腸管マクロファージの形質制御機構の解明
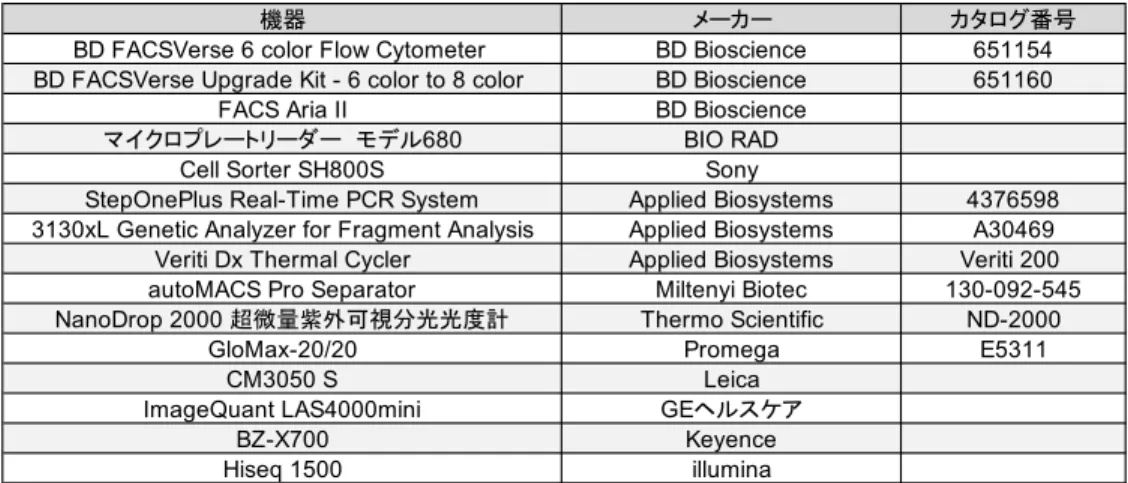
第2 章 実 験 方 法 使 用 機 器 ・ 試 薬 表1,2 に示す。 実 験 動 物 < 野 生 型 マ ウ ス > 日 本 ク レ ア か ら 購 入 し た 。(C57BL/6J、♀、6-12 週齢、体重 30g 以下) <Maf 胎仔肝・骨髄キメラマウス> Maf の機能を調べるためには Maf 完全欠損マウスを解析する必要があるが、当該 マ ウ ス は 出 生 前 後 に 致 死 と な る23。そ こ で 、CD45.1 の Maf+/-マ ウ ス お よ びMaf-/-マ ウ ス 胎 仔 肝 臓 か ら 、 造 血 幹 細 胞 を 含 む 胎 仔 肝 臓 細 胞 を 調 製 し 、 こ れ を 野 生 型 マ ウ ス に 移 植 す る こ と で 、造 血 幹 細 胞 由 来 細 胞 で の みMaf を欠損するキメラマウスを作製し た 。CD45.2 の野生型マウスに 8 Gy の X 線を照射して骨髄細胞を破壊した後、1 x 106か ら 5 x 106 個の Maf+/-胎 仔 肝 臓 細 胞 お よ び Maf-/-胎 仔 肝 臓 細 胞 を 静 脈 内 注 射 に よ り 移 植 し 、胎 仔 肝 キ メ ラ マ ウ ス を 作 製 し た 。移 植 か ら7-10 週間後の末梢血をフロ ー サ イ ト メ ト リ ー 解 析 し 、キ メ ラ 率 が 90%以上の個体を実験に用いた。胎仔肝キメ ラ マ ウ ス は 個 体 数 が 少 な い た め 、 こ の マ ウ ス の 骨 髄 細 胞 を 移 植 し た 二 次 移 植 キ メ ラ マ ウ ス を 作 製 し 、 個 体 数 を 確 保 し た 。 移 植 か ら2 ヶ月以降に、Maf 胎仔肝キメラマ ウ ス か ら 上 腕 骨 、大 腿 骨 及 び 脛 骨 を 摘 出 し 、CD45.1+の 骨 髄 細 胞 を 調 製 し た 。胎 仔 肝 キ メ ラ マ ウ ス と 同 様 の 方 法 で 野 生 型 マ ウ ス に 骨 髄 細 胞 を 移 植 し た 。 <Nrf2 欠損マウス> 東 北 大 学 医化学教室 山本雅之先生から分与を受けた51。 上 記 の 遺 伝 子 改 変 マ ウ ス に つ い て 、 ジ ェ ノ タ イ ピ ン グ に 使 用 し た プ ラ イ マ ー セ ッ ト を 表3 に示す。 炎 症 モ デ ル < デ キ ス ト ラ ン 硫 酸 ナ ト リ ウ ム (DSS)誘導大腸炎>
< 酢 酸 誘 導 大 腸 炎 モ デ ル > 野 生 型 マ ウ ス を6 時間絶食後、さらに 3 ml の下剤を(ニフレック、味の素)を経 口 投 与 針 ( 夏 目 製 作 所 ) で1 時間毎に、3 回投与した。下剤投与から 12 時間後、 軟 性 ゾ ン デ (FUCHIGAMI)で 100 µl 4% 酢酸を注腸した。10 秒後、直ちに 500 µl 生 理 食 塩 水 を3 回注腸し、酢酸を希釈した。 細 胞 の 調 製 < 末 梢 血 ・ 骨 髄 細 胞 > 末 梢 血 細 胞 に つ い て は 、 ヘ マ ト ク リ ッ ト 毛 細 管 を 用 い て 眼 窩 静 脈 叢 か ら 末 梢 血 を 採 取 し た 。氷 上 に て 、全 血20 µl を 1 x Pharm Lyse 200 µl で 5 分溶血後、MACS Running buffer (0.5% BSA / 2 mM EDTE / PBS)で洗浄し、遠心後のペレットをフローサイトメ ト リ ー 解 析 に 使 用 し た 。骨 髄 細 胞 に つ い て は 、マ ウ ス を 十 分 な 麻 酔 下 で 安 楽 死 さ せ 、 上 腕 骨 、 大 腿 骨 及 び 脛 骨 を 摘 出 後 、MACS running buffer が入った 60 mm 無処理デ ィ ッ シ ュ に 回 収 し た 。両 側 の 骨 端 を ハ サ ミ で 切 断 し 、断 面 に1 ml シリンジ (26G 針 付, テルモ) で MEMαを注入し、流出する骨髄細胞懸濁液を回収した。この細胞懸 濁 液 を ナ イ ロ ン メ ッ シ ュ で ろ 過 し て15 ml チューブに回収した。遠心後、上清を除 い た ペ レ ッ ト に2 ml の 1 x Pharm Lyse を加えて溶血し、洗浄した。 < 骨 髄 由 来 マ ク ロ フ ァ ー ジ (BMDM)、骨髄由来樹状細胞 (BMDC) の誘導> 骨 髄 細 胞 を 1 x 106細胞 / ml になるよう、MEMα / 10% M-CSF (細胞株 CMG-14-12 の培 養上清) / 10% FCS / 1% Penicillin-Streptomycin (以降 P/S)、もしくは RPMI-1640 / 10% GM-CSF (細胞株 MGM-5 の培養上清) / 10% FCS / 1% P/S に懸濁し、10 cm コートデ ィ ッ シ ュ 、 も し く は 60 mm 非コートディッシュに播種し、BMDM もしくは BMDC を 誘 導 し た 。 < リ ン パ 節 の マ ク ロ フ ァ ー ジ ・ 樹 状 細 胞 >
< 大 腸 粘 膜 固 有 層 の 細 胞 > マ ウ ス の 大 腸 を 摘 出 し 、内 容 物 をPBS で洗浄した。余分な脂肪や組織を取り除き、 腸 管 を 縦 方 向 に 切 開 し た 。 氷 冷 し た PBS で 再 度 洗 浄 し 、 2-3 等 分 し た 後 、 EDTA solution; 20 mM EDTA pH 7.2 / 2% FCS / 1x HBSS に入れ、湯浴で撹拌しながら 15 分 間 、37℃でインキュベートした。PBS で腸を洗浄し、残存した上皮細胞をピンセッ ト で 鈍 的 に 剥 離 し た 。 断 片 が お よ そ φ5 mm 以 下 に な る ま で 、 腸 を ハ サ ミ で ミ ン チ し 、Collagenase solution; 0.15 mg/ml Liberase TL / 0.5 mg/ml DNase I / 1% Dispase / 10 mM HEPES / 2% FCS / RPMI-1640 で 37℃, 100rpm, 30 分間、震盪した。その後、細 胞 懸 濁 液 を セ ル ス ト レ イ ナ ー で 濾 過 し 、4˚C, 500 x g, 10 分間遠心し、洗浄した。細 胞 を 濃 縮 す る 場 合 、更 にCD11b-microbeads を加え、30 分間、氷上でインキュベート し た 。MACS Running buffer で洗浄し、AutoMACS Pro により CD11b+細 胞 を 粗 精 製 し た(POSSELD モード)。
BMDM ま た は BMDC の 形 質 変 化 の 誘 導
BMDM と BMDC を 4 日間培養後、7.5 x 104 cells / 100 µl / well で 96 well plate に 播 種 し た 。24 時間後にリポ多糖 (LPS, O55:B5、100 ng/ml) またはマレイン酸ジエチ ル (DEM, 100µM) 刺激した。刺激 5 時間または 24 時間後に培養上清を回収しサイ ト カ イ ン 濃 度 をELISA で定量した。さらに細胞から RNA を抽出し定量 PCR に用い た 。 M1・M2 の形質変化を誘導する場合、10 ng/ml LPS と 20 ng/ml IFN-γ(M1 誘導)、 も し く は 20 ng/ml IL-4 存在下(M2 誘導)で培養し、24 時間後の細胞を溶解して RNA を抽出した。 酸 化 ス ト レ ス に 対 す る 応 答 を 評 価 す る 実 験 で は 、BMDM を培養 4 日後、2.0 x 104 cells / 100 µl / well で 96 well plate に播種した。24 時間後に 50 µM、100 µM、200 µM の 濃 度 に な る よ う にtBHP (tert-butyl hydroperoxide, Sigma)を添加した。24 時間後に、 Cell counting kit-8 (DOJINDO)を培養上清の 10%量を加え、2 時間後に 460 nm 吸光度 を 測 定 し て 生 存 率 を 評 価 し た 。
フ ロ ー サ イ ト メ ト リ ー ・ セ ル ソ ー テ ィ ン グ
細 胞 と 蛍 光 色 素 標 識 抗 体 を 含 む 全 て の 試 薬 はMACS Running Buffer に懸濁した。 Fc-Blocker を加えて 5 分後に、細胞を蛍光色素標識抗体の溶液に懸濁し、30 分間氷 上 で イ ン キ ュ ベ ー ト し た 。 二 次 抗 体 が 必 要 な 場 合 は 、MACS Running Buffer で洗浄 後 、更 に 当 該 抗 体 と20 分間、氷上でインキュベートした。その後、細胞を洗浄し 7-AAD もしくは DAPI (いずれも 1 µg/ml)を添加し FACSVerse で解析した。
い はSH-800 により分取し、RPMI-1640 (2 % FCS / 1 % P/S)もしくは TRIzol LS に回 収 し た 。 培 養 液 に 回 収 し た 細 胞 をPBS で洗浄後、RNA を抽出した。
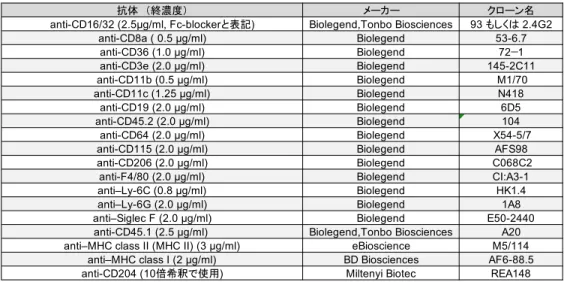
フ ロ ー サ イ ト メ ト リ ー に 用 い た 抗 体 は 表4 の通り。抗 CD169 抗体(Clone: M7)は当 研 究 室 で 作 製 し17、EZ-Link (Sulfo-NHS-LC-LC Biotin, ThermoFisher scientific)により ビ オ チ ン 化 し た 。 ビ オ チ ン 化 抗 体 は PE-Streptavidin (0.2 µg/ml, Biolegend)あるいは Alexa fluor 488-streptavidine (1 µg/ml, ThermoFisher scientific)で検出した。
プ ラ ス ミ ド
<ルシフェラーゼレポータープラスミドの構築>
Ccl8, Ccl8-mut1, Ccl8-mut2, Ccl8-mut3, Ccl8-mut4, Slpi -Luc-pGL4.10 (図 4, 6b,c)
<Maf 発現ベクター(Maf-pBOSEX)の構築>
Maf cDNA-pBlueScript プラスミド(筑波大学 解剖学発生学研究室 高橋智先生
か ら 分 与48) をEcoRI および NheI で制限酵素処理し、pEFBOS-EX プラスミドに挿 入 し た 。
<Maf 発現レトロウイルスベクター(Maf-pMSCV)の構築>
Maf-pBOSEX を、EcoRV および SacI で制限酵素処理し、得られた DNA 断片を
DNA Blunting kit で平滑化した。pMSCV-pgk-Venus ベクター (タカラのベクター: pMSCV-pgk-puro を改変)を HpaI で消化し、平滑化断片を組み込んだ。
<Nrf2 発現ベクター(Nrf2-pBOSEX)の構築>
野 生 型BMDM から作製した cDNA を鋳型に、Nrf2 遺伝子を PCR で増幅した。 PCR 産物および pEFBOS-EX を、KpnI および NheI で制限酵素処理しライゲーショ ン お よ び ク ロ ー ニ ン グ し た 。
<Maf 3´-UTR レポーターベクター(Maf 3´-UTR-psiCHECK2)の構築>
野 生 型BMDM から作製した cDNA を鋳型に、Maf 遺伝子の終止コドン直後の Maf 3´-UTR 領域を PCR で増幅した。PCR 産物および pBluescript II SK(+)を EcoRI お よ びNotI で制限酵素処理し、ライゲーションおよびクローニングした。このプ ラ ス ミ ド を XhoI および NotI で制限酵素処理し、psiCHEK2 (Promega)に組み込ん だ 。 MSCV レ ト ロ ウ イ ル ス 発 現 シ ス テ ム < ウ イ ル ス の 作 製 > パ ッ ケ ー ジ ン グ 細 胞 Platinum-E (Plat-E)細胞(東京大学 医科学研究所 先端医療 研 究 セ ン タ ー 細 胞 療 法 分 野 北 村 俊 雄 先 生 か ら 分 与 52) を ト ラ ン ス フ ェ ク シ ョ ン 前 日 に 、2 x 106 細胞 / 4ml DMEM (10% FCS / 1% P/S) になるよう懸濁し、60 mm コー ト デ ィ ッ シ ュ に 播 種 し た 。24 時間後、Maf-Venus もしくは
pMSCV-pgk-Venus(以後、コントロールと表記)各 5 µg を、Plat-E 細胞に Fugene6 を用いてトラ
抽 出 し た 。
4 日間培養後の BMDM を、5 x 104 cells / 200 µl / well の条件で 48 well plate に播 種 し た 。24 時間後の培地を、MEMα / 10% M-CSF / 10% FCS / 1% P/S / ウイルス培養液 / polybrene (8 µg/ml)に交換し、Maf-pMSCV-pgk-Venus トランスフェクションした。 感 染 16 時間後に培地交換し、さらに 40 時間培養した。この細胞に DEM(終濃度 100 µM)を添加し、さらに 5 時間後に、培地を除去し、LPS (100 ng/ml)で刺激した。 24 時間後に RNA を抽出した。 Total RNA 抽 出
RNeasy Mini kit もしくは miRNeasy Mini kit のプロトコルに従い、RNA を抽出し た 。 細 胞 は Buffer RLT もしくは TRIzol LS で溶解した。TRIzol LS で溶解した場合、 TRIzol LS : 細胞懸濁液が 3 : 1 になるよう液量を調製した。さらにこの液に対して、 1/5 量のクロロホルムを加え、有機層にタンパク質および DNA を、水層に RNA を 溶 解 し た 。 水 層 を 回 収 し 、 以 後 、RNeasy Mini kit の推奨プロトコルに準拠して操作 を 行 っ た 。
定 量RT-PCR (qRT-PCR) <mRNA の定量>
100 ng – 1µg の RNA を ReverTra Ace qPCR RT Kit で逆転写した。得られた cDNA を 鋳 型 と し 、THUNDERBIRD® qPCR Mix のプロトコルに準拠して qRT-PCR を行っ た 。18S rRNA の発現量を内在性コントロールとして、目的遺伝子の mRNA 発現量 を 相 対 的 に 定 量 し た 。qRT-PCR に用いたプライマーは表 6 に示す。
<miRNA の定量>
miRNA は、Taqman Micro RNA Assays (Applied Biosystems)キットのプロトコルに従 い 、逆 転 写 お よ びqRT-PCR を行なった。Sno135 を内在性コントロールとして、miR-129, miR-155 の発現量を相対的に評価した。
DNA マ イ ク ロ ア レ イ
microRNA-sequencing (miRNA-seq)
6 日間培養した野生型 BMDM 5x 105cells / 2ml / well を 6 well plate に播種した。 19 時間インキュベートした後、最終濃度 100 µM DEM を添加した。刺激から 5 時 間 後 、miRNeasy mini kit を用いて miRNA を含む全 RNA を抽出した。Agilent 2100 Bioanalyzer and Quantus Fluorometer (Agilent)により RNA 濃度を測定した。NEBNext Multiplex Small 274 RNA Library Prep Set for Illumina (NEB)を用いてアダプター配列 を 付 与 し た RNA ライブラリーを作製し、3% アガロースカセットをセットした BluePippin (Sage Science)により miRNA を分離した。精製した miRNA を Illumina Hiseq 1500 platform でシークエンスした。Strand NGS software (v2.9, Agilent Technologies)を用い、得られた配列データからアダプター配列をトリミングし、 mm10 ゲノムシークエンスにマッピングした後、2 サンプルの発現量を分位正規化 し て 統 合 し 、 散 布 図 を 作 成 し た 。
ル シ フ ェ ラ ー ゼ ア ッ セ イ
RAW264.7 細胞をもしくは HepG2 細胞を DMEM / 10% FCS / 1% P/S で懸濁し、1 x 105 cells / ml に調製した。細胞懸濁液 500 µl を 24 well plate に播種した。24 時間 後 に 発 現 ベ ク タ ー 、 レ ポ ー タ ー ベ ク タ ー 、 コ ン ト ロ ー ル レ ポ ー タ ー (pRLTK)のミ ッ ク ス をFuGENE6 によりトランスフェクションした。また、HEK293T 細胞 1x 104 個 を96 well plate に播種し、24 時間後に Maf 3´-UTR-psiCHECK2 および miRNA mimic (miRCURY LNA inhibitor, control or miR-155 or miR-129, exiqon)を、Lipofectamine 3000 に よ り ト ラ ン ス フ ェ ク シ ョ ン し た 。 ト ラ ン ス フ ェ ク シ ョ ン 後 24 時 間 に Dual Luciferase Kit (Promega)付属のプロトコルに準拠して細胞溶解液を作製し、ルシ フ ェ ラ ー ゼ 活 性 を GloMax-20/20 で測定した。Ccl8, Slpi, xCT-pGL4 のホタルルシフ ェ ラ ー ゼ 活 性 は 、pRLTK 由来のウミシイタケルシフェラーゼの発光量をコントロー ル と し て 発 光 量 を 相 対 的 に 評 価 し た 。Maf 3´-UTR のルシフェラーゼ活性は、同プラ ス ミ ド に 組 み 込 ま れ て い る ホ タ ル ル シ フ ェ ラ ー ゼ の 発 光 量 を コ ン ト ロ ー ル と し て 相 対 的 に 評 価 し た 。 ELISA
え 、1 時間室温でインキュベートした。5 回洗浄後、Assay Diluent で 250 倍希釈し たHRP-SA (BD Biosciences)を 50 µL/well 加え、30 分間室温でインキュベートし、7 回 洗 浄 し た 。TMB Microwell Peroxidase Substrate System (KPL)を加え、暗所で 30 分 間 、 室 温 で 発 色 さ せ た 。2N の H2SO4 25 µL/well で反応を停止し、450 nm の吸光度 を マ イ ク ロ プ レ ー ト リ ー ダ ー (Bio-Rad)で 測 定 し た 。 IL-6、 IL-10 の ELISA は BD OptEIA ELISA Set (BD Biosciences)を用い、メーカー規定のプロトコル通りに操作 し た 。
免 疫 組 織 化 学 染 色
マ ウ ス か ら 大 腸 を 摘 出 し 、 内 容 物 をPBS で洗浄した。5 mm 大に輪切りにした大 腸 をFrozen section compound (Leica)に浸し、液体窒素で凍結した。凍結した大腸を CM3050 S で 10 µm にセクショニングし、組織切片を 1 時間風乾した。内因性ペル オ キ シ ダ ー ゼ 活 性 を 阻 害 す る た め 、 組 織 切 片 を0.5% 過酸化水素 / メタノールに 浸 し 、 さ ら にBiotin blocking system (Dako)で内因性ビオチンをブロッキングした。 TN blocking buffer (ParkinElmer)でブロッキング後、ビオチン化抗 CD169 (clone: M7、フローサイトメトリーの項で記載した抗体と同様) および 抗α-SMA ウサギ IgG (clone: ab5694, abcam)で標識した。TSA Biotin System (ParkinElmer)を用いてビ オ チ ン 化 抗 体 を 増 感 し た 。 二 次 標 識 はCy3-streptavidin (The Jackson Laboratory)およ びAlexa Fluor 488-抗ウサギ IgG (Invitrogen)を用いて染色した。組織切片を
Fluorsave Reagent (Merck)/ DAPI (1 µg/ml)でマウントし、カバーガラスで封入し、 蛍 光 顕 微 鏡 BZ-X700 で蛍光を観察した。
ウ エ ス タ ン ブ ロ ッ ト
抗 体 (2000 倍希釈で使用、Dako)を加え、室温で 1 時間インキュベートした。Super signal west pico を用いて発色させ、LAS4000mini で検出した。撮影後、抗体除去バ ッ フ ァ ー (2% SDS, 100 mM 2-メルカプトエタノール, 62.5 mM Tris-HCl (pH 6.7))に メ ン ブ レ ン を 浸 し 、50˚C, 70rpm, 30min 震盪した。再度ブロッキング処理し、 HRP- anti-GAPDH (2000 倍希釈で使用、MBL)で 30 分間標識後、内在性コントロー ル を 検 出 し た 。 統 計 処 理 グ ラ フ に 示 す デ ー タ は SD もしくは SEM を記載した。2 群間の比較には Mann-Whitney U test を用いた。2 群以上の比較には one-way もしくは two-way ANOVA を 使 用 し た 。 全 て の 解 析 に お い て 、p 値<0.05 の時に有意差ありと判定した。
表1:使用機器
機器 メーカー カタログ番号
BD FACSVerse 6 color Flow Cytometer BD Bioscience 651154
BD FACSVerse Upgrade Kit - 6 color to 8 color BD Bioscience 651160
FACS Aria II BD Bioscience
マイクロプレートリーダー モデル680 BIO RAD
Cell Sorter SH800S Sony
StepOnePlus Real-Time PCR System Applied Biosystems 4376598
3130xL Genetic Analyzer for Fragment Analysis Applied Biosystems A30469
Veriti Dx Thermal Cycler Applied Biosystems Veriti 200
autoMACS Pro Separator Miltenyi Biotec 130-092-545
NanoDrop 2000 超微量紫外可視分光光度計 Thermo Scientific ND-2000
GloMax-20/20 Promega E5311
CM3050 S Leica
ImageQuant LAS4000mini GEヘルスケア
BZ-X700 Keyence
表2:使用した試薬(ここに記載のないものは本文中に記載する)
試薬 メーカー カタログ番号
MicroAmp® Optical Adhesive Film ABI 4311971
MicroAmp® Optical 96Well Reaction plate Applied Biosystems N8010560
Dispase BD Bioscience 354235
Cell Strainer, 70mm BD Bioscience 352350
OptEIAAssay Diluent BD Biosciences 555213
BD Pharm Lyse Lysing buffer (10x conc.) BD Bioscience 555899
6well Clear TC-Treated Microplates Corning 3516
24well Clear TC-Treated Microplates Corning 3526
48well Clear TC-Treated Microplates Corning 3548
96well Clear TC-Treated Microplates Corning 3596
60mm 無処理ディッシュ Corning 430589
HBSS (10X), calcium, magnesium, no phenol red GIBCO 14065056
96-Well Half-Area Microplates Greiner 675061
BigDye® Terminator v3.1 Cycle Sequencing Kit Invitrogen 4337455
TRIzol LS Reagent Life Technologies 10296010
Immobilon P Merck IPVH00010
デキストラン硫酸ナトリウム MW36,000~50,000 MPBiomedicals 591-18791
Phusion High-Fidelity DNA Polymerase New England Biolabs M0530
QIAprep Spin Miniprep Kit QIAGEN 27104
QIAGEN Plasmid Maxi Kit QIAGEN 12162
RNeasy Mini Kit QIAGEN 74104
miRNeasy Mini Kit QIAGEN 217004
cOmplete、EDTAフリー、プロテアーゼ阻害剤カクテル Roche 11873580001
グリコーゲン Roche 10901393001
Liberase TL Roche 5401020001
D-MEM High Glucose Wako 043-30085
マレイン酸ジエチル Wako 059-02052
MEMα (L-グルタミン、フェノールレッド含有) Wako 135-15175
Penicillin-Streptomycin Solution (×100) Wako 168-23191
RPMI-1640 (L-グルタミン、フェノールレッド含有) Wako 183-02023
デキストラン硫酸ナトリウム5000 Wako 196-13401
Recombinant Murine IL-4 Peprotech 214-14
Recombinant Murine IFN-γ Peprotech 315-05
Dual-Luciferase Reporter Assay System Promega E1910
FuGENE® 6 Transfection Reagent Promega E2691
GoTaq DNA Polymerase Promega M3001
DNaseI Worthington biochemical LS002139
DNaseI Sigma DN125-1G
Collagenase Iis Sigma C1764
臭化ヘキサジメトリン (polybrene) Sigma H9268-5G
リポポリサッカリド 大腸菌055:B5由来 Sigma L2880-10MG
Pierce BCA Protein Assay Kit Thermo Scientific 23227
SuperSignal West Pico PLUS Chemiluminescent Substrate Thermo Scientific 34579
Nunc イージーディッシュ (60 mm, 100 mm) Thermo Scientific 150462, 150464
Lipofectamine 3000 Thermo Scientific L3000001
ReverTra Ace qPCR RT Master Mix TOYOBO FSQ-301
THUNDERBIRD SYBR qPCR mix TOYOBO QPS-201
DNA Blunting kit タカラ 6025
Gel/PCR Purification Mini Kit (100) チヨダサイエンス FAGCK 001
Total RNA Extraction Column チヨダサイエンス FARBC-C50
HEPES ナカライテスク 17557-94
MG132 (Z-Leu-Leu-Leu-H) ペプチド研究所 3175-v
7-AAD Biolegend 420403
表3:ジェノタイピング PCR に使用したプライマー
表4:フローサイトメトリー解析に使用した抗体
表5:プラスミド構築に使用したプライマーセット
Gene name Sequence (5'→3')
Maf WT Fwd CTGCCGCTTCAAGAGGGTGCAGC
Rev TCGCGTGTCACACTCACATG
Maf targeted Fwd TGTTCACGTTCGAGCTTTC
Rev GATTAAGTTGGGTAACGCC
Nrf2 Fwd TGGACGGGACTATTGAAGGCTG
lacZ GCGGATTGACCGTAATGGGATAGG
Rev GCGGATTGACCGTAATGGGATAGG
Gene name Sequence (5'→3')
Nrf2 Fwd ATAGGTACCCGCCCTCAGCATGATGGACT Rev GGCCGCTAGCCTAGTTTTTCTTTGTATCTG Ccl8プロモーター Fwd ATAGGTACCTTCATTTCTATGTTTCAGAATCCCTG Rev TAATCTCGAGTGTTGAAGGCAAAGATTTTGGAGTGAAG Ccl8プロモーター mut1 Fwd CCAGTGGGACTGCTTCCTCCAGAAGAGAGGTTTCAGATGCTTGCCC Rev GGGCAAGCATCTGAAACCTCTCTTCTGGAGGAAGCAGTCCCACTG Ccl8プロモーター mut2 Fwd AGATTTTAGCATCTTATTCTGAAGAGACTGCCTTCCAGCTGCCGGGA Rev TCCCGGCAGCTGGAAGGCAGTCTCTTCAGAATAAGATGCTAAAATCT Ccl8プロモーター mut3 Fwd ATCTCATGATCTGATGACTATCTCTTCTAACAAAGATCTTGCTTTCA Rev TGAAAGCAAGATCTTTGTTAGAAGAGATAGTCATCAGATCATGAGAT Slpiプロモーター Fwd AGCAGGTACCAGGACACCACAGCTCCACGC Rev TAATCTCGAGCAGGGGAGCTCTGATGACCA
Maf 3'-UTR Fwd GTCCGGAATTCGACGCCTACAAGGAGAAATACGAG
Rev ATAAGAATGCGGCCGCCGTCACGCGTGGTTAGTTAGTA
抗体 (終濃度) メーカー クローン名
anti-CD16/32 (2.5µg/ml, Fc-blockerと表記) Biolegend,Tonbo Biosciences 93 もしくは 2.4G2
anti-CD8a ( 0.5 µg/ml) Biolegend 53-6.7 anti-CD36 (1.0 µg/ml) Biolegend 72−1 anti-CD3e (2.0 µg/ml) Biolegend 145-2C11 anti-CD11b (0.5 µg/ml) Biolegend M1/70 anti-CD11c (1.25 µg/ml) Biolegend N418 anti-CD19 (2.0 µg/ml) Biolegend 6D5 anti-CD45.2 (2.0 µg/ml) Biolegend 104 anti-CD64 (2.0 µg/ml) Biolegend X54-5/7
anti-CD115 (2.0 µg/ml) Biolegend AFS98
anti-CD206 (2.0 µg/ml) Biolegend C068C2
anti-F4/80 (2.0 µg/ml) Biolegend CI:A3-1
anti–Ly-6C (0.8 µg/ml) Biolegend HK1.4
anti–Ly-6G (2.0 µg/ml) Biolegend 1A8
anti–Siglec F (2.0 µg/ml) Biolegend E50-2440
anti-CD45.1 (2.5 µg/ml) Biolegend,Tonbo Biosciences A20
anti–MHC class II (MHC II) (3 µg/ml) eBioscience M5/114
anti–MHC class I (2 µg/ml) BD Biosciences AF6-88.5
表6. qRT-PCR に用いたプライマー配列 Gene name Sequence (5'→3')
以 前 の 研 究 で 、 骨 髄 細 胞 を GM-CSF 存在下で培養した細胞(BMDC)は、Maf を 発 現 せ ず 、LPS 刺激しても CCL8 を産生しないことを確認している 17,26。 こ の 細 胞 に 、Maf を強制発現させることで CCL8 産生能を獲得するか検討した。レトロウイ ル ス ベ ク タ ー を 用 い て 、BMDC にコントロールプラスミドもしくは Maf 発現プラス ミ ド を ト ラ ン ス フ ェ ク シ ョ ン し 、 さ ら に LPS 刺激した際の遺伝子発現を定量した。 そ の 結 果 、Maf を強制発現させた BMDC では、LPS 刺激に伴う CCL8 発現レベルが、 コ ン ト ロ ー ル と 比 較 し て 600 倍に亢進した(図 3e)。これらの実験結果は、Maf 発 現 が 、 マ ク ロ フ ァ ー ジ のCCL8 産生に必要十分であることを明確に示している。ま た 、こ こ ま で の 結 果 よ り 、Maf は CD169+マ ク ロ フ ァ ー ジ の 分 化 に は 必 須 で は な い が 、 マ ク ロ フ ァ ー ジ の 機 能 制 御 に 重 要 な 役 割 を 担 っ て い る こ と が 明 ら か と な っ た 。 Maf は CCL8 発 現 を 転 写 レ ベ ル で 促 進 す る
Maf は Maf recognition element, (MARE:5´-TGCTGA (G/C) TCAGCA-3´ もしくは half-MARE; 5´-AT rich- TGCTGA(G/C)- 3´)に結合し、標的遺伝子の発現を制御する
伝 子 群 に は 、Ccl8、Fgl2、Mmp13 や Il10 などが含まれていた(図 5a,b)。これらの遺 伝 子 に 共 通 す る 機 能 に つ い て 遺 伝 子 オ ン ト ロ ジ ー 解 析 を し た 結 果 、Maf 標的遺伝子 に は 炎 症 関 連 遺 伝 子 が 多 く 含 む こ と が 分 か っ た ( 図5c)。一方、Tnf, Arg1 など典型 的 なM1/M2 マーカーの発現は両群で同程度だった(図 5d,e)また、Maf が発現を抑 制 す る 遺 伝 子 に は 、Slpi や Slc7a11 (xCT コーディング遺伝子), Cxcl1 など炎症の回復 期 や 慢 性 期 に 発 現 が 亢 進 す る 遺 伝 子 が 数 多 く 含 ま れ て い た( 図5f)29-32。我 々 は 、こ の よ う な 遺 伝 子 の 多 く が 、酸 化 ス ト レ ス 応 答 の マ ス タ ー 転 写 因 子Nrf2 の制御下にあ る こ と に 着 目 し た ( 図 5g)。最近、Nrf2 は、酸化ストレス応答以外にも、組織保護 や 抗 炎 症 作 用 に 関 与 す る こ と が 明 ら か に な っ て き た 33,34,35。 こ れ ら の 報 告 と 我 々 の 解 析 は 、Maf が、炎症関連遺伝子の発現を正に制御する一方で、組織保護・抗炎症 遺 伝 子 の 発 現 を 負 に 制 御 す る こ と を 示 唆 す る 。Slpi や xCT 以外に、他の Nrf2 標的 遺 伝 子 の 発 現 を Maf が抑制するか検討した。代表的な Nrf2 標的遺伝子として知ら
れ るHmox1, Nqo1 発現レベルを定量したところ、これらの遺伝子は Maf+/- BMDM お
よ びMaf-/- BMDM で差が見られなかった(図 5h)。このことは、Maf が全ての Nrf2 標 的 遺 伝 子 制 御 に は 関 与 し な い こ と を 示 唆 す る 。 続 い て 、Maf の欠損とそれに伴う Nrf2 標的遺伝子の発現亢進が、酸化ストレスに 対 す る 抵 抗 性 を 上 昇 さ せ る か 検 討 し た 。Maf+/-お よ び Maf-/- BMDM に tBHP で酸化 ス ト レ ス を 与 え た 際 の 生 存 率 を 検 討 し た 。こ の 結 果 、Maf-/- BMDM は酸化ストレス に 抵 抗 性 を 持 つ こ と が 分 か っ た ( 図5i)。この結果も、Maf による特定の Nrf2 標的 遺 伝 子 の 転 写 抑 制 機 能 を 反 映 し た も の と 考 え ら れ た 。 組 織 保 護 遺 伝 子 の 発 現 はMaf が 競 合 的 に 抑 制 す る
反 対 に 、Nrf2 が Maf の転写活性を阻害し、急性炎症応答遺伝子の発現を抑制する か 、Nrf2-/-マ ウ ス の 骨 髄 か ら 誘 導 し たBMDM を用いて検討した。これまでの報告で、 マ レ イ ン 酸 ジ エ チ ル(DEM)による酸化ストレス刺激は、Nrf2 を活性化し、その遺 伝 子 発 現 を 亢 進 す る こ と が 知 ら れ て い る 37。 図 6d に示すように、Nrf2+/- BMDM で はDEM 刺激による Slpi, Slc7a11 発現レベルが亢進するが、Nrf2-/- BMDM では DEM で 刺 激 し て も 、こ れ ら の 遺 伝 子 発 現 が 亢 進 し な か っ た( 図6d)。一方で、Nrf2+/-お よ びNrf2-/- BMDM の、LPS 刺激に伴う Maf 標的遺伝子 Ccl8 や Il10 発現レベルは同程 度 で あ っ た ( 図6e)。以上の結果は、マクロファージにおいて、Maf は Nrf2 の転写 活 性 を 拮 抗 的 に 抑 制 す る が 、Nrf2 は Maf の転写活性を抑制しないことを明確に示し て い る 。 Maf の 発 現 量 に 応 じ て マ ク ロ フ ァ ー ジ の 形 質 が 変 化 す る 我 々 は 前 項 の 実 験 で 、DEM に よ る 酸 化 ス ト レ ス 刺 激 が 、 Maf 発 現 レ ベ ル お よ び Maf タンパク質発現量を減弱することに着目した(図 7a)。この結果は、酸化ストレ ス が 、Nrf2 を活性化すると同時に、Maf の活性を抑制することを示唆する。Nrf2 が 制 御 す る 組 織 保 護 ・ 抗 酸 化 ス ト レ ス 応 答 は 、 炎 症 収 束 期 に お け る 組 織 修 復 に 重 要 な 役 割 を 担 う 38。こ の こ と か ら 我 々 は 、炎 症 回 復 期 に お け る 大 腸CD169+マ ク ロ フ ァ ー ジ で は 、 酸 化 ス ト レ ス 刺 激 下 で Maf による Nrf2 の抑制が解除され、組織保護的形 質 に 転 換 す る こ と を 想 定 し た 。 こ の 仮 説 を 証 明 す る た め 我 々 は 、DEM 刺 激 に よ る Maf 発現低下が、マクロファージの形質の変化に関与するか検討した。野生型 BMDM を 、DEM 刺激した後に LPS 刺激し、Maf ならびに Nrf2 の標的遺伝子発現レベルを 定 量 し た 。DEM によりあらかじめ Maf を抑制した BMDM では、Ccl8, Il10 など Maf 制 御 下 に あ る 急 性 炎 症 応 答 遺 伝 子 の 発 現 上 昇 が 抑 制 さ れ た 一 方 で 、Nrf2 制御下にあ
るSlpi, Slc7a11 の発現が劇的に亢進した(図 7b)。この変化が、主として Maf に依
る こ と か ら 、 我 々 は 、 プ ロ テ ア ソ ー ム 経 路 が 関 与 す る 可 能 性 を 考 え た 。 そ こ で 、 DEM 刺激した BMDM に、プロテアソーム阻害剤 MG132 を添加し、Maf のタンパ ク 質 発 現 量 を ウ エ ス タ ン ブ ロ ッ ト で 評 価 し た と こ ろ 、MG132 は DEM による Maf の タ ン パ ク 量 減 少 を 阻 害 し た ( 図8a)。DEM 刺激による Maf 発現の低下は、mRNA レ ベ ル で も 見 ら れ る こ と か らDEM 刺激による Maf タンパク質の発現低下には、プ ロ テ ア ソ ー ム 以 外 に も 、 転 写 抑 制 や 転 写 後 調 節 と い っ た 、 何 ら か の 調 節 機 構 が 関 与 し て い る 可 能 性 が あ る 。T 細胞に発現する Maf がマイクロ RNA(miRNA)-155 に よ っ て 負 の 制 御 を 受 け る こ と が 報 告 さ れ て い る こ と か ら39,40、 マ ク ロ フ ァ ー ジ に お い て も 、 同 様 の 機 序 が 働 い て い る 可 能 性 を 考 え た 。 し か し 、 予 想 に 反 し て 、DEM 刺 激 し たBMDM の Mir155 の発現を PCR で定量したが、定常状態と比較して発現 の 亢 進 は 見 ら れ な か っ た ( 図8b)。マクロファージでは、miR-155 以外のマイクロ RNA が Maf の分解に関与する可能性を考え、BMDM において、DEM 刺激により 発 現 が 変 化 す るmiRNA を miRNA-seq により網羅的に解析したところ、定常状態と 比 べ て10 倍以上亢進する miR-129 を同定した(図 8c,d)。さらに、miR-129 が Maf mRNA に直接結合して遺伝子発現を抑制するか、ルシフェラーゼアッセイにより 検 討 し た 。miRNA は、mRNA の 3´-UTR 領域に結合し、mRNA の分解を促進する 39,40。 ル シ フ ェ ラ ー ゼ 遺 伝 子 の 下 流 に Maf mRNA の 3´-UTR 領域を組み込んだレポ ー タ ー ベ ク タ ー を 作 製 し 、miRNA とともに、細胞株 HEK293T にトランスフェクシ ョ ン し た 。miR-129 導入は、コントロール miRNA を導入した場合と比較して、ル シ フ ェ ラ ー ゼ 活 性 が 有 意 に 減 少 し た ( 図8d)。この結果は、miR-129 が Maf mRNA の3´-UTR に結合し、mRNA 発現抑制や、分解に寄与することを示す。以上より、 酸 化 ス ト レ ス がMaf タンパク質発現を 1) miRNA による mRNA の転写後調節と、 2)翻訳された Maf タンパク質のプロテアソーム経路による分解という、2 つの過 程 で 抑 制 す る こ と を 示 す 。 腸 炎 の 進 展 に 伴 い マ ク ロ フ ァ ー ジ の 形 質 が 変 化 す る 我 々 は 、 こ こ ま で の 研 究 で 、Maf の発現量に応じてマクロファージが形質転換す る こ と を 、in vitro で証明した。そこで次に、同様の現象が腸管マクロファージで も 見 ら れ る か 検 討 し た 。DSS 誘導腸炎モデルを用い、マクロファージの遺伝子発 現 を 定 量 す る こ と で 、 炎 症 時 に お け る マ ク ロ フ ァ ー ジ の 形 質 を 調 べ た 。 腸 炎 の 急 性 期 で あ る4 日目には、Ccl8 発現が亢進していた。一方、腸炎の回復が開始する 8 日 目 に は 、 定 常 状 態 と 比 較 し てMaf の発現が減弱し、それに伴って Ccl8 発現の低
下 と 、Slpi, Slc7a11 発現の亢進が見られた(図 9a)。この結果は、腸炎の急性期に
の で あ り 、 マ ク ロ フ ァ ー ジ の 形 質 変 化 が ど の よ う に 制 御 さ れ て い る の か 、 と い う 免 疫 学 上 の 中 心 課 題 に 一 つ の 答 え を 提 示 し た も の で あ る 。 Maf に よ る 腸 管 マ ク ロ フ ァ ー ジ の 形 質 変 化 Maf は、広範囲の細胞種に発現し、細胞や組織の分化に重要な役割を担うことが 知 ら れ て い た 。 例 え ば 、Maf は、水晶体形成や、C57BL/6 マウスの胎仔肝における 2 次造血に重要な役割を担うことが報告されている48。 ま たAziz らは、Maf と、も う 一 つ のlarge Maf である MafB が、マクロファージの自己複製を阻害していること を 突 き と め 、Maf がマクロファージの分化にも何らかの役割を担うことを示した49,50。 我 々 は 、本 研 究 で 、Maf が、Nrf2 の転写活性を拮抗的に阻害することで、炎症経 過 に 伴 う マ ク ロ フ ァ ー ジ の 形 質 変 化 を 制 御 す る 、 と い う 新 機 能 を 発 見 し た 。 こ の Nrf2 は、もともと抗酸化ストレス応答遺伝子の発現を促進し、組織保護に中心的な 役 割 を 担 う こ と が 知 ら れ て い た が 、 最 近 、 炎 症 性 サ イ ト カ イ ン の 産 生 を 抑 制 す る こ と で 、 抗 炎 症 に も 積 極 的 に 関 与 す る こ と が 明 ら か に な っ て き た 34。 し た が っ て Maf の 阻 害 は 、急 性 炎 症 を 抑 制 す る と 同 時 に 、マ ク ロ フ ァ ー ジ の 性 質 をNrf2 優位な形質 に 転 換 す る こ と で 、 様 々 な 慢 性 炎 症 疾 患 の 治 療 に 貢 献 で き る 可 能 性 が あ る 。
図2. Maf は CD169+マ ク ロ フ ァ ー ジ の 分 化 ・ 局 在 を 制 御 し な い (a-d) 図に示す組織に存在する免疫細胞の構成比をフローサイトメトリー解析し た 。Maf+/-キ メ ラ マ ウ ス お よ び Maf-/-キ メ ラ マ ウ ス は 、 骨 髄 細 胞 を 移 植 後2 ヶ月の 2 次 移 植 キ メ ラ マ ウ ス で 解 析 し た 。Maf を欠損した場合でも、各組織における免疫細 胞 お よ びCD169+マ ク ロ フ ァ ー ジ の 存 在 率 は 、 変 化 し な い 。Maf+/-お よ びMaf-/-キ メ ラ マ ウ ス 各 3 匹のうち、代表的な図を表示した。B; B 細胞、T; T 細胞、Mo; 単 球 、Neu; 好中球、Eos; 好酸球、Mφ; マクロファージを意味する。(e) 大腸に存 在 す る7AAD-CD11b+Ly6G-Ly6C-CD64+マ ク ロ フ ァ ー ジ の 内 の CD169+マ ク ロ フ ァ ー ジ の 存 在 率 ( 左 ) と 細 胞 数 ( 右 ) の 平 均 値 とSD をグラフに示した。Maf の欠損 は 、CD169+マ ク ロ フ ァ ー ジ の 細 胞 数 に も 影 響 し な い 。n.s., not significant, Student’s t-test. (f) 免疫組織化学染色によって、大腸の CD169(赤)、筋肉細胞のマーカーα-SMA(緑)、核(青)を蛍光染色した。Maf の欠損は CD169+マ ク ロ フ ァ ー ジ の 局 在 に 影 響 し な い 。x20, スケールバーは 100µm を示す。2 回の独立した実験のうち、 代 表 的 な 写 真 を 掲 載 し た 。(g) 大腸に存在する 7AAD-CD11b+Ly6G-Ly6C-CD64+マ ク ロ フ ァ ー ジ を 、CD169+お よ びCD169-に 分 け 、 図 に 示 す そ れ ぞ れ の 表 面 マ ー カ ー 発 現 を フ ロ ー サ イ ト メ ト リ ー 解 析 し た 。 灰 色 : ア イ ソ タ イ プ 抗 体 で 染 色 し たMaf+/-マ ク ロ フ ァ ー ジ 、 赤 : 図 示 す る 抗 体 で 染 色 し たMaf+/-マ ク ロ フ ァ ー ジ, 青:Maf-/-の マ ク ロ フ ァ ー ジ を 示 す 。Maf の欠損はマクロファージの表面マーカー発現に影響しな
0 7 0 100 200 300 400 Days CCL8 (ng/ml)
a
Maf+/-Maf-/-colon macrophages
0 3 6 9 Ccl8 CD169- CD169+ * 0 1 2 3 Maf CD169- CD169+ * Fol d c ha nge CCL8 ELISA Serum C C L8 ( ng/ml) DSS day0 day7
b
* * n.s.Maf-/-chimera mouse
Maf +/-0 40 80 120 Ccl8 0 5000 10000 15000 20000 Il6 LPS
-
+
* * * * n.s. Maf +/-Maf-/-BMDMc
0 10 20 30 40 Maf 0 300 600 900 Ccl8 LPS-
+
LPS-
+
コントロール Maf 強制発現 BMDC -40 (h) ↑ ウイルス 感染 ↓ RNA抽出 5 0 ↑ LPSe
LPS-
+
Fol d c ha nge Fol d c ha nge * * Fol d c ha nge 0 50 100 150 Il10 0 1 2 3 CD169 0 10 20 30 Maf Maf+/- Maf-/- LPS-
+
LPS-
+
- + 0 1 2 3 4 CCL8 (ng/ml) LPS - + 0 1 2 3 IL-10 (ng/ml) LPS 0 - + 2 4 6 8 10 IL-6 (ng/ml) LPSCCL8 ELISA IL-10 ELISA IL-6 ELISA
図3. Maf は CD169+マ ク ロ フ ァ ー ジ の 機 能 を 制 御 す る
図4. Maf は CCL8 の 発 現 を 直 接 制 御 す る
0 3 6 9 Cxcl1 * Microarray of BMDM a -8 -4 0 4 8 12 Maf Maf +/-Maf -/-Slpi Ccl8 IL10 Slc7a11 (xCT) Vegfa Mmp13 Cxcl1 Fgl2 Maf-/-BMDM Maf +/-0 50 100 150 Slpi 0 50 100 150 Slc7a11 Fol d cha nge * * 0 5 10 15 Slpi 0 2 4 6 Slc7a11 Fol d cha nge Nrf2-/-BMDM Nrf2 +/-* * 0 50 100 150 0 100 200 300 400 500 600 Maf-/-BMDM Maf +/-Sur vi va l ( % ) tBHP (µM) 0 20 40 60 80 100 Ccl8 0 20 40 60 Fgl2 0 20 40 60 80 Il10 LPS - + - + - + LPS - + - + Fol d cha nge Maf-/-BMDM Maf +/-+ - LPS - + - + 0 10 20 30 Cxcl1 + -* b f g i * * *
GO Term P Value Benjamini
defense response 3.67E-13 1.07E-09
immune response 1.61E-10 2.35E-07
inflammatory response 1.12E-09 1.09E-06 positive regulation of immune system process 3.08E-09 2.25E-06 c * 0 2000 4000 6000 8000 Nos2 Maf+/- Maf -/-Fol d cha nge 0 20 40 60 Il1b 0 0.5 1 1.5 2 Tnf 0 10000 20000 30000 Arg1 0 3000 6000 9000 Fizz1 0 3000 6000 9000 Chil3 Fol d cha nge
Maf+/- Maf-/- Maf+/- Maf-/- Maf+/-Maf-/- Maf+/- Maf-/- Maf+/- Maf
図5. Maf は Nrf2 の 転 写 活 性 と 拮 抗 す る
(a) LPS 刺激した Maf+/-BMDM および Maf-/-BMDM の遺伝子発現をマイクロアレイ解 析 し 、 シ グ ナ ル 値 を 散 布 図 で 示 し た 。 赤 で 示 す プ ロ ッ ト は Maf-/-BMDM で発現が低 下 し た 遺 伝 子 、 青 で 示 す プ ロ ッ ト は Maf-/-BMDM で発現が亢進した遺伝子を意味す る 。2 倍 以 上 発 現 が 変 動 し た 遺 伝 子 を 記 載 し た 。 (b,f,g,h) 図 に 示 し た 遺 伝 子 型 の BMDM における、LPS 刺激 5 時間後の遺伝子発現を qRT-PCR で定量した。Maf +/-BMDM もしくは Nrf2+/-BMDM において、未刺激の BMDM の遺伝子発現に対する相 対 値 を 縦 軸 に 示 し た 。Slc7a11 は xCT のコーディング遺伝子。Hmox1 は HO-1 のコ
0 10 20 30 40
Slpi
0 10 20 30Slc7a11
DEM 0h 5h 24h 0h 5h 24h 0 100 200 300Ccl8
0 200 400 600Il10
LPS 0h 5h 24h 0h 5h 24h DEM LPS Nrf2-/-BMDM Nrf2+/-d
* * * * n.s. n.s. n.s. n.s. ARE consensus sequenceTGA (G/C) NNN GC MARE consensus sequence
TGC TGA (G/C) TCA GCA
図6. Maf は Nrf2 活 性 と 拮 抗 す る が 、 Nrf2 は Maf を 抑 制 し な い
図7. Maf 発 現 量 の 減 少 に よ り マ ク ロ フ ァ ー ジ が 形 質 転 換 す る
0 40 80 120 5 10 15 20 5 10 15 20 Naive D E M 5h miR-129 0 100 200 300 Mir129 DEM 0 5 24 (h) Luc
Luc- Maf 3’UTR Reporter
+1 +2018 +2031
図8. Maf は プ ロ テ ア ソ ー ム お よ び miRNA に よ っ て 分 解 さ れ る (a) 100µM DEM および 10µM MG132 を添加し、5 時間培養した野生型 BMDM にお け る タ ン パ ク 質 発 現 量 を ウ エ ス タ ン ブ ロ ッ ト で 確 認 し た 。DEM 刺激に伴う Maf 発 現 量 の 減 弱 は 、MG132 によってキャンセルされる。写真は 2 回の独立した実験のう ち 、代 表 的 な デ ー タ を 記 載 し た 。(b) DEM 刺激した野生型 BMDM の miR-155 発現を qRT-PCR で定量した。DEM は miR-155 発現レベルを促進しない。縦軸には未刺激の BMDM に対する相対値を示した。n.s., not significant, one-way ANOVA. 2 回の独立し た 実 験 の う ち 、代 表 的 な デ ー タ を 記 載 し た 。(c) 100µM DEM 刺激した野生型 BMDM に お け るmiRNA 発現レベルを microRNA-sequencing 解析した。直線より外側は DEM 刺 激 後 5 時間で 3 倍以上発現が変化した miRNA を示す。(d) DEM 刺激した野生型 BMDM における miR-129 発現を qRT-PCR で確認した。DEM 刺激によって miR-129 発 現 が 亢 進 す る 。 縦 軸 に は 未 刺 激 の BMDM に対する相対値を示した。 (e) Maf 3´-UTR 上に存在する miR-129 結合配列を、microRNA.org で検索した結果(上)。miRNA ミ ミ ッ ク をHEK293T 細胞に導入した際の Maf 3´-UTR レポータープラスミドのルシ フ ェ ラ ー ゼ 活 性 を 示 し た( 下 )。Maf mRNA の 3´-UTR 上の 2 箇所に miR-129 が結合
し 、Maf mRNA 発 現 を 抑 制 す る 。 (b,d,e) 平 均 値 と SD を グ ラ フ に 示 し た 。 解 析 は
図9. 大 腸 マ ク ロ フ ァ ー ジ は Maf 発 現 の 減 少 に よ り 形 質 を 変 え る
第6 章 参 考 文 献
1. Okabe Y, Medzhitov R. Tissue biology perspective on macrophages. Nat Immunol 17, 9-17 (2016).
2. Okabe Y, Medzhitov R. Tissue-Specific Signals Control Reversible Program of Localization and Functional Polarization of Macrophages. Cell 157, 832-844 (2014). 3. Jeremy S. D., et al. Selective depletion of macrophages reveals distinct, opposing
roles during liver injury and repair., J Clin Invest. 115(1), 56-65 (2005).
4. Ming-Zhi Zhang, et al., CSF-1 signaling mediates recovery from acute kidney injury. J Clin Invest. 122(12), 4519-4532 (2012).
5. Charles D. M., et al., M-1/M-2 Macrophages and the Th1/Th2 Paradigm. J Immunol 164, 6166-6173 (2000).Ming-Zhi Zhang, et al., CSF-1 signaling mediates recovery from acute kidney injury. J Clin Invest. 122(12), 4519-4532 (2012).
6. Antonio S.and Alberto M., Macrophage plasticity and polarization: in vivo veritas. J Clin Invest. 122(3), 787-795 (2012).
7. Gordon S., and P. R. Taylor. Monocyte and macrophage heterogeneity. Nat Rev
Immunol 5, 953–964 (2005).
8. Stein M., et al., Interleukin 4 potently enhances murine macrophage mannose receptor activity: a marker of alternative immunologic macrophage activation. J Exp
Med 176, 287–292 (1992).
9. Peter J. M., et al., Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 41, 14-20 (2014)
10. Kohyama, M., et al., Role for Spi-C in the development of red pulp macrophages and splenic iron homeostasis. Nature 457, 318–321 (2009).
11. Malay H., et al., Heme-mediated SPI-C induction promotes monocyte differentiation into iron-recycling macrophages., Cell 156(6), 1223-1234 (2014).
12. Christopher K G. & Gioacchino N., Molecular control of activation and priming in macrophages. Nat Immunol 17(1), 26-33 (2016).
13. Ido A., et al., The role of the local environment and epigenetics in shaping
macrophage identity and their effect on tissue homeostasis. Nat Immunol 17(1), 18-25(2016).
14. Martinez-Pomares L, Gordon S. CD169+ macrophages at the crossroads of antigen presentation. Trends Immunol 33, 66-70 (2012).
2268-2278 (2007).
16. Asano K. et al., CD169-positive macrophages dominate antitumor immunity by crosspresenting dead cell-associated antigens. Immunity 34(1), 85-95 (2011). 17. Asano K, et al. Intestinal CD169(+) macrophages initiate mucosal inflammation by
secreting CCL8 that recruits inflammatory monocytes. Nat Commun 6, 7802 (2015). 18. Ishii Y, et al. Transcription factor Nrf2 plays a pivotal role in protection against
elastase-induced pulmonary inflammation and emphysema. J Immunol 175, 6968-6975 (2005).
19. Sasaki H, et al. Electrophile response element-mediated induction of the
cystine/glutamate exchange transporter gene expression. J Biol Chem 277, 44765-44771 (2002).
20. Qiang Ma, Role of nrf2 in oxidative stress and toxicity. Annu Rev Pharmacol Toxicol 53, 401-426 (2013)
21. KiKuchi K., et al. Macrophages Switch Their Phenotype by Regulating Maf Expression during Different Phases of Inflammation. J Immunol 201(2), 635-651 (2018).
22. Tamoutounour S, et al. CD64 distinguishes macrophages from dendritic cells in the gut and reveals the Th1-inducing role of mesenteric lymph node macrophages during colitis. Eur J Immunol 42, 3150-3166 (2012).
23. Kusakabe M, et al. Maf plays a crucial role for the definitive erythropoiesis that accompanies erythroblastic island formation in the fetal liver. Blood 118, 1374-1385 (2011).
24. Apetoh L., et al., The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T cells induced by IL-27. Nat Immunol 11, 854– 861 (2010).
25. Cao S., et al., The protooncogene c-Maf is an es- sential transcription factor for IL-10 gene expression in macrophages. J Immunol 174, 3484–3492. (2005).
26. Karasawa K, et al. Vascular-resident CD169-positive monocytes and macrophages control neutrophil accumulation in the kidney with ischemia-reperfusion injury. J Am
Soc Nephrol 26, 896-906 (2015).
27. Kataoka K. Multiple mechanisms and functions of maf transcription factors in the regulation of tissue-specific genes. J Biochem 141, 775-781 (2007).
28. Yoshida T, Ohkumo T, Ishibashi S, Yasuda K. The 5´-AT-rich half-site of Maf recognition element: a functional target for bZIP transcription factor Maf. Nucleic
29. Motz G. T., and G. Coukos. The parallel lives of angiogenesis and
immunosuppression: cancer and other tales. Nat Rev Immunol 11, 702–711 (2011). 30. Seifert L., et al. The necrosome pro- motes pancreatic oncogenesis via CXCL1 and
Mincle-induced immune suppression. Nature 532: 245–249 (2016).
31. Voron T., et al., Control of the immune response by pro-angiogenic factors. Front
Oncol 4, 70 (2014).
32. Reardon C., et al., Thymic stromal lymphopoetin-induced expression of the endogenous inhibitory enzyme SLPI mediates recovery from colonic inflammation.
Immunity 35, 223-235 (2011).
33. Ishii T., et al., Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. J Biol Chem 275, 16023-16029 (2000). 34. Kobayashi E. H., et al., Nrf2 suppresses macrophage inflammatory response by
blocking proinflammatory cytokine transcription. Nat Commun 7, 11624 (2016). 35. Banjac A, et al. The cystine/cysteine cycle: a redox cycle regulating susceptibility
versus resistance to cell death. Oncogene 27, 1618-1628 (2008).
36. Kimura M, et al. Molecular basis distinguishing the DNA binding profile of Nrf2-Maf heterodimer from that of Nrf2-Maf homodimer. J Biol Chem 282, 33681-33690 (2007).
37. Dhakshinamoorthy S., and A. K. Jaiswal. c-Maf negatively regulates ARE-mediated detoxifying enzyme genes expression and anti-oxidant induction. Oncogene 21, 5301-5312 (2002).
38. Reddy NM., et al., Disruption of Nrf2 impairs the resolution of hyperoxia-induced acute lung injury and inflammation in mice. J Immunol 182, 7264-7271 (2009). 39. O’Connell RM, Taganov KD, Boldin MP, Cheng G, Baltimore D. MicroRNA-155 is
induced during the macrophage inflammatory response. Proc Natl Acad Sci U S A 104, 1604-1609 (2007).
40. Rodriguez A, et al. Requirement of bic/microRNA-155 for normal immune function.
Science 316, 608-611 (2007).
41. Millar A. D., et al., Evaluating the antioxidant potential of new treatments for inflammatory bowel disease using a rat model of colitis. Gut 39, 407–415 (1996). 42. Stout, R. D., C. Jiang, B. Matta, I. Tietzel, S. K. Watkins, and J. Suttles. 2005.
Macrophages sequentially change their functional phenotype in response to changes in microenvironmental influences. J. Immunol. 175: 342–349.
43. Adams DO, Hamilton TA. The cell biology of macrophage activation. Annu Rev
44. Ginhoux F., et al., New insights into the multidimensional concept of macrophage ontogeny, activation and function. Nat Immunol 17, 34-40 (2016).
45. Bain C. C., et al., Constant replenishment from circulating monocytes maintains the macrophage pool in the intestine of adult mice. Nat Immunol 15, 929–937 (2014). 46. Zigmond E., et al., Ly6C hi monocytes in the inflamed colon give rise to
proinflammatory effector cells and migratory antigen-presenting cells. Immunity 37, 1076–1090 (2012).
47. Bain C. C., et al., Resident and pro-inflammatory macrophages in the colon represent alternative context-dependent fates of the same Ly6Chi monocyte precursors.
Mucosal Immunol 6, 498–510 (2013).
48. Kawauchi S., et al., Regulation of lens fiber cell differentiation by transcription factor c-Maf. J. Biol. Chem. 274, 19254–19260 (1999).
49. Aziz A., et al., MafB/c-Maf deficiency enables self-renewal of differentiated functional macrophages. Science 326, 867–871 (2016).
50. Soucie E. L., et al. Lineage-specific enhancers activate self-renewal genes in macrophages and embryonic stem cells. Science 351, aad5510 (2016).
51. Itoh, K., et al.、An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem Biophys
Res Commun 236, 313–322 (1997).