakabayashi N, Saunders B, Shen Y, Fujimura T, Su LK and Levin B. The effect of celecoxib, a cyclooxygenase-2 inhibitor, in familial adenomatous polyposis. N. Eng.l J. Med. 342 (2000) 1946-1952.
15. Sakoguchi-Okada N, Takahashi-Yanaga F, Fukada K, Shiraishi F, Taba Y, Miwa Y, Morimoto S, Iida M, Sasaguri T. Celecoxib inhibits the expression of survivin via the suppression of promoter activity in human colon cancer cells.
Biochem. Pharmacol. 73 (2007) 1318-1329.
16. Takahashi-Yanaga F, Yoshihara T, Jingushi K, Miwa Y, Morimoto S, Hirata M, Sasaguri T. Celecoxib-induced degradation of T-cell factors-1 and -4 in human colon cancer cells. Biochem. Biophys. Res. Commun. 377 (2008) 1185-1190.
17. Tuynman JB, Vermeulen L, Boon EM, Kemper K, Zwinderman AH, Peppelenbosch MP, Richel DJ. Cyclooxygenase-2 inhibition inhibits c-Met kinase activity and Wnt activity in colon cancer. Cancer Res. 68 (2008) 1213-1220
18. Gerstenfeld LC, Al-Ghawas M, Alkhiary YM, Cullinane DM, Krall EA, Fitch JL, Webb EG, Thiede MA, Einhorn TA. Selective and nonselective
24
cyclooxygenase-2 inhibitors and experimental fracture-healing. Reversibility of effects after short-term treatment. J. Bone Joint Surg. Am. 89 (2007) 114-125
19. Goodman S, Ma T, Trindade M, Ikenoue T, Matsuura I, Wong N, Fox N, Genovese M, Regula D, Smith RL. COX-2 selective NSAID decreases bone ingrowth in vivo. J. Orthop. Res. 20 (2002) 1164-1169
20. Harder AT, An YH. The mechanisms of the inhibitory effects of nonsteroidal anti-inflammatory drugs on bone healing: a concise review. J. Clin.
Pharmaco.l 43 (2003) 807-815
21. Vuolteenaho K, Moilanen E. Non-steroidal anti-inflammatory drugs, cyclooxygenase-2 and the bone healing process. Basic. Clin. Pharmacol.
Toxicol. 102 (2008) 10-14
22. Matsuzaki E, Takahashi-Yanaga F, Miwa Y, Hirata M, Watanabe Y, Sato N, Morimoto S, Hirofuji T, Maeda K, Sasaguri T. Differentiation-inducing factor-1 alters canonical Wnt signaling and suppresses alkaline phosphatase expression in osteoblast-like cell lines. J. Bone Miner. Res. 21 (2006) 1307-1316.
23. Henriquez B, Hepp M, Merino P, Sepulveda H, van Wijnen AJ, Lian JB, Stein GS, Stein JL, Montecino M. C/EBPβ binds the P1 promoter of the Runx2 gene and up-regulates Runx2 transcription in osteoblastic cells. J Cell. Physiol.
226 (2011) 3043-3052
24. Grösch S, Tegeder I, Niederberger E, Bräutigam L, Geisslinger G. COX-2 independent induction of cell cycle arrest and apoptosis in colon cancer cells by the selective COX-2 inhibitor celecoxib. FASEB J. 15 (2001) 2742-2744
25
25. Kazanov D, Dvory-Sobol H, Pick M, Liberman E, Strier L, Choen-Noyman E, Deutsch V, Kunik T, Arber N. Celecoxib but not rofecoxib inhibits the growth of transformed cells in vitro. Clin. Cancer Res. 10 (2004) 267-271
26. Gaur T, Lenger CJ, Hovhannisyan H, Bhat RA, Bodine PVN, Komm BS, Javed A, van Wijnen AJ, Stein JL, Stein GS, Lian JB. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 280 (2005) 33132-33140
27. Rosen CJ. Bone remodeling, energy metabolism, and the molecular clock. Cell. Metab. 7 (2008) 7-10
28. Deschaseaux F, Sensébé L, Heymann D. Mechanisms of bone repair and regeneration.
Trends Mol. Med. 15 (2009) 417-429
29. Casrellone MD, Teramoto H, Williams BO, Druey KM, Gutkind JS. Prostaglandin E2 promotes colon cancer cell growth through a Gs-axin--catenin signaling axis. Science 310 (2005) 1504-1510
30. Clevers H. Colon cancer--understanding how NSAIDs work. N. Engl. J. Med. 354 (2006) 761-763
Maier TJ, Schilling K, Schmidt R, Geisslinger G, Grösch S. Cyclooxygenase-2 (COX-2)-dependent and -independent anticarcinogenic effects of celecoxib in human colon carcinoma cells. Biochem. Pharmacol. 67 (2004) 1469-1478.
32. Naot D, Chhana A, Matthews BG, Callon KE, Tong PC, Lin JM, Costa JL, Watson M, Grey AB, Cornish J. Molecular mechanisms involved in the mitogenic effect of lactoferrin in osteoblasts. Bone 49 (2011) 217-224