①海域の栄養塩環境が低次生産に及ぼす影響解明
イ.大阪湾における基礎生産簡易測定技術の開発
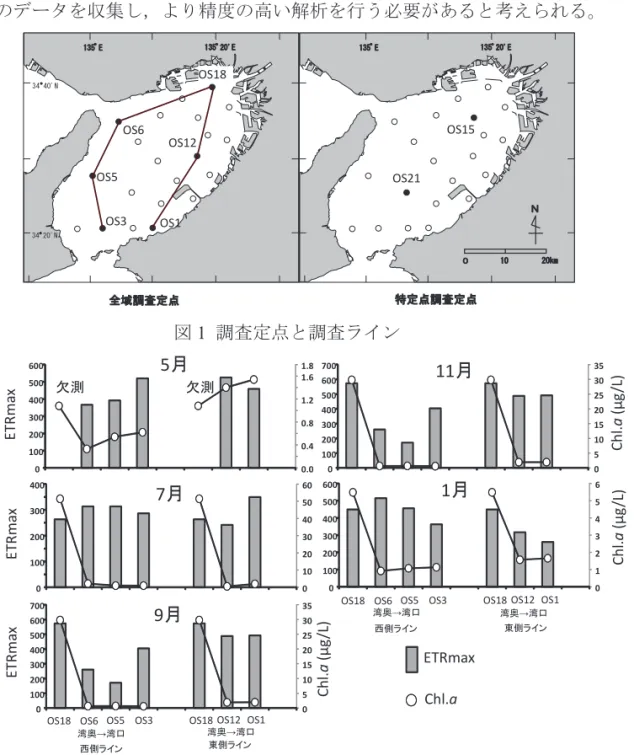
大阪府立環境農林水産総合研究所水産技術センター 山本 圭吾,秋山 諭,田中咲絵 1 全体計画 (1) 目的 近年,瀬戸内海海域では,貧栄養化が進行したことによる水産資源への影響が指摘され ている。同海域の低次生物生産に関しては,各種モニタリングによって現存量のデータは 整備されつつあるものの,生産速度や転換効率に関する知見は断片的であり,瀬戸内海各 海域においてどのような生物生産構造が成り立っているのかについては明らかになって いない。そこで,瀬戸内海東部の大阪湾において,栄養塩等の海域の水質環境調査を実施 するとともに動・植物プランクトンの低次生物生産に関する調査を行い,水質環境が海域 の生産力に及ぼす影響を把握するとともに,基礎生産速度等を簡易に測定する技術の開発 を目指す。 (2) 試験等の方法 瀬戸内海東部の大阪湾において定点(図1)を設置し,年 5 回の現場観測調査を実施す る。水温,塩分,光条件等の鉛直プロファイルを獲得するとともに,採水調査およびネ ット採集調査を実施し,低次生物生産量を算定する。また,光合成活性を測定する機器 (パルス変調型光合成測定装置:PAM)を活用し現場海域での水柱基礎生産量を推定す るとともに,従来法(疑似現場培養法)による測定結果と比較検討する。 2 平成 28 年度計画および結果 (1) 目的 全体計画と同じ。 (2) 試験等の方法 全体計画と同じ。詳細を以下に示す。 調査実施月日 平成 28 年 5 月 9,12 日(全域調査のみ),7 月 4,5 日,9 月 6 日,11 月 7 日, 平成29 年 1 月 4,6 日,3 月 1,2 日(予定) 調査回数は周年でのデータ獲得を目指して奇数月に計6 回実施した。ただし,従来法 との比較については計画通り5 回の実施となった。なお,本報告では平成 29 年 1 月調査 までの4 回分をとりまとめた。 調査定点(図1)全域調査(6 定点;Stn. OS1, OS3, OS5, OS6, OS12, OS18) 特定点調査(2 定点;Stn. OS15 及び OS21)
1) 全域調査 ・CTD 観測(水温・塩分・DO),透明度 多項目式CTD(ASTD102,JFE アドバンテック),セッキ板を用いて測定した。 ・動物プランクトン採集(改良型ノルパックネット,100μm メッシュ) 濾水計を装着したネットを海底まで垂下し,おもり着底後から海面まで約 1m/秒 の速度で鉛直曳網した。採取したネットプランクトンサンプルは,中性ホルマリン (最終濃度約10%)で固定し,課題 4-①-エの解析に供した。 ・栄養塩(DIN,DIP,D-Si),Chl.a,植物プランクトン組成 栄養塩濃度は,自動分析装置(swAAt,ビーエルテック株式会社)を用いた比色 法によって測定した。Chl.a 濃度は,アセトン抽出蛍光法を用いた。植物プランクト ン組成は,原則として表層試水1mL に含まれる全数を計数した。 ・PAM 測定
光合成測定装置(WATER-PAM CUVETTE-Version,WALZ 社,以下 PAM と称する) を用いた。測定用の試水は表層から採水し,500mL 褐色瓶に入れて遮光した後,船 上に設置した大型のクーラーボックス内に海水を満たし,その中で保管することに よって温度を一定に保った。 光合成の測定は,研究室に戻り次第実施した。測定条件は,試水量 1.2mL(専用 キュベットを使用),ML レベル 12 以下,Gain レベルはデフォルト,PM-gain レベ ルは測定可能な範囲で可能な限り14 以下とした。PAR(光合成有効放射)の測定間 隔は30 秒とした。測定 PAR 値は,広島大学(課題 4-①-ウ)の設定(0~2743 の 9 段階)を使用し,それぞれの PAR 条件で得られた rETR(=PAR×ΦII,相対電子伝 達 速 度 ) と の デ ー タ セ ッ ト か ら , 光 合 成 曲 線 :rETR = ETRmax×(1-exp(-(a × PAR)/ETRmax))にあてはめた場合の残差平方和が最小となるように 2 種類のパラメー タ(ETRmaxおよびa)を算出した。
2) 特定点調査 ・PAM 測定 現場の透明度から光量子量が表層の48%(以下,中層 1),33%(中層 2),14%(中 層3),8%(中層 4)にそれぞれ減衰する 4 層を推定し,表層を加えた 5 層のそれぞれ から採取した試水について全域調査と同様に光合成を測定した。 ・光量子量測定 光量子計(JFE アドバンテック社小型メモリー光量子計 DEFI2-L を小型メモリー圧 力計DEFI2-DHG と併用)を多項目式 CTD に装着し,鉛直垂下により現場観測した。 ・明暗瓶法(酸素法)による基礎生産量推定(PAM との比較) PAM 測定に供した海水(表層,中層 1,中層 2,中層 3,中層 4 の計 5 層分)と同 じ試水を用いた。それぞれの試水を明瓶と暗瓶(各 duplicate)にてインキュベータ内 で12 時間培養した後,溶存酸素濃度の差から算出した。培養温度は,調査日の大阪湾 南部(大阪府泉南郡岬町地先,水産技術センター定置観測)の水温とした。光条件は, 蛍光灯を用いて,表層海水は480 μmol/m2/s,中層 1 はその 48%,中層 2 は 33%,中層 3 は 14%,中層 4 は 8%となるように黒色遮光幕を使って減衰設定した。各 2 本の平均
値の明暗瓶差から酸素発生量を算出し,その値に0.375(モル比)を乗じ光合成商(1.3) で除することによって(炭素)光合成速度(μgC/L/h)を求めた。 (3) 結果および考察 本年は周年データが得られなかった項目もあるため,主にPAM による光合成の測定値 から得られた結果と従来法(明暗瓶法:酸素法)で測定した光合成速度の関係について 検討した結果を報告する。 1) 現場観測調査 ①全域調査 大阪湾奥から湾口に延びるラインを西側と東側に2 本設定し,これらのライン上におけ る表層の光合成曲線パラメータ(ETRmax)と傾きa,および Chl.aの変化を検討した(図 2,図 3)。5 月調査では Chl.a は全域で 2μg/L 未満と低い傾向であった。ETRmaxは湾奥の OS18 で収束せず欠測となったが,東側のラインでは湾口にかけて低く,西側のラインで は,湾口にかけて高くなっていた。傾きa は西側では ETRmaxとは逆に湾口にかけて低く なっていたが,東側ではETRmaxと同様湾口にかけて低くなっていた。7 月,9 月は,Chl.a は湾奥のOS18 で 10μg/L を超える高い濃度で観測されたが,その他の定点では低い値で あった。ETRmaxは 7 月には各定点でほぼ同じレベルであったが,9 月は西側のラインで 低い傾向がみられた。また傾きa は 7 月には 5 月同様西側で ETRmaxと逆の,東側で同様 の増減傾向であったが,9 月は東側,西側とも ETRmaxと同様の増減傾向であった。11 月 は7 月,9 月と同様に湾奥で高い傾向があったが,全体的に濃度は低く,6μg/L 未満であ った。1 月の調査では Chl.a は東側,西側とも湾中部で高かったが,ETRmaxは湾奥で高い 傾向であった。各月における優占種は,5 月は Leptocylindrus danicus,7 月が Skeletonema spp.,9 月は Skeletonema spp.,Chaetoceros spp.,Leptocylindrus danicus の混合,11 月は
Skeletonema spp.主体,Chaetoceros spp., Skeletonema spp.であった。
②特定点調査
Chl.a は 7 月,9 月,1 月に高い傾向で,定点別には 1 月を除き,OS15 で高い値となっ た(図4)。明暗瓶法による光合成速度も 7 月,9 月,1 月に比較的高い傾向であったが, 7 月,9 月で特に高く,Chl.a 濃度の傾向と比較すると 1 月の光合成速度は低かった(図 5)。 PAM による測定値から計算された ETRmaxは7 月の両定点と 9 月の OS21 で高い値となっ たが定点間の差は小さかった(図6)。一方,傾き a は 9 月を除き OS21 で高い傾向であ った。鉛直的な光合成速度(図7)は,7 月は OS15 で有光層がかなり浅かったが,両定 点とも中層1 で高く,深くなるに従って低くなった一方で,Chl.a あたりの光合成速度(以 下,光合成活性とする)は表層で最も高く,深くなるに従って低くなった。9 月は,OS21 においては7 月と同じ傾向であったが,OS15 では光合成速度も表層で最も高かった。11 月もOS21 では過去 2 回の調査と同様の傾向であったが,OS15 では中層 3 まで光合成速 度の差が小さく,光合成活性の差も同様に小さかった。1 月は 7 月と類似した傾向を示し, OS15 では表層で,OS21 では中層 1 で光合成速度,光合成活性とも高い傾向であった。 2) 明暗瓶法(酸素法)による基礎生産量推定と PAM との比較
明暗瓶法の結果と推定 rETR を比較した結果,全層の結果,上層 3 層の結果とも光合 成活性との関係が生産速度との関係に比べ相関が高い傾向が見られた(全層の光合成 速度,全層,上層 3 層の光合成活性でそれぞれ R2=0.24,0.41,0.22,P<0.01,0.01, 0.05 で有意)(図 8)。そこで,光合成活性との関係を層別に検討したところ(図 9), 中層1 において有意な関係が確認された(R2=0.51, p<0.05)。一方,中層 3,4 につい ては明暗瓶法の光合成速度が非常に低く,rETR との相関性も低いことから,PAM を 活用した基礎生産のモニタリングにおいては中層3(光量が表層の 33%)あたりまで の測定で条件を満たすと推測された。以上のように,本年の結果から PAM による簡 易な基礎生産モニタリングの可能性が示唆された。しかしながら,本年の結果では周 年の観測が完結しておらず,層別の観測データも少ないことから,次年度においても 同様のデータを収集し,より精度の高い解析を行う必要があると考えられる。 34 40'N° 135 E° 135 20'E° 34 20'N° OS1 OS3 OS5 OS18 OS12 OS6 135 E° 135 20'E° 0 N 10 20km OS15 OS21 全域調査定点 特定点調査定点 図1 調査定点と調査ライン ET Rm ax 欠測 欠測 0.0 0.4 0.8 1.2 1.6 1.8 0 100 200 300 400 500 600 0 10 20 30 40 50 60 0 100 200 300 400 ET Rm ax 0 5 10 15 20 25 30 35 0 100 200 300 400 500 600 700 ETRmax 0 5 10 15 20 25 30 35 0 100 200 300 400 500 600 700 0 1 2 3 4 5 6 0 100 200 300 400 500 600 Ch l.a (μ g/ L) Chl .a (μg /L) Chl.a 7月 9月 11月 1月 5月
OS18 OS6 OS5 OS3 OS18 OS12 OS1
東側ライン 西側ライン
OS18 OS6 OS5 OS3 OS18 OS12 OS1
東側ライン 西側ライン ETRm ax Ch l.a (μ g/ L) 湾奥→湾口 湾奥→湾口 湾奥→湾口 湾奥→湾口
図2 PAM 測定によって推定された表層試水の光合成曲線パラメータ(ETRmax)とア セトン抽出蛍光法により求めたChl.a
図 3 PAM 測定によって推定された表層試水の光合成曲線パラメータ(傾き a)とア セトン抽出蛍光法により求めたChl.a
図4 特定点(OS15 と OS21)における各層の Chl.a(左から表層~中層 4)
図5 特定点(OS15 と OS21)における明暗瓶法による各層の光合成速度 (左から表層~中層4) a a a 0.0 0.4 0.8 1.2 1.6 1.8 0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.75 0 10 20 30 40 50 60 0.40 0.45 0.50 0.55 0.60 0 5 10 15 20 25 30 35 0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0 1 2 3 4 5 6 0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 0.80 0 2 4 6 8 10 12 0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70 Ch l.a (μg /L) Chl .a (μg /L) Chl. a (μg /L) a Chl.a
OS18 OS6 OS5 OS3 OS18 OS12 OS1
東側ライン 西側ライン
OS18 OS6 OS5 OS3 OS18 OS12 OS1
東側ライン 西側ライン 湾奥→湾口 湾奥→湾口 湾奥→湾口 湾奥→湾口 7月 9月 11月 1月 5月 0 2 4 6 8 10 12 14 16
Chl.
a
(μg
/L
)
OS15
OS21
表層→中層47月
9月
11月
1月
5月
80 0 10 20 30 40 50 60 70光合成速度
(μgC/L/h)
7月
9月
11月
1月
5月
表層→中層4OS15
OS21
図6 特定点(OS15 と OS21)における各層の光合成曲線パラメータ(ETRmaxとa) (左から表層~中層4) 図7 特定点(OS15とOS21)における光合成速度,Chl.a 濃度,光合成活性の鉛直 プロファイル;中層は,透明度から表層光量子量の 48%,33%,14%,8%に減 衰する層を現場で推定し,採水した。 0.00 0.20 0.40 0.60 0.80
a
0 1,000 2,000 3,000 4,000ETRma
x
OS15
OS21
表層→中層47月
9月
11月
1月
5月
0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 0 50 100 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 0 5 10 15 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 0 5 10 15 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0 0 50 100 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0 0 5 10 15 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0 0 5 10 15 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0 0 10 20 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0 0 1 2 0.0 2.0 4.0 6.0 8.0 10.0 12.0 14.0 16.0 0 10 20 0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0 0 10 20 30 0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0 0 1 2 0.0 1.0 2.0 3.0 4.0 5.0 6.0 7.0 8.0 9.0 10.0 0 10 20 光合成速度 Chl.a 光合成活性 OS15 OS21 水 深( m ) 水 深( m ) 水深 (m ) ( μg/L ) (μgC/L/h ) ( μgC/Chl.a/h ) ( μg/L ) (μgC/L/h ) ( μgC/Chl.a/h ) ( μg/L ) (μgC/L/h ) ( μgC/Chl.a/h ) ( μg/L ) (μgC/L/h ) ( μgC/Chl.a/h )7月
9月
11月
1月
y = 1.78 x + 57.73 R² = 0.24, p<0.01 0 50 100 150 200 250 0 20 40 60 80 y = 0.61 x + 113.23 R² = 0.05 0 50 100 150 200 250 0 50 100 y = 7.15 x + 93.34 R² = 0.22, p<0.05 0 50 100 150 200 250 0 10 20 y = 12.72 x + 43.21 R² = 0.41, p<0.01 0 50 100 150 200 250 0 5 10 15 20
全層
上
3層
光合成速度(μgC/L/h) 光合成活性 (μgC/Chl.a/h) rETR rETR n=48 n=24 n=48 n=24 図 8 明暗瓶法(酸素法)で測定した光合成速度,光合成活性とPAM 測定結果から推 定したrETR との関係;*相関が有意であったものに p 値を追加した。単位 rETR:μmol electron mg/Chl.a/s
図9 明暗瓶法(酸素法)で測定した光合成活性とPAM 測定結果から推定した rETR の層別の関係;*相関が有意であったものに p 値を追加した。 y = ‐1.07 x + 201.65 R² = 0.03 0 50 100 150 200 250 0 5 10 15 20 y = 6.74 x + 79.36 R² = 0.51, p<0.05 0 50 100 150 200 0 5 10 15 20 y = 1.02 x + 77.97 R² = 0.01 0 20 40 60 80 100 120 0 5 10 15 20 y = 0.02 x + 35.82 R² = 0.00 0 5 10 15 20 25 30 35 40 45 0 5 10 15 20 y = 2.43 x + 17.26 R² = 0.17 0 10 20 30 40 50 60 0 5 10 15 20 rE TR rETR rETR rE TR rE TR 表層(n=8) 中層1(n=8) 中層2(n=8) 中層3(n=8) 中層4(n=8) 光合成活性 (μgC/Chl.a/h) 光合成活性 (μgC/Chl.a/h)
①海域の栄養塩環境が低次生産に及ぼす影響解明
ウ.広島県海域における基礎生産簡易測定技術の開発
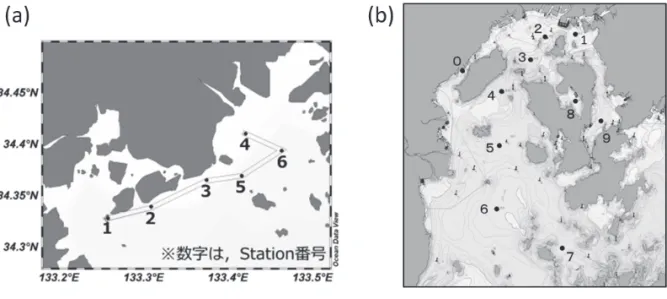
広島大学 生物圏科学研究科 小池一彦 水産研究・教育機構 瀬戸内海区水産研究所 阿保勝之,阿部和雄,松原 賢 1 全体計画 (1)目的 近年,瀬戸内海海域では,貧栄養化が進行したことによる水産資源への影響が指摘されて いる。同海域の基礎生産に関しては,各種モニタリングによって一次生産者の現存量データ は整備されつつあるものの,その生産速度に関する知見は断片的であり,瀬戸内海各海域に おいてどのように生物生産構造の基礎が成り立っているのかについては明らかになっていな い。そこで,瀬戸内海中西部の広島県海域において,栄養塩等の海域の水質環境調査を実施 するとともに植物プランクトンの基礎生産に関する調査を行い,水質環境が海域の生物生産 力に及ぼす影響を把握するとともに,基礎生産速度等を簡易に測定する技術の開発を目指す。 (2)試験等の方法 瀬戸内海中西部の広島県福山市沖において,横島~田島~福山港にいたる定線を設置し, この定線上に 6 定点(図 1a)を設置する。基本的には毎月一回(計 12 回程度)の基礎生産 調査を実施する。加えて,田島沖は広島県でほぼ唯一のノリ養殖海域であるので,海域の環 境とノリの色落ちとの関係を調べるべく,12 月~1 月にかけては高頻度調査(1~2 週に一回 程度)も実施する。さらに,海域の基礎生産を向上させる試みとして,福山港内において漁 業者の協力を得ながら海底耕耘を行う。このことによって,海底の貧酸素化を改善すると共 に,海底付近の高濃度栄養塩や珪藻類の休眠期細胞を水中に拡散させ,基礎生産の土台部分 の拡大を狙う。 広島湾において定点を設置し(図 1b),年 4 回の現場観測調査を実施する。水温,塩分, 光条件等の鉛直プロファイルを獲得するとともに,採水調査およびネット採集調査を実施し, 動・植物プランクトンの低次生物生産に関する調査を行う。また,光合成活性を測定する機器 (パルス変調型光合成測定装置)を活用し現場海域での水柱基礎生産速度を推定する。これ により,基礎生産の簡易測定技術の確立に資する。 2 平成28 年度計画及び結果 (1)目的 全体計画と同じ。 (2)試験等の方法 全体計画と同じ。詳細を以下に記す。 1)福山市沖 福山市沖に6 定点を設置し(図 1a),平成 28 年 4 月以降,平成 29 年 2 月まで毎月一回実施 した。本報告では平成28 年 12 月までの 9 回分の調査結果を取り纏めた。調査分析項目と分析方法は以下のとおり。 ・CTD 観測(水温・塩分・溶存酸素量・蛍光強度・光量子量) 多項目水質計(ハイドロラボ製,DS-5)を用いて測定した。 ・栄養塩(DIN,DIP,D-Si),クロロフィル a,植物プランクトン組成 海面下1m,3m,10mにおいて採水を行い,海水中の無機態栄養塩濃度およびクロロ フィル a 濃度を測定した。栄養塩濃度は,自動分析装置(SWAAT,ビーエルテック)を 用いた比色法によって測定した。クロロフィル a 濃度は,DMF で抽出した試料を Holm-Hansen 法により測定した。植物プランクトン組成は,1 m 層,3 m 層,10 m 層の試 水1 mL に含まれる細胞数を検鏡した。 ・PAM 測定 光合成測定装置(WATER-PAM CUVETTE-Version,WALZ 社)を用いた。測定用の試 水は海面下1m,3m,10mにおいて採水し,褐色瓶(200 ml)に入れて遮光した後,採 水温度を保ち研究室に輸送した。 光合成の測定の際の測定条件は,試水量 1.2mL(専用キュベットを使用),ML レベル 12 以下,Gain レベルはデフォルト,PM-gain レベル 14 以下,PAR(光合成有効放射)の 測定間隔は30 秒または 60 秒とした。測定 PAR 値は,機器固有の設定(0~2743 の 9 段 階)を使用し,それぞれのPAR 条件で得られた rETR(=PAR×ΦII,相対電子伝達速度) とのデータセットから,光合成曲線:rETR=ETRmax×(1-exp(-(a×PAR)/ETRmax))を求めた。 ・基礎生産速度の推定 PAM の測定結果を用いて調査日を含む 1 ヶ月間の基礎生産速度を推定した。まず,西 日本農業研究センター(広島県福山市西深津町)における全天日射量の時別データを用 いて1 ヶ月間の各時刻の平均全天日射量を求めた。PAR を全天日射量の 47%,海面反射 率を15%と仮定し,光量子計により測定した現場の光減衰係数から各時刻における PAR の鉛直分布を求めた。つぎに,PAM 測定で求めた光合成曲線を用いて rETR の鉛直分布 を推定し,光化学系II への分配係数 0.768(珪藻群の代表値;Johnsen and Sakshaug, 2007) およびGoto et al (2008)に基づいて測定したクロロフィル吸収断面(a*)を乗じて,ETR (電子伝達速度,µmol electron mg Chl. a -1 sec-1)を計算した。得られたETR をクロロフ ィル a あたりの酸素発生速度に換算し,Iriarte et al (1991)よるクロロフィル a 濃度に基づ く植物プランクトンの呼吸速度を減じて純酸素発生量を求めた。さらに,光合成商を1.3 として炭素同化速度に変換し,最終的に水柱の炭素同化速度を積算して基礎生産速度(gC m-2 day-1)とした。生産層(有光層深度)は純光合成速度がゼロになる水深とした。これ らの計算は MS エクセルシート上の所定セルにデータを入力することにより,モデル式 の係数決定も含め自動的に算出されるように工夫した。 2)広島湾 広島湾に10 定点を設置し(図 1b), 平成28 年 4 月 25 日~27 日,7 月 2 日~4 日,10 月 14 日~16 日および平成 29 年 1 月 31 日~2 月 1 日の計 4 回実施した。なお,本報告では平成 28 年 10 月までの 3 回分の調査結果を取り纏めた。調査分析項目と分析方法は以下のとおり。 ・CTD 観測(水温・塩分・蛍光強度・DO・光量子量),透明度, 多項目水質計(JFE アドバンテック社製,AAQ127),セッキ板を用いて測定した。 ・動物プランクトン採集(改良型ノルパックネット,100μm メッシュ)
濾水計を装着したネットを海底まで垂下し,おもり着底後から海面まで毎秒約1m の速 度で鉛直曳網した。採取したネットプランクトンサンプルは,中性ホルマリン(最終濃度 約5%)で固定し,課題 4-①-エの解析に供した。 ・栄養塩(DIN,DIP,D-Si),クロロフィル a,植物プランクトン組成 海面下1m,3m,10m(St.0 は 7m)において採水を行い,海水中の無機態栄養塩濃 度およびクロロフィルa 濃度を測定した。栄養塩濃度は,自動分析装置(TRAACS 2000, ビーエルテック株式会社)を用いた比色法によって測定した。クロロフィル a 濃度は, DMF で抽出した試料を Holm-Hansen 法により測定した。植物プランクトン組成は,1m 層,3m 層,10m 層の試水 100~1,000 mL に含まれる細胞数を検鏡した。 ・PAM 測定 光合成測定装置(WATER-PAM CUVETTE-Version,WALZ 社)を用いた。測定用の試 水は海面下1m,3m,10m(St.0 は 7m)において採水し,褐色瓶(100ml または 500ml) に入れて遮光した後,採水温度に保った水槽内に保管した。 光合成の測定は,10 分以上水槽内に保管した後,採水後 2 時間以内に実施した。測定 条件は,試水量1.2mL(専用キュベットを使用),ML レベル 12 以下,Gain レベルはデフ ォルト,PM-gain レベル 14 以下,PAR(光合成有効放射)の測定間隔は 30 秒または 60 秒とした。測定PAR 値は,機器固有の設定(0~2678 の 9 段階)を使用し,それぞれの PAR 条件で得られた rETR(=PAR×ΦII,相対電子伝達速度)とのデータセットから,光 合成曲線:rETR=ETRmax×(1-exp(-(a×PAR)/ETRmax))を求めた。
・基礎生産速度の推定 PAM の測定結果を用いて調査日を含む 1 ヶ月間の基礎生産速度を推定した。まず,広 島気象台における全天日射量の時別データを用いて 1 ヶ月間の各時刻の平均全天日射量 を求めた。PAR を全天日射量の 47%,海面反射率を 15%と仮定し,光量子計により測定 した現場の光減衰係数から各時刻における PAR の鉛直分布を求めた。つぎに,PAM 測 定で求めた光合成曲線を用いてrETR の鉛直分布を推定し,0.5 a*を乗じてETR(電子伝
達速度,mol electron mg Chl. a -1 sec-1)を計算した。ここで,a*はクロロフィル吸収断
面であり海水中のクロロフィル a 濃度(µg L-1)から簡易式(a* = 0.0403×(Chl. a)-0.332) により求めた。得られたETR をクロロフィル a あたりの酸素発生速度に換算し,濃度か ら推定した植物プランクトンの呼吸速度を減じてクロロフィル a あたりの純酸素発生量 を求めた。さらに,光合成商を1.3 として炭素同化速度に変換し,最終的に水柱の炭素同 化速度を積算して基礎生産速度(gC m-2 day-1)とした。 (3)結果及び考察 1)福山市沖 ①環境 図に定点を繋いだ側線上の鉛直水温(図 2)・塩分(図 3)・溶存酸素(図 4)・クロロフ ィル a 濃度(図5)の鉛直・季節分布を示す。4 月から例年にない早い水温上昇が見られ, 8 月には著しい高水温下による顕著な成層化,それに引き続く 9,10 月の高水温化が認め られた。10 月と 11 月は秋雨前線の活発な動きのため降雨が多く,海域全体で塩分 30 台の 低塩分化が認められた。6 月~8 月には成層による底層の低酸素化が認められた。
無機態窒素(図 6)は概ね福山港湾奥の芦田川由来と思われ,St 4 において高濃度であ った。10 月から 11 月は降雨の影響により高濃度化した。無機態リン(図 7)は河川から の目立った供給は無く,夏期の底泥からの供給に依存しているのか,この時期のみに底層 において高濃度化した。この海域は通年にわたり N/P が 20 を超えるリン制限海域である が,2016 年は夏期の顕著な成層化による底層からのリン供給が多かったため,後述するよ うに秋期の珪藻のブルームが見られた。 ②基礎生産量と植物プランクトン出現量 4 月~1 月にかけての基礎生産速度は 0.40~6.14 gC m-2 day-1の範囲内であった(図8)。 基礎生産は5 月から顕著な上昇を示し,7 月に最大を示した。5 月の基礎生産は,全点平 均で4.39 gC m-2 day-1であった。ただしクロロフィル a 濃度としては低く,平均4.24 µg L-1 であった。図9 には植物プランクトン分類群毎の細胞数構成比を示した。この海域では 5 月に有害プランクトンの Vicicitus globosus が多く出現するが,2016 年の出現量は少なく, 最大でも2 cells mL-1であり,珪藻類,特に Guinardia flaccida が主に出現した。クロロフィ ル a 濃度が低いにもかかわらずこの様に高い基礎生産速度を与えた要因としては,特に St.5 と St.6 において,減衰係数 0.185 および 0.238 に代表されるような透明度の高い水塊 がみられたこと,それに伴い,15 m を超えるような補償深度を記録したこと,さらに,光 合成の最大量子収率である Fv/Fm が0.7 以上を記録するような高い活性の植物プランクト ンが占めていたことだと考えられる。 6~7 月にかけても同様に高い基礎生産が記録されたが,このとき特に高いクロロフィル
a 濃度を示した6 月の St.4 にはラフィド藻類 Heterosigma akashiwo が最高細胞密度 1,008 cell
mL-1で出現し,7 月の St.3~St.6 には Vicicitus globosus が最高細胞密度 61 cells mL-1で出現 した。また7 月は,Chattonella spp.が最高細胞密度 18 cells mL-1(St.6 の水深 10 m)が出現 しており,今回の月例サンプリングでは捉えられなかったが,調査の6 日後である,7 月 21 日には Chattonella spp.の赤潮警報も発令され,St.2 付近では,Chattonella spp.の 3 つを 合わせて,最高細胞密度178 cells mL-1の出現が確認されている。8 月には,その名残とも 言うべき Chattonella 細胞群が認められ,C. ovata が最高細胞密度130 cells mL-1で出現し, St.1,St.2,St.5 で出現細胞数の 55.1~84.7 %を占めた(図 9)。このうち St.1 と St.2 では 特に C. ovata が両点の高い基礎生産速度(それぞれ2.94, 3.13 gC m-2 day-1)をもたらした。 この様に,当年は6~8 月にかけ,H. akashiwo や Chattonella spp.など有害ラフィド藻類の 出現が目立った。 9 月に入るとクロロフィル a 濃度,基礎生産速度が共に一旦減少し,10 月に再び急上昇 した。この,10 月の鉛直混合期における高い基礎生産(平均 3.33 gC m-2 day-1)は特にこ の年に顕著で,全点平均のクロロフィル a 濃度は2016 年度内においても最高値(24.56 µg L-1)を示した。この時点でも極低密度の C. ovata が認められたが,出現の主体は珪藻類で, 特に Coscinodiscus wailesii や Chaetoceros socialis 等の出現が顕著だった。その一方,例年 12 月から上昇する基礎生産およびクロロフィル a 濃度の上昇は低いレベルに留まった。こ れは珪藻の増殖がみられなかったことに起因し,12 月には,この時期に珍しく渦鞭毛藻類
Akashiwo sanguinea が大量に出現し,St.4 において赤潮を形成すると共に(最高細胞密度
141 cells mL-1),出現種の主体を構成していた。 ③PAM 蛍光法の利点・問題点
以上のように,PAM 蛍光法を用いることによって,これまでの13C 法では成し得ない高 頻度・多定点での測定が可能となった。しかし,得られる基礎生産速度は既報と比べて高 く,同海域で調べられた過去の基礎生産量と比べ4 倍~7 倍の数値を示した。これは,今 回の調査点が既報に比べて沿岸に近いこと,生産力の高い福山港も対象としたことに加え, PAM 蛍光法が;(1) 基本的には光化学系の上流部から下流への電子伝達から酸素発生量の みを推定していること,(2) 実際の光合成においては発生した酸素の一部が光呼吸などに よって消去されるので,PAM 蛍光法において酸素発生量を過大評価していること,(3) 酸 素発生量からの炭素固定の換算に理論値を用いており実際とは異なること,(4) ボトルイ ンキュベーション法では無視してしまう溶存態有機物としての逸脱を加味していること, (5) ボトルインキュベーションにおいて過小評価につながるインキュベーション中の栄養 の枯渇やボトルエフェクトがないことなどが挙げられる。これらの問題は測定原理の違い にもとづく問題であり解決は難しい。しかし,その簡便さを生かし,多頻度・多定点での 測定を継続して実施し,湾・灘毎のきめ細やかな比較という面で大きな利点がある。 ④海洋環境と基礎生産向上の試み;海底耕耘 夏場に悪化しやすい海底環境の改善と,底層のリンの表層への供給,珪藻休眠期細胞の 拡散による基礎生産向上を狙って,2016 年 6 月 6,7 日,2016 年 7 月 7,8 日,2016 年 8 月1,2 日の合計 6 日において広島県福山市鞆において海底耕耘作業を行った。図 10 に耕 耘範囲と調査点の経度緯度を示す。耕耘区域内(耕耘区)に3 点,耕耘区域外(対象区) に3 点の定点を設定した。福山市漁業青年部の小型底引き網船 3 隻が戦車こぎ網を改良し た耕耘具を用い,原則として9 時から 12 時に海底耕耘を行った。耕耘実施面積は 1.5 km2 であった。調査は,耕耘前として6 月 6 日,7 月 7 日,8 月 1 日に,耕耘後調査として 6 月7 日,7 月 8 日,8 月 2 日に行った。さらに 2 日後調査として 7 月 11 日,8 月 4 日,7 日後調査として6 月 14 日,7 月 15 日,8 月 9 日に行った。耕耘前調査は 8 時から,耕耘 後調査は耕耘終了後に,2 日後と 7 日後調査は 8 時から行った。なお,本報告書では,最 も環境が悪化しやすい8 月の結果を述べる。それ以外の月のデータについては要望に応じ て提供可能である。 図11 に海底直上水(海底-0.5 m)濁度の変化を示した。耕耘区のみで,耕耘後に濁度 が FTU=3.5±1.3 が 6.5±8.5 に上昇し,耕耘による海底攪拌効果が認められた。海底泥を採 取し,強熱減量,酸化還元電位(Eh),揮発性硫化物量(AVS)を調べた(図 12)。なお 対象区と比べ耕耘区の何れの項目も,耕耘前から良好な数値を示しているが,これは前月 の耕耘の効果が持続しているものと解釈される。 強熱減量は,耕耘前(8 月 1 日)には耕耘区で 6.7±0.9%,対照区で 7.9±1.2%であった。 耕耘直後(8 月 2 日)には両区とも若干増加し,耕耘区で 7.9±1.1%,対照区で 9.5±0.5%と なった。耕耘区において増加幅が小さかった。 海底泥酸化還元電位は,耕耘前において耕耘区-0.67±15.3 mV,対照区-10.0±0 mV で あったが,耕耘直後に耕耘区は+40.0±14.0 mV まで上昇したのに対して,対照区では-36.0 ±40.0 mV まで低下した。耕耘 8 日後には耕耘区では+31.0±6.0 mV と比較的高い値を維持 し,対照区では-22.7±100 mV と比較的低い値を維持した。 硫化物量は,耕耘前は耕耘区0.26 ±0.02 mg g-1,対照区0.36± 0.08 mg g-1であった。耕耘 直後は耕耘区で0.21 ± 0.05 mg g-1に減少し,対照区で0.51 ± 0.14 mg g-1に増加した。耕耘8
日後は計測を行っていない。以上のことから,海底耕耘は特に海底の還元状態の還元,硫 化物の低減に効果をもたらしたことが確認された。
海底耕耘前後の植物プランクトン数の変化について図 13 に示した。珪藻類は耕耘直後 に増加し,耕耘3 日後にはさらに大きく増加した。優占種は Skeletonema sp.であった。渦 鞭毛藻類は0 m,4 m において耕耘 8 日後に増加した。水深 2 m においては耕耘 8 日後に 0 cells mL-1から1295 cells mL-1に大きく増加した。優占種は Gyrodinium dominans であった。 ラフィド藻類は耕耘3 日後に水深 0 m において 1140 cells mL-1,水深4m において 700 cells mL-1まで増加し,耕耘8 日後には両水深で消滅した。水深 2m においては耕耘直後に 930 cells mL-1に増加し,耕耘3 日後には消滅した。優占種は Chattonella antiqua と Chattonella
ovata であった。なお,この時福山沖全域で Chattonella 属が赤潮化しており,耕耘が原因 となり増加を招いたとは考えていない。むしろ,耕耘後に珪藻が増加し,それと同じくし て Chattonella が大きく増殖することなく減少しているので,耕耘によって珪藻休眠期細胞 が巻き上げられ,それが Chattonella を押さえたと捉えている。なお,珪藻は4 日目以降大 きく減少したが,これは Gyrodinium dominans の捕食によると考えられる。 2)広島湾 ①栄養塩およびクロロフィル a 各点,各層における栄養塩濃度およびクロロフィル a 濃度を図14 に示した。溶存態無機窒 素(DIN)濃度は,湾奥で高く湾口で低い傾向がみられた。4 月には湾奥の表層では高めだっ たが,湾口よりでは全層で低く0.5~2.4μM であった。7 月には 10m 層では比較的高い点が 多かったが,1m,3m 層では低かった。10 月には各点とも全層で比較的高く 2.7~10.2μM で あった。無機態リン濃度(PO4-P)は,4 月には各点とも低めであり,St.0 を除くと 0.10~0.28 μM であった。7 月には 10m 層では比較的高かったが,1m,3m 層では 0.04~0.24μM と低 かった。10 月は各点とも高めで 0.50~1.42μM であった。ケイ酸態ケイ素(SiO2-Si)は,DIN と同様に,湾奥で高く湾口で低い傾向がみられた。Si/N 比,Si/P 比はそれぞれ 2~37,15~ 927 であり,ケイ藻にとって Si は十分量存在していたと考えられる。クロロフィル a 濃度は 湾奥で高く,特に7 月の St.1,St.2 の 1m,3m 層では約 20 µg L-1と高濃度であった。 ②植物プランクトンの動態 各定点における植物プランクトンの総細胞密度を見ると,St. 0~3 の沿岸域で 4 月,7 月, 10 月のいずれの月でも高く,江田島湾内の St. 8 では 7 月および 10 月に高密度であった(図 15)。1m 層の植物プランクトン組成を見ると,4 月の St. 0,1,8,9 では鞭毛藻類が多くみ られ,St. 0,1,8 はクリプト藻類,St. 9 はラフィド藻類の Fibrocapsa japonica であった。ま た,珪藻類では全体的に Chaetoceros 属の優占度が高かった。しかし7 月には Chaetoceros 属 に替わり,全体的に Leptocylindrus 属が優占するようになった。そして10 月になると,St. 0 ~3 では Skeletonema 属が優占し,St. 7~8 では Chaetoceros 属が優占した。層別の組成を見る と,各定点とも概ね1m 層および 3m の組成は類似していた。しかし,特に海域全体的に成層 が発達した7 月の 10m 層については,1,3m 層とは組成が大きく異なっていた。 ③PAM 測定 各点,各層で実施したPAM 測定により求めた光合成曲線のパラメータを図 16 に示した。
ETRmaxはrETR(相対電子伝達速度)の最大値を表すパラメータであり海水中の光合成活性の 指標となる。また,aは光合成曲線の傾きを表すパラメータでありaが高いと光量が少なく ても光合成活性が比較的高いことを示す。4 月,7 月,10 月の ETRmaxはそれぞれ 332~673, 321~1031,336~755 の範囲にあり,7 月には定点による差が大きかった。また採水層による 違いはあまりなかった。aは0.39~0.73 の範囲にあり,月別,層別の差はあまりなかった。 ④基礎生産速度 PAM 測定結果から計算した各月の基礎生産速度(gC m-2 day-1)を図17 に示した。4 月,7 月,10 月の基礎生産速度はそれぞれ 2.5~6.4,2.6~8.3,1.4~5.2 gC m-2 day-1の範囲にあり, 各月の平均値はそれぞれ4.3,4.5,3.4 gC m-2 day-1であった。基礎生産速度は湾奥で大きい傾 向があり,特に7 月の湾奥で大きかった。また,各点における基礎生産速度と各層のクロロ フィル a 濃度はよく対応しており(図18),広島湾内の基礎生産速度の大きさは植物プラン クトンの現存量である程度決まることが示唆された。ただし,ここで推定した基礎生産速度 は,クロロフィル吸収断面a*の推定方法などに誤差が大きい可能性がある。今後は推定方法 について再検討するとともに,他海域の推定結果と比較して議論する必要がある。 3)引用文献
Goto N, Miyazaki H, Nakamura N, Terai H, Ishida N, Mitamura O. Relationships between electron transport rates determined by pulse amplitude modulated (PAM) chlorophyll fluorescence and photosynthetic rates by traditional and common methods in natural freshwater phytoplankton. Archiv für Hydrobiologie, 172, 121-134, 2008.
Iriarte A, Daneri G, Garcia VMT, Purdie DA, Crawford DW. Plankton community respiration and its relationship to chlorophyll a concentration in marine coastal waters. Oceanologica Acta, 14, 379-388, 1991.
Johnsen G, Sakshaug E. Biooptical characteristics of PSII and PSI in 33 species (13 pigment groups) of marine phytoplankton, and the relevance for pulse-amplitude-modulated and
fast-repetition-rate fluorometry. J. Phycol. 43, 1236–1251, 2007.
Platt T, Gallegos CL, Harrison WG. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 38, 687-701, 1080.