The Japanese Society for Plant Systematics
NII-Electronic Library Service
The JapaneseSociety forPlant Systematics
ISSNOOOI-6799 Acta Phytotax. Geobot 44 (2):113-122 (1993)
Cytotaxonomic Aspects of Annonaceae,
with Special Reference to Differences
in Proportion of Polyploids between Asia and America
HIROSHI OKADA
Department ofBiology,
Collegeof
GeneralEducation,Osaka
Uhiversity,
Tbyenaka, Cisaka560Abstract The diversificationofbasic chromosome numbers, i.e.,7,8and 9,within theAnnonaeeae ispresumed tohaveoccurred atthevery earlystage of evolution of thefamily,
i,
e.,atleast before
thcseparation of the continents of Asia,Africaand Arnerica,The factthatxT=8
is
themostfrequent
number supports theassumption thatx=8 istheoriginal basicchromosome number
for
thisfamily.Judging from differences in
the proportion ofbasic
chromosome numbers between Asia andAmerica, diversMcation
of the AsianAnnonaceae is
considered tohave
proceeded mainiy at thex=:9 level,while these in Americafrequently diversified
at the xr=7 level.Further,conspicuousdifferencesintheproportionof polyploidsbetweenAsia and America were observed.
(Received
September 20,1993; Accepted October
18,1993)Key words: Annonaceae, Asia,America, basicchromoseme number, distribution,polyploidy,
,troplcs
The
species ofAnnonaceae
areinteresting
subjectsfor biosystematic
studies, especially when oneintends
to clarify the evolutionarytrends in
members of tropicalforests. The Annonaceae has diverged into
about130 genera
and morethan 24oo
species(Cronquist, 1980)
majnlyin
tropical and sub-tropical
forests. Their distribution is pan-tropical. In
contrast withthe fact
that
manyPolycarpicae
are monotypic or relict,the Annonaceae
are extremelydiyerse.
Almostal1 genera
ofthe Annonaceae
are endemicto
a particularregion;Asia, Africa
orAmerica. Only
afew
genera occurin
two or three regions.In
other words, aimostal1 genera
and specieshave
diversified
within a single region.What kinds
ofkaryological
eventshave
occurredin
theevolutionary
process? hre
thereanykaryological
differencesbetween
thethreeregions?To
answer thesequestions, karyological
surveys were carried outin the Asian
tropics(Okada
and Ueda,1984)
and compared with published reports of similar studiesin
theAmerica.Little karyologicai
information is
availablefor Africa, this
article willfocus
mainly ondifferences between Asia
and America.Materials
alldMethods
wereFresh
materials used
in
thisstudy arelisted in Table 1. Young leaves
collectedin
thefield
pretreatedwith O.03-O.05%
colchicine aqueous solution at17-200C. Root
tips were114 Acta Phytotax. Geobot. vol. "
obtained
from
seedlings grown atOsaka
university.Material gathered in the field
was cooledby dissolution
of ammonium nitrate(NH4N03). Materials
werefixed in
modifiedCarnoy's
solution(absolute EtOH:ch]oroform:glacial
acetic acid=2:1:1)for
more than1 hour.
Youngleayes
were stained with amixture of2%
aceto-orceinin 111O
volume1N HCI for
about20
minutes,heated
to600C for
several seconds and squashed.Materials
not stainingby
thismethod were treatedusingFuelgen's
reaction, then stained with 2% aceto-orceinfor
about1 hour,
and squashed.Root tips
were transferredto
45%
acetic acid at5eC for
about20
minutes, macerated withIN HCI
at600C
for
about20
seconds, then stained with2%
aceto-orceinfor
about15
minutes and squashed.Voucher
specimens arekept in Osaka University, KYO, TI, BO, BRI, LAE (Table 1).
Results
andDiscussion
Chromosomes
were countedfor 19 genera
and24
species(Fig. 1, Table 1). Almost
all arenewly reported
here. Chromosome
counts of sevengenera; Cleistopetaium. Disepalum,
Enicosanthum. Mezzettiopsis, Oncodostigma, Polyaulex
andRyramidunthe,
are reportedfor
thefirst
time.Information
on thebasic
chromosome numberis
now availablefor 35
of51 genera
from Asia
and16
of36 genera from America (Tables 2, 3) (Ehrendorfer
etal.,1968; Walker, 1972;
Bawa, 1973; Okada
andUeda, 1984; Morawetz,
1984a, b, c, 1986a,b, 1988; Sauer
andEhrendorfer, 1984; Morawetz
andWaha, 1985; Morawetz
andLe Thomas, 1988; Okada, 1990;
Datta
andDe, 1990). These
observationsprovide
an opportunity todiscuss
evolutionary trendsin
theAnnonaceae based
on chromosome numbers.The
majority of chromosome counts sofar indicate
abasic
chromosome number of8 (29
of61 genera).
Thispattern is
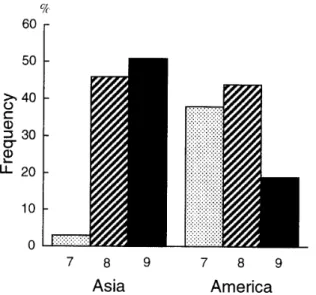
constant throughout the threeregions(Fig. 2). Genera
with x=7 arerather
few
exceptin the Americas,
supporting theconclusionsdrawn by Okada
andUeda (1984)
and
Morawetz (1986)
thatthe originalbasic
chromosome numberfor
theAnnonaceae is 8.
Although
nearlyal1
genera are endemic to one ofthe three
main regions(cf. Fries, 1959;
Walker, 1972),
somegenera
aredistributed
widely throughout two orthree
regions.These genera have basic
chromosome number, x=7in Annona, 8 in Antzxagorea, Artabott:ys, Uvaria
andJb,lopia,
and9 in
Polyalthia.This
suggests thatdiversification in basic
chromosome number,from 8 to 9
andfrom 8
to7,
occurred at an early stage ofdiversification in
thefamily,
atleast before the three
regions were separatedfrom
each other, after whichmigration
to the otherregions
became impossible. It is dithcult
toconsider thatparallelism in
chromosome evolution orin
morphological changes occurredindependently in
manygenera. The fact that
thechromosome nurnber of the majority of pan-tropicalgenerais
x=8
also supportsthe
conciusions ofOkada
andUeda (1984)
andMorawetz (1986).
It
has become
clear thatevolutionary trendsin
chromosome numbers,i.
e.,aneuploidy and polyploidy,have played
animportant
rolein
thediyersification
of thefamily. Aiieuploid
changes,
8
to9
and8
to7,
appear tohave influenced
genericdiversification in
thefamily. Like
morphological criteria
(for
example;Walker, 1971;
Le Thomas,1981; Klucking, 1986; Metcalfe,
1987; Heusden,
1992),
these changes canalso be
used as reliable criteria to understandphylogenetic
relationshipsbetween genera. Judging from
the occurrence ofdistinct basic
The Japanese Society for Plant Systematics
NII-Electronic Library Service
The JapaneseSociety for Plant Systematics
December 1993 OKADA:Cytotaxonomic Aspects
of Annonaceae115 Table 1.
genusChrornosome
counts of Annonaceae
from
Malesia, Indicatesnew recordfor
**, A: Australia,I:Indonesia,M: Malaysia,
P: Papua New Guinea,
T:Taiwan.species' andspecles 2n source voucher
Alphonsea
orthQpetala Okada'Ancana
stenopetala Muell.Artabotrys
cf,scortechinii King*Cleistopetalumsumatranum
Okada"*
Clyathoealyx
sp, 1*CY.
sp.2*Disepatum plaijLpetalumMerr."
E7iicosanthumcf.
congnetatum
(King)
Nry-Shaw'*
Goniothalamus
amayon(Blanco)
Merr.*
Go.
sp.'Hbplostichanthtts
novoguineensis Okada*
Mlezzettiopsiscreaghii Ridl,@'"
Mitrephora gtabra
Scheff.*OncodostigmamicrcV7brum
Okada"*
On, monospetma
(Hk. f. & Th.)
Sincl.'*
Orophea
sp.'Rhaeanthus
macropedus Diels' Ph, cf.crcassipetalusBecc."
Polyaithiasp."
Polyaulaxcf.cylinch'ecatpa**
Rseuduvaria villosaJessup*
Rseuduvaria sp.*
Ib,ramidantheprismatica
(Hk.
f.&
Th.)Sincl."
Uvariasp.*
181816
16l61616 P;Madang
A; New
South
WalesI; W. Sumatra
I;W. Sumatra
I;S.Kalimantan I;W.Surnatra
I;W.Sumatra
18 I; E.
Kalimantan 1616181818
18
1818181818181818
1616 TP;
Morobe P; Morobe
I;
Bogor (cult.)
I;E.
Kalimantan
I;W. SumatraM;
Tarnan NegaraM; Tarnan
NegaraP; Morobe
I;W,
Sumatra
P;'Morobe P; MorobeA; N.
Queensland
P;Merobe
I;E.
Kalimantan
I;E.KalimantanO,
Ke, Ka. 4362(KYO,
TI,LAE, BRI)O,Im, U. 733
(TI,
BRI)H, O.198
(KYO,
BO)O.4607
(KYO,
BO)M & al. 26117
(TI, BO)
H, O. 190
(KYO,
BO)H, O,
It.654(KYO,
BO)K
&
al,20704(TI,
BO)C. 835
(Osaka
U.)O, Ka. 4318
(KYO,
LAE,BRI)
O, Ka. 4202
(KYO,
TI,LAE, BRI)O. 3541
(KYO, BO)
K & al. 20732
(TI, BO)
H, O, It. 263 (KYO,
BO) O&
al,82 (TI,
BO)O & al. 907
(TI,
BO)W & al. 1
(KYO,
TI,LAE,
BRI)H, O.6S
(KYO,
BO)O, Ka.4314
(KYO,
LAE)O, Ka.4296
(KYO,
TI,LAE, BRI)J.733
(BRI)
O,
K,4340(KYO, TI, LAE)
K & al,20643
(TI,
BO)K &
al.20731 (TI,
BO)@:Misidentified as Rseuduvaria reticulata
(Bl.)
Miq. byOkada and Ueda<1984).
chromosome numbers
in
each of three regions, aneuploid changes mayhave
occurredindependently in
each region,but
evolutionary trendsin
AsianAnnonaeeae proceeded difflerently
from thosein
theAmericas (Table
2,3, Fig. 2). The American Annonaceae
contain ahigh proportion
of generawith
x=7, whileAsian Annona
¢eaehave
manygenera
with x=9, and only onegenus
with x=7.In
other words,diversification
ofgenerahas
mainly proceeded atthe
x==7level in America, but
atthe
x=9level in Asia. These
events mayhave
occurred afterthe
continents separated.
It
wouldbe interesting
toknow
whether or not othergroups
exhibitVol. 個
Acta
PhVtotax.
Geobot , 116
牌 臼
慣
鷹 瀬
譲
軌 冊
惣U
無 怨 甲
韃臨
醗
“
疑醗
解
制
tt
齪 内
窮鞭
鴫
離ぎ難
訟
轄 腫
舞
匸
痒蕨
「
獣
霞
選
篷
齪 鴃
照ぼ
籌
じ
艇
鰻 … 虞 岸 勲
撫 輳 演
朧 購 轄 卩 臼 硫 鯛
煕 減
饒 摩
輩、 宅 宅 へ 劉 , 駐
盤
轍
嬉〜
酣
蠱
臣
縛
眞
維 凄
お
瀚 ぼ 慮
菖
ヒ
麟 瓣
窪 難
凝
F
芒
臨 誤 P 獄
灘
乱 F距
舞
彎
蝿
照
離
・
ー庫 晦 p
影
駕 罵 距 囁 む 野
ま
ま
ヨ
熟駆
篷 曩
跚 照 广
隔
艱
湾
に
躍
盤 竇 蓐 儲
畦
F
…
惣韈
. 囁 广
繍 飜
鑼
籌
騰 攤
攤
攤 鱈
韃
濕
お
黴
韆
爨 黙
舜
鰍
難
膨
緊
ー
广
「
巽
雛
戀
鯲
鑵
篝 ・
簿
蠡
興鍵
鰻 .
舗
、
肩
誓
藻嘸
、 撫 、
醜 瀬
讐 藍
ド
ゆ
脚
蘿
藩
・
懿 麟
靉
・
欝
灘 糶
灘 孅
購
劣
懿
照
・
. .
嚢
籠
燕
1
,
Photomicrographs of chromosomes of some Annonaceae,
A:Gon ’othalamus amayon(
2n=
16) ,
B: Pseuduvaria v ’〃osa(
2n=18 ) ,
C
:1llphonsea orthopetala(
2n=
18)
, D: Polyau
’
のc cf.
cytindrocarpa( 2
・− 18 ) ,
E・ Dゆ
・鰍卿 typet
・’
嫦( 2n− 16 ) ,
F・ C勉・ガ・脚
・tum
・um ・贓 砌(
2n=
16) ・
G・ 0”−
codostigma monosperma
(
2n; 18) ,
H :Polya” hia
sp, (
2n=
18) 」
ゴ)yramidanthe prismat’ca(
2n=
16) 」
:Phaea 2n
= 8 . Bar 5 .
Fig
The Japanese Society for Plant Systematics
NII-Electronic Library Service
The JapaneseSociety for Plant Systematics
December 1993 OKADA: Cytotaxonomic Aspects
ofAnnonaceae 117
ff60
50
>. 40oco=
30cr9L 20 10
o
7s9 789
Asia America
Fig.2. Proponion ofgenera with
basic
chromosome number of x=7(dotted),
8
(stripes)
and 9(solid),
patterns
similar tothatin
theAnnonaceae. Such
analyses might clarify theprocess
of speciesdiversification in tropical forests in
each region.Difflerences in
theproportion
ofpolyploids between Asia
andAmerica
mightindicate
therole of polyploidyin
speciation(Fig. 3). Polyploids
are observedfrequently in American
species ofAnnonaceae, but
are very rarein Asian
ones.According to
previous reports(Okada
andUeda,
1984; Morawetz, 1984a, 1986a, b, 1988), karyological
obseryations of American plants were mainly carriedout on speciesfrorn
rather xerichabitats,
asin
theBrazilian Cerrado,
whileAsian
materials were collected mainly
from
thewet tropicswith veryhigh humidity. In general, floras
athigh Iatitudes, high
altitudes andin harsh
environments contain ahigher proponion
of polyploids(cf. Funabiki, 1967; Grant,
1981).Polyploids
are composed of multiplegenome
sets, ofwhich twofundarnental
setsassurethe
originalgenetic
system, while theadditional setshave
the ability to changegenetic
systems to allow the organism to adapt toharsher
environments.Another
%
>oco]croLL
1OO
75
50
25
o
Asia America
Fig.3. Proportionof
diploids
and polyploidsinAsiaand America.118
Acta Phytotax.Geobot. Vol. 44
Tablc2. PloidylevelsandbasicchromosomenumbersofAnnonaceaeinAsiaandAmerica. Apomictictaxaexcluded.
Nurnerals in
parenthesesindicatenumber of species observed and totalnumber ofspecies.ploidy
basicnumber Asia(51
genera)America
(36genera)
2x x==7
Mlezzettia (217)
Annona(121125)
Rollina
(2165)
7letrameranthus (112)
x=8 Antzxagorea
(1129)
Artabot,}ts
(6floo)
Cananga
(2f2)
Cleistopetatum
(112) Cyathoealyx (6125)
Desmos
(S/2S)
Disepalum
(118)
Mssistigma
(2180)
1#}'ieyodielsia
(2140)
Goniothalamus
(6180)
Mlelodorum(314)
Mitrella(11S)
Ilyramidanthe(111) Ra"wenhqt77a
(2f5)
Ctvaria
(21175)
Jkylopia
(2f170)
Anczxagorea
(2129)
Asimina
(7!8)
Bocageopsis (113)
Desm
opsis(1/16)
Duguetia
(15174)
Htsaea
(113)
Alylopia (87170)
x=9
Aiphonsea (3130)
Ancana (213)
Enicosanthum
(1116)
FVtzalania
(1/2)
Hdptostichanthus
(5!6)
Mbzzettiopsis(111)
Miiiusa(3140)
Mitrephora (3140)
IVbouvaria
(ll3)
Oncodostigma
(215)
Orophea (2160)
Phaeanthus
(2120)
Platymitra
(112)
Polyalthia
(141150)
Polyaulax(112?) Pqpowia (1140)
Ilseuduvaria
(6135)
StelechocaJ;ptts(212S)
Cymbqpetalum (2113)
Porcelia (lfS)
Sapranthus(1112)
3x
x=8 Asimina(118)x=9
Ombopetalum (1113)
The Japanese Society for Plant Systematics
NII-Electronic Library Service
The JapaneseSociety for Plant Systematics
December 1993 OKADA:Cytotaxonomic Aspects
ofAnnonaceae 119 Table 2.
continuedploidy basicnumber
Asia(51genera)
America
(36genera)
4x x==7 Annona
(2/125)
Guatteria
(241250)
Guatteriella
(112)
Guatteric\isis
(415)
RoUinia (2165)
7letrameranthus
(112)
x=8 Duguetia
(3174)
x=9 Enieosanth"m
(1116)'
Polyalthia
<11150)
6x x=7 RoUinia
(3!65)
x=8
Anczxtrgorea (1129)
Duguetia
(1174)
8x x=:7 RoUina
(116S)
x=8
Cyathocalyx (112S)
x=9 Clymbopetatum
(1113)
" :
Polyalthia
grandifbliais
treated as a synonym of Enicosanthum grand(fbtiumTable3.Number of generawith basicchromosome numbeT ef 7,8and
9.
basicnumber
Asia
(51)
number of genera
Africa
(40) America (36)
totalx==7x=8x=9 11618 361 673 102922
total
35
10 1661
explanation
is
thatnew regions, where the original vegetation mayhave been destroyed,
supplyhabitats for
newcomers.Some
newcomers mayhave
originatedfrom hybrids between parental
species which grow
in
stablehabitats. The hybrids
are thought tohave
a greaterability toadaptto harsher
environment than theparental
speciesbecause
they contain aheterogeneous
genomeinherited
fromboth parental
species.But
usually they are sterilebecause
ofhybrid
weakness or some other reasons.The polyploidization guarantees
continuation offurther
generationsby
formation
offertile gametes (Stebbins, 1984, 1985). In
any cases,polyploids
seem to tolerateharsher
environments.A high
proportion of polyploidsin the Brazilian Cerrado
mayhave
arisen from an evolutionary strategy toadapt toxeric environments, notjust because
of thelong history
of the
family. It is
thereforepresumed
thatif
we observethe
chromosomes of species of wetforests in America,
alow frequency
ofpolyploidy
mightbe
observed.120 Acta Phytotax. Geobot. Vol. 44
I
wouldlike
toexpress my cordial thanks toDrs. M. Hotta, Kagoshima University,
andM.
Kato, University
ofTokyo,
whoprovided
opportunities tostudyin Indonesia. Thanks
arealso
due to
Mr. Mohd Khanbin
MominKhan, Director General for Wildlife
andNational Parks, Malaysia, Dr. Fransis Ng,
theformer Deputy Director General, Forest Research Institute
ofMalaysia, Malaysia, for kindly
arrangingfor the
survey atTaman Negara, MaEaysia,
and toMr.
T. Hainald, LIPI, Indonesia, and Dr.
J. R. Croft, Acting Director, Division
ofBotany, Department
ofForests, Lae, Papua New Guinea, for
theirkind
support and arrangement of myfield
studies.Dr. L. W. Jessup, Senior Botanist, Queensland Herbarium, Australia, kindly
permitted
me touseherbarium
equipment andgave
mefresh
seeds.Dr. S.-M. Chaw, Institute
ofBotany, Academia Sinica, Taiwan, provided fresh
seeds.References
Bawa, K.S.1973.Chromosome numbers of treespecies of a lowlandtropical community. J.Arnold Arb.
54:422-434.
Cronquist,A. 1981.An IntcgratedSystemofClassMcation ofFlowering Plants.Columbia Univ,Press,New
York.1262pp.
Datta,P.C.and De, B. 1990.Karyologyof some IndianAnnonaceae.
Cytologia
5S:187-196,Ehrendorfer, F., Krendl, F.,
Habeler, E.andSauer,
W. 1968.Chromosomc numbers and evolutionin
primitive
angiosperrns. Taxon 17:337-353.Fries,R. E. 1959.Annonaceae.
in
Melchior,H.(ed.)
Die NaturlichenPflanzenfamilien2.Aufl,17all.Berlin,Duncker and Humblot.
Funabiki,K. 1967.A study on the relationship
between
chromosomalfeatures
of angiosperm fioraandlatitude,temperature and vegetation zones alongthe Japanese
islands.
J.Jap.For.Soc,
49:379-385, Grant,V. 1981.PlantSpeciation,
2nd ed.563pp.Columbia Univ.Press,New York.Housden, E.C. H. van. 1992,Flowers of
Annonaceae:
morphology, classificationand evolution, Blumeasuppl, 7:1-218.
Klucking, E,
P. 1986.Leaf VenationPatterns.vol. 1.Annonaceae. J,Cramer, Berlin.Le Thomas, A, 1981. Ultrastructural
characters of pollengrainsof AfricanAnnonaceae and theirsignificancefor
thephylogeny of primitiveAngiosperms.
Pollenand Spores22:265-342,and 23:5-36.Metcalfe,C.R, 19S7,Annonaceae. Anatomy of the Dicotyledons.2nd Ed. vol. 3.34-48.ClarendonPress,
Oxford.
Morawetz, W. 1984a.Karyologicalraces and ecology of theBrazilianDuguetia
juditracea
as compared withX),lopiaaromatica
(Annonaceae).
Flora175:195-209.
.
1984b.How stable are genomes of trepicalwoody plants?HeterozygosityinC-banded karyotypes of Porceliaas comparcd with Annona(Annonaceae)
and Drinu,s(Winteraceae).
Pl. Syst.Evol.145:29-39.
1984c.6.Karyology. inMaas, P. J.M. and Westra,L. Y.
Th, Studies in Annonaceae II. A
monograph of thegenus Anczxagorea A. St, Hil.
part1. Bot.Jahrb.Syst.105:73-134..
1986a.Remarks on karyologicaldifferentiation
patternsintTopical woody plants. Pl.Syst.Evol.152:49-1oo.
,
1986b.Systematicsandkaryoevelution in Magnoliidae,
7letrameranthusas compared with other Annonaeeae generaof thesame chromosome number. Pl,Syst.Evol.154:147-173.
.
1988.Karyosystematicsand eyolution ofAustralianAnnonaceae as compared with Eupomatiaeeae,Himantandraceae and Austrobaileyaceae.
Pl. Syst.
Evol,IS9:49-80..
and LeThomas, A. 1988.Karyologyand systematics of thegenusAmbavia and other Annonaceae from Madagascar. Pl.Syst.Evol,158:155-160.
.
and Waha, M. 1985.A
new pollentype,C-banded
and fiuorochromecounterstained chromosomes,and evolution inGuatteriaand related genera
(Annonaceae).
Pl.Syst.Evol.150:119-141.The Japanese Society for Plant Systematics
NII-Electronic Library Service
The Japanese Sooiety for Plant Systematios
December 1993
OKADA
:Cytotaxonomic
Aspects
ofAnnonaceae 121
Okada , H.
1990.
Reproductive biology of Polyalthialittoralis (
Annonaceae) .
P1.
Syst
』 voL 170:237−245.
and
Ueda
,K 。
1984.
Cytotaxonomical
studies on Asian Annonaceae.
P1.
Syst.
Evol.
144:165−
177.
Sauer,
W.
a皿d Ehrendorfer, F,
1984.
Notes on the karyosystematics of Annonaceae.
PL Syst.
EvoL146:47
−
55.
Stebbins ,
L1984.
Polyploidy and thedistribution
of the arCtic−
alpine且
ora: new evidence and a newapproach
.
Botanica Helvetica 94:1−
13.
.
1985,
Polyploidy, hybridization,
and the invasion of new habitats,
Ann,
Missouri Bot,
Gard.
72:
824−832.
Walker,
J.
W .1971.Pollen
morphology,
phytogeography and phylogeny of theAnnonaceae,
Contr.
Gray
Herb
.
202:1−
123,
摘 要
岡
田
博 :ア ジ ア
産バ
ンレ
イシ
科 植 物の
細 胞 分 類 学 的 特 質と ア メ リ カ
産の も の と の
倍 数 体 頻 度の 相違
バ ン レ
イ
シ科植物 は 日
本に は 沖
縄地 方
(八
重 山諸島
)に た だ 1
種, ク ロボ ウ
モド キ
(Po
/yalthia
liukiuensis
Hatusima
)カ ミ
分布
する だ け で 我
々に な じ み
の薄
い植物 で
ある
。し か し ,
こ の科 は ア
ジ
ア, ア
フリ カ ,
ア メリ カ の 熱帯
,亜 熱帯 に 広
く分布 し ,約130属
,2300種
(Cronquist
1982
)を 含 み ,
原 始 的被
子植物
の中 で
も大き な
1群 で あ る
。 こ の科
の染
色体
基本数 に は 7,8, 9
が 知ら れ る
。 これ ら の 染
色体基
本数 は
, 原 始 的 被 子植物 と
言わ
れる 植物群 の 中 で
も最
も少 な い
もの の 1
つ
に
上 げら
れる
(例え
ばモ ク レ ソ科 は
x=
19, デ ゲ ネ リ
ア科 は
x=
12,
シキ
ミ モド キ 科 は
x=13,
ユウ ポ
マチ
ア科は
x=10 な ど で い ず れ
もパ ン レイ
シ科よ り大
きい)。細胞
学 的な
観 点か
ら
する と , こ
の 植 物 群は
原 始 的被
子植物
の中 で も 根幹 を 占 め る
べき も
のと
言わ
ざる を 得 な
い 。従っ
て
, その
細 胞 分 類 学 的 特 質は
原 始 的 被 子 植 物の
系 統 進 化を
考え る 上 で
無 視で
きな
い 重 要な
情報 の 1
つで
ある
。一 方
,こ の
科は
熱 帯に お
いて 上
記の よ
うに
多 くの
分 類 群に
分 化し た ,
いわ
ぽ熟帯 で
成功 し て い る
科で
あり ,
熱 帯に お
ける
被 子 植 物の
進 化 的 傾 向,
種 分 化の
機 構 な どを 研 究 する
のに
是 非 取 り扱いた
い 植 物 群の 1
つで
ある
。こ の研
究 は
以前 の
研究
(Okada & Ueda ,1984
)に
続い て
行 なわ
れた
もので ,
東 南ア ジ ア
に分 布 する
パ ン レイ
シ科植物
, 19属24
種の
染 色 体 数 を 観察 し た
(Table
1
)。こ の
うち 8
属の
染 色 体 数は
今 回
初め て 明 ら か に さ れ た
もの で
ある
。 今 回の
もの
も含め
現 在ま で に ア ジ ア
地 域に分 布 する
約51 属 の
うち 35
属,ア
フリ カ
地 域に
分 布 する
約40
属の
うち 10
属,
そし て ア
メリ カ
地 域に
分 布 する
約 36属
のう ち
16属 の 染色体数 が
明ら か に な
って
いる 。
これら
のう ち ,
アジ
アと
ア メリ カ 地
域の染
色体 に 関 す る 特徴 を 比
較した
結 果 (Table
2 )
, 以 下に
記 すよ
うなこ と が わ か
った
。 なお
,ア
フ
リ カ
地 域の ものは
報 告 数が
少な く 比 較
の 対 象に し な か
った
。1
) 染 色 体 基 本 数は
x=8 を 示
す 属が
い ずれ
の地
域に お
いて も
高い 頻度 を 示 し た
。 この
事 実は Okada & Ueda
(1984
),
Morawetz
(1986
)が
提案 し た 「
こ の科
の 原 始染 色 体
基 本数は 8 で
ある
」と い
う主
張を
支 持 する
もの で
ある
。一
般に
, ある グ
ルー プ
が発
生し た と き
の 原初 に 持
って い た
特質 は , そ
の後 そ
のグ
ルー プ の 発
展に と
もな
って
多 くの派 生 形質 が
生じた 時 で
も維持 さ れ
続 ける
可能
性 が高
い 。
2
) アジ
ア,
ア フ リ カ , ア メリ
カの 2
〜3
地 域に ま た が
って
分 布 する
属の染 色 体 基 本 数に は 7
( Annona )
,8
(Anaxagorea ,
Artabotrys
,Uvaria
,Xytopi
の, 9
(Polyaithia ) の
い ず れ もが
ある 。
これ は
バ ソ レイ
シ科 植 物の染
色体
基 本数
の変化 が 3 地
域が 分
離 する
前に
起 こ った
こと
を示 唆 する
。ま た ,1
)の主
張の
繰 り返し に
なる が , こ
れら
の属に
染 色 体 基 本 数,8
が 多くみ ら れ る
こと は ,
N工 工
一
Eleotronio Library122 Acta
Phytotax.
Geobot . Vo1 .44
は や
りx; 8
がこの
科の
原 初の 特質 で
あった
こと を
示唆
する
。3
) アジ
アと
ア メリ カ の
バ ソ レイ
シ科植物 の 染
色体
基 本数 の
頻 度を 見 る と , 両 地
域の 間で
進 化の 傾 向の 違いが み
られる
。 アジ
アで は
x= 9
の属 が
多 く派生 し
,一 方
ア メリ
カ では ’
x
=
7の属
が多
く派 生し て い る
。こ の 違 い は
両地 域
が隔離 さ れ た 後 に 起
こ った
もので
あろ う
。 こ の様 な 進
化の
傾 向の
違い が
何 を 意 味 する
もの か
今 回の
研 究で は 解
明し 得 な い が
, 興味深 い 現象 で
ある
。4
)
アジ
アと
ア メ リ カ の 倍 数体
の 頻 度は 明 ら か に
違う 。
アジ
ア では ほ と ん
どの 種が 2
倍 体のま ま で
分化 し て
いる
のに
対し て ,
ア メリ カ
のも
ので は
多 くの 倍 数 体が
報 告さ れ て
いる 。 こ
れは
ア
ジ
アで
の 報 告は
主に 湿 潤 熱 帯,
ある
いは
そ れに 準 ずる
雨 緑 林に
分 布 する
種 を 用いた
もの で
ある
のに
対し て , ア
メリ カ で
の報 告 が 主に湿 潤 熱 帯 林に
分 布 する
種で は
な く,や
や 乾 燥し た
セラ ー
ド
地帯
(ブ ラ ジ
ル にみ ら
れる サ
バ ンナ植
生 )に 分布
する 植物 に 基 づ い て い る 事 が
大 き く影 響し て
い る よ
うに
考 え られる
。 ある
地 域の
植 生に
占め る
倍 数 体の
頻 度は
低緯
度か
ら高 緯 度に
なる に
従っ
て ,
ある い は
標 高が
高 くなる に
従って だ ん だ ん
増 えて
行 く傾 向が
ある こ と が
報告 さ れ て い る
(
Grant
1981
)。
そのよ
うな
環 境は
生 物の
生 育に と
って
きび し い
もの と
思 わ れる が ,
倍 数 体に は
ゲ
ノ ム セ ット が
複 数 あり ,
基 本 的な
遺 伝 子の セ ット に よ
って
最 低 限の 生 活が
保 証さ
れ,
他の セッ ト の 遺 伝 子
が
無 方 向に突 然 変 異 を起こし て ,
その うち
きび し
い 環 境に も耐 えて
行 くこ と
の 出 来る よ
うな
遺 伝 子の セ ット が
形 成 される か
らと
考 え ら れて
いる
。 ある
いは
別の 見 解 も ある
(